- Проведение потенциалов в мембранах нервных клеток

Содержание
- 2. Рецепторный потенциал фоторецептора Под действием энергии раздражителя в сенсорных нейронах, связанных с рецепторами (или в самих
- 3. Рецепторный потенциал волосковой клетки
- 4. Рецепторный потенциал волосковой клетки Волосковая клетка деполяризуется при отклонении стереоцилий в одну сторону и гиперполяризуется при
- 5. Рецепторный потенциал волосковой клетки Рецепторный потенциал обычно сохраняется в течение действия стимула, немного уменьшаясь по амплитуде.
- 6. Постсинаптический потенциал возникает в дендритах и соме нейронов в результате синаптической передачи - открытия лиганд-зависимых каналов.
- 7. Потенциал действия возникает в основном в проводящих структурах – аксонах нейронов, а иногда и в дендритах
- 8. Потенциал действия Для передачи информации (возбуждения) на большие расстояния в ЦНС генерируются ПД, которые распространяются по
- 11. Пассивное распространение электрических сигналов
- 12. Пассивное распространение электрических сигналов
- 15. Постоянная длины волокна (λ) Падение амплитуды потенциала носит экспоненциальный характер, так что потенциал (Vx) при данном
- 16. Постоянная длины волокна (λ) Максимальная разность потенциалов V0 определяется величиной стимулирующего тока. Уменьшение разности потенциалов определяется
- 17. Постоянная длины волокна (λ) λ возрастает с увеличением сопротивления мембраны rm, препятствующего утечке тока во внеклеточное
- 18. Пассивное распространение электрических сигналов При пассивном распространении сигналов по нервным клеткам определяющими факторами являются емкость (cm
- 19. Удельное сопротивление Рассчитанные значения rm и ri, характеризуют сопротивление модельного цилиндрического сегмента аксона длиной 1 см.
- 20. Удельное сопротивление мембраны Удельное сопротивление Rm отражает сопротивление мембраны площадью 1 см2 и измеряется в Ом•см2.
- 21. Удельное сопротивление аксоплазмы Удельным сопротивлением аксоплазмы Ri считают внутреннее продольное сопротивление сегмента аксона длиной 1 см
- 22. Влияние диаметра кабеля на постоянную длины волокна = (rm/ri )½, rm = Rm / (2πα )
- 23. Зависимость λ от диаметра волокна С увеличением диаметра волокна потенциалы затухают на одну и ту же
- 24. Зависимость λ от диаметра волокна Объект Диаметр (мкм) λ (мм) Аксон кальмара 1000 13 Мышечное волокно
- 25. Постоянная времени (τ ) Мембрана обладает свойством емкости, накапливая заряд на своей внешней и внутренней поверхности.
- 26. Постоянная времени (τ ) Из-за емкости мембраны фаза роста амплитуды потенциала в ответ на ток развивается
- 27. Постоянная времени (τ ) Ii = Im - Ic
- 28. Постоянная времени (τ ) Замедление фаз роста и спада потенциалов с удалением от места раздражения (сверху
- 33. Распространение нервного импульса по немиелинизированному волокну
- 34. Распространение нервного импульса по немиелинизированному волокну
- 36. Скорость проведения возбуждения по нервному волокну Скорость проведения V ~ λ2 ~ 1/τ3 Поскольку λ ~
- 37. Зависимость λ от диаметра волокна Объект Диаметр (мкм) λ (мм) Аксон кальмара 1000 13 Мышечное волокно
- 39. Распространение нервных импульсов по миелинизированному волокну Миелиновая оболочка прерывается участками без миелина (перехватами Ранвье). Расстояние между
- 40. Advantages of myelin for conductance of spikes. Myelin increases resistance by factor of 5,000. Decreases capacitance
- 43. Распространение нервных импульсов по миелинизированному волокну Миелинизированные аксоны способны проводить нервные импульсы с более высокой частотой
- 44. Распространение нервных импульсов по миелинизированному волокну
- 45. Распространение нервных импульсов по миелинизирован-ному волокну Сальтаторное проведение ПД по миелинизированному волокну происходит значительно быстрее, чем
- 46. Длина участка между перехватами Ранвье
- 47. Длина участка между перехватами Ранвье С образованием нового слоя (миелиновой оболочки) поперечное сопротивление мембраны Rм увеличивается,
- 48. Длина участка между перехватами Ранвье L/D у волокон лягушки-быка ~ 205, в нервах боковой линии Rana
- 49. Длина участка между перехватами Ранвье В области перехвата цилиндр у толстых волокон сужается, а у тонких
- 50. Длина участка между перехватами Ранвье Соотношение диаметра волокна (Fibre diameter) и длины межперехватного участка (Internodal distance)
- 51. Распространение нервных импульсов по миелинизированному волокну Скорость проведения любого электрического кабеля зависит от соотношения поперечного (Rm)
- 52. Распространение нервных импульсов по миелинизированному волокну Оптимальная толщина миелина достигается, когда диаметр аксона составляет около 0,7
- 53. Скорость распространение нервных импульсов по миелинизированному волокну Зависимость скорости от диаметра волокна
- 54. Распространение нервных импульсов по миелинизированному волокну Постоянство соотношения d/D справедливо только для волокон диаметром (D) >
- 55. Скорость распространение нервных импульсов по миелинизированному волокну
- 56. Распространение нервных импульсов по миелинизированному волокну Позже были найдены миелинизированные волокна с D=0,2 мкм Соотношение d/D
- 57. Распространение нервных импульсов по миелинизированному волокну После некоторого критического значения диаметра скорость проведения по миелинизированному волокну
- 58. Распространение нервных импульсов по миелинизированному волокну После некоторого критического значения диаметра скорость проведения по миелинизированному волокну
- 59. Ионные каналы в миелинизированных волокнах В миелинизированных волокнах Na+-каналы сконцентрированы в перехватах Ранвье, а К+-каналы локализованы
- 60. Ионные каналы в миелинизированных волокнах В миелинизированных волокнах Na+-каналы сконцентрированы в перехватах Ранвье, а К+-каналы локализованы
- 61. Ионные каналы в миелинизированных волокнах Мембрана перехвата Ранвье в аксоне (Node) содержит только Na+-каналы (Nav1.6), K+-каналы
- 62. Распределение ионных каналов в различных функциональных частях миелинизированных аксонов
- 63. Ионные каналы в миелинизированных волокнах При возбуждении в перехватах Ранвье возникает лишь потенциал-зависимый входящий Na+-ток, а
- 64. .
- 65. Концентрация ионов Na+ в разных частях миелинизированного аксона при его возбуждении Changes in intracellular Na+ during
- 66. Амплитуда Na+-токов в разных частях миелинизированного аксона Left) Top, schematic diagram of the outside-out recording from
- 68. Фактор надежности проведения ПД Фактор надежности проведения – это отношение амплитуды деполяризации, которую создает ток активированного
- 69. Фактор надежности проведения ПД На участках, где исчезает миелиновая оболочка (в непосредственной близости от аксонной терминали
- 70. Фактор надежности проведения ПД Снижение плотности тока на мембране производит меньшую деполяризацию, чем в перехватах Ранвье.
- 75. Скачать презентацию
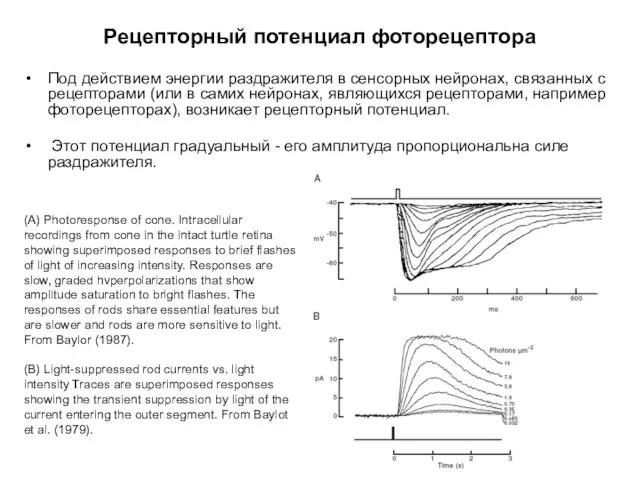
Рецепторный потенциал фоторецептора
Под действием энергии раздражителя в сенсорных нейронах, связанных с
Рецепторный потенциал фоторецептора
Под действием энергии раздражителя в сенсорных нейронах, связанных с
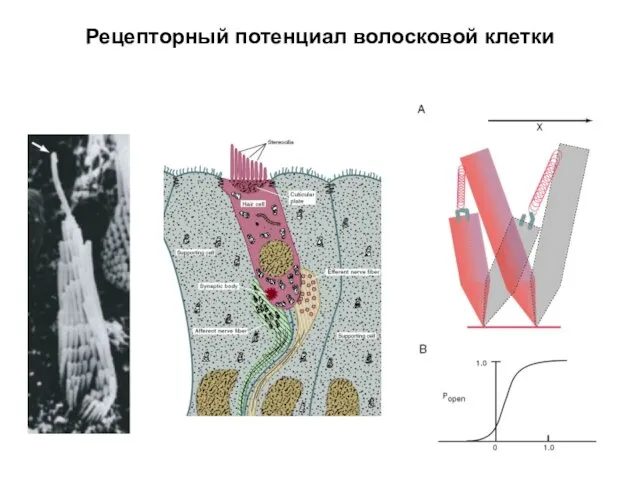
Рецепторный потенциал волосковой клетки
Рецепторный потенциал волосковой клетки
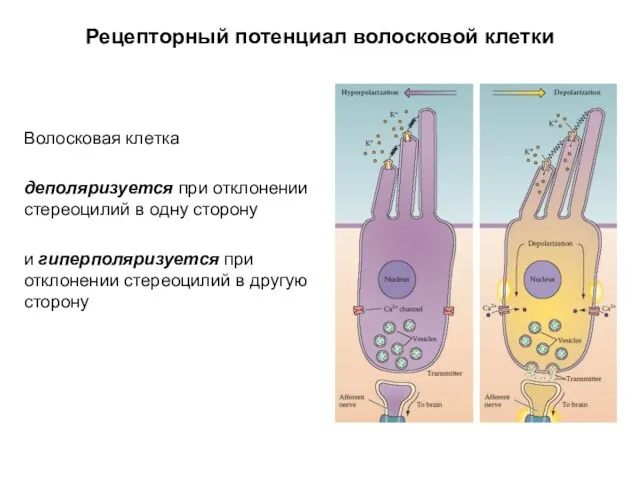
Рецепторный потенциал волосковой клетки
Волосковая клетка
деполяризуется при отклонении стереоцилий в одну
Рецепторный потенциал волосковой клетки
Волосковая клетка
деполяризуется при отклонении стереоцилий в одну
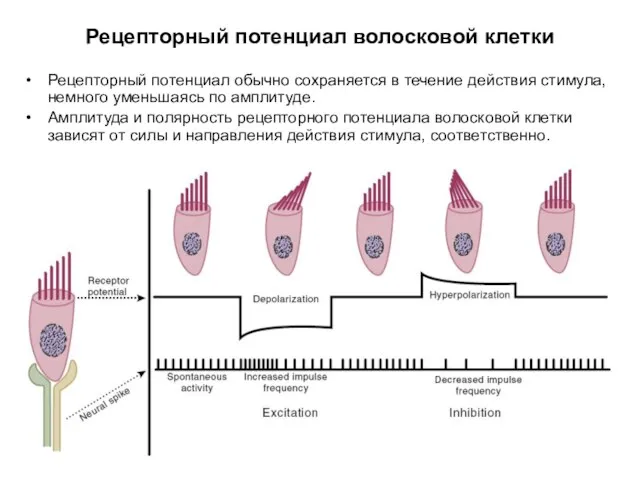
Рецепторный потенциал волосковой клетки
Рецепторный потенциал обычно сохраняется в течение действия стимула,
Рецепторный потенциал волосковой клетки
Рецепторный потенциал обычно сохраняется в течение действия стимула,
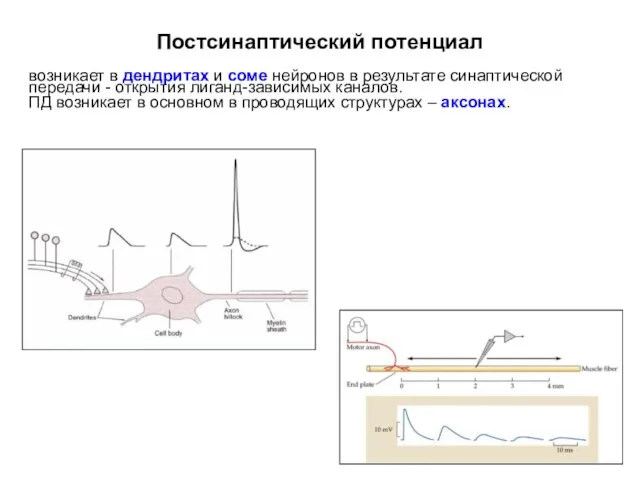
Постсинаптический потенциал
возникает в дендритах и соме нейронов в результате синаптической передачи
Постсинаптический потенциал
возникает в дендритах и соме нейронов в результате синаптической передачи
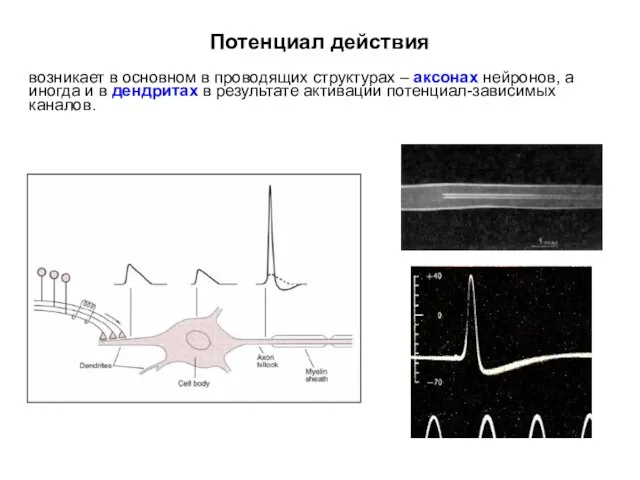
Потенциал действия
возникает в основном в проводящих структурах – аксонах нейронов, а
Потенциал действия
возникает в основном в проводящих структурах – аксонах нейронов, а

Потенциал действия
Для передачи информации (возбуждения) на большие расстояния в ЦНС генерируются
Потенциал действия
Для передачи информации (возбуждения) на большие расстояния в ЦНС генерируются
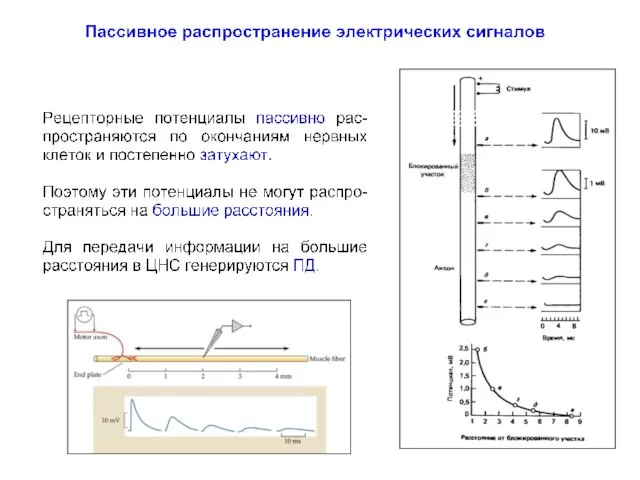
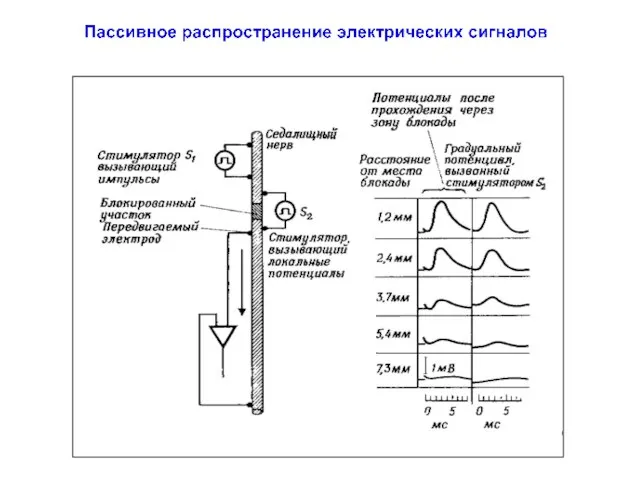
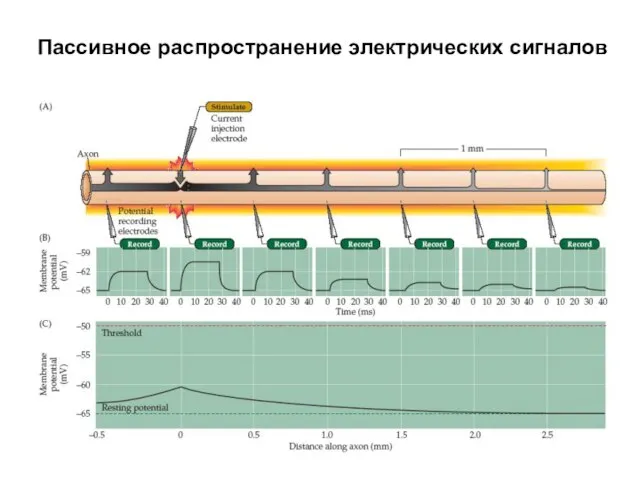
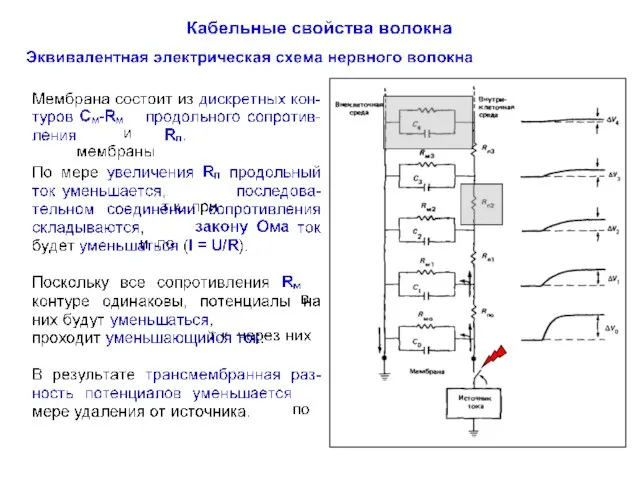
Пассивное распространение электрических сигналов
Пассивное распространение электрических сигналов
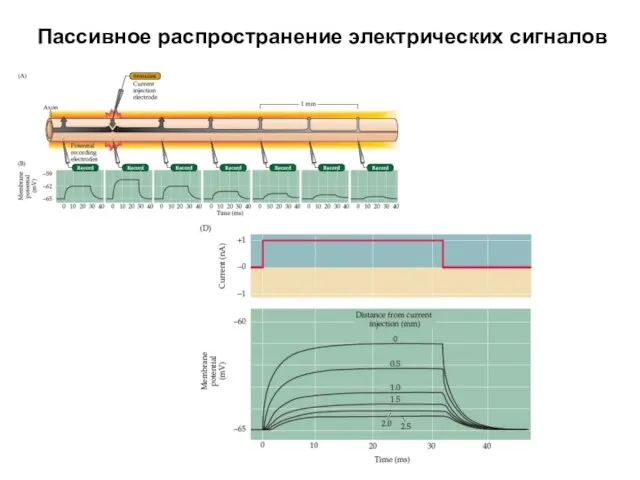
Пассивное распространение электрических сигналов
Пассивное распространение электрических сигналов
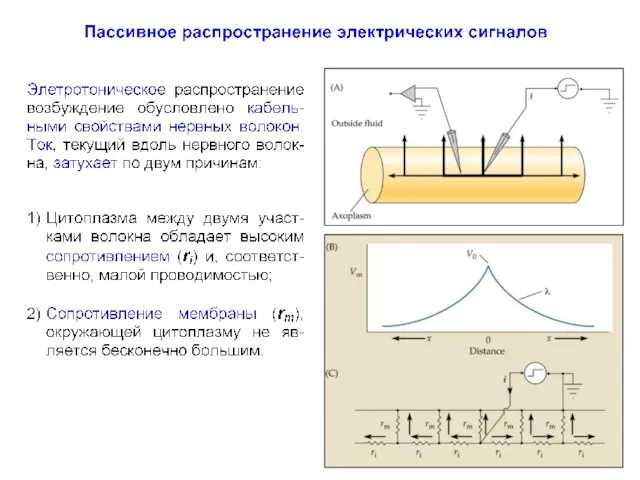
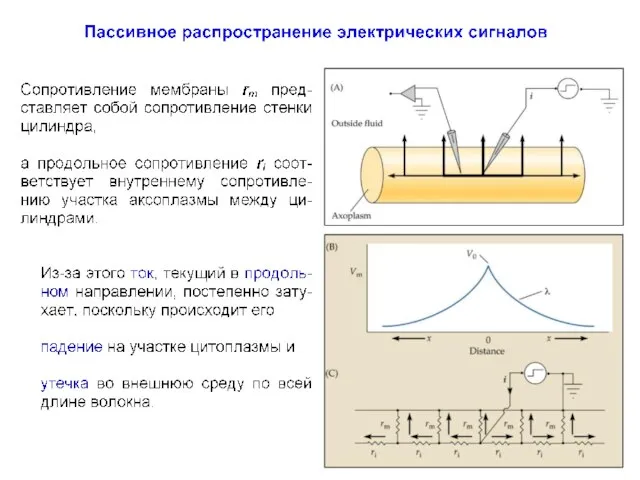
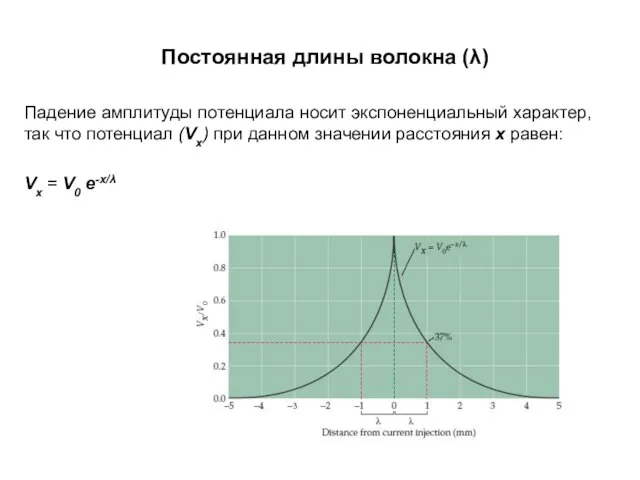
Постоянная длины волокна (λ)
Падение амплитуды потенциала носит экспоненциальный характер, так что
Постоянная длины волокна (λ)
Падение амплитуды потенциала носит экспоненциальный характер, так что
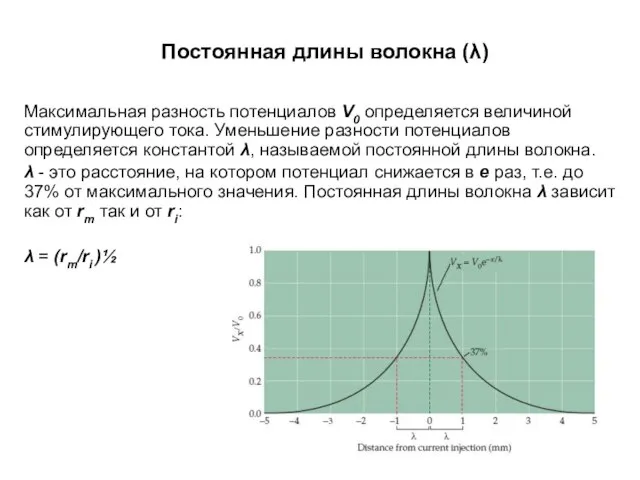
Постоянная длины волокна (λ)
Максимальная разность потенциалов V0 определяется величиной стимулирующего тока.
Постоянная длины волокна (λ)
Максимальная разность потенциалов V0 определяется величиной стимулирующего тока.
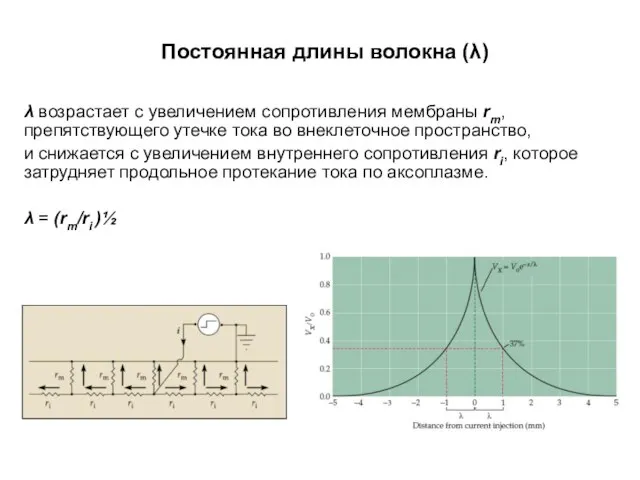
Постоянная длины волокна (λ)
λ возрастает с увеличением сопротивления мембраны rm, препятствующего
Постоянная длины волокна (λ)
λ возрастает с увеличением сопротивления мембраны rm, препятствующего
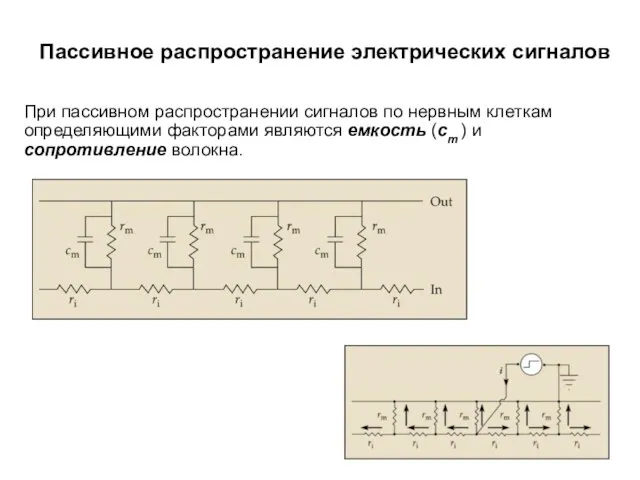
Пассивное распространение электрических сигналов
При пассивном распространении сигналов по нервным клеткам определяющими
Пассивное распространение электрических сигналов
При пассивном распространении сигналов по нервным клеткам определяющими
Удельное сопротивление
Рассчитанные значения rm и ri, характеризуют сопротивление модельного цилиндрического сегмента
Удельное сопротивление
Рассчитанные значения rm и ri, характеризуют сопротивление модельного цилиндрического сегмента
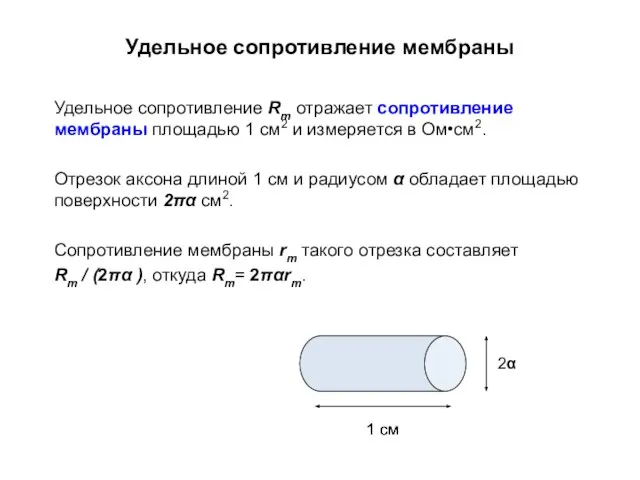
Удельное сопротивление мембраны
Удельное сопротивление Rm отражает сопротивление мембраны площадью 1 см2
Удельное сопротивление мембраны
Удельное сопротивление Rm отражает сопротивление мембраны площадью 1 см2

Удельное сопротивление аксоплазмы
Удельным сопротивлением аксоплазмы Ri считают внутреннее продольное сопротивление сегмента
Удельное сопротивление аксоплазмы
Удельным сопротивлением аксоплазмы Ri считают внутреннее продольное сопротивление сегмента

Влияние диаметра кабеля
на постоянную длины волокна
= (rm/ri )½,
rm =
Влияние диаметра кабеля
на постоянную длины волокна
= (rm/ri )½,
rm =
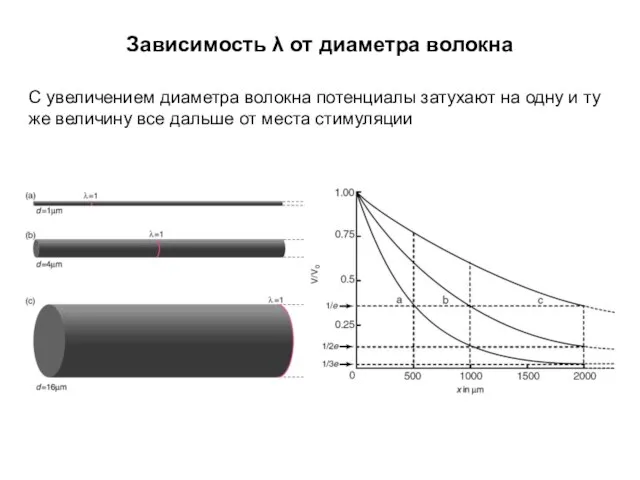
Зависимость λ от диаметра волокна
С увеличением диаметра волокна потенциалы затухают на
Зависимость λ от диаметра волокна
С увеличением диаметра волокна потенциалы затухают на
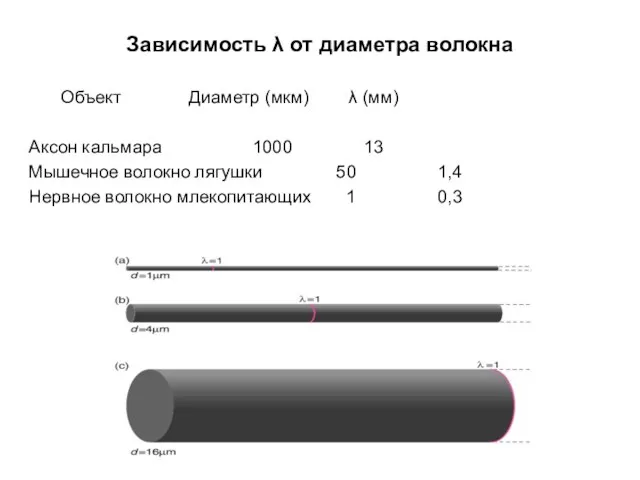
Зависимость λ от диаметра волокна
Объект Диаметр (мкм) λ (мм)
Аксон кальмара 1000 13
Мышечное волокно
Зависимость λ от диаметра волокна
Объект Диаметр (мкм) λ (мм)
Аксон кальмара 1000 13
Мышечное волокно
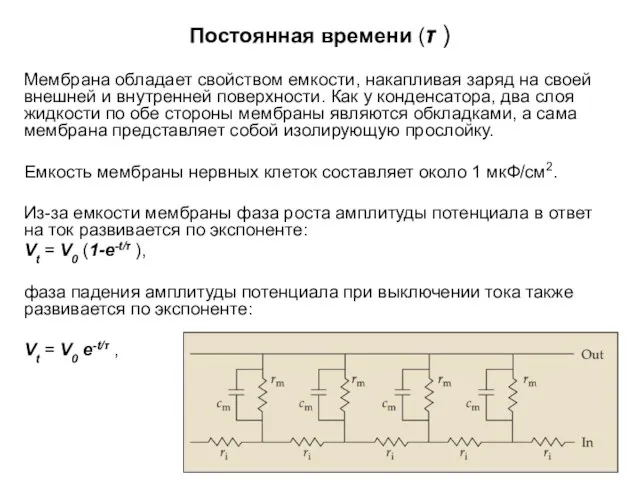
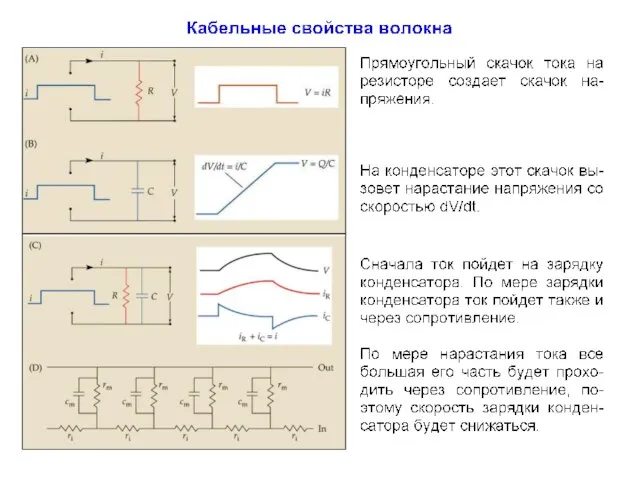
Постоянная времени (τ )
Мембрана обладает свойством емкости, накапливая заряд на своей
Постоянная времени (τ )
Мембрана обладает свойством емкости, накапливая заряд на своей
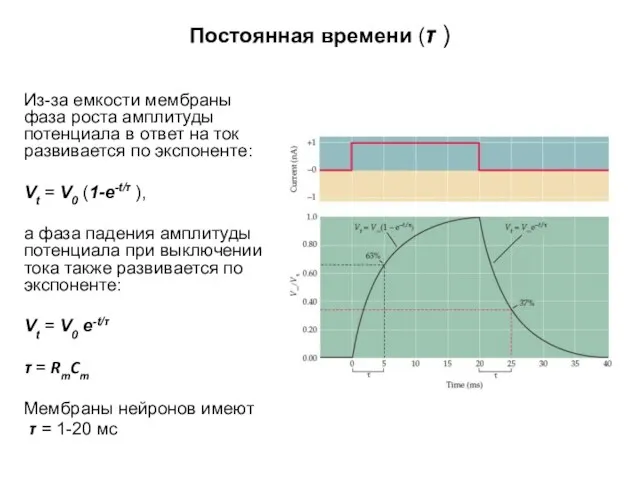
Постоянная времени (τ )
Из-за емкости мембраны фаза роста амплитуды потенциала в
Постоянная времени (τ )
Из-за емкости мембраны фаза роста амплитуды потенциала в
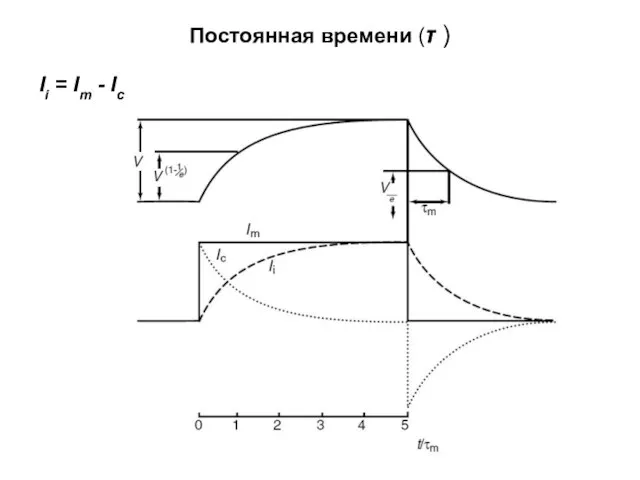
Постоянная времени (τ )
Ii = Im - Ic
Постоянная времени (τ )
Ii = Im - Ic
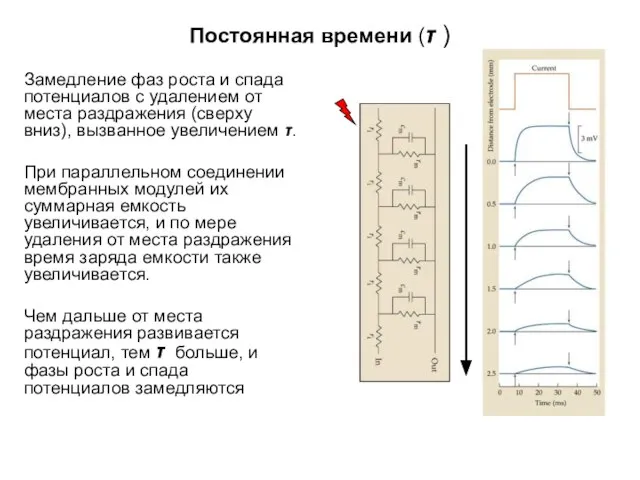
Постоянная времени (τ )
Замедление фаз роста и спада потенциалов с удалением
Постоянная времени (τ )
Замедление фаз роста и спада потенциалов с удалением
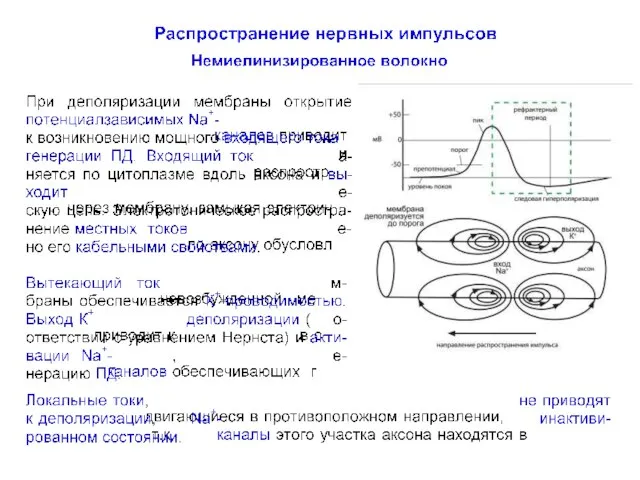
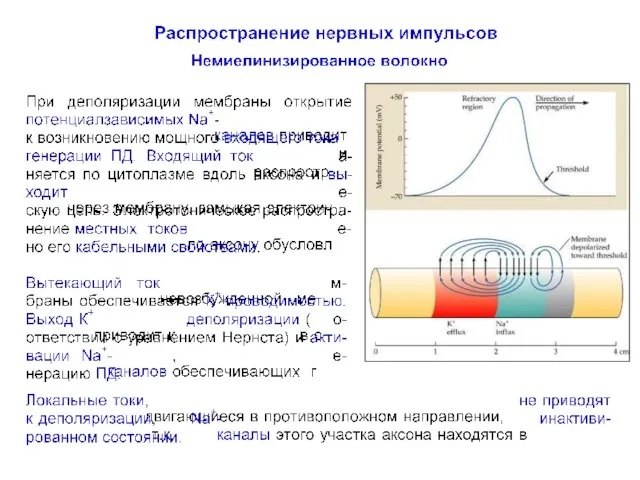
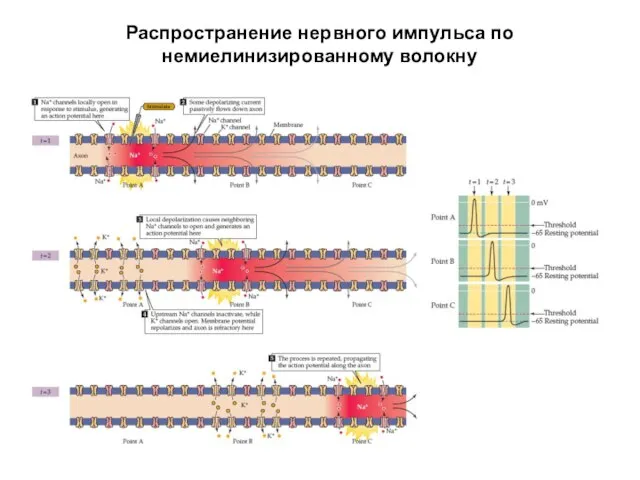
Распространение нервного импульса по немиелинизированному волокну
Распространение нервного импульса по немиелинизированному волокну
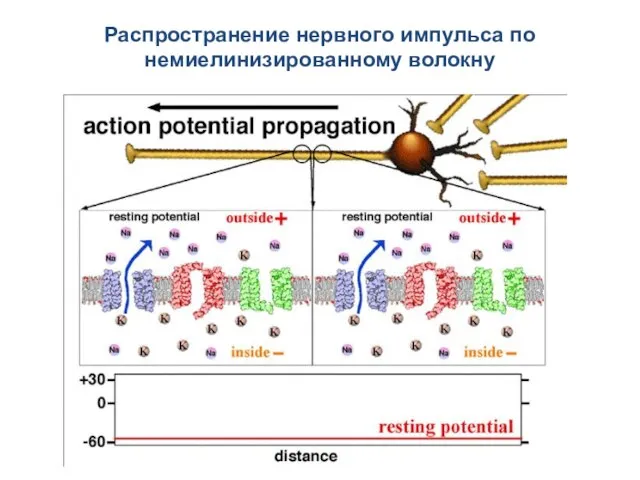
Распространение нервного импульса по немиелинизированному волокну
Распространение нервного импульса по немиелинизированному волокну

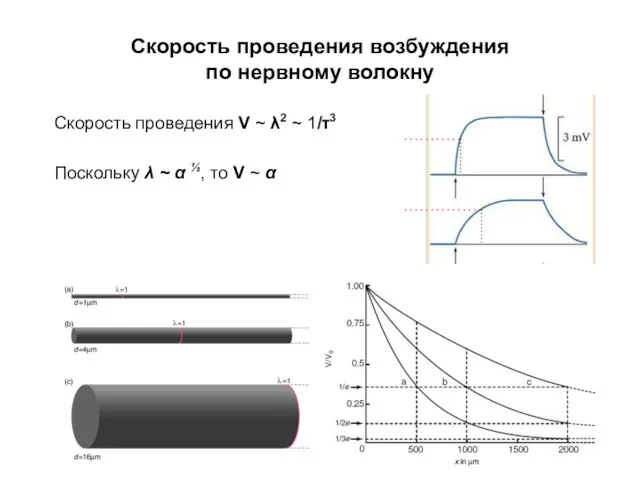
Скорость проведения возбуждения
по нервному волокну
Скорость проведения V ~ λ2 ~ 1/τ3
Поскольку
Скорость проведения возбуждения
по нервному волокну
Скорость проведения V ~ λ2 ~ 1/τ3
Поскольку
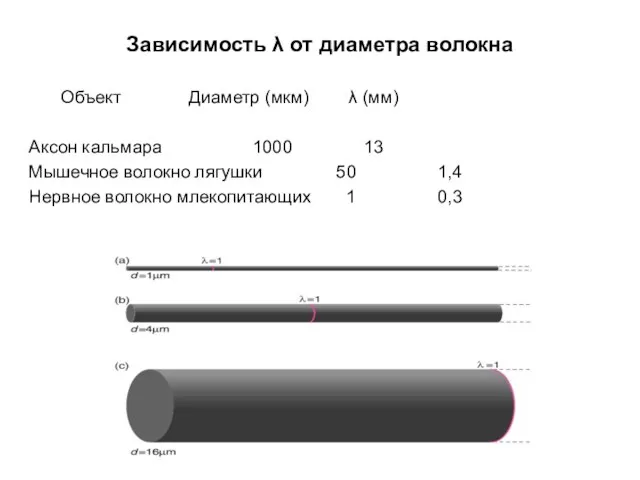
Зависимость λ от диаметра волокна
Объект Диаметр (мкм) λ (мм)
Аксон кальмара 1000 13
Мышечное волокно
Зависимость λ от диаметра волокна
Объект Диаметр (мкм) λ (мм)
Аксон кальмара 1000 13
Мышечное волокно
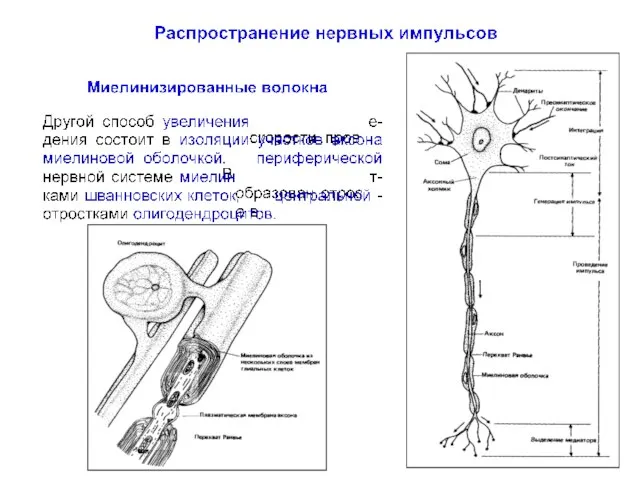
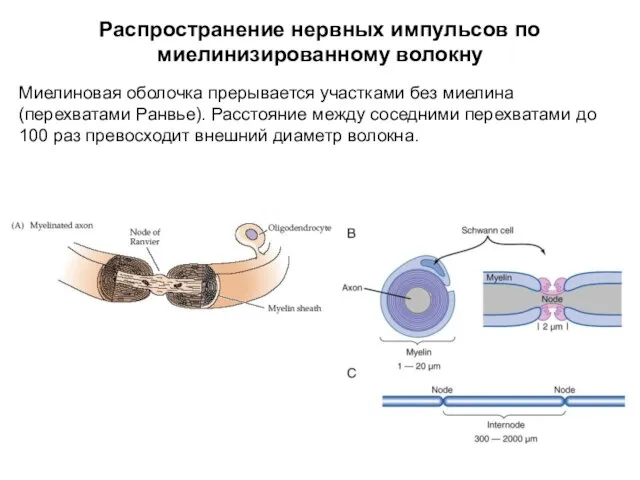
Распространение нервных импульсов по миелинизированному волокну
Миелиновая оболочка прерывается участками без миелина
Распространение нервных импульсов по миелинизированному волокну
Миелиновая оболочка прерывается участками без миелина
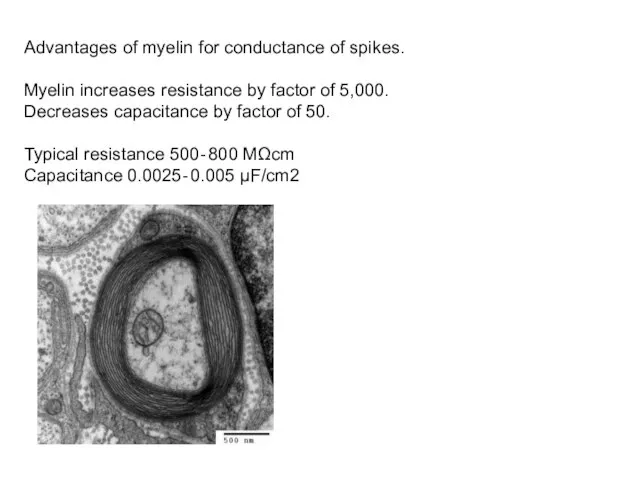
Advantages of myelin for conductance of spikes.
Myelin increases resistance by
Advantages of myelin for conductance of spikes.
Myelin increases resistance by
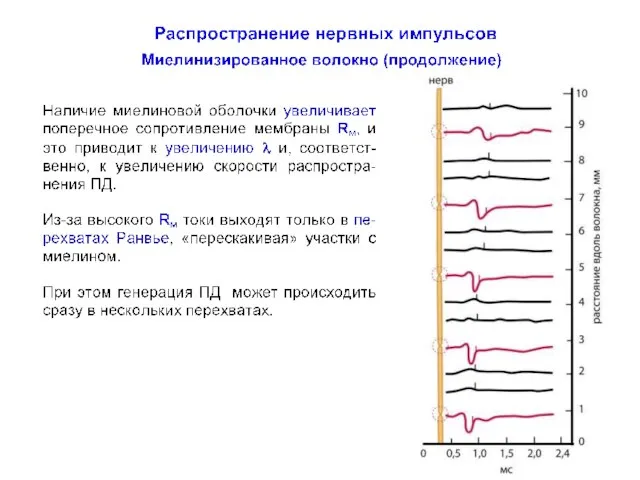
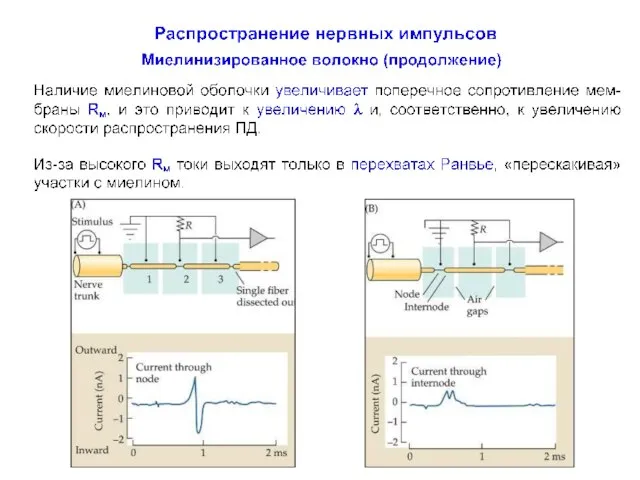

Распространение нервных импульсов по миелинизированному волокну
Миелинизированные аксоны способны проводить нервные импульсы
Распространение нервных импульсов по миелинизированному волокну
Миелинизированные аксоны способны проводить нервные импульсы
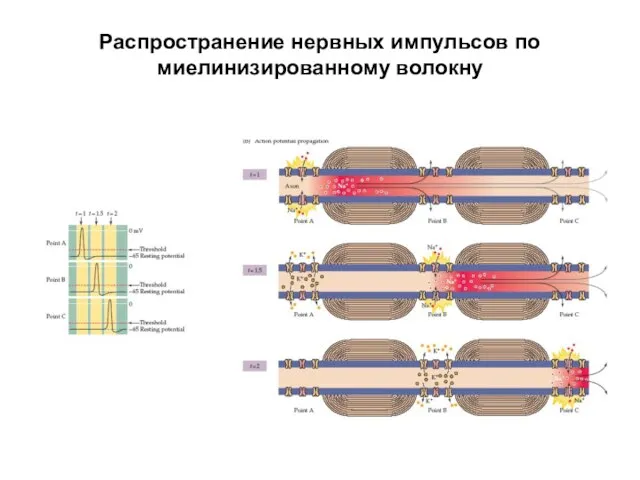
Распространение нервных импульсов по миелинизированному волокну
Распространение нервных импульсов по миелинизированному волокну
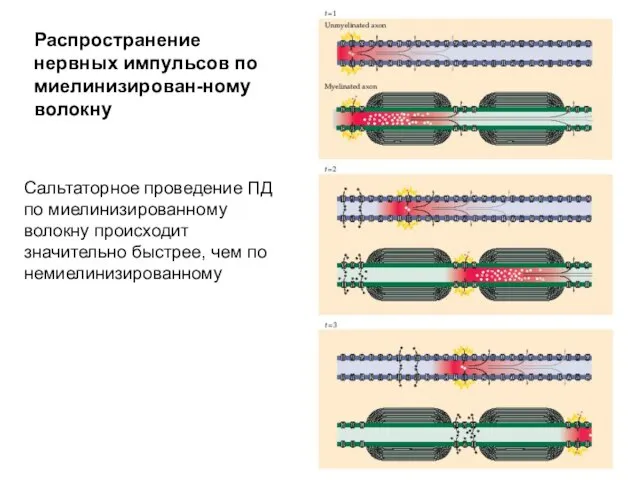
Распространение нервных импульсов по миелинизирован-ному волокну
Сальтаторное проведение ПД по миелинизированному волокну
Распространение нервных импульсов по миелинизирован-ному волокну
Сальтаторное проведение ПД по миелинизированному волокну

Длина участка между перехватами Ранвье
Длина участка между перехватами Ранвье

Длина участка между перехватами Ранвье
С образованием нового слоя (миелиновой оболочки) поперечное
Длина участка между перехватами Ранвье
С образованием нового слоя (миелиновой оболочки) поперечное

Длина участка между перехватами Ранвье
L/D
у волокон лягушки-быка ~ 205,
в нервах боковой
Длина участка между перехватами Ранвье
L/D
у волокон лягушки-быка ~ 205,
в нервах боковой
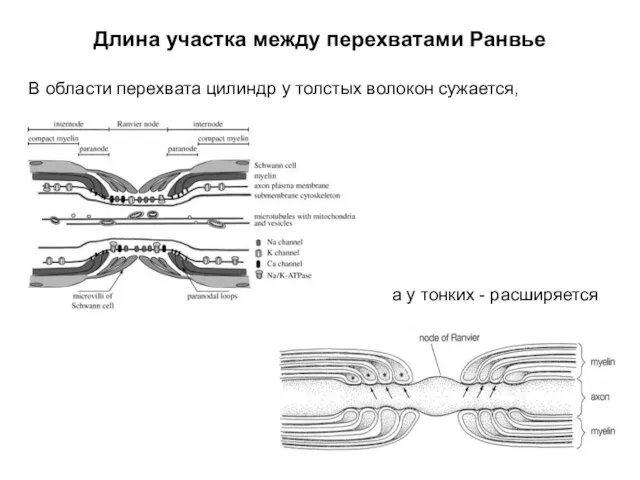
Длина участка между перехватами Ранвье
В области перехвата цилиндр у толстых волокон
Длина участка между перехватами Ранвье
В области перехвата цилиндр у толстых волокон
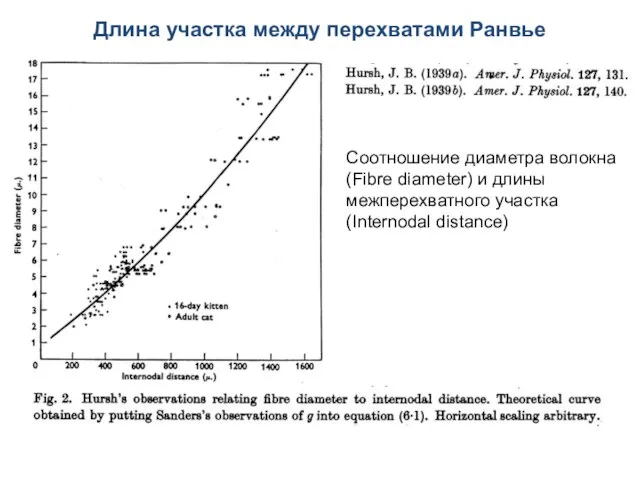
Длина участка между перехватами Ранвье
Соотношение диаметра волокна (Fibre diameter) и длины
Длина участка между перехватами Ранвье
Соотношение диаметра волокна (Fibre diameter) и длины

Распространение нервных импульсов по миелинизированному волокну
Скорость проведения любого электрического кабеля зависит
Распространение нервных импульсов по миелинизированному волокну
Скорость проведения любого электрического кабеля зависит
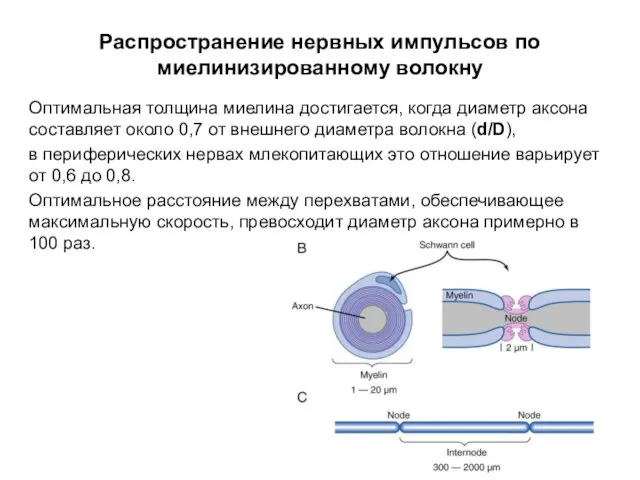
Распространение нервных импульсов по миелинизированному волокну
Оптимальная толщина миелина достигается, когда диаметр
Распространение нервных импульсов по миелинизированному волокну
Оптимальная толщина миелина достигается, когда диаметр
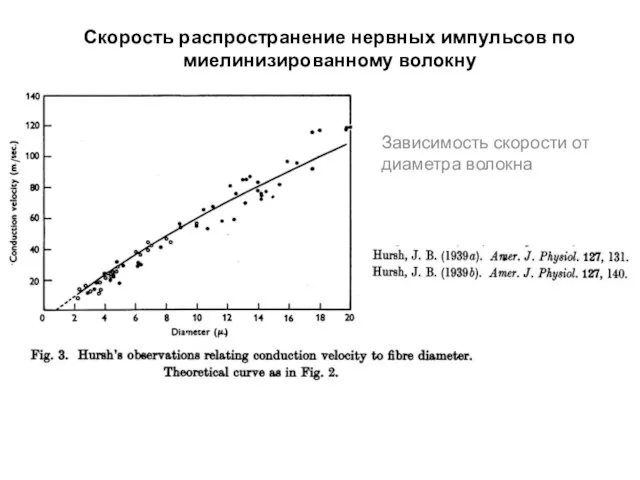
Скорость распространение нервных импульсов по миелинизированному волокну
Зависимость скорости от диаметра волокна
Скорость распространение нервных импульсов по миелинизированному волокну
Зависимость скорости от диаметра волокна
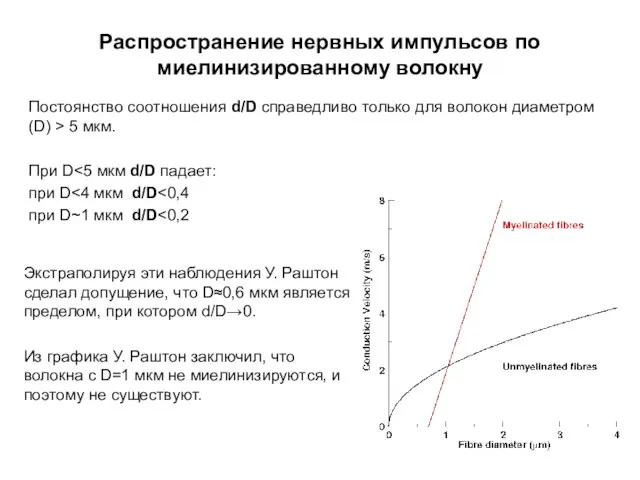
Распространение нервных импульсов по миелинизированному волокну
Постоянство соотношения d/D справедливо только для
Распространение нервных импульсов по миелинизированному волокну
Постоянство соотношения d/D справедливо только для
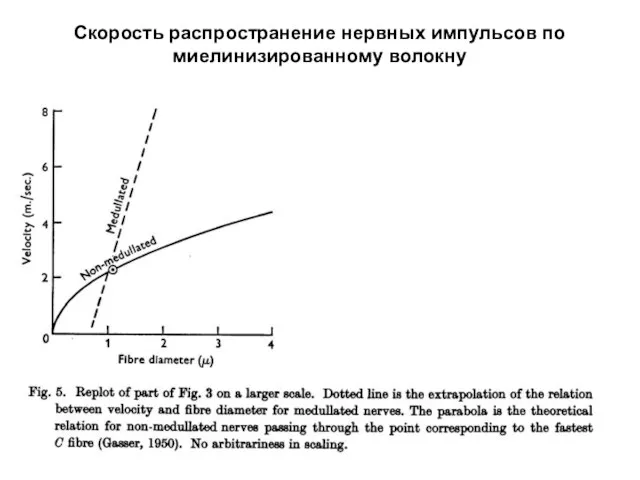
Скорость распространение нервных импульсов по миелинизированному волокну
Скорость распространение нервных импульсов по миелинизированному волокну
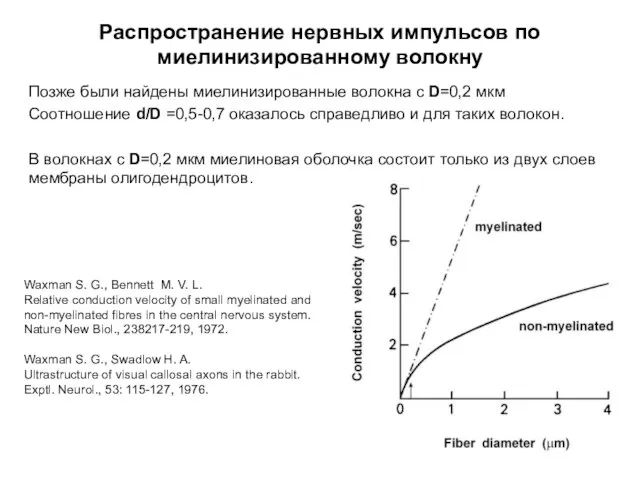
Распространение нервных импульсов по миелинизированному волокну
Позже были найдены миелинизированные волокна с
Распространение нервных импульсов по миелинизированному волокну
Позже были найдены миелинизированные волокна с
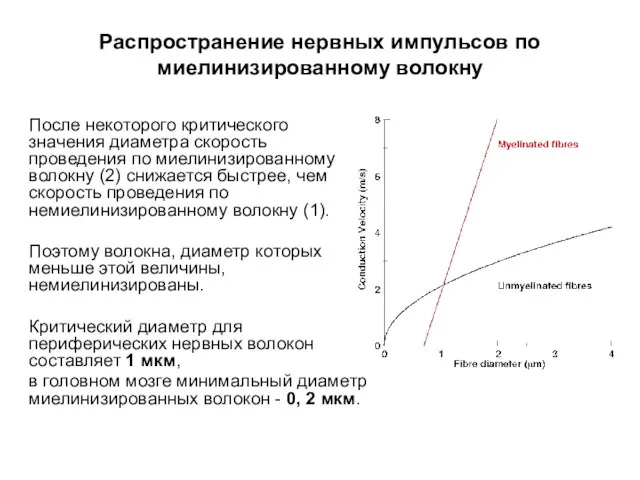
Распространение нервных импульсов по миелинизированному волокну
После некоторого критического значения диаметра скорость
Распространение нервных импульсов по миелинизированному волокну
После некоторого критического значения диаметра скорость
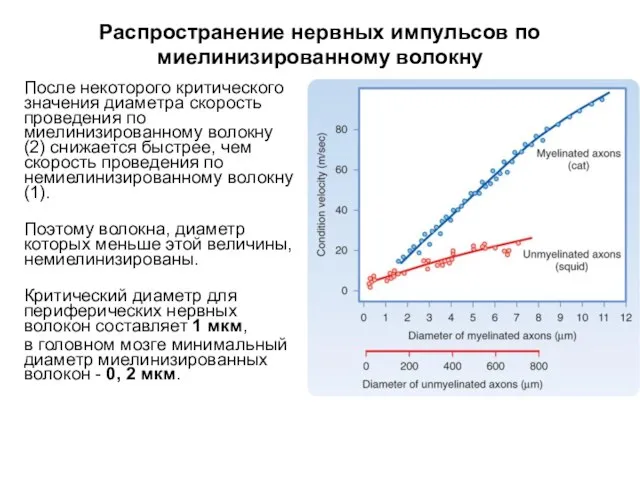
Распространение нервных импульсов по миелинизированному волокну
После некоторого критического значения диаметра скорость
Распространение нервных импульсов по миелинизированному волокну
После некоторого критического значения диаметра скорость
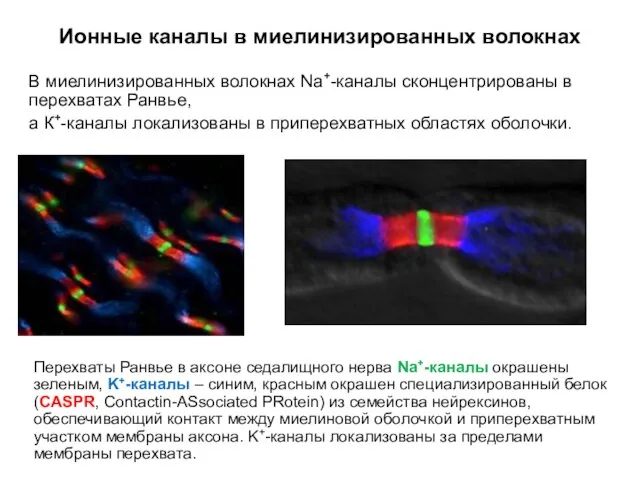
Ионные каналы в миелинизированных волокнах
В миелинизированных волокнах Na+-каналы сконцентрированы в
Ионные каналы в миелинизированных волокнах
В миелинизированных волокнах Na+-каналы сконцентрированы в

Ионные каналы в миелинизированных волокнах
В миелинизированных волокнах Na+-каналы сконцентрированы в
Ионные каналы в миелинизированных волокнах
В миелинизированных волокнах Na+-каналы сконцентрированы в
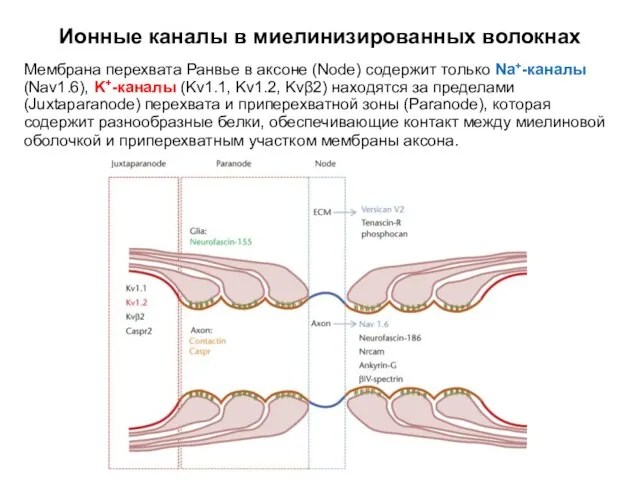
Ионные каналы в миелинизированных волокнах
Мембрана перехвата Ранвье в аксоне (Node)
Ионные каналы в миелинизированных волокнах
Мембрана перехвата Ранвье в аксоне (Node)
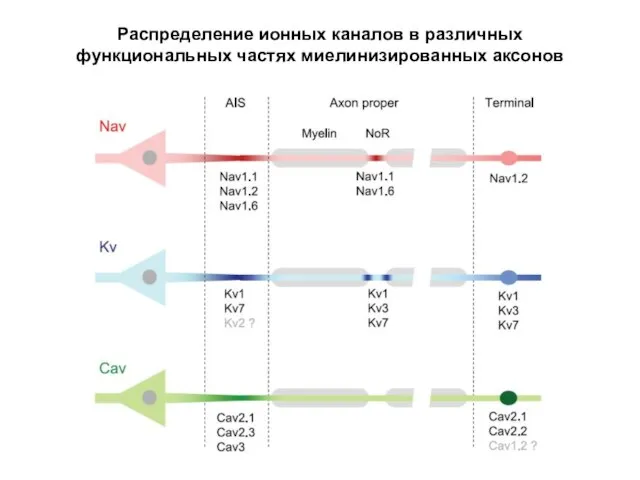
Распределение ионных каналов в различных функциональных частях миелинизированных аксонов
Распределение ионных каналов в различных функциональных частях миелинизированных аксонов

Ионные каналы в миелинизированных волокнах
При возбуждении в перехватах Ранвье возникает
Ионные каналы в миелинизированных волокнах
При возбуждении в перехватах Ранвье возникает
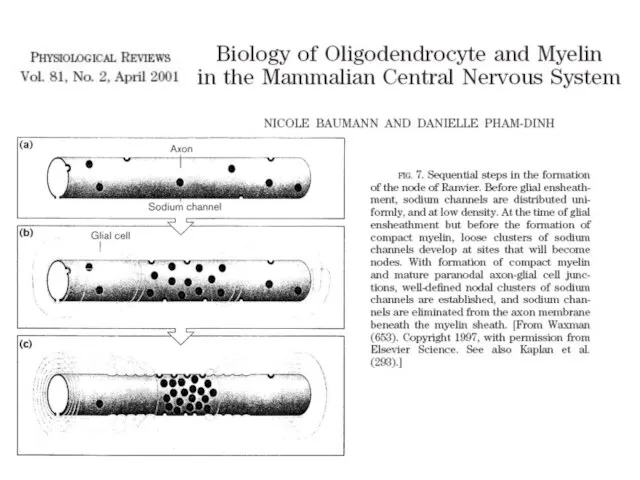
.
.
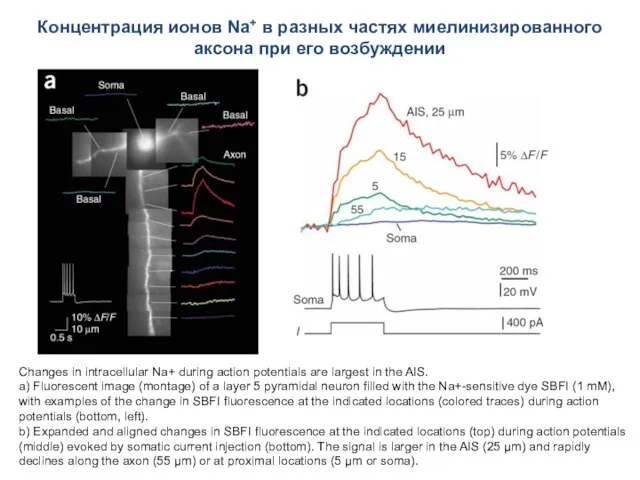
Концентрация ионов Na+ в разных частях миелинизированного аксона при его возбуждении
Концентрация ионов Na+ в разных частях миелинизированного аксона при его возбуждении
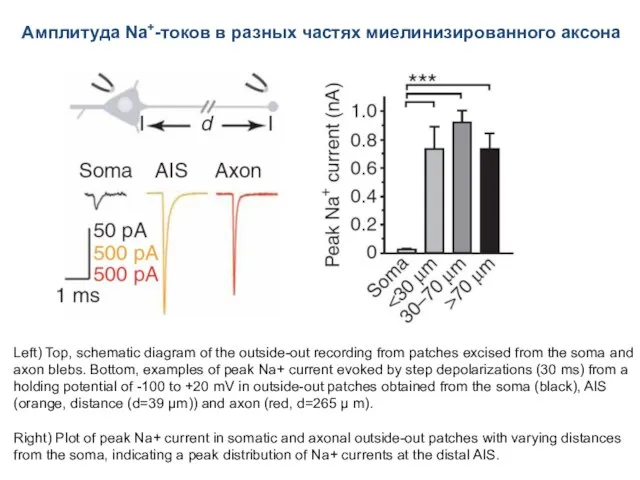
Амплитуда Na+-токов в разных частях миелинизированного аксона
Left) Top, schematic diagram
Амплитуда Na+-токов в разных частях миелинизированного аксона
Left) Top, schematic diagram
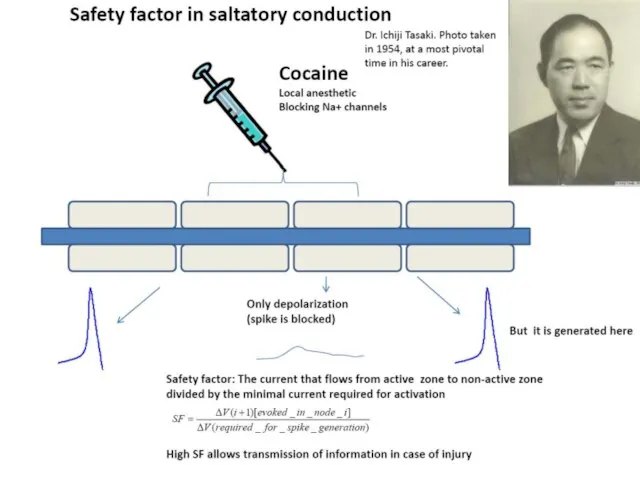
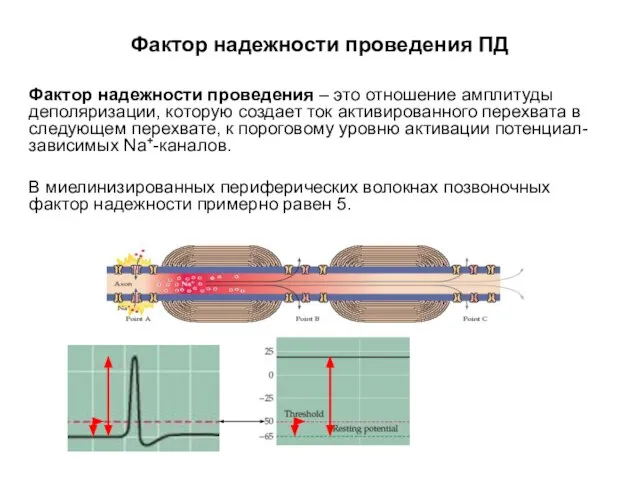
Фактор надежности проведения ПД
Фактор надежности проведения – это отношение амплитуды деполяризации,
Фактор надежности проведения ПД
Фактор надежности проведения – это отношение амплитуды деполяризации,
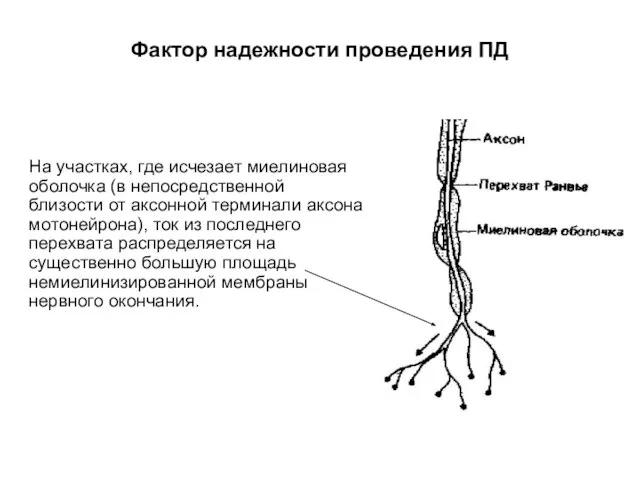
Фактор надежности проведения ПД
На участках, где исчезает миелиновая оболочка (в непосредственной
Фактор надежности проведения ПД
На участках, где исчезает миелиновая оболочка (в непосредственной
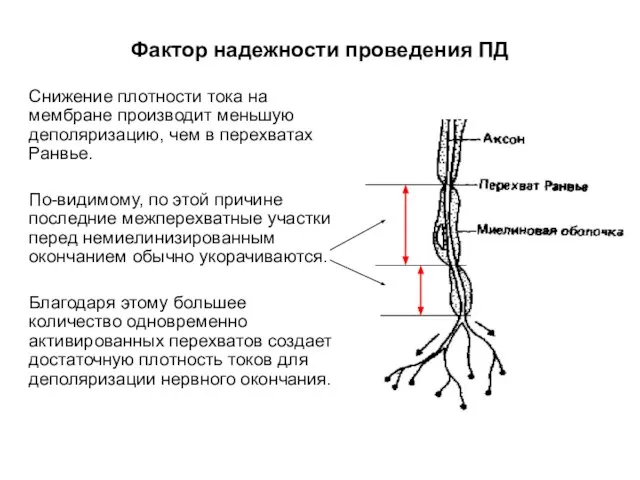
Фактор надежности проведения ПД
Снижение плотности тока на мембране производит меньшую деполяризацию,
Фактор надежности проведения ПД
Снижение плотности тока на мембране производит меньшую деполяризацию,
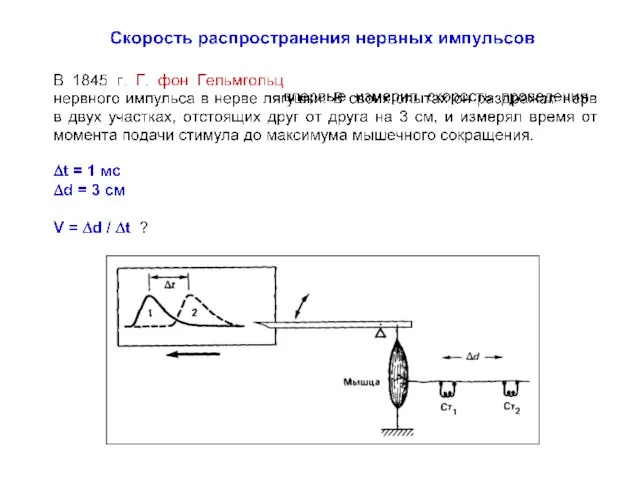
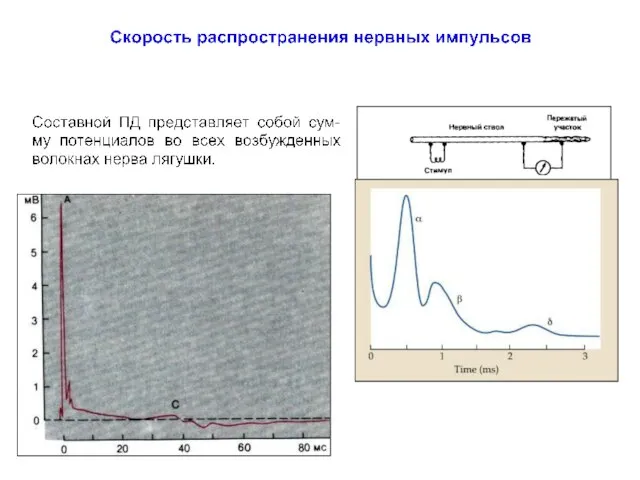

 Полевая мышь
Полевая мышь Аквариумные рыбки
Аквариумные рыбки Метаболические основы функций
Метаболические основы функций Головня. Грибы - паразиты
Головня. Грибы - паразиты Психофизиология детского сна
Психофизиология детского сна Флора. Методы сбора и анализа флористических данных
Флора. Методы сбора и анализа флористических данных Мы выбираем жизнь! Презентация учащихся 9 класса Лихобабина Валерия и Андреева Андрея , выполненная в рамках акции «Гимн жизни», пр
Мы выбираем жизнь! Презентация учащихся 9 класса Лихобабина Валерия и Андреева Андрея , выполненная в рамках акции «Гимн жизни», пр Клетка – элементарная единица живого
Клетка – элементарная единица живого Влияние качества воды реки Еруслан на здоровье школьников Выполнил: ученик 7 класса Довыденко Алексей МОУ - СОШ с.Карпенк
Влияние качества воды реки Еруслан на здоровье школьников Выполнил: ученик 7 класса Довыденко Алексей МОУ - СОШ с.Карпенк Дигибридное и полигибридное скрещивание. Третий закон Менделя – закон независимого комбинирования Урок по основам генетики 10 кл
Дигибридное и полигибридное скрещивание. Третий закон Менделя – закон независимого комбинирования Урок по основам генетики 10 кл Умная домашняя теплица “EcoGrow”
Умная домашняя теплица “EcoGrow” Презентация на тему "Тема: Розмноження та розвиток людини.Вагітність." - скачать бесплатно презентации по Биологии
Презентация на тему "Тема: Розмноження та розвиток людини.Вагітність." - скачать бесплатно презентации по Биологии Jolies plumes, belles ailes
Jolies plumes, belles ailes Фрукты. Игры
Фрукты. Игры ТЕМА УРОКА: «Классы рыб: Хрящевые, Костные». ЦЕЛЬ УРОКА: Познакомиться с многообразием рыб; Дать общую характеристику класс
ТЕМА УРОКА: «Классы рыб: Хрящевые, Костные». ЦЕЛЬ УРОКА: Познакомиться с многообразием рыб; Дать общую характеристику класс Презентация на тему Соцветия
Презентация на тему Соцветия Трахея, trachea
Трахея, trachea Презентация на тему "Строение и значение цветка" - скачать презентации по Биологии
Презентация на тему "Строение и значение цветка" - скачать презентации по Биологии Места обитания и внешнее строение земноводных
Места обитания и внешнее строение земноводных Ветер Ветры всегда влияли на человеческую цивилизацию. Благодаря парусным суднам, которые плыли за счет ветра, впервые появила
Ветер Ветры всегда влияли на человеческую цивилизацию. Благодаря парусным суднам, которые плыли за счет ветра, впервые появила Механизмы транспорта ксенобиотиков
Механизмы транспорта ксенобиотиков Брачное поведение птиц
Брачное поведение птиц Водные обитатели
Водные обитатели Все знают, кто такой гепард, и что он способен развивать огромную скорость. А точнее до 115 км/час, он как лучшая спортивная маши
Все знают, кто такой гепард, и что он способен развивать огромную скорость. А точнее до 115 км/час, он как лучшая спортивная маши Психогенетика
Психогенетика Репродукция вирусов
Репродукция вирусов Власне сполучні тканини
Власне сполучні тканини Видоизменения побегов (6 класс) - презентации по Биологии
Видоизменения побегов (6 класс) - презентации по Биологии