- Уровни организации наследственного материала у про- и эукариот

Содержание
- 2. Ядро клетки было открыто в 1831 г. английским ботаником Робертом Брауном. Он открыл его в клетках
- 3. Наследственный материал про- и эукариотической клетки Прокариоты: Кольцевая молекула ДНК, образующая нуклеоид Генетический материал находится в
- 4. Ядро клетки Основные функции ядра 1. Хранение и передача наследственной информации Репликация ДНК Репарация ДНК Кроссинговер
- 5. Доказательства роли ядра в передаче наследственной информации Опыты Геммерлинга Объект опыта: одноклеточная водоросль (Acetabularia), имеющая форму
- 6. Опыты с яйцеклетками лягушек Объект: два подвида лягушек. У одного из них (1 подвид) из яйцеклетки
- 7. Опыты Астаурова с тутовым шелкопрядом Астауров Борис Львович Объект: два подвида тутового шелкопряда. У одного подвида
- 8. Основные структурные компоненты эукариотических клеток.
- 9. ядерная оболочка (кариолема), ядерный сок (или кариоплазма), ядрышки хроматин. Структурные компоненты ядра
- 10. Ядерная оболочка Наружная ядерная мембрана Внутренняя ядерная мембрана Перинуклеарное пространство (10 - 30 нм) Наружная мембрана
- 11. Ядерная ламина Внутренняя мембрана связана с ядерной ламиной, которая состоит из трех типов белков A, B,
- 12. Ядерные поры Ферментативная воронка, которая пропускает вещества. Образована 3 рядами глобулярных белков.
- 13. Функция поры: Барьерная Регуляторная Транспортная фиксирующая
- 14. Ядрышко (непостоянный компонент ядра) возникновение ядрышек связано с ядрышковыми организаторами, расположенными в области вторичных перетяжек спутничных
- 15. хроматин метафаз интерфаза Хроматин - это одно из возможных структурно-функциональных состояний наследственного материала Хромосома
- 16. Гетерохроматин – спирализованный, конденсированный, неактиывный, нетранскрибируемый, более интенсивно окрашен. Эухроматин – деспирализованный, активный, транскрибируемый, менее окрашенный.
- 17. Химический состав хроматина (хромосом) 40% - ДНК, 60% - белков: - 40% гистоновых белков (Н1, Н2а,
- 18. Уровни укладки ДНК в хромосому Нуклеосомный Хроматиновые фибриллы (соленоид) 30 nm (нуклеомерный) Хроматиновые филаменты (Хроматиновые петли-домены)
- 19. H1 Нуклеосома - наименьшая единица хроматина и хромосомы Нуклеосомный кор Линкерный участлк H2A, H2B, H3, H4
- 20. Нуклесомный уровень дает формирование «цепочки из бусинок» (соленоидный и нуклеомерный типы укладки). 6-7 нуклеосом сближаются и
- 21. Третий уровень – хромомерный Хроматиновые филаменты (Хроматиновые петли-домены) Фибриллы формируют петли-домены, которые фиксируются негистоновым белком (scaffold).
- 22. Четвертый уровень – хромонемный Сближенные хромомеры образуют толстые нити – хромонемы Пятый уровень – хроматидный (хромосомный)
- 23. Минибенд содержит около 18 петель ДНК укорачивается в 10.000 раз. Минибенды при дальнейшей компактизации дают формирование
- 25. Строение метафазной хромосомы
- 26. Морфологические типы хромосом. Метацентрические Субметацентрические Акроцентрические Спутничные Телоцентрические
- 27. Хромосомы подразделяются на аутосомы (одинаковые у обоих полов). и гетеросомы, или половые хромосомы (разные для мужских
- 28. Совокупность числа и морфологии хромосом данного вида называется - КАРИОТИП
- 29. Классификация хромосом
- 30. 11-я хромосома человека HBB — β-субъединица гемоглобина 16-я хромосома человека 2-я хромосома человека
- 33. После утверждения в 20-х годах ХХ в. хромосомной теории наследственности биологи более сорока лет считали, что
- 34. Нуклеиновые кислоты Это природные высокомолекулярные органические биополимеры, обеспечивающие хранение и передачу наследственной информации. Ф. Левен, Д.
- 35. Строение нуклеотида Углевод Азотистое основание Остаток фосфорной кислоты
- 36. Углевод (сахар, рентоза) рибоза Две группы: дезоксирибоза Только водород Гидроксильная группа
- 37. Азотистое основание Пиримидиновые: тимин цитозин урацил Пуриновые: аденин гуанин
- 38. 3' конец 5' конец
- 40. ДНК двухцепочечный высокомолекулярный биополимер. Является носителем генетической информации. Мономер - дезоксирибонуклеотид РНК Одноцепочечный высокомолекулярный биополимер, мономером
- 42. Описана в 1953 James Watson и Francis Crick Вторичная структура ДНК
- 43. Свойства ДНК Цепи ДНК соединены посредством водородных связей между комплементаными азотистыми основаниями A=T G≡C Комплементарность Антипараллельность
- 44. Антипараллельность ДНК
- 45. Доказательства роли ДНК в передаче наследственной информации (опыты по трансформации и трансдукции). Трансформация - изменение наследственных
- 46. Вывод: под действием трансформирующего фактора живые авирулентные пневмококки приобрели вирулентные свойства штамма S2. В 1944г Эвери
- 48. Трансдукция (от лат. transduction - перемещение) – процесс переноса фрагмента бактериальной ДНК из клетки – донора
- 49. Первый из экспериментов был выполнен в 1952 году американскими генетиками Джошуа Ледербергом и Нортоном Циндлером. Нобелевская
- 50. Для эксперимента была использована U-образная трубка, которая в нижней части посредине была разделена бактериальным фильтром, через
- 51. Свойства ДНК Функции ДНК: хранение, передача, реализация репликация репарация
- 52. Вся масса ДНК Ядро (98-99%) Ядерный геном Цитоплазма (1-2%) Плазмон Линейная ДНК, связанная с белками Митохондриальная
- 54. Скачать презентацию
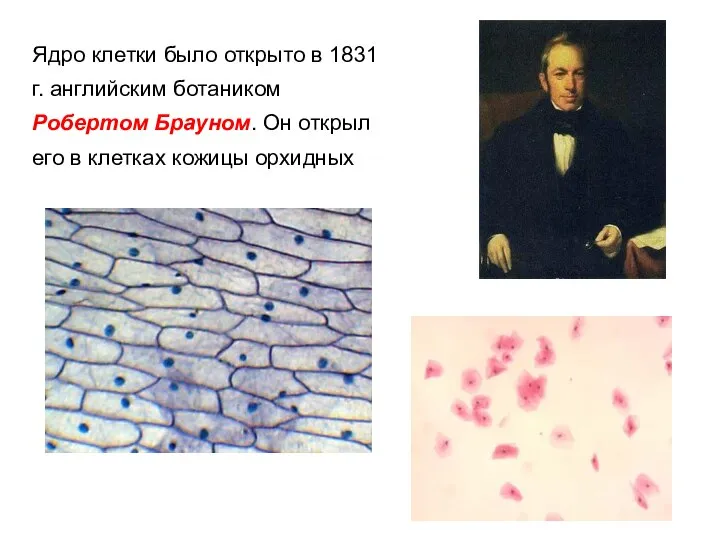
Ядро клетки было открыто в 1831 г. английским ботаником Робертом Брауном.
Ядро клетки было открыто в 1831 г. английским ботаником Робертом Брауном.
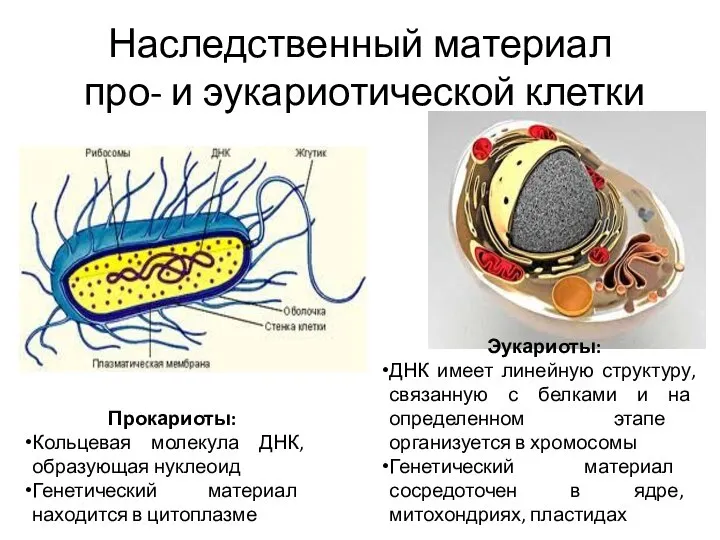
Наследственный материал
про- и эукариотической клетки
Прокариоты:
Кольцевая молекула ДНК, образующая нуклеоид
Генетический материал
Наследственный материал
про- и эукариотической клетки
Прокариоты:
Кольцевая молекула ДНК, образующая нуклеоид
Генетический материал
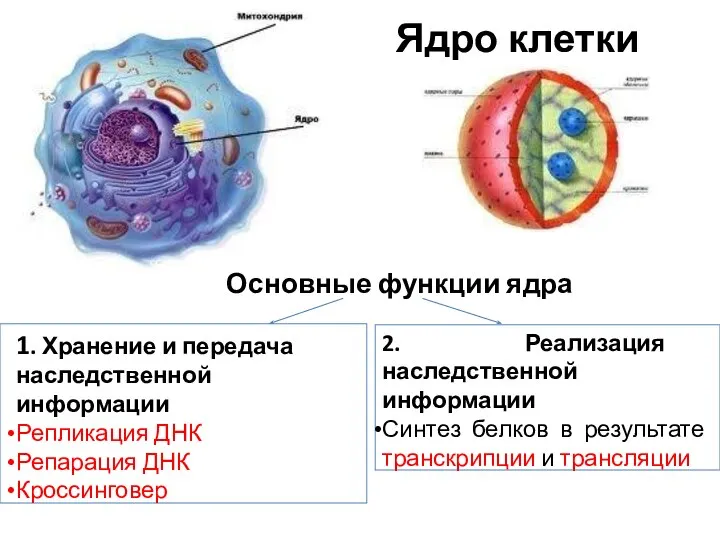
Ядро клетки
Основные функции ядра
1. Хранение и передача наследственной информации
Репликация ДНК
Репарация ДНК
Кроссинговер
2.
Ядро клетки
Основные функции ядра
1. Хранение и передача наследственной информации
Репликация ДНК
Репарация ДНК
Кроссинговер
2.
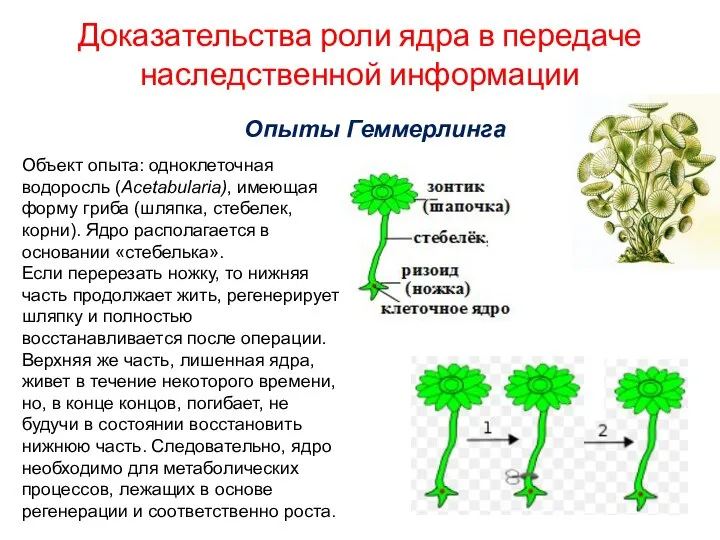
Доказательства роли ядра в передаче наследственной информации
Опыты Геммерлинга
Объект опыта: одноклеточная водоросль
Доказательства роли ядра в передаче наследственной информации
Опыты Геммерлинга
Объект опыта: одноклеточная водоросль
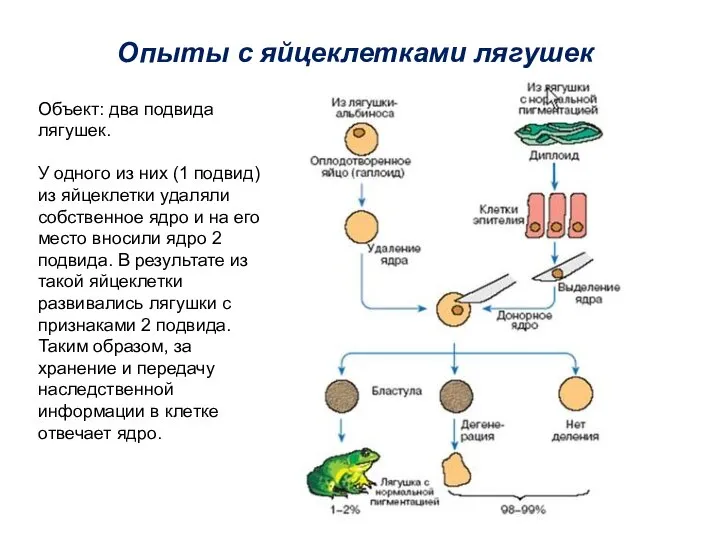
Опыты с яйцеклетками лягушек
Объект: два подвида лягушек.
У одного из них
Опыты с яйцеклетками лягушек
Объект: два подвида лягушек.
У одного из них

Опыты Астаурова с тутовым шелкопрядом
Астауров
Борис Львович
Объект: два подвида тутового шелкопряда.
Опыты Астаурова с тутовым шелкопрядом
Астауров
Борис Львович
Объект: два подвида тутового шелкопряда.
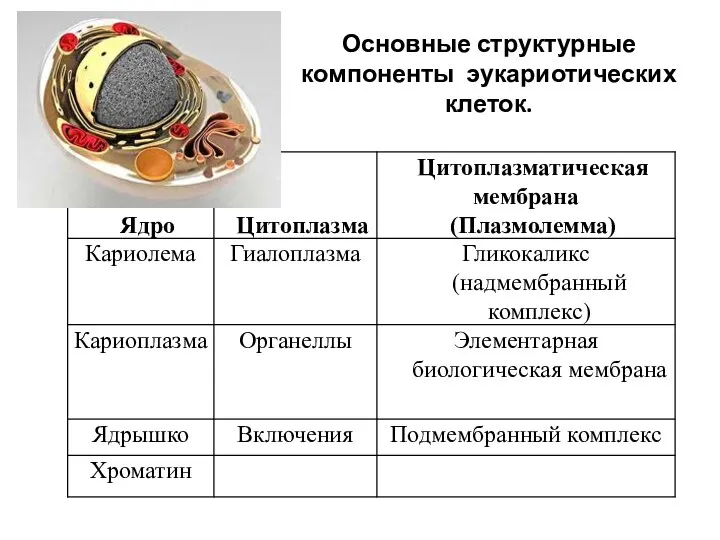
Основные структурные компоненты эукариотических клеток.
Основные структурные компоненты эукариотических клеток.

ядерная оболочка (кариолема),
ядерный сок (или кариоплазма),
ядрышки
хроматин.
Структурные компоненты ядра
ядерная оболочка (кариолема),
ядерный сок (или кариоплазма),
ядрышки
хроматин.
Структурные компоненты ядра
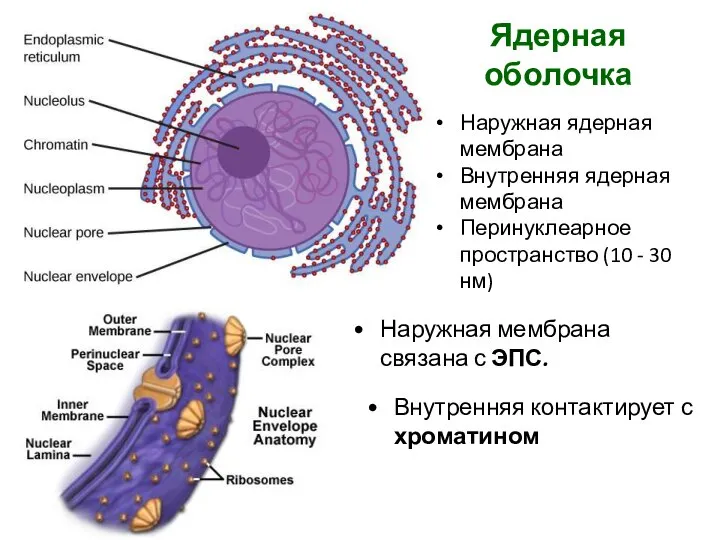
Ядерная оболочка
Наружная ядерная мембрана
Внутренняя ядерная мембрана
Перинуклеарное пространство (10 - 30 нм)
Наружная
Ядерная оболочка
Наружная ядерная мембрана
Внутренняя ядерная мембрана
Перинуклеарное пространство (10 - 30 нм)
Наружная
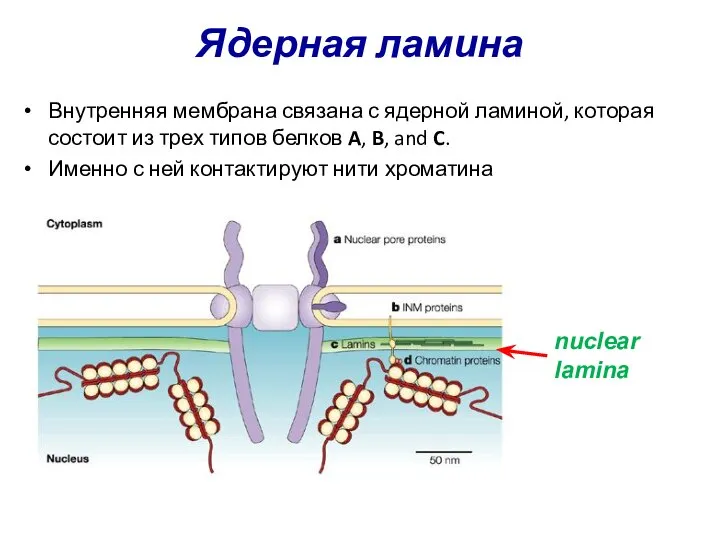
Ядерная ламина
Внутренняя мембрана связана с ядерной ламиной, которая состоит из трех
Ядерная ламина
Внутренняя мембрана связана с ядерной ламиной, которая состоит из трех
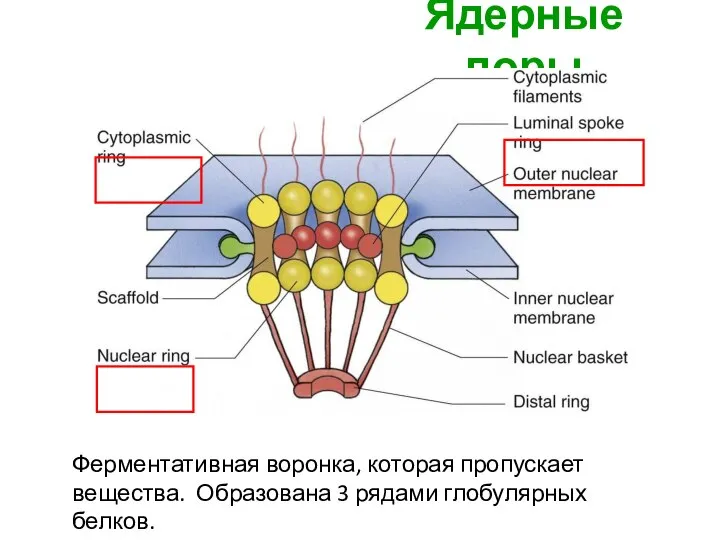
Ядерные поры
Ферментативная воронка, которая пропускает вещества. Образована 3 рядами глобулярных белков.
Ядерные поры
Ферментативная воронка, которая пропускает вещества. Образована 3 рядами глобулярных белков.
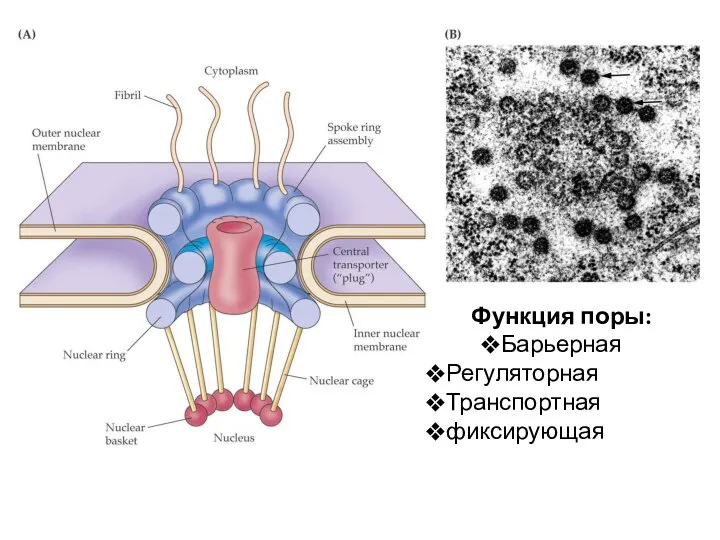
Функция поры:
Барьерная
Регуляторная
Транспортная
фиксирующая
Функция поры:
Барьерная
Регуляторная
Транспортная
фиксирующая
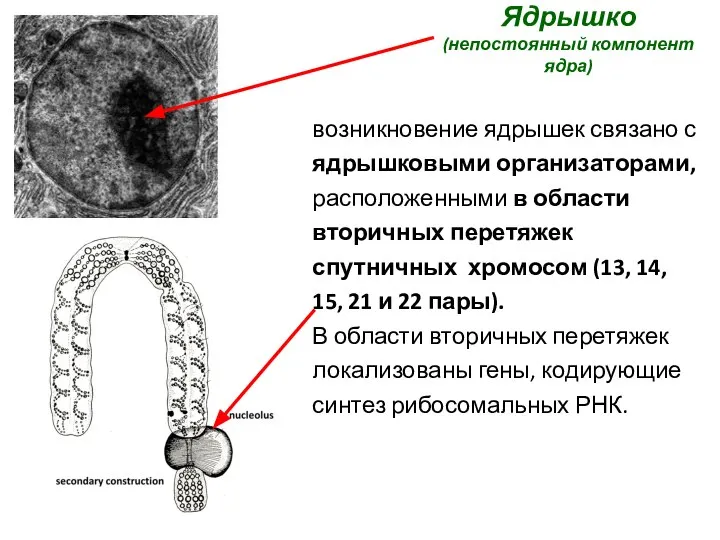
Ядрышко
(непостоянный компонент ядра)
возникновение ядрышек связано с ядрышковыми организаторами, расположенными в
Ядрышко
(непостоянный компонент ядра)
возникновение ядрышек связано с ядрышковыми организаторами, расположенными в
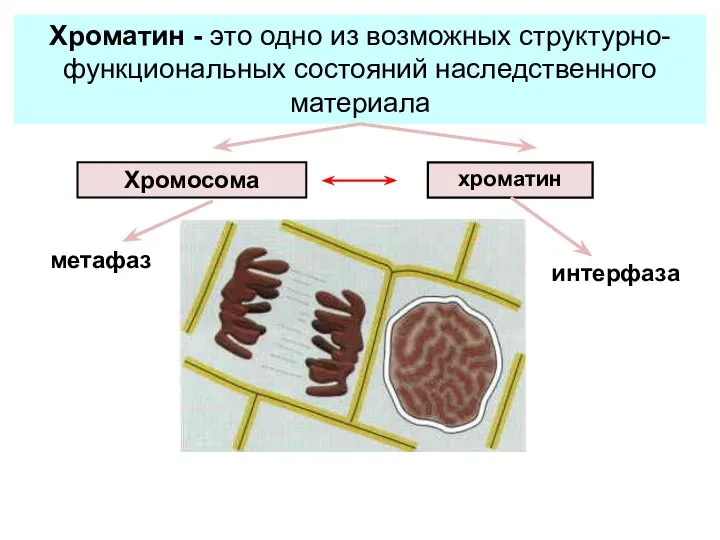
хроматин
метафаз
интерфаза
Хроматин - это одно из возможных структурно-функциональных состояний наследственного материала
Хромосома
хроматин
метафаз
интерфаза
Хроматин - это одно из возможных структурно-функциональных состояний наследственного материала
Хромосома
Гетерохроматин –
спирализованный,
конденсированный,
неактиывный,
нетранскрибируемый, более
интенсивно окрашен.
Эухроматин –
Гетерохроматин –
спирализованный,
конденсированный,
неактиывный,
нетранскрибируемый, более
интенсивно окрашен.
Эухроматин –
Химический состав хроматина (хромосом)
40% - ДНК,
60% - белков:
- 40% гистоновых
Химический состав хроматина (хромосом)
40% - ДНК,
60% - белков:
- 40% гистоновых
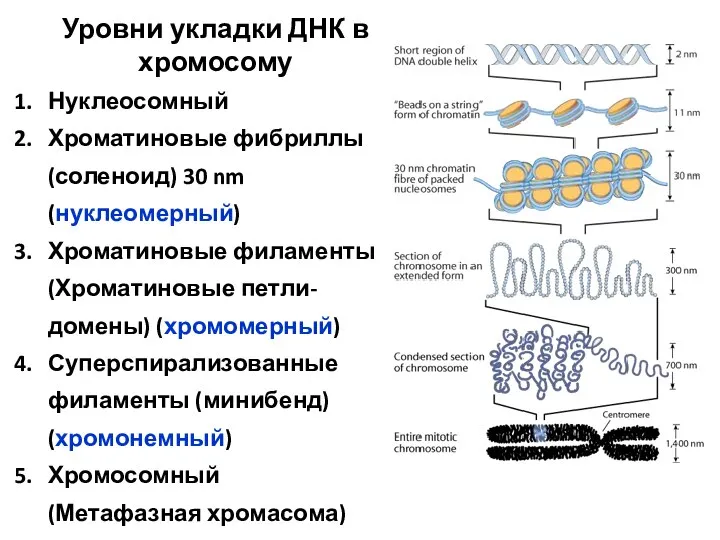
Уровни укладки ДНК в хромосому
Нуклеосомный
Хроматиновые фибриллы (соленоид) 30 nm (нуклеомерный)
Хроматиновые филаменты
Уровни укладки ДНК в хромосому
Нуклеосомный
Хроматиновые фибриллы (соленоид) 30 nm (нуклеомерный)
Хроматиновые филаменты
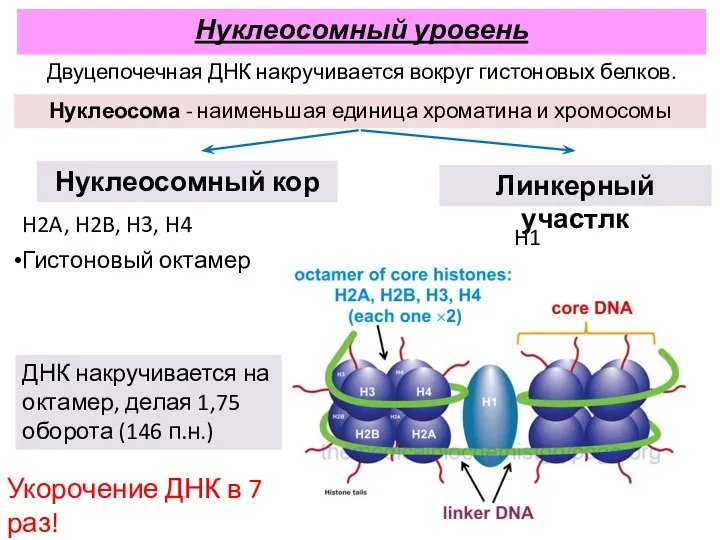
H1
Нуклеосома - наименьшая единица хроматина и хромосомы
Нуклеосомный кор
Линкерный участлк
H2A, H2B, H3,
H1
Нуклеосома - наименьшая единица хроматина и хромосомы
Нуклеосомный кор
Линкерный участлк
H2A, H2B, H3,

Нуклесомный уровень дает формирование «цепочки из бусинок» (соленоидный и нуклеомерный типы
Нуклесомный уровень дает формирование «цепочки из бусинок» (соленоидный и нуклеомерный типы
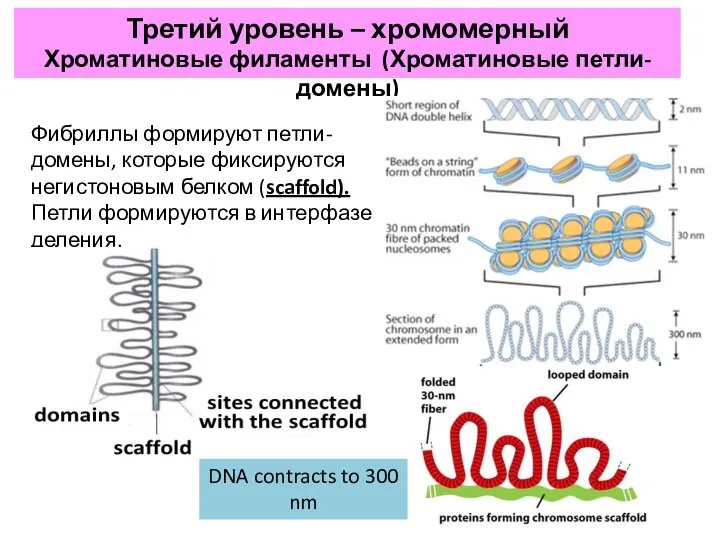
Третий уровень – хромомерный
Хроматиновые филаменты (Хроматиновые петли-домены)
Фибриллы формируют петли-домены, которые
Третий уровень – хромомерный
Хроматиновые филаменты (Хроматиновые петли-домены)
Фибриллы формируют петли-домены, которые
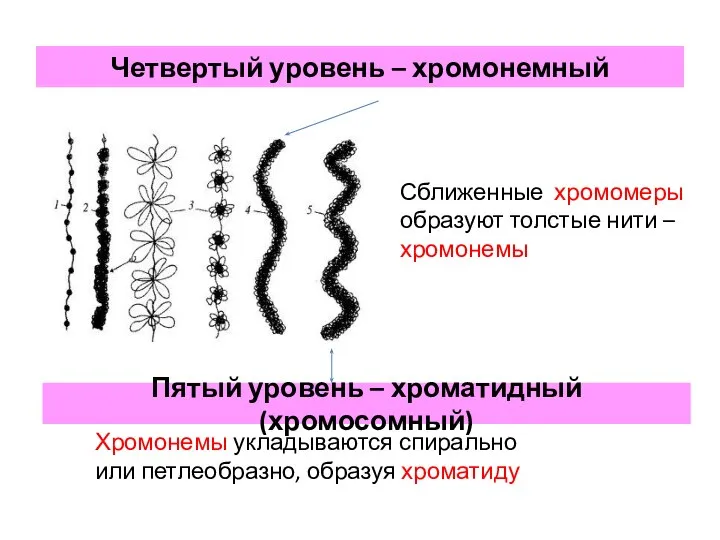
Четвертый уровень – хромонемный
Сближенные хромомеры образуют толстые нити – хромонемы
Пятый
Четвертый уровень – хромонемный
Сближенные хромомеры образуют толстые нити – хромонемы
Пятый
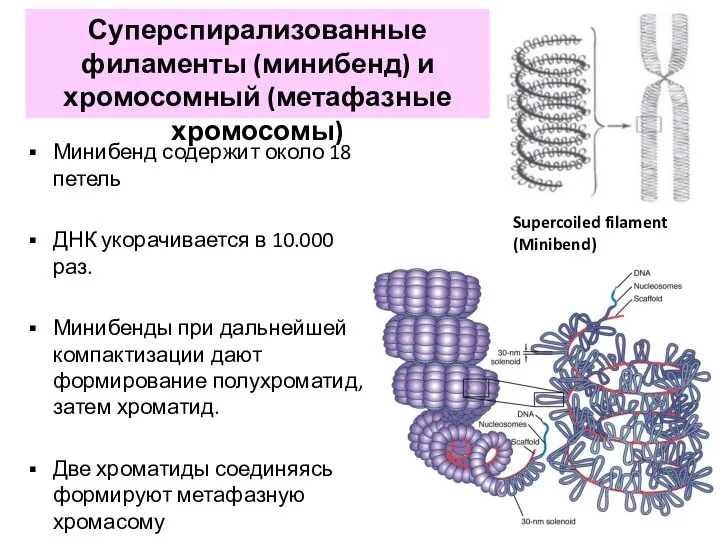
Минибенд содержит около 18 петель
ДНК укорачивается в 10.000 раз.
Минибенды при дальнейшей
Минибенд содержит около 18 петель
ДНК укорачивается в 10.000 раз.
Минибенды при дальнейшей
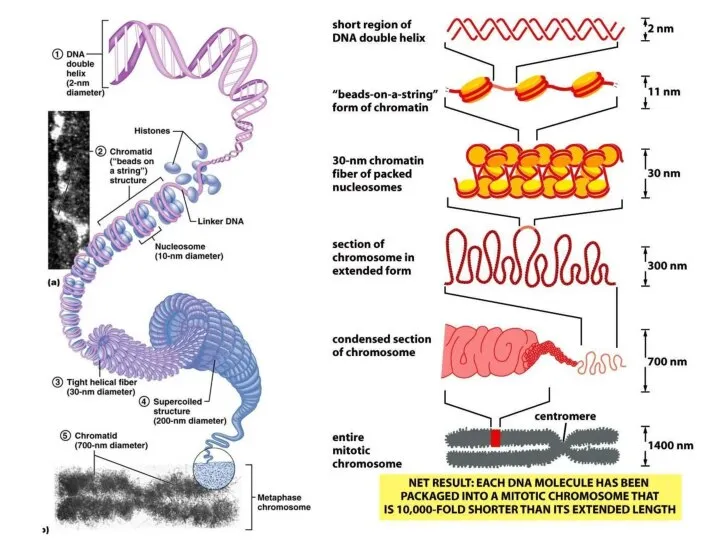
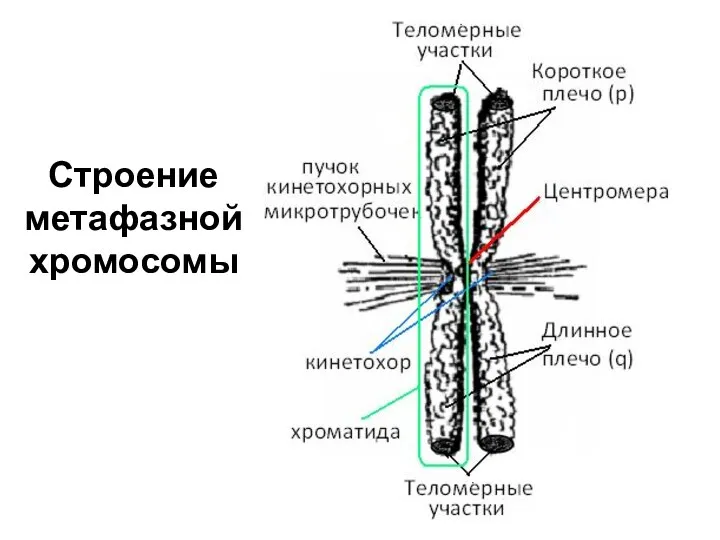
Строение метафазной хромосомы
Строение метафазной хромосомы
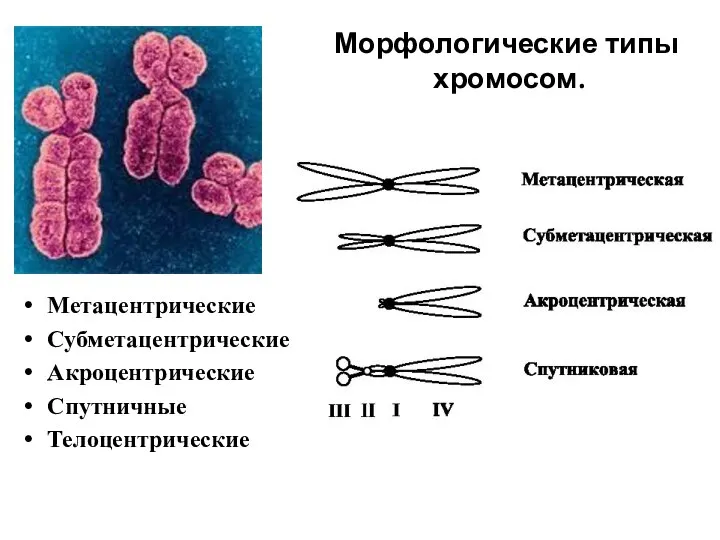
Морфологические типы
хромосом.
Метацентрические
Субметацентрические
Акроцентрические
Спутничные
Телоцентрические
Морфологические типы
хромосом.
Метацентрические
Субметацентрические
Акроцентрические
Спутничные
Телоцентрические
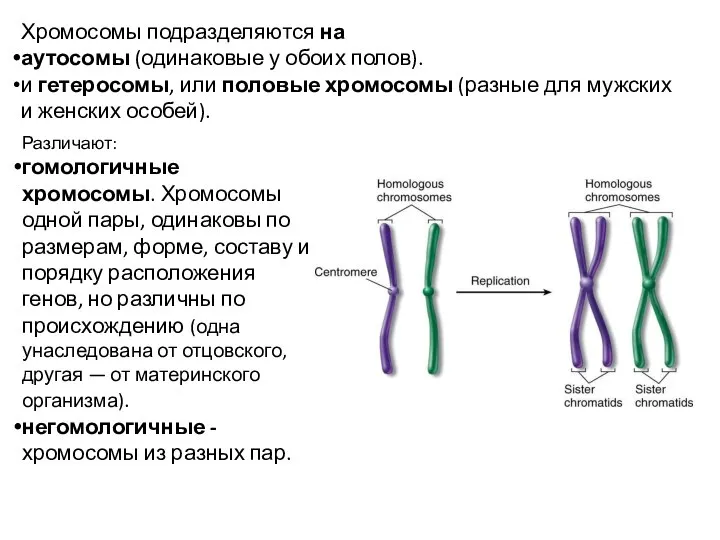
Хромосомы подразделяются на
аутосомы (одинаковые у обоих полов).
и гетеросомы, или
Хромосомы подразделяются на
аутосомы (одинаковые у обоих полов).
и гетеросомы, или
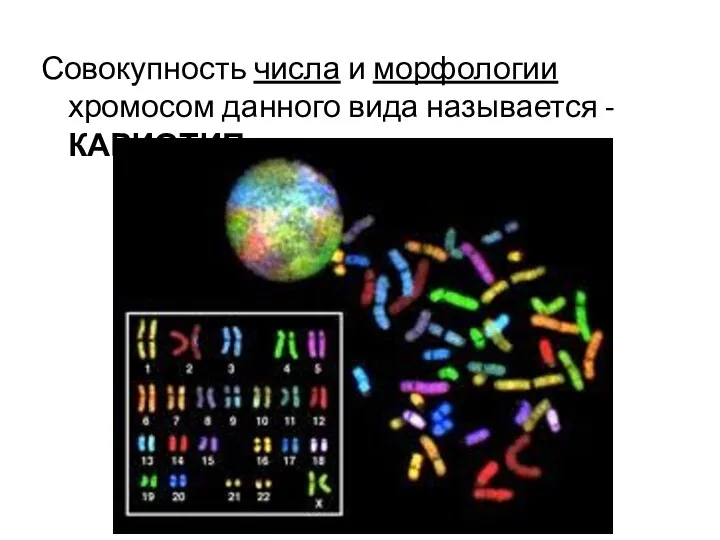
Совокупность числа и морфологии хромосом данного вида называется - КАРИОТИП
Совокупность числа и морфологии хромосом данного вида называется - КАРИОТИП
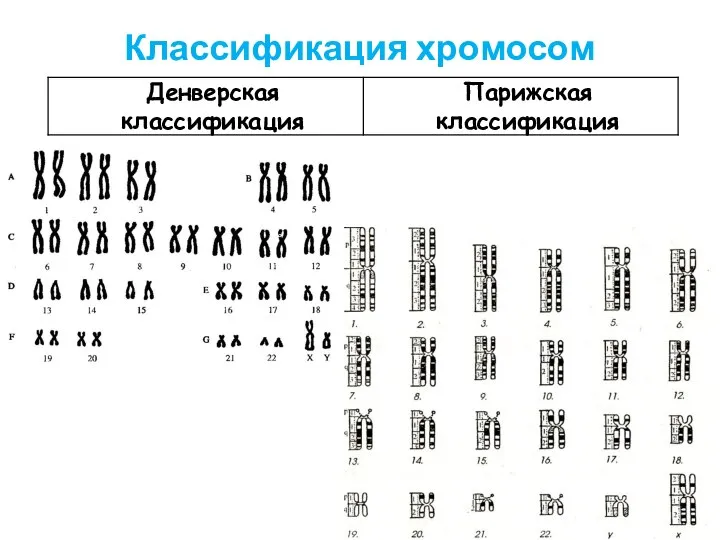
Классификация хромосом
Классификация хромосом

11-я хромосома человека
HBB — β-субъединица гемоглобина
16-я хромосома
человека
2-я хромосома
человека
11-я хромосома человека
HBB — β-субъединица гемоглобина
16-я хромосома
человека
2-я хромосома
человека



После утверждения в 20-х годах ХХ в. хромосомной теории наследственности биологи
После утверждения в 20-х годах ХХ в. хромосомной теории наследственности биологи

Нуклеиновые кислоты
Это природные высокомолекулярные органические биополимеры, обеспечивающие хранение и передачу наследственной
Нуклеиновые кислоты
Это природные высокомолекулярные органические биополимеры, обеспечивающие хранение и передачу наследственной
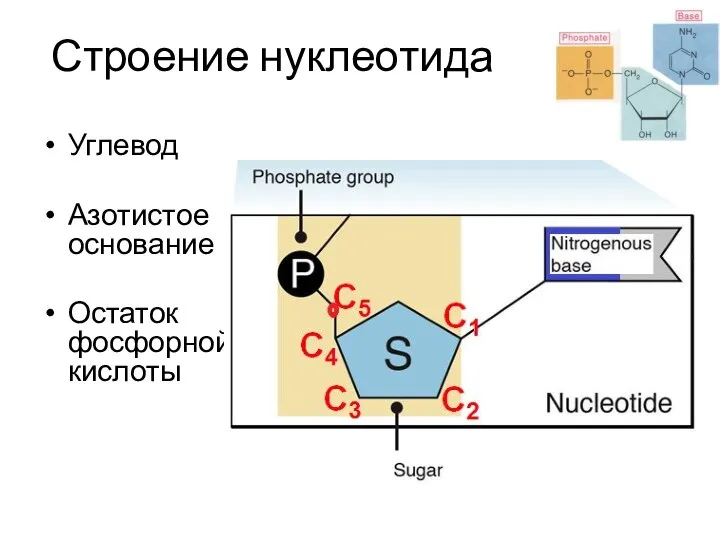
Строение нуклеотида
Углевод
Азотистое основание
Остаток фосфорной кислоты
Строение нуклеотида
Углевод
Азотистое основание
Остаток фосфорной кислоты
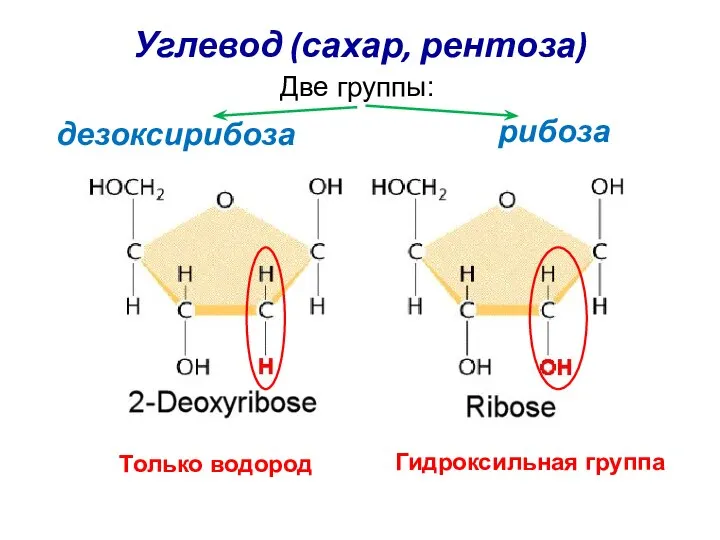
Углевод (сахар, рентоза)
рибоза
Две группы:
дезоксирибоза
Только водород
Гидроксильная группа
Углевод (сахар, рентоза)
рибоза
Две группы:
дезоксирибоза
Только водород
Гидроксильная группа
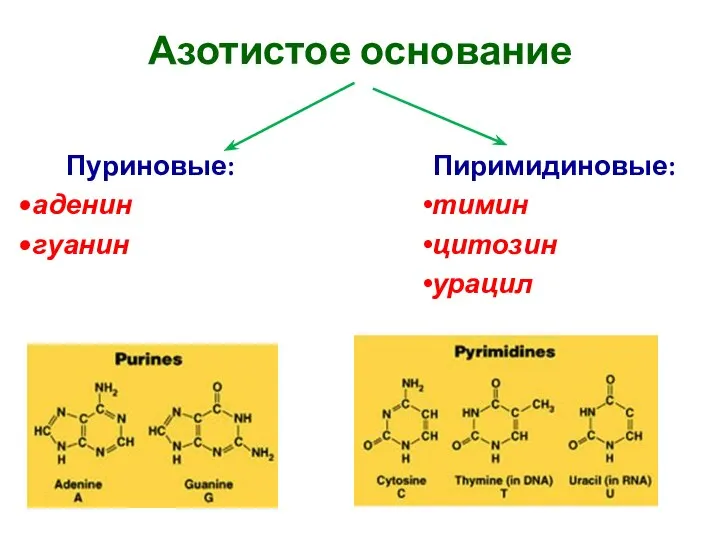
Азотистое основание
Пиримидиновые:
тимин
цитозин
урацил
Пуриновые:
аденин
гуанин
Азотистое основание
Пиримидиновые:
тимин
цитозин
урацил
Пуриновые:
аденин
гуанин
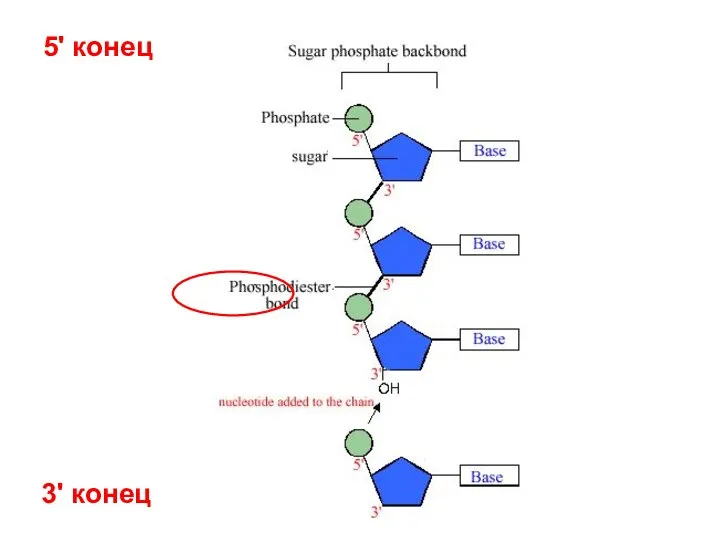
3' конец
5' конец
3' конец
5' конец

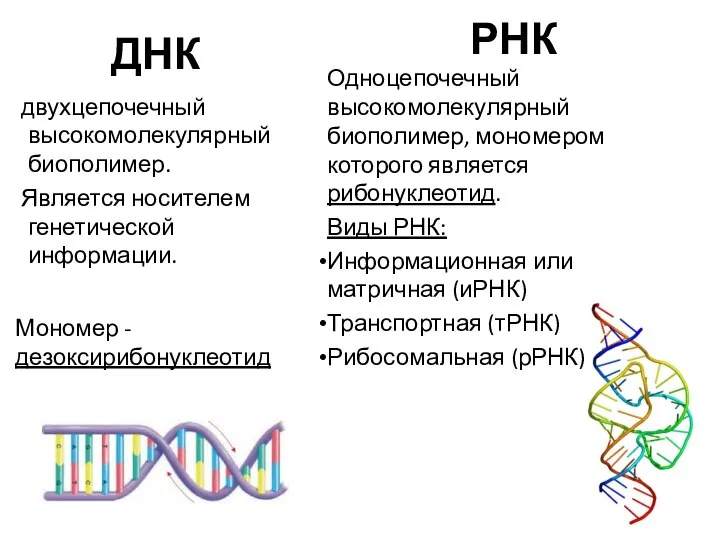
ДНК
двухцепочечный высокомолекулярный биополимер.
Является носителем генетической информации.
Мономер - дезоксирибонуклеотид
РНК
Одноцепочечный высокомолекулярный биополимер,
ДНК
двухцепочечный высокомолекулярный биополимер.
Является носителем генетической информации.
Мономер - дезоксирибонуклеотид
РНК
Одноцепочечный высокомолекулярный биополимер,

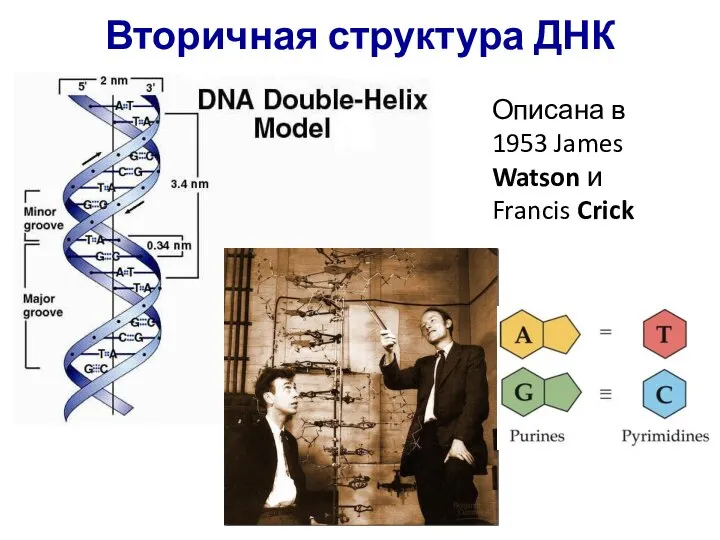
Описана в 1953 James Watson и Francis Crick
Вторичная структура ДНК
Описана в 1953 James Watson и Francis Crick
Вторичная структура ДНК
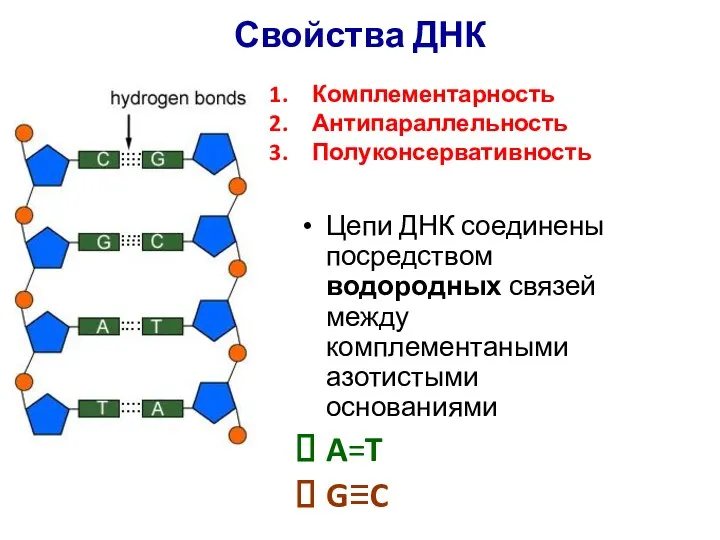
Свойства ДНК
Цепи ДНК соединены посредством водородных связей между комплементаными азотистыми основаниями
A=T
G≡C
Комплементарность
Антипараллельность
Полуконсервативность
Свойства ДНК
Цепи ДНК соединены посредством водородных связей между комплементаными азотистыми основаниями
A=T
G≡C
Комплементарность
Антипараллельность
Полуконсервативность
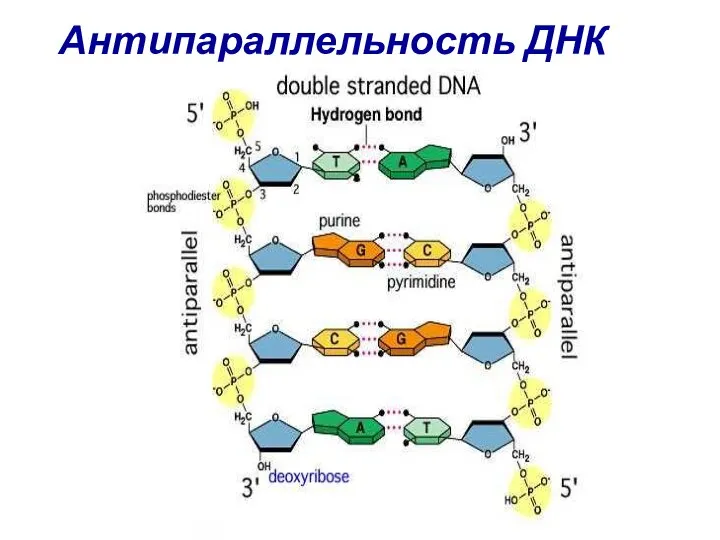
Антипараллельность ДНК
Антипараллельность ДНК

Доказательства роли ДНК
в передаче наследственной информации
(опыты по трансформации и
Доказательства роли ДНК в передаче наследственной информации (опыты по трансформации и
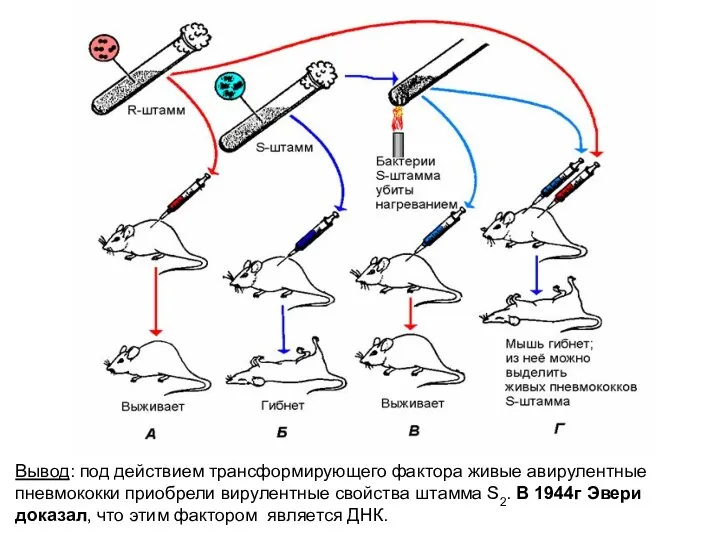
Вывод: под действием трансформирующего фактора живые авирулентные пневмококки приобрели вирулентные свойства
Вывод: под действием трансформирующего фактора живые авирулентные пневмококки приобрели вирулентные свойства


Трансдукция (от лат. transduction - перемещение) – процесс переноса фрагмента бактериальной
Трансдукция (от лат. transduction - перемещение) – процесс переноса фрагмента бактериальной

Первый из экспериментов был выполнен в 1952 году американскими генетиками Джошуа
Первый из экспериментов был выполнен в 1952 году американскими генетиками Джошуа
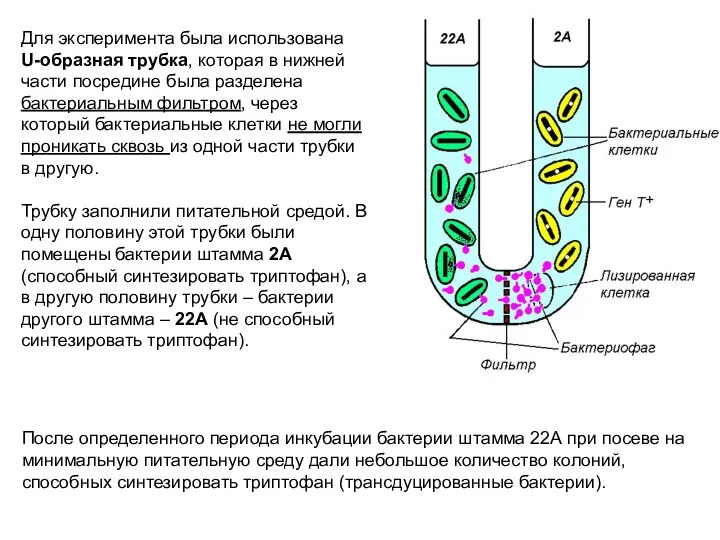
Для эксперимента была использована U-образная трубка, которая в нижней части посредине
Для эксперимента была использована U-образная трубка, которая в нижней части посредине
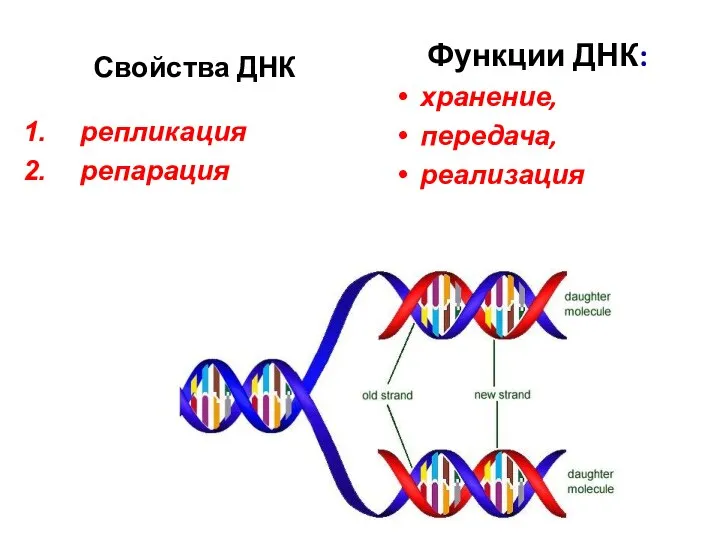
Свойства ДНК
Функции ДНК:
хранение,
передача,
реализация
репликация
репарация
Свойства ДНК
Функции ДНК:
хранение,
передача,
реализация
репликация
репарация
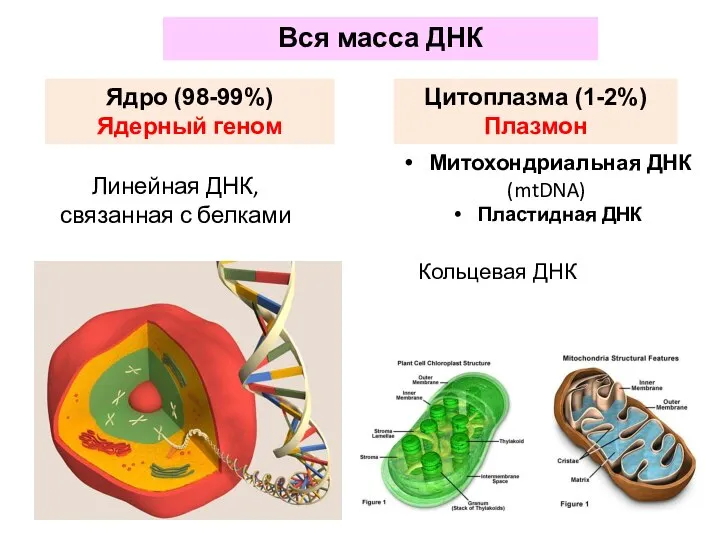
Вся масса ДНК
Ядро (98-99%)
Ядерный геном
Цитоплазма (1-2%)
Плазмон
Линейная ДНК, связанная с белками
Митохондриальная ДНК
(mtDNA)
Пластидная
Вся масса ДНК
Ядро (98-99%)
Ядерный геном
Цитоплазма (1-2%)
Плазмон
Линейная ДНК, связанная с белками
Митохондриальная ДНК
(mtDNA)
Пластидная
 Место птиц в системе живой природы
Место птиц в системе живой природы Животный мир. Тайга
Животный мир. Тайга Эволюция строения и функций органов и их систем.
Эволюция строения и функций органов и их систем. Механизмы регуляции сердечной деятельности
Механизмы регуляции сердечной деятельности African animals
African animals Лікарські засоби, препарати і способи для надання першої допомоги при отруєннях
Лікарські засоби, препарати і способи для надання першої допомоги при отруєннях  Слагаемые биотехнологического производства
Слагаемые биотехнологического производства Размножение можжевельника казацкого
Размножение можжевельника казацкого Интерактивное пособие по теме «Кровь и кровообращение» Часть 1 «Внутренняя среда организма» (для подготовки к ЕГЭ,ГИА и 8 класс
Интерактивное пособие по теме «Кровь и кровообращение» Часть 1 «Внутренняя среда организма» (для подготовки к ЕГЭ,ГИА и 8 класс Круглые черви и прогресс
Круглые черви и прогресс Природная зональность
Природная зональность Как сохранить обоняние
Как сохранить обоняние Влияние 6-гидроксидофамина при разных путях его введения на поведение мышей
Влияние 6-гидроксидофамина при разных путях его введения на поведение мышей Углеводы (Сn(H2O)m)
Углеводы (Сn(H2O)m) Анатомия и морфология листа
Анатомия и морфология листа  Чарльз Роберт Дарвин
Чарльз Роберт Дарвин Возникновение жизни на земле в Архее и Протерозое На протяжении тысячелетий людям казалось очевидным, что живая природа была созд
Возникновение жизни на земле в Архее и Протерозое На протяжении тысячелетий людям казалось очевидным, что живая природа была созд Авторы: Мартюкова Наталья – 5 класс Чухрова Юлия – 4 класс
Авторы: Мартюкова Наталья – 5 класс Чухрова Юлия – 4 класс Орёл-птица великая
Орёл-птица великая Фокина Ольга Степановна, учитель биологии
Фокина Ольга Степановна, учитель биологии Обитатели водоемов: рыбы, земноводные
Обитатели водоемов: рыбы, земноводные Влияние биологически активных добавок Трансфер факторов на организм человека Голубева Карина
Влияние биологически активных добавок Трансфер факторов на организм человека Голубева Карина  Презентация на тему "Воздействие человека и его деятельности на животных" - скачать презентации по Биологии
Презентация на тему "Воздействие человека и его деятельности на животных" - скачать презентации по Биологии Индивидуальное развитие организмов
Индивидуальное развитие организмов Способы размножение растений. Размножение споровых растений
Способы размножение растений. Размножение споровых растений Мейоз
Мейоз Строение, размножение, развитие млекопитающих
Строение, размножение, развитие млекопитающих Жизненный цикл клетки. Деление клетки. Митоз. Мейоз. Амитоз. Эндомитоз
Жизненный цикл клетки. Деление клетки. Митоз. Мейоз. Амитоз. Эндомитоз