- Реплікація, транскрипція для РНК-вмісних вірусів та вірусів з амбісенсовим геномом

Содержание
- 2. Родини длРНК вірусів Reoviridae – велика родина, мають 10-12 сегменів, інфікує хребетних, безхребетних, рослини і гриби
- 3. Family Reoviridae
- 4. Геном Reovirus 10-12 сегментів dsRNA Пакується 1 копія в віріон Загальний розмір геному 22-28 kb (0.8-4.5
- 5. Консервативні кінцеві послідовності сегментів геномів роду Оrbivirus (+ ланцюг) BTV5 5-GUUAAA............................ACUUAC-3 EHDV 5 -GUUAAA..........................A/GCUUAC-3 AHSV 5
- 6. Модифіковано за Flint et al., Principles of Virology 2nd Ed., ASM Press Структура і організація геному
- 7. Structural and nonstructural proteins encoded by Mammalian reovirus 1
- 8. Модифіковано за Alan Cann by BIH dsRNA 1 dsRNA 2 dsRNA 3 dsRNA 4 dsRNA 5
- 9. mRNAs, ймовірно, переписуються в комплексах транскрипції в кожному з 12 незалежних фрагментів двадцятигранника Модифіковано за Alan
- 10. Кепування та метилювання mRNAs при транскрипції відбувається в корі реовірусної часточки Модифіковано за Alan Cann by
- 11. Транскрипція/Реплікація: RNA транскрибується консервативно: Використовується тільки (‑)смисловий ланцюг; В результаті синтезується (+)смислова mRNAs, Кепувапння відбувається в
- 12. RNA реплікація Геном реплікується в цитоплазмі за консервативним механізмом Виробляється надлишок (+) сенсових ланцюгів, які виступають
- 13. Reovirus: dsRNA Virus Strategy Субвірусні часточки в цитоплазмамі є місцями синтезу мRNA мРНК витісняється в цитоплазму
- 14. Totiviridae віруси - “killer” фунгі Members of the family Totiviridae Не викликає інфекції в заражених клітинах
- 15. Figure 2 Genome organization of Saccharomyces cerevisiae virus L-A (ScV-L-A). The virion-associated RNA polymerase catalyzes in
- 16. Рослинні Reoviruses Три головних роди відрізняються 5’ і 3’ кінцевими ділянками и і в кодованих протеїнах.
- 17. Ambisense genomes
- 18. Організація геному ВПЗТ
- 20. Подібності реплікативного процесу у (+) РНК вірусів, длРНК вірусів і зворотньо-транскрибуючих вірусів
- 21. 7 класів вірусів за стратегією реплікації геному та енкапсидації
- 23. Фундаментальні зв'язки між класами Виявлено паралелі у процесі реплікації геному між: 1) (+) РНК вірусів, 2)
- 24. Паралелі між (+) РНК вірусами і ретровірусами: роль тРНК-послідовностей в ініціації синтезу (-) ланцюга Ініціація реплікації
- 25. Паралелі між (+) РНК вірусами і ретровірусами: комплекси реплікації РНК та капсиди Реплікація (+) РНК вірусів
- 26. Паралелі між (+) РНК вірусами і ретровірусами: формування сферул та капсидів (brome mosaic virus) 1а –
- 27. Регуляція Pol у ретровірусів та (+) РНК вірусів Gag/Gag-Pol = 20 Зменшення співвідношення інгібує збірку віріону
- 28. Паралелі між длРНК вірусами і (+) РНК вірусами: фактори реплікації
- 29. Паралелі між (+) РНК вірусами і длРНК вірусами:
- 31. НЕКАНОНІЧНІ ВІРУСИ
- 32. Сателіти Геном приблизно 500-2000 нуклеотидів з одноланцюгової РНК Геном сателіту не схожий за своїми нуклеотидними послідовностями
- 33. Вірус гепатиту дельта (HDV) Вірус гепатиту дельта – унікальна молекула РНК,, яка схожа на віроїд, однак
- 34. Геном вірусу гепатиту дельта (HDV) Віроїдоподібний регіон Білок-кодуючий регіон (δ - антиген)
- 35. Віріони HBV та HDV
- 36. Віроїди
- 37. Віроїди Дуже малі, ковалентно замкнені, кільцеві РНК молекули, здатні до автономної реплікації (не потребують віруса-хелпера) та
- 38. Відкриття Перший виявлений віроїд Potato spindle tuber viroid (PSTVd) 1967 Dr. Ted Diener
- 39. Potato spindle tuber viroid (PSTVd)
- 40. Avsunviroidae та Pospiviroidae
- 41. Структура віроїдів
- 42. Структура віроїдів
- 43. Реплікація віроїда В ядрі (PSTVd) або хлоропласті (ASBVd) В хлоропласті розрізання рибозимопосередковане, у ядрі - ферментом
- 44. Локалізація +ланцюг віроїдів локалізується і в ядерці, і в нуклеоплазмі -ланцюг – тільки у нуклеоплазмі
- 45. Переміщення віроїда для проникнення в крізь ядерну пору зв’язується з білком VirP1 Через плазмодесми Через флоему
- 46. Основні питання Які молекулярні сигнали примушують РНК-полімеразу хазяїна сприймати віроїд як матрицю для побудови комплементарного ланцюга?
- 47. Організація геному 5 доменів: термінальні петлі Tr і Тl, патогенності (Р), центральний з консервативною ділянкою (С),
- 48. Реплікація віроїду реплікація за механізмом кільця, що котиться сайт ініціації – U359 або C1 у Тl,
- 49. -РНК локалізуються у нуклеоплазмі, де, очевидно, відбувається транскрипція, у той час як +РНК розподілені між нуклеоплазмою
- 50. відсутність movement-протеїнів мутації у правому Т-домені перешкоджають нормальному міжклітинному транспорту мутації у петлі Е блокують рух
- 51. Взаємодія з компонентами клітини хазяїна щонайменше 3 детермінанти патогенезу у доменах Т, Р і V петля
- 52. Взаємодія з компонентами клітини хазяїна інтерференція РНК господаря (суперечливі результати досліджень) взаємодія з протеїнами: - неспецифічна
- 54. Скачать презентацию

Родини длРНК вірусів
Reoviridae – велика родина, мають 10-12 сегменів, інфікує хребетних,
Родини длРНК вірусів
Reoviridae – велика родина, мають 10-12 сегменів, інфікує хребетних,
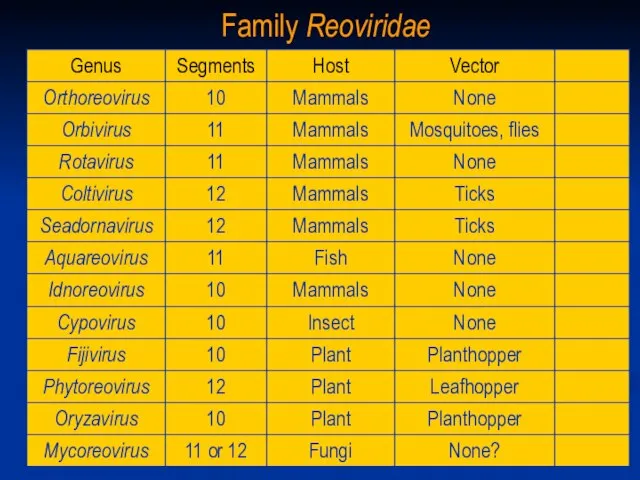
Family Reoviridae
Family Reoviridae

Геном Reovirus
10-12 сегментів dsRNA
Пакується 1 копія в віріон
Загальний розмір геному
Геном Reovirus
10-12 сегментів dsRNA
Пакується 1 копія в віріон
Загальний розмір геному

Консервативні кінцеві послідовності сегментів геномів роду Оrbivirus (+ ланцюг)
BTV5 5-GUUAAA............................ACUUAC-3
EHDV
Консервативні кінцеві послідовності сегментів геномів роду Оrbivirus (+ ланцюг)
BTV5 5-GUUAAA............................ACUUAC-3
EHDV
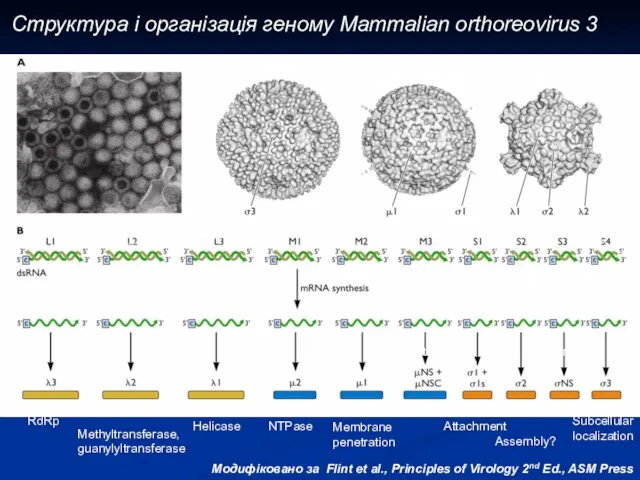
Модифіковано за Flint et al., Principles of Virology 2nd Ed., ASM
Модифіковано за Flint et al., Principles of Virology 2nd Ed., ASM
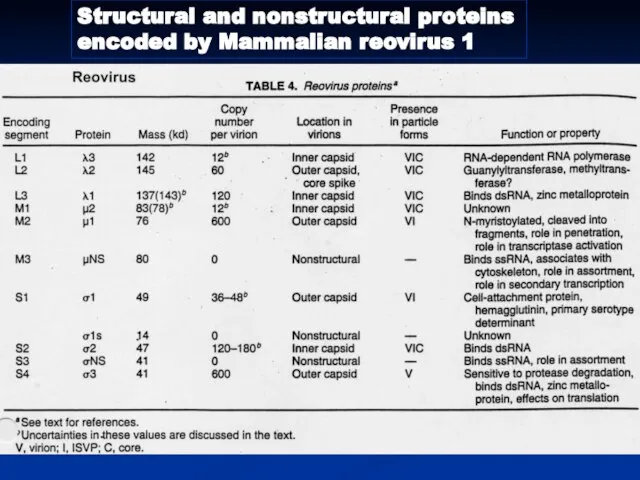
Structural and nonstructural proteins
encoded by Mammalian reovirus 1
Structural and nonstructural proteins
encoded by Mammalian reovirus 1
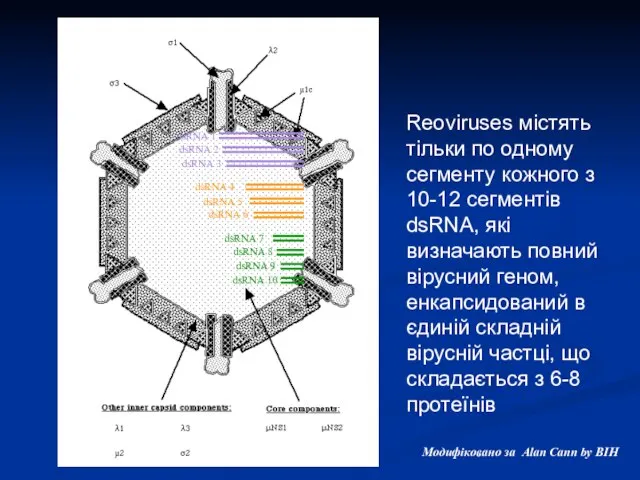
Модифіковано за Alan Cann by BIH
dsRNA 1
dsRNA 2
dsRNA 3
dsRNA 4
dsRNA 5
dsRNA
Модифіковано за Alan Cann by BIH
dsRNA 1
dsRNA 2
dsRNA 3
dsRNA 4
dsRNA 5
dsRNA
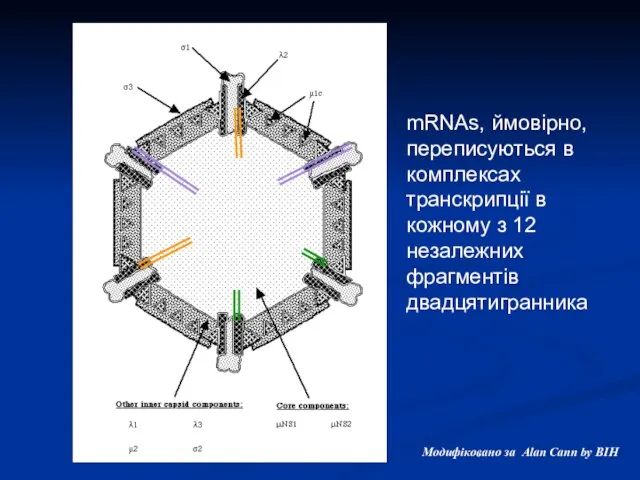
mRNAs, ймовірно, переписуються в комплексах транскрипції в кожному з 12 незалежних
mRNAs, ймовірно, переписуються в комплексах транскрипції в кожному з 12 незалежних
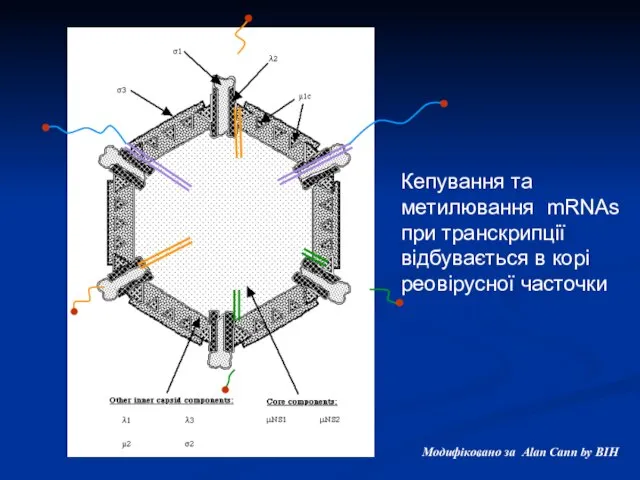
Кепування та метилювання mRNAs при транскрипції відбувається в корі реовірусної часточки
Модифіковано
Кепування та метилювання mRNAs при транскрипції відбувається в корі реовірусної часточки
Модифіковано

Транскрипція/Реплікація:
RNA транскрибується консервативно:
Використовується тільки (‑)смисловий ланцюг;
В результаті синтезується (+)смислова
Транскрипція/Реплікація:
RNA транскрибується консервативно:
Використовується тільки (‑)смисловий ланцюг;
В результаті синтезується (+)смислова
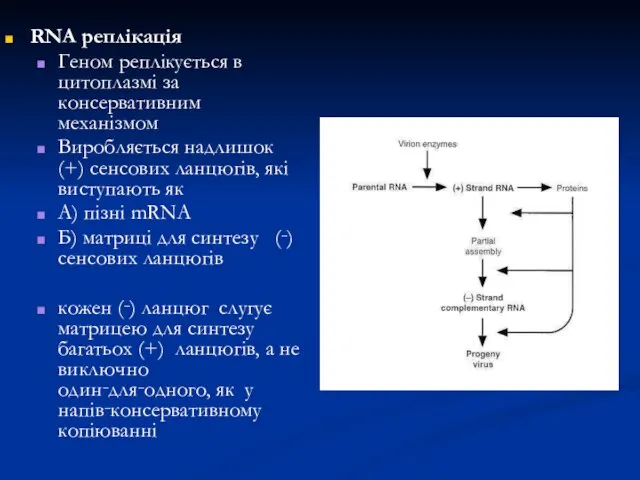
RNA реплікація
Геном реплікується в цитоплазмі за консервативним механізмом
Виробляється надлишок (+)
RNA реплікація
Геном реплікується в цитоплазмі за консервативним механізмом
Виробляється надлишок (+)
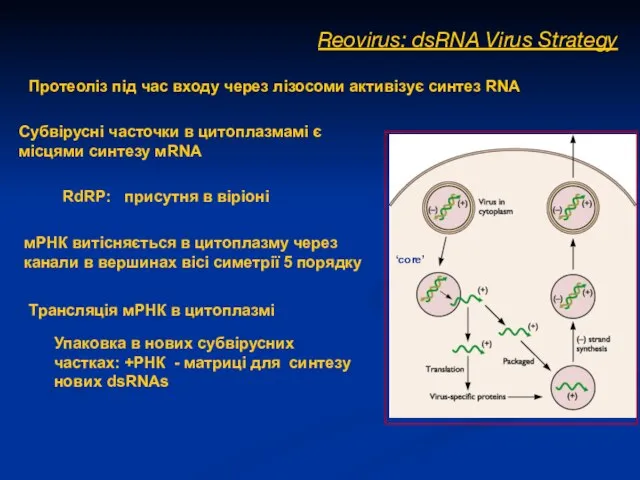
Reovirus: dsRNA Virus Strategy
Субвірусні часточки в цитоплазмамі є місцями синтезу мRNA
Reovirus: dsRNA Virus Strategy
Субвірусні часточки в цитоплазмамі є місцями синтезу мRNA

Totiviridae
віруси - “killer” фунгі
Members of the family Totiviridae
Не викликає інфекції
Totiviridae
віруси - “killer” фунгі
Members of the family Totiviridae
Не викликає інфекції
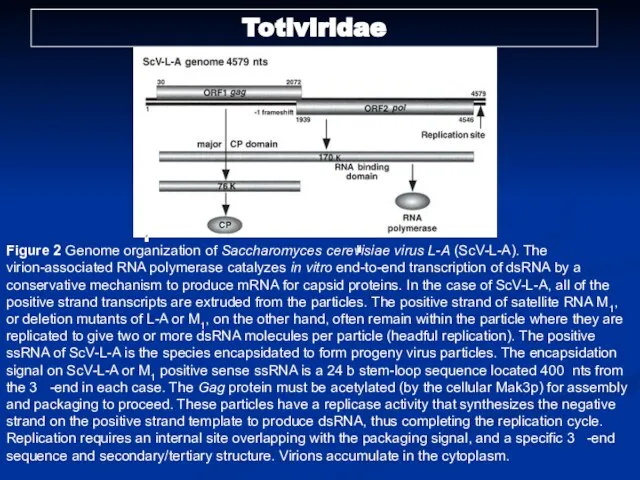
Figure 2 Genome organization of Saccharomyces cerevisiae virus L-A (ScV-L-A). The
Figure 2 Genome organization of Saccharomyces cerevisiae virus L-A (ScV-L-A). The
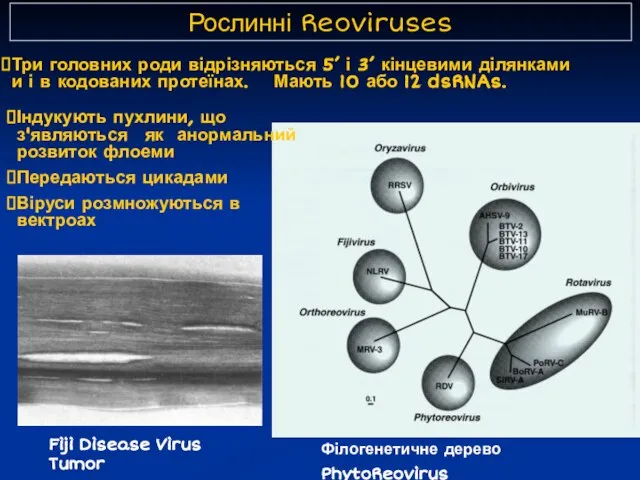
Рослинні Reoviruses
Три головних роди відрізняються 5’ і 3’ кінцевими ділянками
Рослинні Reoviruses
Три головних роди відрізняються 5’ і 3’ кінцевими ділянками
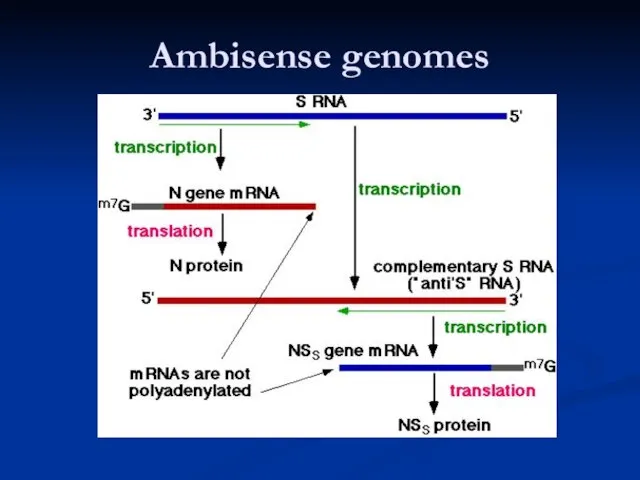
Ambisense genomes
Ambisense genomes
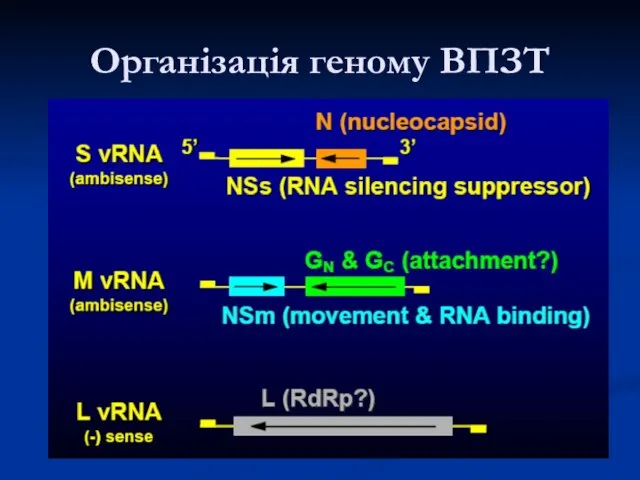
Організація геному ВПЗТ
Організація геному ВПЗТ
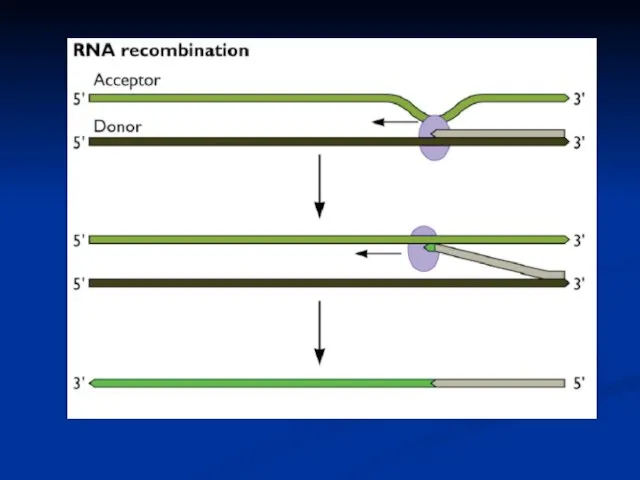

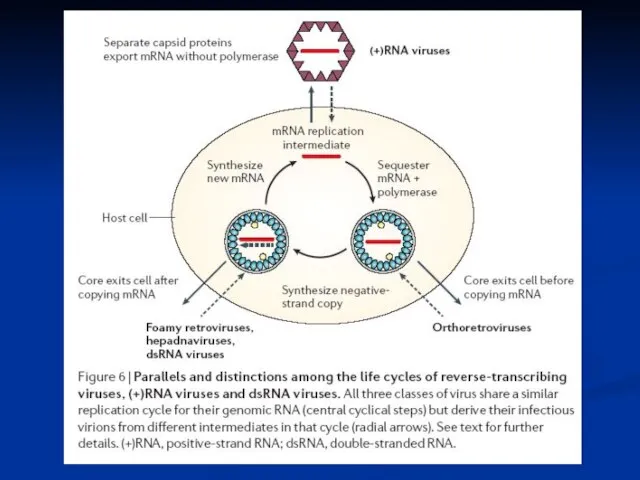
Подібності реплікативного процесу
у (+) РНК вірусів, длРНК вірусів і зворотньо-транскрибуючих вірусів
Подібності реплікативного процесу
у (+) РНК вірусів, длРНК вірусів і зворотньо-транскрибуючих вірусів
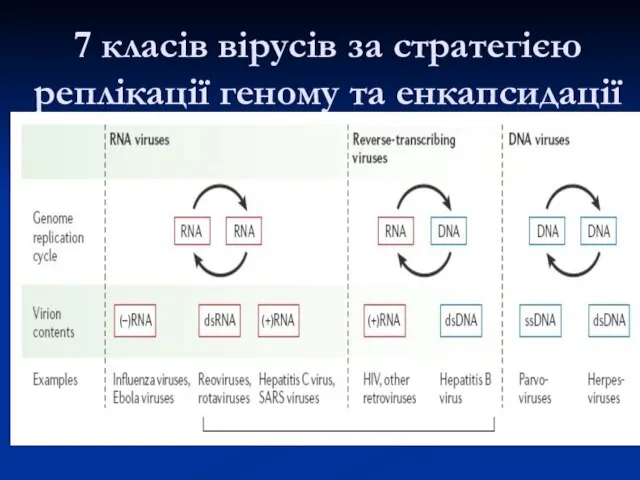
7 класів вірусів за стратегією реплікації геному та енкапсидації
7 класів вірусів за стратегією реплікації геному та енкапсидації
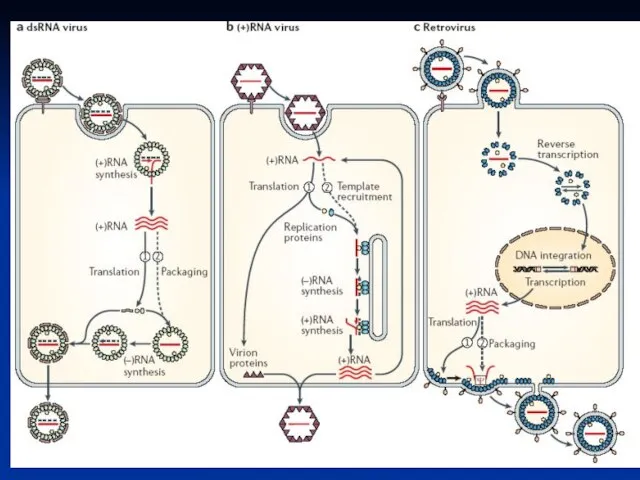

Фундаментальні зв'язки між класами
Виявлено паралелі у процесі реплікації геному між:
1) (+)
Фундаментальні зв'язки між класами
Виявлено паралелі у процесі реплікації геному між:
1) (+)
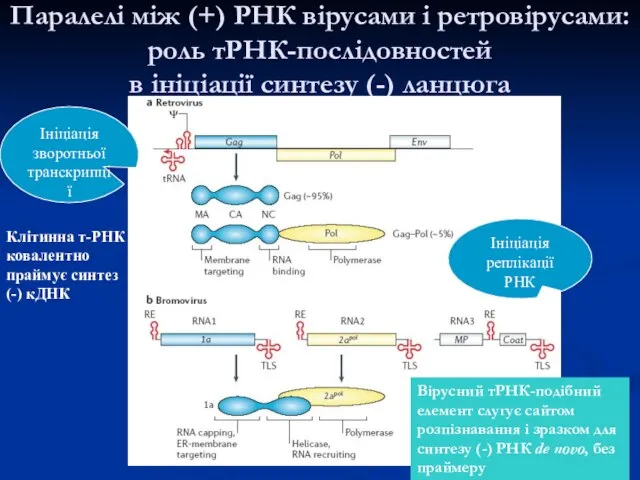
Паралелі між (+) РНК вірусами і ретровірусами: роль тРНК-послідовностей
в ініціації
Паралелі між (+) РНК вірусами і ретровірусами: роль тРНК-послідовностей в ініціації
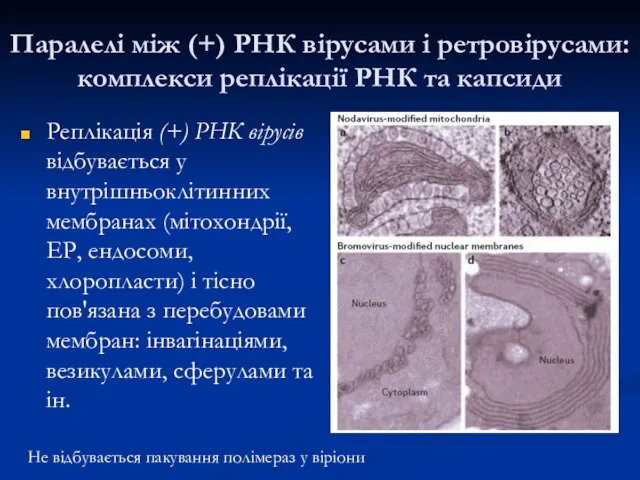
Паралелі між (+) РНК вірусами і ретровірусами: комплекси реплікації РНК та
Паралелі між (+) РНК вірусами і ретровірусами: комплекси реплікації РНК та
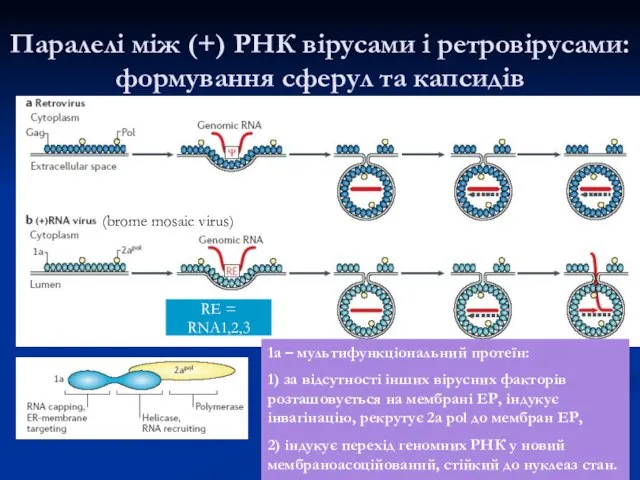
Паралелі між (+) РНК вірусами і ретровірусами: формування сферул та капсидів
(brome
Паралелі між (+) РНК вірусами і ретровірусами: формування сферул та капсидів
(brome
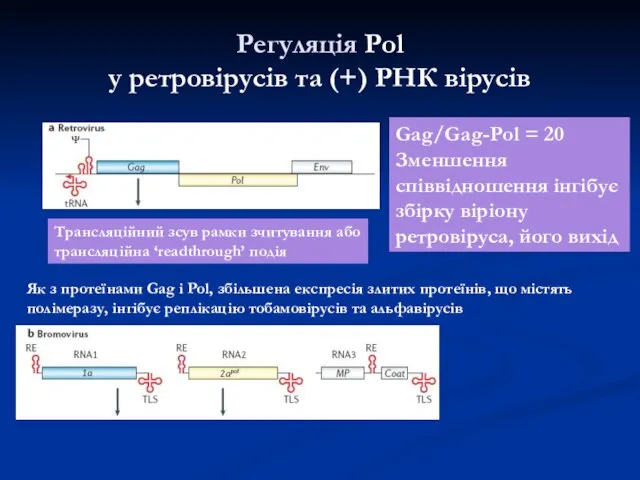
Регуляція Pol
у ретровірусів та (+) РНК вірусів
Gag/Gag-Pol = 20
Зменшення співвідношення
Регуляція Pol
у ретровірусів та (+) РНК вірусів
Gag/Gag-Pol = 20
Зменшення співвідношення
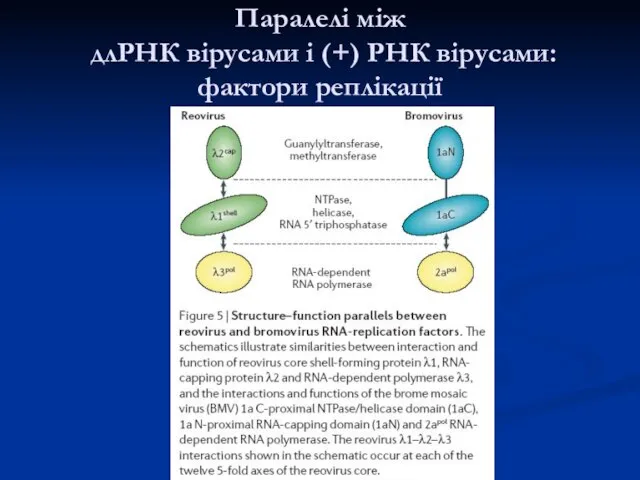
Паралелі між длРНК вірусами і (+) РНК вірусами:
фактори реплікації
Паралелі між длРНК вірусами і (+) РНК вірусами:
фактори реплікації
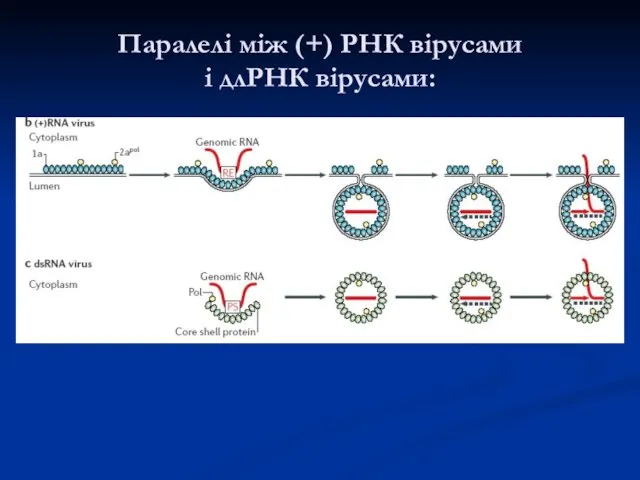
Паралелі між (+) РНК вірусами
і длРНК вірусами:
Паралелі між (+) РНК вірусами
і длРНК вірусами:

НЕКАНОНІЧНІ ВІРУСИ
НЕКАНОНІЧНІ ВІРУСИ

Сателіти
Геном приблизно 500-2000 нуклеотидів з одноланцюгової РНК
Геном сателіту не схожий за
Сателіти
Геном приблизно 500-2000 нуклеотидів з одноланцюгової РНК
Геном сателіту не схожий за

Вірус гепатиту дельта (HDV)
Вірус гепатиту дельта – унікальна молекула РНК,,
Вірус гепатиту дельта (HDV)
Вірус гепатиту дельта – унікальна молекула РНК,,
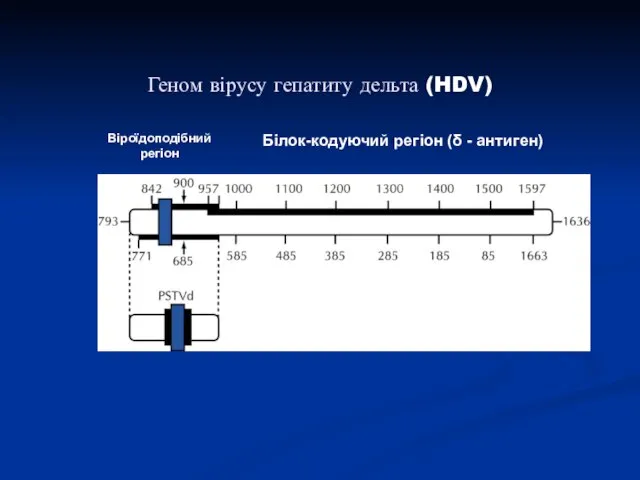
Геном вірусу гепатиту дельта (HDV)
Віроїдоподібний
регіон
Білок-кодуючий регіон (δ - антиген)
Геном вірусу гепатиту дельта (HDV)
Віроїдоподібний
регіон
Білок-кодуючий регіон (δ - антиген)

Віріони HBV та HDV
Віріони HBV та HDV

Віроїди
Віроїди

Віроїди
Дуже малі, ковалентно замкнені, кільцеві РНК молекули, здатні до автономної реплікації
Віроїди
Дуже малі, ковалентно замкнені, кільцеві РНК молекули, здатні до автономної реплікації
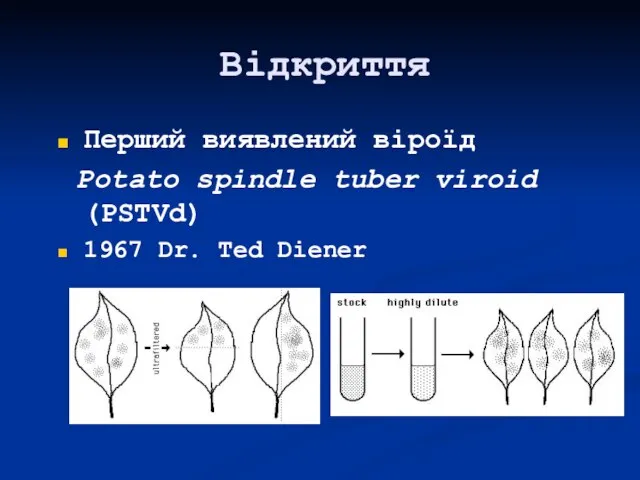
Відкриття
Перший виявлений віроїд
Potato spindle tuber viroid (PSTVd)
1967 Dr. Ted Diener
Відкриття
Перший виявлений віроїд
Potato spindle tuber viroid (PSTVd)
1967 Dr. Ted Diener

Potato spindle tuber viroid (PSTVd)
Potato spindle tuber viroid (PSTVd)
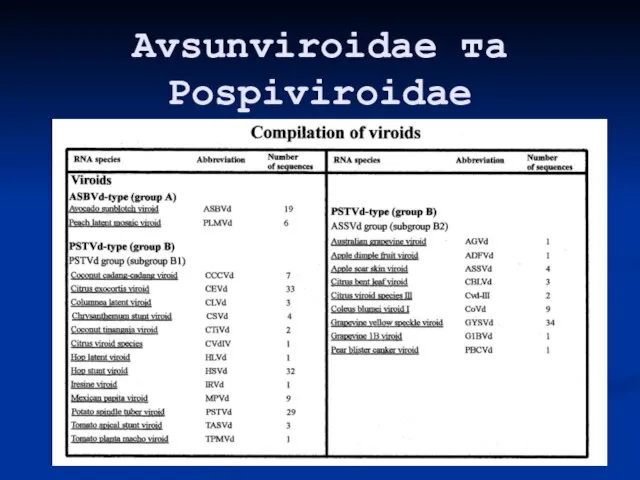
Avsunviroidae та Pospiviroidae
Avsunviroidae та Pospiviroidae
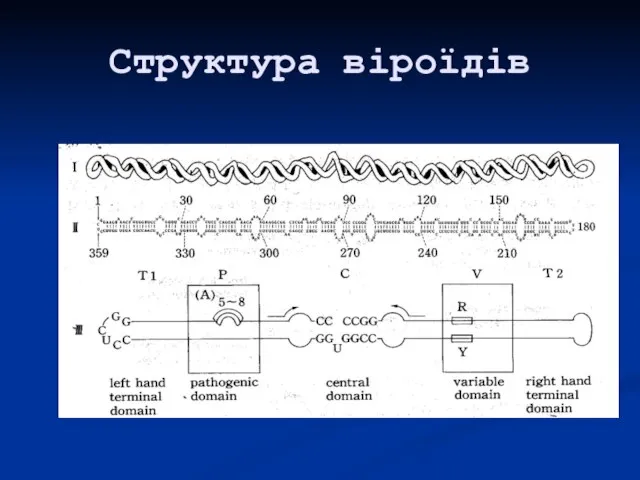
Структура віроїдів
Структура віроїдів
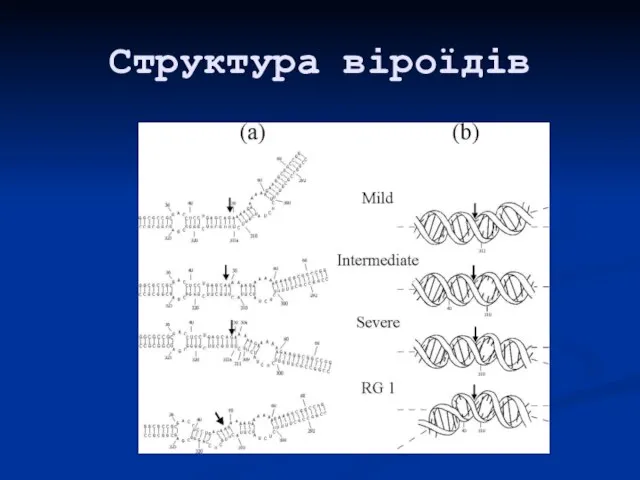
Структура віроїдів
Структура віроїдів
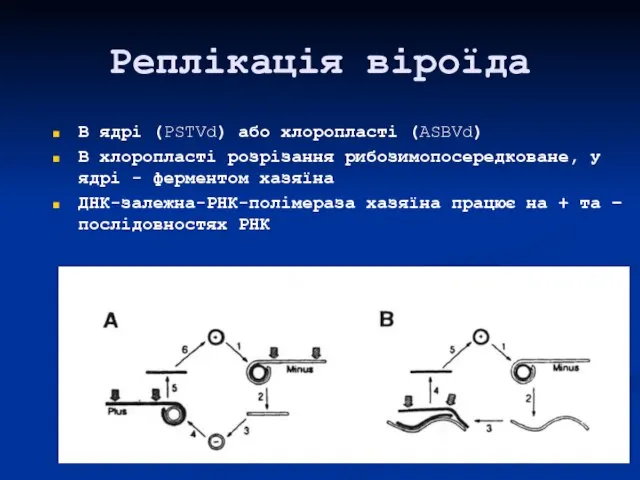
Реплікація віроїда
В ядрі (PSTVd) або хлоропласті (ASBVd)
В хлоропласті розрізання рибозимопосередковане, у
Реплікація віроїда
В ядрі (PSTVd) або хлоропласті (ASBVd)
В хлоропласті розрізання рибозимопосередковане, у

Локалізація
+ланцюг віроїдів локалізується і в ядерці, і в нуклеоплазмі
-ланцюг –
Локалізація
+ланцюг віроїдів локалізується і в ядерці, і в нуклеоплазмі
-ланцюг –

Переміщення віроїда
для проникнення в крізь ядерну пору зв’язується з білком VirP1
Через
Переміщення віроїда
для проникнення в крізь ядерну пору зв’язується з білком VirP1
Через

Основні питання
Які молекулярні сигнали примушують РНК-полімеразу хазяїна сприймати віроїд як матрицю
Основні питання
Які молекулярні сигнали примушують РНК-полімеразу хазяїна сприймати віроїд як матрицю
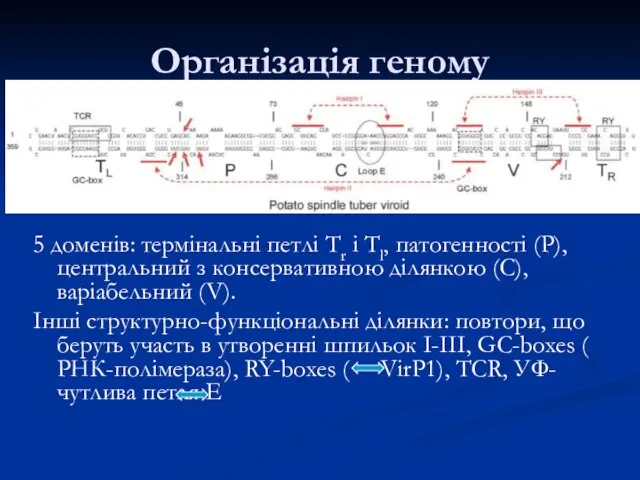
Організація геному
5 доменів: термінальні петлі Tr і Тl, патогенності (Р), центральний
Організація геному
5 доменів: термінальні петлі Tr і Тl, патогенності (Р), центральний
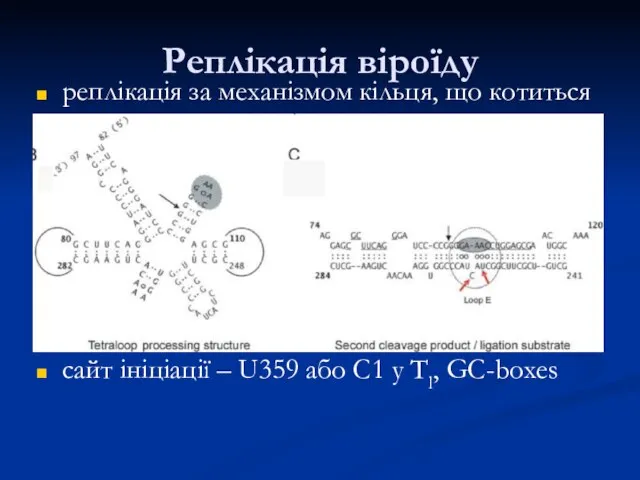
Реплікація віроїду
реплікація за механізмом кільця, що котиться
сайт ініціації – U359 або
Реплікація віроїду
реплікація за механізмом кільця, що котиться
сайт ініціації – U359 або

-РНК локалізуються у нуклеоплазмі, де, очевидно, відбувається транскрипція, у той час
-РНК локалізуються у нуклеоплазмі, де, очевидно, відбувається транскрипція, у той час

відсутність movement-протеїнів
мутації у правому Т-домені перешкоджають нормальному міжклітинному транспорту
мутації у петлі
відсутність movement-протеїнів
мутації у правому Т-домені перешкоджають нормальному міжклітинному транспорту
мутації у петлі

Взаємодія з компонентами клітини хазяїна
щонайменше 3 детермінанти патогенезу у доменах Т,
Взаємодія з компонентами клітини хазяїна
щонайменше 3 детермінанти патогенезу у доменах Т,

Взаємодія з компонентами клітини хазяїна
інтерференція РНК господаря (суперечливі результати досліджень)
взаємодія з
Взаємодія з компонентами клітини хазяїна
інтерференція РНК господаря (суперечливі результати досліджень)
взаємодія з
 Многообразие насекомых Отряды: Чешуекрылые Двукрылые Равнокрылые Блохи
Многообразие насекомых Отряды: Чешуекрылые Двукрылые Равнокрылые Блохи Клонирование. Молекулярное клонирование
Клонирование. Молекулярное клонирование Отряд Вши
Отряд Вши Агрессия собак
Агрессия собак Функциональные блоки мозга
Функциональные блоки мозга Презентация на тему "Половое и бесполое размножение организмов" - скачать презентации по Биологии
Презентация на тему "Половое и бесполое размножение организмов" - скачать презентации по Биологии Отруєння їстівними грибами
Отруєння їстівними грибами  Биология – наука о живой природе
Биология – наука о живой природе Птицы жарких стран
Птицы жарких стран Биохимия крови
Биохимия крови Водный и минеральный обмен
Водный и минеральный обмен Жүректі тыңдау жүрек саздары. Жүректің қақпашаларының нақты орны
Жүректі тыңдау жүрек саздары. Жүректің қақпашаларының нақты орны Презентация на тему "Прорастание" - скачать презентации по Биологии
Презентация на тему "Прорастание" - скачать презентации по Биологии Биотехнология, ее достижения, перспективы развития
Биотехнология, ее достижения, перспективы развития Нервная ткань
Нервная ткань Морж
Морж Тип Кишечнополостные Презентацию составила: Живоглазова Н.А.
Тип Кишечнополостные Презентацию составила: Живоглазова Н.А. Программированный опрос Соотнесите каждое из утверждений с правильным ответом, соединив линиями утверждение и ответ. По
Программированный опрос Соотнесите каждое из утверждений с правильным ответом, соединив линиями утверждение и ответ. По Артерии таза и нижней конечности
Артерии таза и нижней конечности Попугаи
Попугаи Ледовских Ильи 11 а класса http://vkontakte.ru/id36052741
Ледовских Ильи 11 а класса http://vkontakte.ru/id36052741 Презентация на тему Биологические катализаторы
Презентация на тему Биологические катализаторы Сравнение Японской и Техасской белой породы перепелов
Сравнение Японской и Техасской белой породы перепелов Интегрированный урок по биологии и ОБЖ на тему «Беспозвоночные животные и сохранение здоровья человека» Презентацию подготовил
Интегрированный урок по биологии и ОБЖ на тему «Беспозвоночные животные и сохранение здоровья человека» Презентацию подготовил Кольчатые черви. Общие сведения
Кольчатые черви. Общие сведения Анатомия и физиология системы пищеварения. План строения пищеварительной системы
Анатомия и физиология системы пищеварения. План строения пищеварительной системы Домашние животные
Домашние животные Одноклеточные животные или Простейшие
Одноклеточные животные или Простейшие