- Патофизиология углеводного метаболизма

Содержание
- 2. ПАТОФИЗИОЛОГИЯ УГЛЕВОДНОГО МЕТАБОЛИЗМА
- 3. УГЛЕВОДЫ ПИЩИ крахмал сахароза лактоза глюкоза фруктоза
- 4. В сбалансированной диете примерно 50% энергетических потребностей организма должны быть удовлетворены за счет углеводов. Аскорбиновая кислота
- 5. Для поступления глюкозы в клетку нужен переносчик, поскольку молекула глюкозы гидрофильна, а ПМ – гидрофобна.
- 6. Первый переносчик был выделен из эритроцитов человека. Он представляет собою полипептидную цепь из 492 аминокислотных остатков,
- 7. ПРЕДПОЛАГАЕМОЕ УСТРОЙСТВО ПЕРЕНОСЧИКА ГЛЮКОЗЫ
- 8. Белок-переносчик имеет две конформации: одна связывает глюкозу на внеклеточной стороне мембраны, другая – на внутриклеточной.
- 9. Конформационные изменения, изолирующие контакт вне- и внутриклеточного компартментов, направлены на предотвращение проникновения ионов натрия вместе с
- 10. ПРОНИКНОВЕНИЕ ГЛЮКОЗЫ В КЛЕТКУ
- 11. ПРЕДПОЛАГАЕМАЯ ПОРА ДЛЯ ГЛЮКОЗЫ
- 12. Переносчики глюкозы обозначаются GluT и пронумерованы по мере их открытия. Все пять белков осуществляют перенос глюкозы
- 13. GluT 2 обнаружен в органах, выделяющих глюкозу в кровь (кишечник, печень, почки) и в В-клетках панкреатических
- 14. GluT 3 имеется в нейронах мозга. Обладая большим, чем GluT 1, сродством к глюкозе, переносчик GluT
- 15. GluT 4 – главный переносчик глюкозы в мио- и липоцитах. Его особенность - этот белок перемещается
- 16. ПЕРЕНОСЧИКИ ГЛЮКОЗЫ В ЛИПОЦИТАХ
- 17. T3 индуцирует формирование переносчика глюкозы GLUT 4, беспечивая и базальный, и инсулинзависимый транспорт глюкозы в
- 18. GluT 5 встречается главным образом в тонкой кишке. Подробности его функционирования пока неизвестны.
- 19. Семейство белков-переносчиков GluT отлично от ко-транспортера, белка-переносчика, транспортирующего глюкозу против градиента концентрации.
- 20. Энергию для переноса глюкозы ко-транспортером обеспечивает сопряженный перенос ионов Na по градиенту концентрации Na. Благодаря такому
- 21. НАИБОЛЕЕ ВЫРАЖЕН ОТВЕТ НА ИНСУЛИН У GLUT 4 (МИО- И ЛИПОЦИТЫ): СВЯЗЫВАНИЕ ИНСУЛИНА СО СВОИМ РЕЦЕПТОРОМ
- 22. Выступающая часть рецептора к инсулину связывает инсулин (1), а внутренняя часть (2) вызывает фосфорилирование белка-мишени, что
- 23. Тироидные гормоны и всасывание глюкозы в ЖКТ Тироидные гормоны ускоряют всасывание глюкозы в ЖКТ. При
- 24. Комментарий к патогенезу ИНСД Животные модели ИНСД могут иметь следующие дефекты: ?сниженное содержание GluT 2 в
- 25. ? дефект хотя бы одного из белков-переносчиков в ПМ миоцитов (80% утилизации глюкозы в условиях нагрузки
- 26. ? другие возможные пострецепторные дефекты: ⮚на этапе сигнализации о движении-переброске переносчиков к мембране, ⮚ на этапах
- 27. Метаболизм углеводов и его гормональный контроль
- 28. Главное место превращения углеводов пищи – гепатоциты. ГЛЮ диффундирует в клетку через GluT2 и сразу подвергается
- 29. Гепатоцит способен превращать избыток глюкозы в гликоген, что позволяет создать запас недиффундирующей глюкозы без сопутствующего осмотического
- 30. Гексокиназная (глюкокиназная) реакция стимулируется Инсулином, который «конкурирует» за гексокиназу с глюкокортикоидами (стимулируют распад Глю-6-фосфата,
- 31. Гликогеногенез ? Стимулируют : инсулин переводит фосфорилазу в неактивную форму; активируя гексокиназу, обеспечивает гликогеногенез субстратом
- 32. ? Тормозят: катехоламины «противодействуют» инсулину, глюкагон (антагонист активации гликогенсинтетазы), тироидные гормоны (стимулируют гликогенолиз,
- 33. Гормональный контроль гликогенолиза ? стимулируют: глюкагон (клетки-мишени: гепатоцит и липоцит) - цАМФ - зависимая активация
- 34. Гликолиз (путь превращения 1 моля Глю в 2 моля лактата с образованием 2 молей АТФ в
- 35. Гормональный контроль реакций гликолиза : ?стимулируют: Инсулин (стимулятор фосфофруктокиназы (ФФК) и пируваткиназы(ПВК) ,
- 36. Тироидные гормоны – (стимуляторы активности ключевых ферментов гликолиза). ? Тормозят: Соматотропин – (ингибитор ферментов
- 37. Пируват ДГ- система реакций (образовавшийся в гликолизе Пир диффундирует в митохондрии, где превращается в ацетил-Ко А,
- 38. ЦТК (Кребса) (Ацетил Ко А подвергается декарбоксилированию и дегидрированию. Водород акцептируется НАД: атомы водорода становятся донорами
- 39. Другие регуляторные ферменты: цитратсинтаза (угнетается АТФ и НАД.Н) изоцитрат ДГ (стимулируется АДФ) сукцинат ДГ.
- 40. Гормональный контроль ЦТК осуществляет инсулин через повышение активности цитратсинтазы, изоцитрат ДГ, СДГ.
- 41. Пентозный цикл (прямое окисление глюкозы, гексозомонофосфатный шунт; в гепатоците 1/3 глюкозы окисляется по этому пути). Прямое
- 42. Гормональный контроль пентозного цикла осуществляют: ? стимуляция: Инсулин 1) окислительного этапа цикла, ? активность Глю-6-фосфат
- 43. Окислительное фосфорилирование (процесс, при котором выделение энергии при окислении субстратов сопряжено с синтезом АТФ): первый пункт
- 44. Гормональный контроль окислительного фосфорилирования определяется, прежде всего, уровнем гормонов щитовидной железы: стимуляция активности Na-K-АТФазы,
- 45. стимуляция активности митохондриальной альфа-глицерофосфатДГ, стимуляция активности СДГ и цитохромоксидазы,
- 46. разобщение дыхания и фосфорилирования в первом пункте сопряжения, «латентное» разобщение второго пункта сопряжения.
- 47. Глюконеогенез (образование глюкозы de novo из углеводных и неуглеводных продуктов: лактат анаэробного гликолиза, аминокислоты, глицерин, оксалоацетат,
- 48. Гормональный контроль глюконеогенеза: ? Глюкагон (стимулирует ключевые ферменты, гормончувствительную липазу в гепатоцитах, тормозит синтез
- 49. ? Глюкокортикоиды (стимулируют ключевые ферменты глюконеогенеза, распад жиров (стимуляция липазы), распад белков (аминокислоты используются как
- 50. ? СТГ ( стимулирует ключевые ферменты, липолиз, угнетает гликолиз).
- 51. ? Т3,Т4 стимулируют глюконеогенез, ускоряют распад инсулина в гепатоцитах.
- 52. ОСОБЕННОСТИ МЕТАБОЛИЗМА УГЛЕВОДОВ В РАЗЛИЧНЫХ ОРГАНАХ И ТКАНЯХ
- 53. Мышцы 1. Цель миоцита – мышечное сокращение, для которого следует наиболее эффективно использовать глюкозу для образования
- 54. Основные энергетические потребности всех типов мышц удовлетворяются в основном за счет окисления продуктов метаболизма жиров. 2.
- 55. 5.В миоците отсутствует Глю-6-фосфатаза и использование глюкозы ограничено исключительно внутриклеточным метаболизмом. 6.Различны субъединицы изоферментов ЛДГ в
- 56. Кардиомиоциты содержат так много митохондрий, что работают по сути в аэробных условиях. Кардиомиоцит окисляет не только
- 57. Напротив, в скелетной мышце максимальна активность ферментов гликолиза и пируват превращается в лактат. Образовавшийся лактат диффундирует
- 58. Мозг Если в крови, поступающей к мозгу, уровень глюкозы снижается в 2 раза, то в течение
- 59. Мозг более высокочувствителен к гипоксемии, чем к гипогликемии. В условиях длительного голодания для обеспечения ЦТК субстратами
- 60. Наследственные ферментопатии
- 61. Синдромы мальабсорбции – «ферментный блок» на этапе всасывания. Эти синдромы относят к категории наследственных аутосомно-рецессивных ферментопатий.
- 62. В клинике такого рода патологии всегда имеют ведущими симптомами: ?бродильный понос, ? эксикоз, ? вторичная гипотрофия.
- 63. Понос вызывают нерасщепленные дисахариды или невсосавшиеся моносахариды, поступившие в более дистальные отделы кишечника. Изменение осмотического давления
- 64. При врожденной алактазии и врожденном нарушении всасывания глюкозы и галактозы симптомы проявляются после первых прикладываний к
- 65. При врожденном недостатке инвертазы или мальтазы симптоматика появляется при переходе на искусственное вскармливание (введение в организм
- 66. Синдромы мальабсорбции – «ферментный блок» в межуточном обмене углеводов. Эти синдромы редки, но опасны в связи
- 67. Галактоза – продукт переваривания лактозы. Причин этой патологии две: ☹ недостаток галактокиназы или ☹ недостаток галактозо-1-фосфат-уридилтрансферазы.
- 68. В организме накапливаются нерасщепленные и токсичные галактоза и галактозо-1-фосфат. Избыток галактозы превращается в токсичный галактитол, влияющий
- 69. Исключение из рациона молока и других источников галактозы позволяет предотвратить развитие патологических признаков!!!
- 70. Лабораторно: резко увеличено содержание галактозы (до 1г/л), галактозурия, билирубинемия, протеинурия, гипераминоацидурия.
- 71. Фруктоземия (непереносимость фруктозы). Аутосомно-рецессивная форма патологии. Причина – отсутствие альдолазы фруктозо -1- фосфата. Накопление фруктозо-1-фосфата тормозит
- 72. В клинике фруктоземии после приема фруктового сахара или фруктов возникает гипогликемия, дрожь, потливость, рвота, нарушение сознания;
- 73. Доброкачественная эссенциальная фруктозурия Характеризуется повышенным содержанием фруктозы в моче, что м.б. основанием ошибочного диагноза – диабет.
- 74. ГЛИКОГЕНОЗЫ
- 75. ГЛИКОГЕНОЗЫ – генетически детерминированные формы патологии, при которых нарушение нормальных ферментативных реакций в вызывает избыточное отложение
- 76. I типа, гепаторенальный тип, болезнь Гирке Edgar Otto Conrad von Gierke (9 February 1877, Breslau -
- 77. АУТОСОМНО-РЕЦЕССИВНЫЙ ТИП НАСЛЕДОВАНИЯ
- 78. Организм б-го неспособен превратить глюкозо-6-фосфат в глюкозу. накопление гликогена в гепато- и нефроцитах связано с подавлением
- 79. Недостаток глюкозы крови? низкие уровни инсулина ? усиление липолиза ? липемия ? ацетонемия (ацидоз) ? ацетонурия.
- 80. Для таких б-х характерно: ✈низкие натощаковые уровни глюкозы и редуцированный подъем сахара в ответ на глюкагон
- 81. Даже непродолжительное голодание ведет к гипогликемии. В тяжелых случаях гипогликемия может привести к судорогам. Результат хронически
- 82. Больные дети имеют короткое туловище, большой живот и увеличенную печень (гепатомегалия – наиболее заметный признак! Обусловлена
- 83. II типа, по-преимуществу гликогеноз мышц, б-нь Помпе (Pompe), cardiomegalia glycogenica diffusa, GAA deficiency, acid maltase deficiency,
- 84. Ген, кодирующий кислую глюкозидазу, картирован на 17 хромосоме (локус c17q25). Наледование – аутосомно-рецессивное.
- 85. Гликоген накапливается в лизосомах клеток различных тканей, особенно часто поражаются лизосомы ●кардиомиоцитов, ● гладкой мускулатуры и
- 86. III типа, гепатомышечный тип, ограниченный декстриноз, болезнь Форбса, болезнь Кори (Примерно ¼ гликогенозов, протекающих с увеличением
- 87. Дефект амило-1,6-глюкозидазы (катализирует гидролиз 1-6 связей в молекуле гликогена = debranching enzyme) ? накопление гликогена ?
- 88. Для б-го характерна гепатомегалия как результат накопления гликогена. Гликоген имеет короткие наружные ветви. Гликогенолиз возможен только
- 89. IV типа, болезнь Андерсена Проявляется в раннем детстве как печеночная недостаточность (желтуха, асцит). Сопровождается циррозом печени
- 90. Патогенез: дефицит ветвящего фермента. Содержание гликогена в печени обычно нормально, но структура гликогена нарушена: гликоген имеет
- 91. VI типа, болезнь Херса Недостаток или отсутствие фосфорилазы ? накопление гликогена нормальной структуры. Больные подвержены гипогликемии
- 92. Мышечные гликогенозы II типа, генерализованный гликогеноз, гликогенная кардиомегалия, болезнь Помпе. Около 10% от всех гликогенозов. Наиболее
- 93. V типа, болезнь Мак-Ардля. Поражаются только скелетные мышцы. Встречется наиболее редко. Полное отсутствие фосфорилазы в миоцитах,
- 94. Поскольку активность фосфорилазы в печени соответствует норме, гипогликемия не обнаруживается. Симптомы б-ни обычно выражены незначительно, за
- 95. VII типа Редкие случаи. В мышцах резко снижены уровни фосфофруктокиназы (1-3% от нормы).
- 97. Скачать презентацию

ПАТОФИЗИОЛОГИЯ УГЛЕВОДНОГО
МЕТАБОЛИЗМА
ПАТОФИЗИОЛОГИЯ УГЛЕВОДНОГО
МЕТАБОЛИЗМА

УГЛЕВОДЫ ПИЩИ
крахмал
сахароза
лактоза
глюкоза
фруктоза
УГЛЕВОДЫ ПИЩИ
крахмал
сахароза
лактоза
глюкоза
фруктоза

В сбалансированной диете примерно 50% энергетических потребностей организма должны быть удовлетворены
В сбалансированной диете примерно 50% энергетических потребностей организма должны быть удовлетворены

Для поступления глюкозы в клетку нужен переносчик, поскольку молекула глюкозы гидрофильна,
Для поступления глюкозы в клетку нужен переносчик, поскольку молекула глюкозы гидрофильна,

Первый переносчик был выделен из эритроцитов человека. Он представляет собою полипептидную
Первый переносчик был выделен из эритроцитов человека. Он представляет собою полипептидную
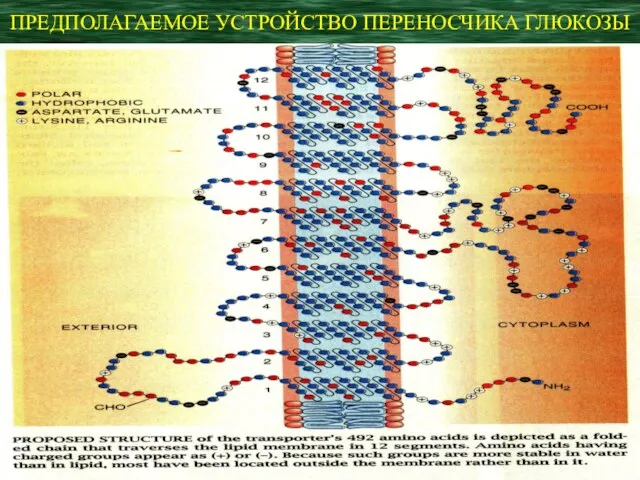
ПРЕДПОЛАГАЕМОЕ УСТРОЙСТВО ПЕРЕНОСЧИКА ГЛЮКОЗЫ
ПРЕДПОЛАГАЕМОЕ УСТРОЙСТВО ПЕРЕНОСЧИКА ГЛЮКОЗЫ

Белок-переносчик имеет две конформации:
одна связывает глюкозу на внеклеточной стороне мембраны,
Белок-переносчик имеет две конформации: одна связывает глюкозу на внеклеточной стороне мембраны,

Конформационные изменения, изолирующие контакт вне- и внутриклеточного компартментов, направлены на предотвращение
Конформационные изменения, изолирующие контакт вне- и внутриклеточного компартментов, направлены на предотвращение
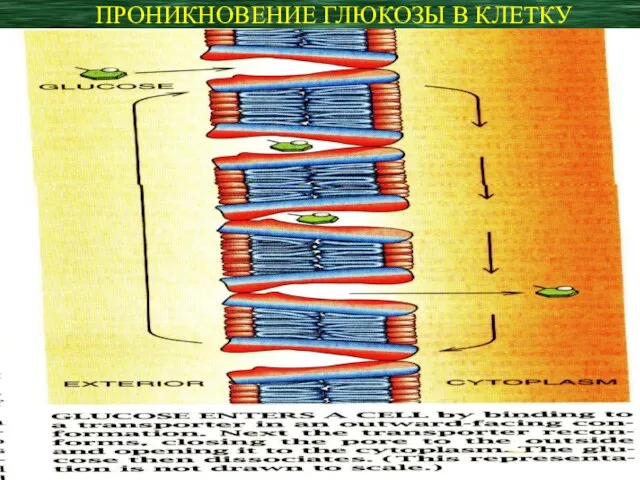
ПРОНИКНОВЕНИЕ ГЛЮКОЗЫ В КЛЕТКУ
ПРОНИКНОВЕНИЕ ГЛЮКОЗЫ В КЛЕТКУ
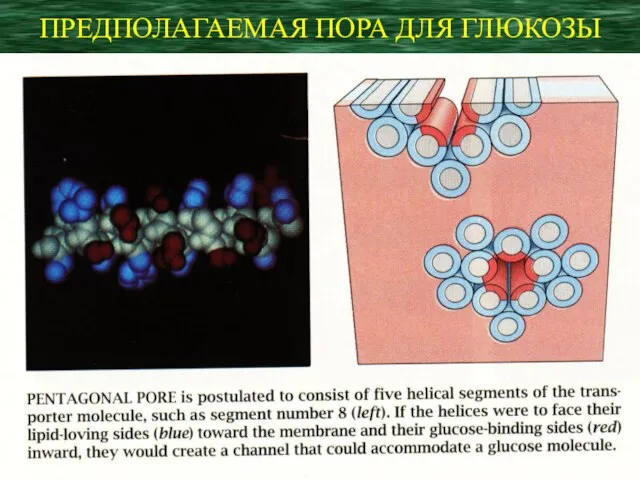
ПРЕДПОЛАГАЕМАЯ ПОРА ДЛЯ ГЛЮКОЗЫ
ПРЕДПОЛАГАЕМАЯ ПОРА ДЛЯ ГЛЮКОЗЫ

Переносчики глюкозы обозначаются GluT и пронумерованы по мере их открытия. Все
Переносчики глюкозы обозначаются GluT и пронумерованы по мере их открытия. Все

GluT 2 обнаружен в органах, выделяющих глюкозу в кровь (кишечник, печень,
GluT 2 обнаружен в органах, выделяющих глюкозу в кровь (кишечник, печень,

GluT 3 имеется в нейронах мозга.
Обладая большим, чем GluT 1, сродством
GluT 3 имеется в нейронах мозга. Обладая большим, чем GluT 1, сродством

GluT 4 – главный переносчик глюкозы в мио- и липоцитах.
Его
GluT 4 – главный переносчик глюкозы в мио- и липоцитах. Его
ПЕРЕНОСЧИКИ ГЛЮКОЗЫ В ЛИПОЦИТАХ
ПЕРЕНОСЧИКИ ГЛЮКОЗЫ В ЛИПОЦИТАХ

T3 индуцирует формирование переносчика глюкозы GLUT 4, беспечивая и базальный,
T3 индуцирует формирование переносчика глюкозы GLUT 4, беспечивая и базальный,

GluT 5 встречается главным образом в тонкой кишке.
Подробности его функционирования
GluT 5 встречается главным образом в тонкой кишке. Подробности его функционирования

Семейство белков-переносчиков GluT отлично от ко-транспортера,
белка-переносчика, транспортирующего глюкозу
против градиента
Семейство белков-переносчиков GluT отлично от ко-транспортера, белка-переносчика, транспортирующего глюкозу против градиента

Энергию для переноса глюкозы
ко-транспортером обеспечивает сопряженный перенос ионов Na по
Энергию для переноса глюкозы ко-транспортером обеспечивает сопряженный перенос ионов Na по

НАИБОЛЕЕ ВЫРАЖЕН ОТВЕТ НА ИНСУЛИН У GLUT 4 (МИО- И ЛИПОЦИТЫ):
СВЯЗЫВАНИЕ
НАИБОЛЕЕ ВЫРАЖЕН ОТВЕТ НА ИНСУЛИН У GLUT 4 (МИО- И ЛИПОЦИТЫ): СВЯЗЫВАНИЕ
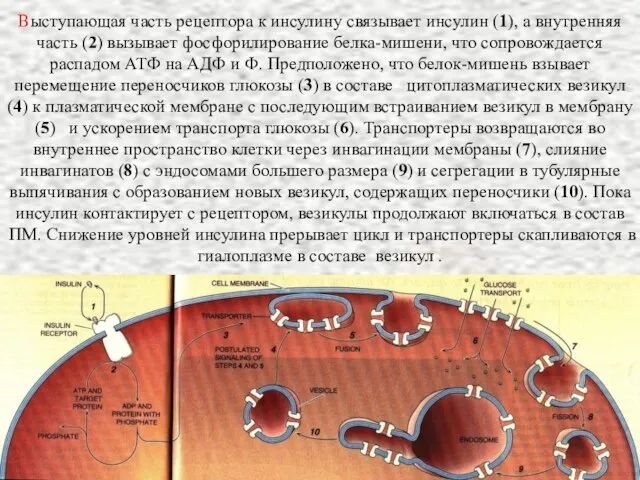
Выступающая часть рецептора к инсулину связывает инсулин (1), а внутренняя часть
Выступающая часть рецептора к инсулину связывает инсулин (1), а внутренняя часть

Тироидные гормоны и всасывание глюкозы в ЖКТ
Тироидные гормоны ускоряют всасывание глюкозы
Тироидные гормоны и всасывание глюкозы в ЖКТ Тироидные гормоны ускоряют всасывание глюкозы

Комментарий к патогенезу ИНСД
Животные модели ИНСД могут иметь следующие дефекты:
?сниженное содержание
Комментарий к патогенезу ИНСД Животные модели ИНСД могут иметь следующие дефекты: ?сниженное содержание

? дефект хотя бы одного из белков-переносчиков в ПМ миоцитов
(80%
? дефект хотя бы одного из белков-переносчиков в ПМ миоцитов (80%

? другие возможные пострецепторные дефекты:
⮚на этапе сигнализации о движении-переброске переносчиков к
? другие возможные пострецепторные дефекты: ⮚на этапе сигнализации о движении-переброске переносчиков к

Метаболизм углеводов и его гормональный
контроль
Метаболизм углеводов и его гормональный
контроль

Главное место превращения углеводов пищи – гепатоциты.
ГЛЮ диффундирует в клетку
Главное место превращения углеводов пищи – гепатоциты. ГЛЮ диффундирует в клетку

Гепатоцит способен превращать избыток глюкозы в гликоген, что позволяет создать запас
Гепатоцит способен превращать избыток глюкозы в гликоген, что позволяет создать запас

Гексокиназная (глюкокиназная)
реакция стимулируется
Инсулином, который «конкурирует» за гексокиназу с
Гексокиназная (глюкокиназная) реакция стимулируется Инсулином, который «конкурирует» за гексокиназу с

Гликогеногенез
? Стимулируют :
инсулин
переводит фосфорилазу в неактивную форму;
активируя
Гликогеногенез ? Стимулируют : инсулин переводит фосфорилазу в неактивную форму; активируя

? Тормозят:
катехоламины «противодействуют» инсулину,
глюкагон (антагонист активации гликогенсинтетазы),
? Тормозят: катехоламины «противодействуют» инсулину, глюкагон (антагонист активации гликогенсинтетазы),

Гормональный контроль гликогенолиза
? стимулируют:
глюкагон (клетки-мишени: гепатоцит и липоцит)
Гормональный контроль гликогенолиза ? стимулируют: глюкагон (клетки-мишени: гепатоцит и липоцит)

Гликолиз
(путь превращения 1 моля Глю в 2 моля лактата с
Гликолиз (путь превращения 1 моля Глю в 2 моля лактата с

Гормональный контроль реакций гликолиза :
?стимулируют:
Инсулин (стимулятор фосфофруктокиназы (ФФК)
Гормональный контроль реакций гликолиза : ?стимулируют: Инсулин (стимулятор фосфофруктокиназы (ФФК)

Тироидные гормоны –
(стимуляторы активности ключевых ферментов гликолиза).
? Тормозят:
Тироидные гормоны – (стимуляторы активности ключевых ферментов гликолиза). ? Тормозят:

Пируват ДГ- система реакций
(образовавшийся в гликолизе Пир диффундирует в митохондрии, где
Пируват ДГ- система реакций (образовавшийся в гликолизе Пир диффундирует в митохондрии, где

ЦТК (Кребса)
(Ацетил Ко А подвергается декарбоксилированию и дегидрированию. Водород акцептируется НАД:
ЦТК (Кребса) (Ацетил Ко А подвергается декарбоксилированию и дегидрированию. Водород акцептируется НАД:

Другие регуляторные ферменты:
цитратсинтаза (угнетается АТФ и НАД.Н)
изоцитрат ДГ (стимулируется
Другие регуляторные ферменты: цитратсинтаза (угнетается АТФ и НАД.Н) изоцитрат ДГ (стимулируется

Гормональный контроль ЦТК осуществляет инсулин через повышение активности
цитратсинтазы,
изоцитрат ДГ,
СДГ.
Гормональный контроль ЦТК осуществляет инсулин через повышение активности
цитратсинтазы,
изоцитрат ДГ,
СДГ.

Пентозный цикл
(прямое окисление глюкозы, гексозомонофосфатный шунт; в гепатоците 1/3 глюкозы окисляется
Пентозный цикл (прямое окисление глюкозы, гексозомонофосфатный шунт; в гепатоците 1/3 глюкозы окисляется

Гормональный контроль пентозного цикла осуществляют:
? стимуляция:
Инсулин
1) окислительного этапа
Гормональный контроль пентозного цикла осуществляют: ? стимуляция: Инсулин 1) окислительного этапа

Окислительное фосфорилирование
(процесс, при котором выделение энергии при окислении субстратов сопряжено с
Окислительное фосфорилирование (процесс, при котором выделение энергии при окислении субстратов сопряжено с

Гормональный контроль окислительного фосфорилирования определяется, прежде всего, уровнем гормонов
щитовидной
железы:
Гормональный контроль окислительного фосфорилирования определяется, прежде всего, уровнем гормонов щитовидной железы:

стимуляция активности митохондриальной альфа-глицерофосфатДГ,
стимуляция активности СДГ и
стимуляция активности митохондриальной альфа-глицерофосфатДГ, стимуляция активности СДГ и

разобщение дыхания
и фосфорилирования в первом пункте
сопряжения,
«латентное»
разобщение дыхания и фосфорилирования в первом пункте сопряжения, «латентное»

Глюконеогенез
(образование глюкозы de novo из углеводных и неуглеводных продуктов: лактат анаэробного
Глюконеогенез (образование глюкозы de novo из углеводных и неуглеводных продуктов: лактат анаэробного

Гормональный контроль глюконеогенеза:
? Глюкагон
(стимулирует ключевые ферменты,
гормончувствительную липазу в
Гормональный контроль глюконеогенеза: ? Глюкагон (стимулирует ключевые ферменты, гормончувствительную липазу в

? Глюкокортикоиды
(стимулируют ключевые ферменты глюконеогенеза,
распад жиров (стимуляция липазы),
распад
? Глюкокортикоиды (стимулируют ключевые ферменты глюконеогенеза, распад жиров (стимуляция липазы), распад

? СТГ
( стимулирует ключевые ферменты,
липолиз,
угнетает гликолиз).
? СТГ
( стимулирует ключевые ферменты,
липолиз,
угнетает гликолиз).

? Т3,Т4
стимулируют глюконеогенез,
ускоряют распад инсулина в гепатоцитах.
? Т3,Т4
стимулируют глюконеогенез,
ускоряют распад инсулина в гепатоцитах.

ОСОБЕННОСТИ МЕТАБОЛИЗМА УГЛЕВОДОВ В РАЗЛИЧНЫХ ОРГАНАХ И ТКАНЯХ
ОСОБЕННОСТИ МЕТАБОЛИЗМА УГЛЕВОДОВ В РАЗЛИЧНЫХ ОРГАНАХ И ТКАНЯХ

Мышцы
1. Цель миоцита – мышечное сокращение, для которого следует наиболее эффективно
Мышцы 1. Цель миоцита – мышечное сокращение, для которого следует наиболее эффективно

Основные энергетические потребности всех типов мышц удовлетворяются в основном за счет
Основные энергетические потребности всех типов мышц удовлетворяются в основном за счет

5.В миоците отсутствует
Глю-6-фосфатаза и использование глюкозы ограничено исключительно внутриклеточным метаболизмом.
6.Различны
5.В миоците отсутствует Глю-6-фосфатаза и использование глюкозы ограничено исключительно внутриклеточным метаболизмом. 6.Различны

Кардиомиоциты содержат так много митохондрий, что работают по сути в аэробных
Кардиомиоциты содержат так много митохондрий, что работают по сути в аэробных

Напротив, в скелетной мышце максимальна активность ферментов гликолиза и пируват превращается
Напротив, в скелетной мышце максимальна активность ферментов гликолиза и пируват превращается

Мозг
Если в крови, поступающей к мозгу, уровень глюкозы снижается в 2
Мозг Если в крови, поступающей к мозгу, уровень глюкозы снижается в 2

Мозг более высокочувствителен к гипоксемии, чем к гипогликемии.
В условиях длительного голодания
Мозг более высокочувствителен к гипоксемии, чем к гипогликемии. В условиях длительного голодания

Наследственные
ферментопатии
Наследственные
ферментопатии

Синдромы мальабсорбции –
«ферментный блок» на этапе всасывания.
Эти синдромы относят к категории
Синдромы мальабсорбции – «ферментный блок» на этапе всасывания. Эти синдромы относят к категории

В клинике такого рода патологии всегда имеют ведущими симптомами:
?бродильный понос,
?
В клинике такого рода патологии всегда имеют ведущими симптомами: ?бродильный понос, ?

Понос вызывают нерасщепленные дисахариды или невсосавшиеся моносахариды, поступившие в более дистальные
Понос вызывают нерасщепленные дисахариды или невсосавшиеся моносахариды, поступившие в более дистальные

При врожденной алактазии и врожденном нарушении всасывания глюкозы и галактозы симптомы
При врожденной алактазии и врожденном нарушении всасывания глюкозы и галактозы симптомы

При врожденном недостатке инвертазы или мальтазы симптоматика появляется при переходе на
При врожденном недостатке инвертазы или мальтазы симптоматика появляется при переходе на

Синдромы мальабсорбции –
«ферментный блок» в межуточном обмене углеводов.
Эти синдромы редки, но
Синдромы мальабсорбции – «ферментный блок» в межуточном обмене углеводов. Эти синдромы редки, но

Галактоза – продукт переваривания лактозы.
Причин этой патологии две:
☹ недостаток галактокиназы
Галактоза – продукт переваривания лактозы. Причин этой патологии две: ☹ недостаток галактокиназы

В организме накапливаются нерасщепленные и токсичные галактоза и галактозо-1-фосфат. Избыток галактозы
В организме накапливаются нерасщепленные и токсичные галактоза и галактозо-1-фосфат. Избыток галактозы

Исключение из рациона молока и других источников галактозы позволяет предотвратить развитие
Исключение из рациона молока и других источников галактозы позволяет предотвратить развитие

Лабораторно:
резко увеличено содержание галактозы (до 1г/л),
галактозурия,
билирубинемия, протеинурия,
Лабораторно: резко увеличено содержание галактозы (до 1г/л), галактозурия, билирубинемия, протеинурия,

Фруктоземия
(непереносимость фруктозы).
Аутосомно-рецессивная форма патологии. Причина – отсутствие альдолазы фруктозо -1-
Фруктоземия (непереносимость фруктозы). Аутосомно-рецессивная форма патологии. Причина – отсутствие альдолазы фруктозо -1-

В клинике фруктоземии
после приема фруктового сахара или фруктов возникает
гипогликемия, дрожь,
В клинике фруктоземии после приема фруктового сахара или фруктов возникает гипогликемия, дрожь,

Доброкачественная эссенциальная фруктозурия
Характеризуется повышенным содержанием фруктозы в моче, что м.б. основанием
Доброкачественная эссенциальная фруктозурия Характеризуется повышенным содержанием фруктозы в моче, что м.б. основанием

ГЛИКОГЕНОЗЫ
ГЛИКОГЕНОЗЫ

ГЛИКОГЕНОЗЫ –
генетически детерминированные формы патологии, при которых нарушение нормальных ферментативных реакций
ГЛИКОГЕНОЗЫ – генетически детерминированные формы патологии, при которых нарушение нормальных ферментативных реакций

I типа, гепаторенальный тип, болезнь Гирке
Edgar Otto Conrad von Gierke
(9
I типа, гепаторенальный тип, болезнь Гирке Edgar Otto Conrad von Gierke (9
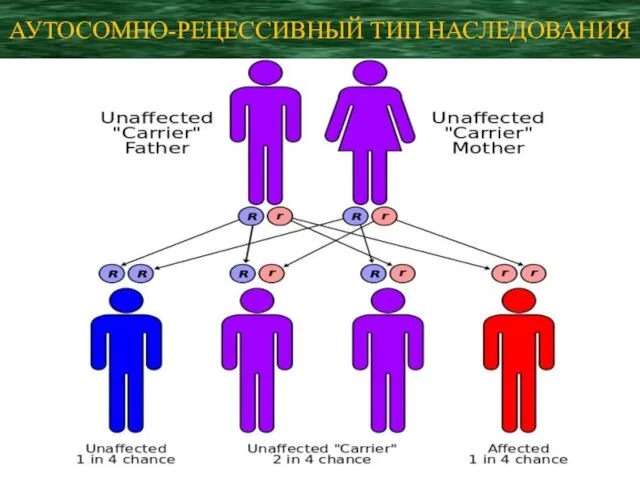
АУТОСОМНО-РЕЦЕССИВНЫЙ ТИП НАСЛЕДОВАНИЯ
АУТОСОМНО-РЕЦЕССИВНЫЙ ТИП НАСЛЕДОВАНИЯ

Организм б-го неспособен превратить глюкозо-6-фосфат в глюкозу.
накопление гликогена в гепато-
Организм б-го неспособен превратить глюкозо-6-фосфат в глюкозу. накопление гликогена в гепато-

Недостаток глюкозы крови?
низкие уровни инсулина ?
усиление липолиза ?
липемия ?
Недостаток глюкозы крови? низкие уровни инсулина ? усиление липолиза ? липемия ?

Для таких б-х характерно:
✈низкие натощаковые уровни глюкозы и редуцированный подъем
Для таких б-х характерно: ✈низкие натощаковые уровни глюкозы и редуцированный подъем

Даже непродолжительное голодание ведет к гипогликемии. В тяжелых случаях гипогликемия может
Даже непродолжительное голодание ведет к гипогликемии. В тяжелых случаях гипогликемия может

Больные дети имеют короткое туловище, большой живот и увеличенную печень
(гепатомегалия
Больные дети имеют короткое туловище, большой живот и увеличенную печень (гепатомегалия

II типа,
по-преимуществу гликогеноз мышц,
б-нь Помпе (Pompe),
cardiomegalia glycogenica diffusa,
II типа, по-преимуществу гликогеноз мышц, б-нь Помпе (Pompe), cardiomegalia glycogenica diffusa,

Ген, кодирующий
кислую глюкозидазу,
картирован на 17 хромосоме (локус c17q25).
Наледование –
Ген, кодирующий кислую глюкозидазу, картирован на 17 хромосоме (локус c17q25). Наледование –

Гликоген накапливается в лизосомах клеток различных тканей, особенно часто поражаются лизосомы
Гликоген накапливается в лизосомах клеток различных тканей, особенно часто поражаются лизосомы

III типа, гепатомышечный тип,
ограниченный декстриноз, болезнь Форбса, болезнь Кори
(Примерно ¼
III типа, гепатомышечный тип, ограниченный декстриноз, болезнь Форбса, болезнь Кори (Примерно ¼

Дефект амило-1,6-глюкозидазы
(катализирует гидролиз 1-6 связей в молекуле гликогена = debranching enzyme)
?
Дефект амило-1,6-глюкозидазы (катализирует гидролиз 1-6 связей в молекуле гликогена = debranching enzyme) ?

Для б-го характерна гепатомегалия как результат накопления гликогена.
Гликоген имеет короткие наружные
Для б-го характерна гепатомегалия как результат накопления гликогена. Гликоген имеет короткие наружные

IV типа, болезнь Андерсена
Проявляется в раннем детстве как печеночная недостаточность
IV типа, болезнь Андерсена Проявляется в раннем детстве как печеночная недостаточность

Патогенез: дефицит ветвящего фермента.
Содержание гликогена в печени обычно нормально, но
структура гликогена
Патогенез: дефицит ветвящего фермента. Содержание гликогена в печени обычно нормально, но структура гликогена

VI типа, болезнь Херса
Недостаток или отсутствие фосфорилазы ?
накопление гликогена нормальной
VI типа, болезнь Херса Недостаток или отсутствие фосфорилазы ? накопление гликогена нормальной

Мышечные гликогенозы
II типа, генерализованный гликогеноз, гликогенная кардиомегалия, болезнь Помпе. Около 10%
Мышечные гликогенозы II типа, генерализованный гликогеноз, гликогенная кардиомегалия, болезнь Помпе. Около 10%

V типа, болезнь Мак-Ардля.
Поражаются только скелетные мышцы.
Встречется наиболее редко.
V типа, болезнь Мак-Ардля. Поражаются только скелетные мышцы. Встречется наиболее редко.

Поскольку активность фосфорилазы в печени соответствует норме, гипогликемия не обнаруживается.
Симптомы б-ни
Поскольку активность фосфорилазы в печени соответствует норме, гипогликемия не обнаруживается. Симптомы б-ни

VII типа
Редкие случаи.
В мышцах резко снижены уровни фосфофруктокиназы (1-3% от
VII типа Редкие случаи. В мышцах резко снижены уровни фосфофруктокиназы (1-3% от
 Презентация по биологии Хвойные растения
Презентация по биологии Хвойные растения  ЦАРСТВО ПРОКАРИОТЫ ПОДЦАРСТВО БАКТЕРИИ
ЦАРСТВО ПРОКАРИОТЫ ПОДЦАРСТВО БАКТЕРИИ  Презентация на тему Дыхательная система Зачем нашему организму кислород?
Презентация на тему Дыхательная система Зачем нашему организму кислород? Кровь и остальные компоненты внутренней среды организма
Кровь и остальные компоненты внутренней среды организма Полезные свойства обычных продуктов
Полезные свойства обычных продуктов Чиполлино и все-все-все… Что без боли и печали к слезам приводит?
Чиполлино и все-все-все… Что без боли и печали к слезам приводит?  ОВ пород и его преобразование в ходе литогенеза
ОВ пород и его преобразование в ходе литогенеза Презентация на тему Защита гидросферы
Презентация на тему Защита гидросферы  Фауна позвоночных. Класс Лучепёрые рыбы
Фауна позвоночных. Класс Лучепёрые рыбы Ткани. Эпителиальная ткань
Ткани. Эпителиальная ткань Класс Птицы (Aves). Подкласс Веерохвостые (Neornithes). Надотряд Типичные птицы (Neognathae)
Класс Птицы (Aves). Подкласс Веерохвостые (Neornithes). Надотряд Типичные птицы (Neognathae) Мочевыделительная система
Мочевыделительная система Живое вещество биосферы и его функции
Живое вещество биосферы и его функции Ботаника. Введение. Корень
Ботаника. Введение. Корень Нервная ткань
Нервная ткань Надцарство прокариоты (Одноклеточные безъядерные организмы)
Надцарство прокариоты (Одноклеточные безъядерные организмы) Презентация на тему "Фармакология" - скачать презентации по Биологии
Презентация на тему "Фармакология" - скачать презентации по Биологии Презентация на тему "Появление человека разумного" - скачать презентации по Биологии
Презентация на тему "Появление человека разумного" - скачать презентации по Биологии Презентация на тему "Золотистое чудо - щедрый подарок природы" - скачать презентации по Биологии
Презентация на тему "Золотистое чудо - щедрый подарок природы" - скачать презентации по Биологии Проверим наши знания
Проверим наши знания …Е121…Е123..Е216…Е217…Е240… Пищевые добавки- полезные, вкусные и запрещённые Московских Анастасия 9 А класс МОУ СОШ № 27 города Бело
…Е121…Е123..Е216…Е217…Е240… Пищевые добавки- полезные, вкусные и запрещённые Московских Анастасия 9 А класс МОУ СОШ № 27 города Бело Живой урок с бабочками
Живой урок с бабочками Удивительная Плесень
Удивительная Плесень Влияние света на интенсивность дыхания
Влияние света на интенсивность дыхания Бойся меня!
Бойся меня! Кожа, строение и функции
Кожа, строение и функции Эволюция жизни на Земле (лекции 4,5)
Эволюция жизни на Земле (лекции 4,5) Решение генетических задач. Генеалогический метод
Решение генетических задач. Генеалогический метод