Проприорецепторы. Моторные рефлексы спинного мозга. Вестибулярный анализатор. Моторные рефлексы ствола мозга
- Проприорецепторы. Моторные рефлексы спинного мозга. Вестибулярный анализатор. Моторные рефлексы ствола мозга

Содержание
- 2. Это самый большой орган – 30-40% массы тела. Благодаря повлению скелетных мышц животные стали отличаться от
- 3. Моторные центры мозга По мере формирования мозга в каждом из его отделов повлялись моторные центры
- 4. Сокращение скелетных мышц направлено не только на выполнение истинных целенаправленных локомоций, но и на противодействие гравитационным
- 5. Проприорецепторы, Вестибулярный, рецепторы кожи, зрительный, слуховой и другие. Рецепторы, участвующие в регуляции моторики
- 6. Нервные центры Многоуровневый (расположен в различных структурах ЦНС) нервный центр обеспечивает человеку возможность выполнения всей разнообразной
- 7. Спинной мозг В сегментах спинного мозга пул мотонейронов каждой мышцы находится в определенном месте, причем: сгибатели
- 8. Сухожильный рефлекс При ударе молоточком по сухожилию (любой мышцы) мышца сокращается. Этот эффект обусловлен рефлексом, замыкающимся
- 9. Проприорецепторы Схема сухожильного органа (а), мышечного веретена (б). 1, 12- афферентные волокна, 2 – интрафузальное мышечное
- 10. Количество мышечных веретен и сухожильных органов в различных мышцах неодинаково. Чем более сложную, более значимую для
- 11. Взаимодействие нейронов в спинном мозге Рецепторы возбуждаются при их растяжении. Импульсы от веретен в спинном мозге
- 12. Сухожильный орган Cухожильные органы - контролируют напряжение мышцы (силу сокращения). А если сокращение слишком сильное (можно
- 13. Мышечные веретена - контролируют длину мышцы. Эта информация поступает в головной мозг (мы всегда знаем в
- 14. В естественных условиях возбуждение α- и γ-мотонейронов одной мышцы возникает чаще всего одновременно. Но вследствие различной
- 15. Гамма-регуляция мышечного сокращения (γ - петля): механизм усиления сокращения мышц Влияние ретикулярной формации ствола мозга указано
- 16. Рефлексы спинного мозга (начинаются с различных рецепторов или автономных программ) Сегментарные: моносинаптические и полисинаптические. Межсегментарные: Сгибательный,
- 17. Центры вышележащих отделов мозга координируют рефлексы спинного мозга. А ретикулярная формация ствола еще и тонизирует центры
- 18. Центры ствола мозга
- 19. Двигательные центры ствола головного мозга. 1 - красное ядро, 2 - ядро ретикулярной формации моста, 3
- 20. В ориентации человека в пространстве помимо проприорецепторов большую роль играет вестибулярная сенсорная система. Она вместе с
- 21. Лабиринт 1 - эндолимфатический мешочек, 2, 3, 4 - полукружные каналы, 5 - улитка, 6 -
- 22. Рецепторная клетка вестибулярного аппарата Реснички двух типов: Одна длинная – киноцилия и много коротких – стероцилий.
- 23. Схема строения, механизм возбуждения и торможения рецепторных клеток Клетка обладает пейсмекерными свойствами (спонтанное возбуждение около 200
- 24. Восприятие изменения движения головы рецепторной клеткой вестибулярного анализатора и появление электрических ответов в окончании сенсорного нейрона
- 25. Схема строения купулы и отолитового аппарата
- 26. Стимулом для рецепторов полукружных каналов являются угловые ускорения. Здесь рецепторные клетки в каждом канале сгруппированы в
- 27. Раздражаются они при изменении скорости движения (начало и конец движений), при так называемых линейных ускорениях. В
- 28. Рецепторы маточки реагируют на изменение скорости горизонтального движения, а мешочка – вертикального (лифтные рефлексы) Механизм возникновения
- 29. Прежде всего: все центры ствола тесно связаны между собой. Рефлексы стволовых моторных ядер реализуются через нисходящие
- 30. Стволовые центры обеспечивают сохранение равновесия и нормальное вертикальное положение тела в условиях действия гравитационного поля Земли
- 31. Изменение тонуса мышц при нарушении взаимодействия центров ствола
- 32. Рефлексы ствола мозга Позные рефлексы: Шейные, Вестибулярные, глазо-двигательные. Статокинетические рефлексы: Статические, Тонические, Ориентировочные. Начинаются они с
- 33. При «неправильном» положении головы с проприорецепторов мышц шеи запускаются движения мышц туловища направленные на сохранение более
- 34. Проявление позных рефлексов у кошки при падении Начинаются движения с поворота головы, т.е. вначале включаются шейные
- 35. Перераспределение тонуса мышц кошки при наклонах головы У животных при наклоне головы вниз (рис. б) повышается
- 36. При наклоне головы вперед повышается тонус мышц-сгибателей и в результате улучшается возможность группировки. При откидывании головы
- 37. При повороте головы повышается тонус мышц сгибателей на стороне поворота, а разгибателей на противоположной стороне. В
- 39. Скачать презентацию

Это самый большой орган – 30-40% массы тела.
Благодаря повлению скелетных мышц
Это самый большой орган – 30-40% массы тела.
Благодаря повлению скелетных мышц
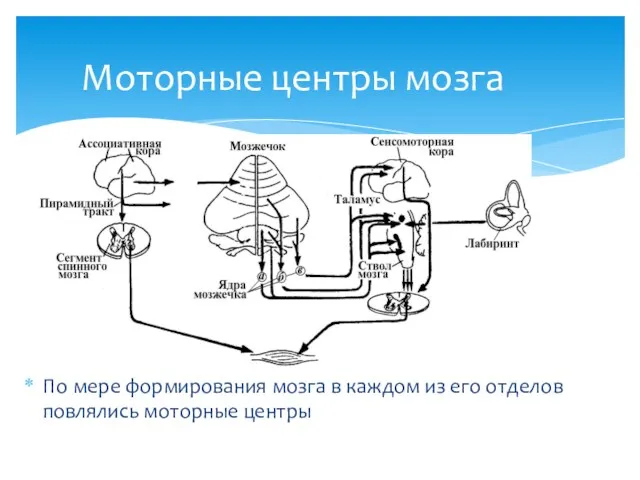
Моторные центры мозга
По мере формирования мозга в каждом из его отделов
Моторные центры мозга
По мере формирования мозга в каждом из его отделов

Сокращение скелетных мышц направлено не только на выполнение истинных целенаправленных локомоций,
Сокращение скелетных мышц направлено не только на выполнение истинных целенаправленных локомоций,

Проприорецепторы,
Вестибулярный,
рецепторы кожи,
зрительный,
слуховой
и другие.
Рецепторы, участвующие в регуляции моторики
Проприорецепторы,
Вестибулярный,
рецепторы кожи,
зрительный,
слуховой
и другие.
Рецепторы, участвующие в регуляции моторики
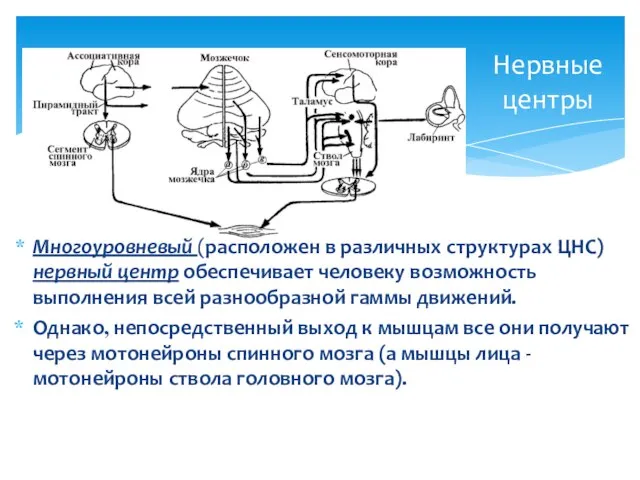
Нервные центры
Многоуровневый (расположен в различных структурах ЦНС) нервный центр обеспечивает человеку
Нервные центры
Многоуровневый (расположен в различных структурах ЦНС) нервный центр обеспечивает человеку
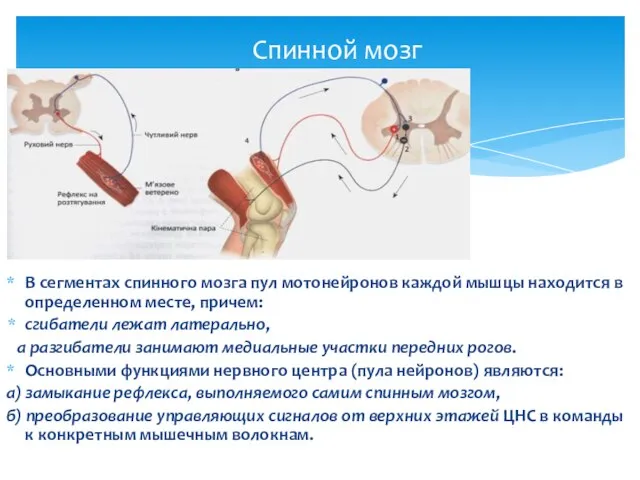
Спинной мозг
В сегментах спинного мозга пул мотонейронов каждой мышцы находится в
Спинной мозг
В сегментах спинного мозга пул мотонейронов каждой мышцы находится в
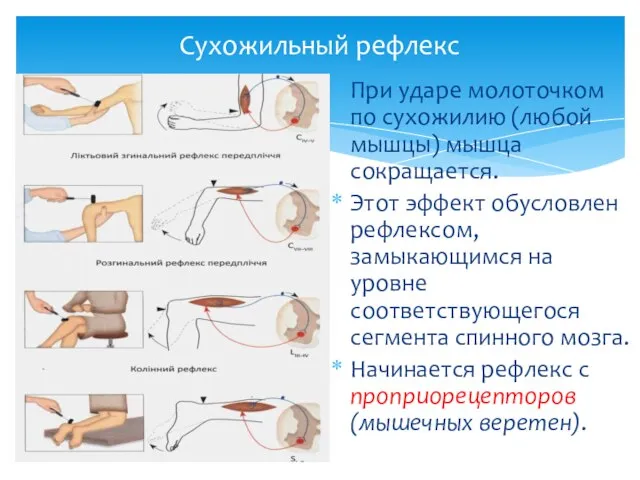
Сухожильный рефлекс
При ударе молоточком по сухожилию (любой мышцы) мышца сокращается.
Этот эффект
Сухожильный рефлекс
При ударе молоточком по сухожилию (любой мышцы) мышца сокращается.
Этот эффект
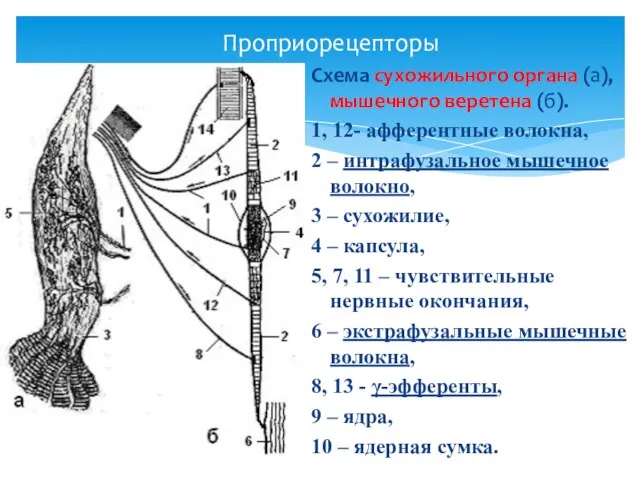
Проприорецепторы
Схема сухожильного органа (а),
мышечного веретена (б).
1, 12- афферентные
Проприорецепторы
Схема сухожильного органа (а),
мышечного веретена (б).
1, 12- афферентные

Количество мышечных веретен и сухожильных органов в различных мышцах неодинаково. Чем
Количество мышечных веретен и сухожильных органов в различных мышцах неодинаково. Чем
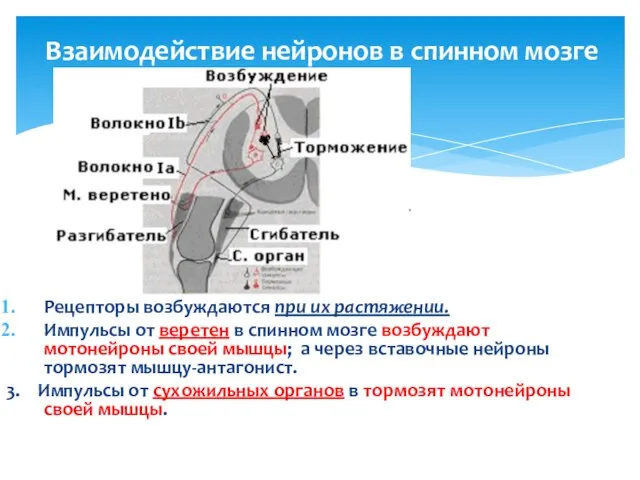
Взаимодействие нейронов в спинном мозге
Рецепторы возбуждаются при их растяжении.
Импульсы от веретен
Взаимодействие нейронов в спинном мозге
Рецепторы возбуждаются при их растяжении.
Импульсы от веретен
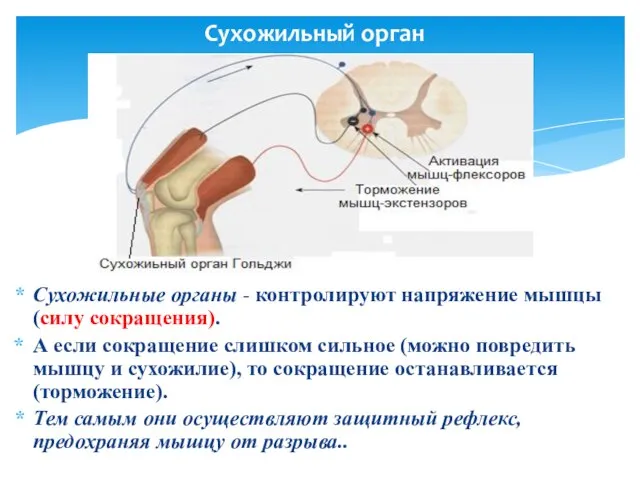
Сухожильный орган
Cухожильные органы - контролируют напряжение мышцы (силу сокращения).
А если
Сухожильный орган
Cухожильные органы - контролируют напряжение мышцы (силу сокращения).
А если
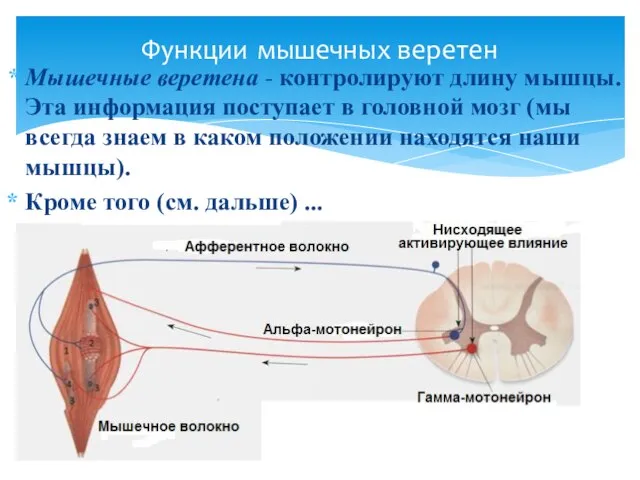
Мышечные веретена - контролируют длину мышцы. Эта информация поступает в головной
Мышечные веретена - контролируют длину мышцы. Эта информация поступает в головной

В естественных условиях возбуждение α- и γ-мотонейронов одной мышцы возникает чаще
В естественных условиях возбуждение α- и γ-мотонейронов одной мышцы возникает чаще
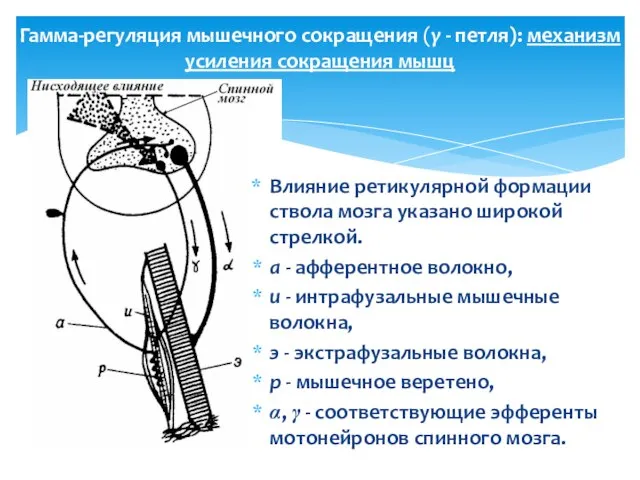
Гамма-регуляция мышечного сокращения (γ - петля): механизм усиления сокращения мышц
Влияние
Гамма-регуляция мышечного сокращения (γ - петля): механизм усиления сокращения мышц
Влияние

Рефлексы спинного мозга
(начинаются с различных рецепторов или автономных программ)
Сегментарные:
моносинаптические и
полисинаптические.
Межсегментарные:
Сгибательный,
Перекрестный
разгибательный,
Чесательный,
Шагательный.
Кроме
Рефлексы спинного мозга
(начинаются с различных рецепторов или автономных программ)
Сегментарные:
моносинаптические и
полисинаптические.
Межсегментарные:
Сгибательный,
Перекрестный
разгибательный,
Чесательный,
Шагательный.
Кроме

Центры вышележащих отделов мозга координируют рефлексы спинного мозга.
А ретикулярная формация ствола
Центры вышележащих отделов мозга координируют рефлексы спинного мозга.
А ретикулярная формация ствола
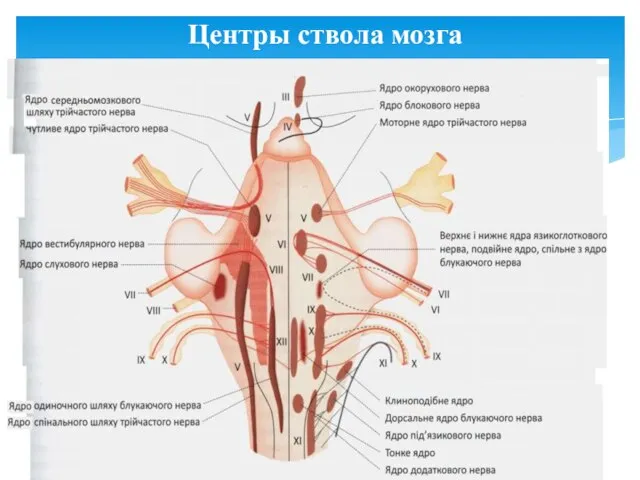
Центры ствола мозга
Центры ствола мозга
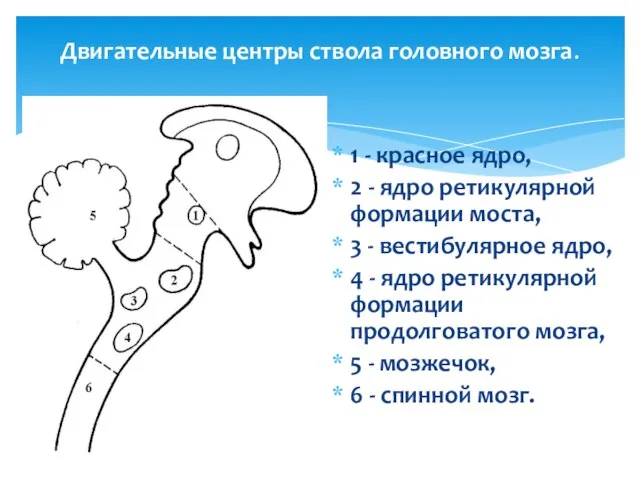
Двигательные центры ствола головного мозга.
1 - красное ядро,
2 -
Двигательные центры ствола головного мозга.
1 - красное ядро,
2 -

В ориентации человека в пространстве помимо проприорецепторов большую роль играет вестибулярная
В ориентации человека в пространстве помимо проприорецепторов большую роль играет вестибулярная

Лабиринт
1 - эндолимфатический мешочек,
2, 3, 4 - полукружные каналы,
5
Лабиринт
1 - эндолимфатический мешочек,
2, 3, 4 - полукружные каналы,
5
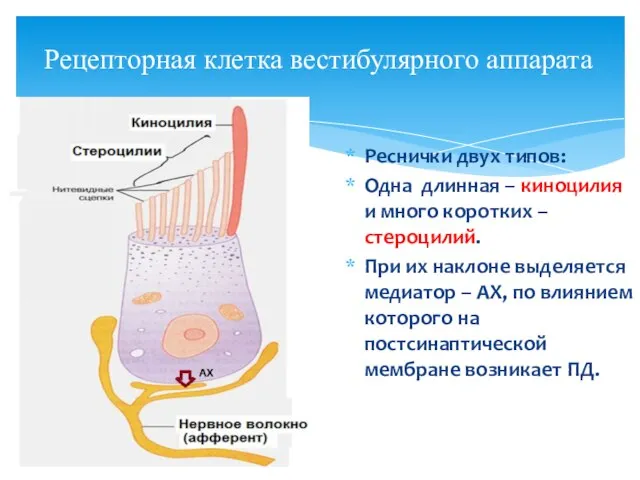
Рецепторная клетка вестибулярного аппарата
Реснички двух типов:
Одна длинная – киноцилия и
Рецепторная клетка вестибулярного аппарата
Реснички двух типов:
Одна длинная – киноцилия и
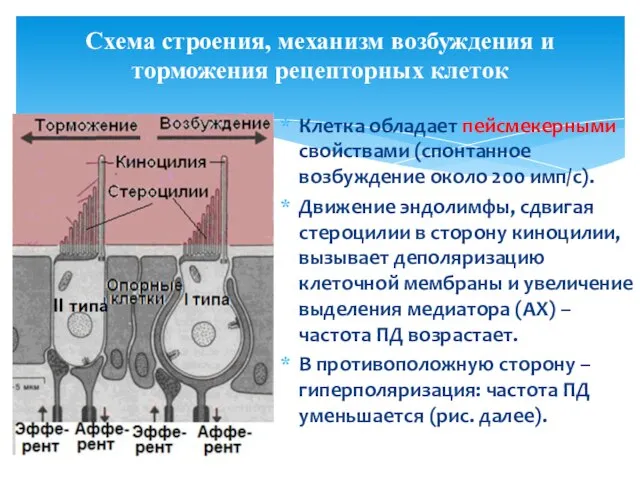
Схема строения, механизм возбуждения и торможения рецепторных клеток
Клетка обладает пейсмекерными свойствами
Схема строения, механизм возбуждения и торможения рецепторных клеток
Клетка обладает пейсмекерными свойствами
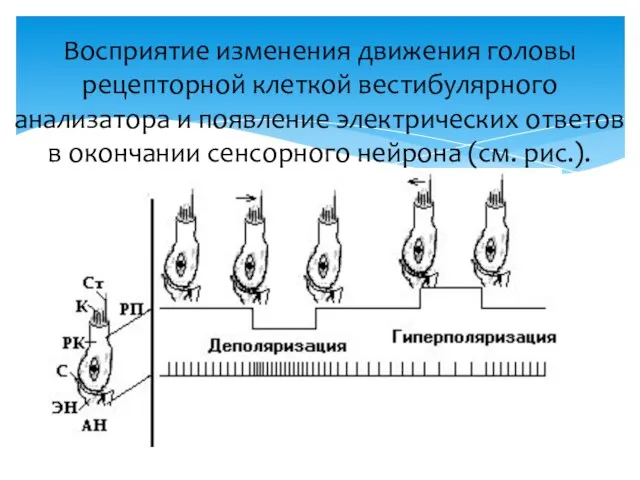
Восприятие изменения движения головы рецепторной клеткой вестибулярного анализатора и появление электрических
Восприятие изменения движения головы рецепторной клеткой вестибулярного анализатора и появление электрических
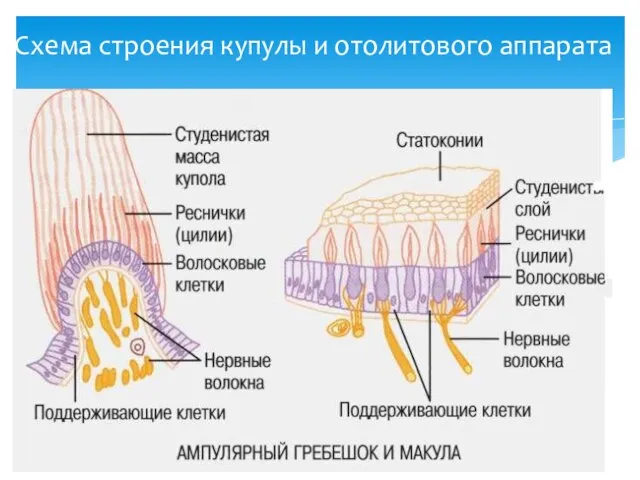
Схема строения купулы и отолитового аппарата
Схема строения купулы и отолитового аппарата
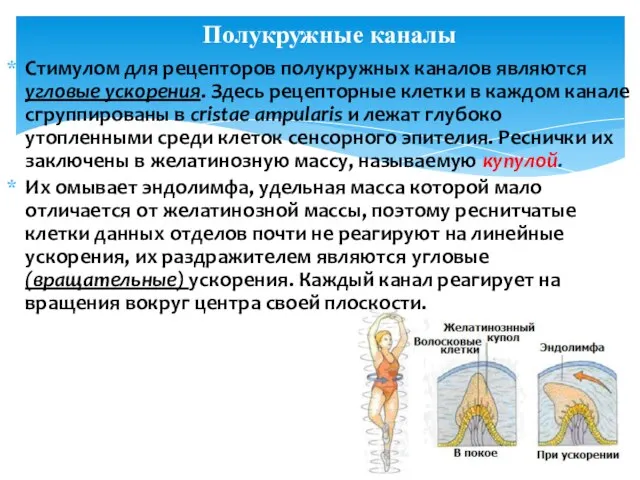
Стимулом для рецепторов полукружных каналов являются угловые ускорения. Здесь рецепторные клетки
Стимулом для рецепторов полукружных каналов являются угловые ускорения. Здесь рецепторные клетки

Раздражаются они при изменении скорости движения (начало и конец движений), при
Раздражаются они при изменении скорости движения (начало и конец движений), при
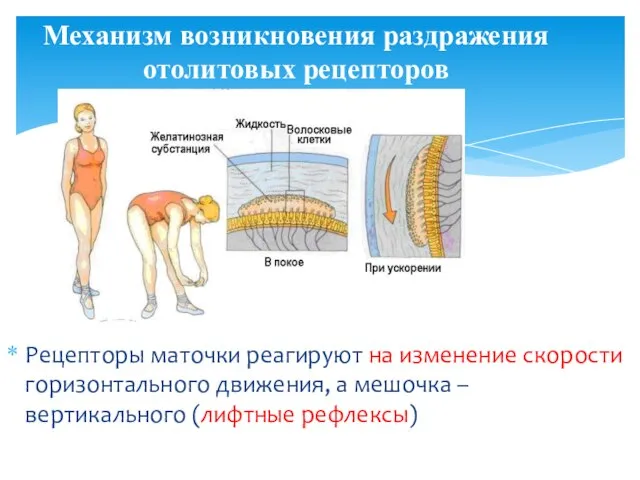
Рецепторы маточки реагируют на изменение скорости горизонтального движения, а мешочка –
Рецепторы маточки реагируют на изменение скорости горизонтального движения, а мешочка –

Прежде всего: все центры ствола тесно связаны между собой.
Рефлексы стволовых моторных
Прежде всего: все центры ствола тесно связаны между собой.
Рефлексы стволовых моторных

Стволовые центры обеспечивают сохранение равновесия и нормальное вертикальное положение тела в
Стволовые центры обеспечивают сохранение равновесия и нормальное вертикальное положение тела в
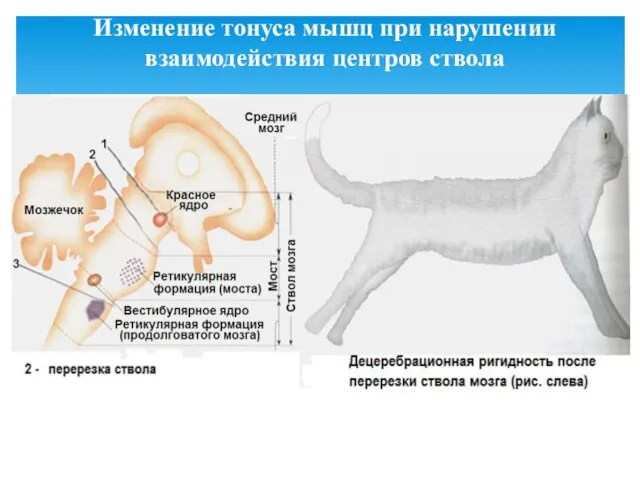
Изменение тонуса мышц при нарушении взаимодействия центров ствола
Изменение тонуса мышц при нарушении взаимодействия центров ствола

Рефлексы ствола мозга
Позные рефлексы:
Шейные,
Вестибулярные,
глазо-двигательные.
Статокинетические рефлексы:
Статические,
Тонические,
Ориентировочные.
Начинаются
Рефлексы ствола мозга
Позные рефлексы:
Шейные,
Вестибулярные,
глазо-двигательные.
Статокинетические рефлексы:
Статические,
Тонические,
Ориентировочные.
Начинаются

При «неправильном» положении головы с проприорецепторов мышц шеи запускаются движения мышц
При «неправильном» положении головы с проприорецепторов мышц шеи запускаются движения мышц
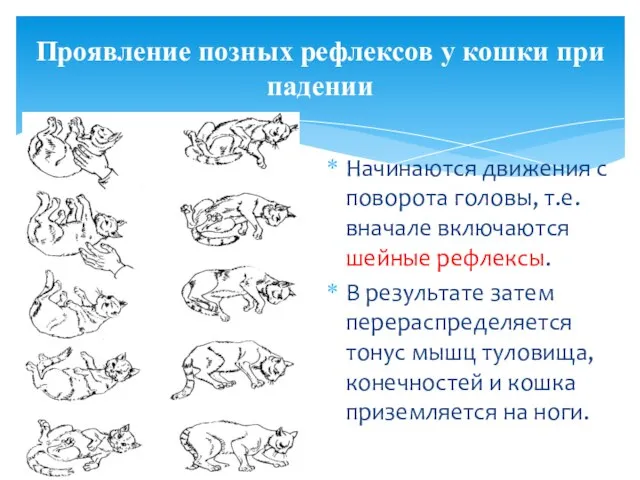
Проявление позных рефлексов у кошки при падении
Начинаются движения с поворота
Проявление позных рефлексов у кошки при падении
Начинаются движения с поворота

Перераспределение тонуса мышц кошки при наклонах головы
У животных при наклоне головы
Перераспределение тонуса мышц кошки при наклонах головы
У животных при наклоне головы

При наклоне головы вперед повышается тонус мышц-сгибателей и в результате улучшается
При наклоне головы вперед повышается тонус мышц-сгибателей и в результате улучшается

При повороте головы повышается тонус мышц сгибателей на стороне поворота, а
При повороте головы повышается тонус мышц сгибателей на стороне поворота, а
 Культурные растения моего дома. Выполнил ученик 5Б класса МОУ «СОШ №6» Павлюткин Никита.
Культурные растения моего дома. Выполнил ученик 5Б класса МОУ «СОШ №6» Павлюткин Никита. Эволюция. Систематика
Эволюция. Систематика Врожденные формы поведения
Врожденные формы поведения Методы изучения регуляторных районов генов
Методы изучения регуляторных районов генов Чешуекрылые насекомые
Чешуекрылые насекомые Клеточная теория
Клеточная теория Многообразие водорослей
Многообразие водорослей  Жизнедеятельность клетки Презентация подготовлена Кассихиной Татьяной Александровной - учителем биологии, химии МОУ СОШ д.Горо
Жизнедеятельность клетки Презентация подготовлена Кассихиной Татьяной Александровной - учителем биологии, химии МОУ СОШ д.Горо Жизнь хомяка Джеффи
Жизнь хомяка Джеффи Общее представление о строении ЦНС
Общее представление о строении ЦНС Чистопородное разведение
Чистопородное разведение ТИП ПЛОСКИЕ ЧЕРВИ
ТИП ПЛОСКИЕ ЧЕРВИ  Три месяца весны
Три месяца весны Презентация на тему "Питание и здоровье" - скачать презентации по Биологии
Презентация на тему "Питание и здоровье" - скачать презентации по Биологии Наследственность и здоровье
Наследственность и здоровье Презентация на тему "Фуксия." - скачать бесплатно презентации по Биологии
Презентация на тему "Фуксия." - скачать бесплатно презентации по Биологии Презентация на тему "Работа мышц" - скачать бесплатно презентации по Биологии_
Презентация на тему "Работа мышц" - скачать бесплатно презентации по Биологии_ Красная книга растений Челябинской области. Подготовила учитель биологии МОУ СОШ № 101 Пылина Т.А.
Красная книга растений Челябинской области. Подготовила учитель биологии МОУ СОШ № 101 Пылина Т.А.  Модификационная изменчивость
Модификационная изменчивость Маисовые полоза. Обзор комбинированных морф
Маисовые полоза. Обзор комбинированных морф Синичкин день
Синичкин день Отряд Стрекозы
Отряд Стрекозы О - ханами
О - ханами СРС НА ТЕМУ: Гипоталамо-гипофизарно-надпочечниковая система Карагандинский Государственный Медицинский Университет Кафедра фи
СРС НА ТЕМУ: Гипоталамо-гипофизарно-надпочечниковая система Карагандинский Государственный Медицинский Университет Кафедра фи Зоопсихология (психология животных)
Зоопсихология (психология животных) Презентация на тему "Строение органоидов клетки" - скачать презентации по Биологии
Презентация на тему "Строение органоидов клетки" - скачать презентации по Биологии Железы внутренней секреции
Железы внутренней секреции Значение опорно-двигательного аппарата
Значение опорно-двигательного аппарата