- Введение в физиологию

Содержание
- 2. Механизмы регуляции 1. Биологически активные соединения (гуморальная регуляция). 2. Нейрогенная регуляция.
- 3. Пути влияния механизмов регуляции Все воздействия механизмов регуляции осуществляются через клеточную мембрану.
- 4. Схематическое изображение клетки
- 5. Схема клеточной мембраны бислой липидов – основа мембраны. интегративный белок, периферический белок, гликокаликс.
- 6. Пути чрезмембранного транспорта 1 – 3 идут по физико-химическим законам для которых необходима разница концентрации. Они
- 7. Na-K-насос Последовательные этапы работы насоса, который запускается поступившим внутрь клетки натрием: а) захват 3 Na+ б)
- 8. Мембранный потенциал (МП, ПП) В покое проницаемость мембран клеток немного выше для К+, чем для Na+.
- 9. Определение заряда мембраны с помощью внутриклеточного микроэлектрода
- 10. Возникновение потенциала действия (ПД) А -Фазы развития ПД 1 – деполяризации, 2 – овершут, 3 –
- 11. Функциональные изменения каналов при развитии ПД а – закрыты активационные ворота, б – открыты активационные ворота,
- 12. Состояние проницаемости мембраны к ионам при развитии потенциала действия При одновременном начале открытия натриевые каналы открываются
- 13. Соотношение ПД и рефрактерности 1 – фаза абсолютной рефрактерности, 2 – ф. относительной рефрактерности, 3 -
- 14. Проводимость ПД возникает между деполяризованной областью мембраны и ее невозбужден-ным участком. Разность потенциалов здесь во много
- 15. Проведение ПД по миелинизированному нервному волокну: сальтаторно - прыжками
- 16. Физиология скелетных мышц
- 17. Схема строения мышечного волокна Саркомер - с двух сторон ограничен Z – мембранами. Толстые – миозиновые,
- 18. Саркоплазма В саркоплазме находится весь набор типичных для любой клетки органоидов. Особо следует подчеркнуть наличие: -
- 19. Актиновые миофиламенты Актиновые филаменты, скомпанованы из двух актиновых нитей, представляющих собой как бы бусинки глобулярных молекул
- 20. Схема строения и взаимосвязь актиновых и миозиновых филаментов Миозиновые филаменты образуются более чем двумястами молекулами миозина.
- 21. Двигательные единицы К каждому мышечному волокну подходит отросток мотонейрона. Как правило, 1 мотонейрон иннервирует несколько мышечных
- 22. Нервно-мышечный синапс 1 - пресинаптическая мембрана, 2 - пузырьки с ацетилхолином, 3 - митохондрии, 4 -
- 23. Передача ПД через синапс 1 – везикула, 2 – медиатор (ацетилхолин, АХ), 3 – холинорецептор, 4
- 24. Взаимодействие медиатора с постсинаптической мебраной Медиатор (АХ) диффундирует по синаптической жидкости и большая часть молекул его
- 25. МП Мышечное волокно имеет мембранный потенциал -80 - -90 мВ. Для того, чтобы вызвать возникновение возбуждения
- 26. Явление суммации. Обычно для передачи одного ПД высвобождается до миллиона молекул АХ (200-300 везикул). Но при
- 27. Для чего необходимо поступление ПД к мышце? ВПСП распространяется по сарколемме. Проскакивает по мембране саркоплазматического ретикулума.
- 28. Депо кальция – саркоплазматический ретикулум 1- миофибриллы, 2 – саркоплазматический ретикулум, 3 – цистерны, 4 –
- 29. Роль кальция в мышечном сокращении Последовательные этапы: а – расслабление, б – соединение миозиновых головок с
- 30. «Шаговый» механизм
- 31. Различные режимы сокращения мышц А - одиночное сокращение, Б – неполный тетанус, В – полный тетанус.
- 32. Анатомический и физиологический поперечники мышц В естественных условиях на проявление силы мышцы оказывает влияние не только
- 33. Роль АТФ в мышце АТФ в мышце необходима для: а) сокращения (образования мостиков); б) расслабления (разрыва
- 34. Максимальная мощность путей ресинтеза АТФ: а) фосфагенный (КФ) - 3,6 моль АТФ/мин, б) гликолитический - 1,2
- 35. Типы ДЕ (двигательные единицы) - процентное соотношение врожденное и у разных людей различное (спринтеры, стайеры) Быстрые
- 36. Гладкие мышцы Гладкие мышцы находятся в стенке внут-ренних органов, сосудов, коже. Структурной единицей их является вытянутой
- 37. Компановка сократимых миофиламентов внутри клетки. Актиновые филаменты сгруппированы в пучки, которые время от времени образуют уплотнения
- 38. Разновидности деполяризации гладко-мышечных клеток Один из них (а) напоминает ПД скелетной мышцы, отличаясь от нее большей
- 39. Пейсмекеры Среди гладкомышечных клеток, образующих функциональный синцитий, имеются такие, которые обладают пейсмекерными свойствами (от англ. рacemaker
- 40. Нейронная регуляция 1. Отличие нейронной регуляции от гуморальной. 2. Рефлекторный принцип регуляции. 3. Физиологическая характеристика нерва.
- 41. Отличие нейронной регуляции от гуморальной. Точность «адресата». Рефлекторный принцип регуляции. Включение на конечном этапе гуморальное звено
- 42. Нейроны 1 - мультиполярный нейрон; 2 - биполярный нейрон; 3 - псевдополярный нейрон; 4 - униполярный
- 43. Рефлекторный принцип организации нейронной регуляции Рефлексом называется стереотипная реакция организма или его отдельных органов на сенсорный
- 44. Первично чувствующие рецепторы Это нервные окончания чувствительных нейронов. При действии на них раздражителя возникает ПД. Суммация
- 45. Вторично чувствующие рецепторы Это специализированные клетки, особо чувствительные к действия какого-либо раздражителя. При возникновении в них
- 46. Основной принцип рефлекторной регуляции Обеспечивается точность регуляции, в основе которой лежит получение информации от органа, ее
- 47. В ЦНС нейронов лишь 10%, 90% - нейроглия Глиальные клетки: Астроциты Резорбция ряда медиаторов Временное поглощение
- 48. Астроцит и схема гематоэнцефалического барьера Астроцит создает преграду между нервом и кровеносным капилляром, поэтому к нервам
- 49. Функциональные показатели нейронов ПП – от –60 мВ до –90 мВ Аксонный холмик (начало аксона): ПП
- 50. Рефрактерность и лабильность Абсолютный рефрактерный период примерно такой же, как и длительность ПД. В крупных нейронах
- 51. Синапсы ЦНС Межнейронные синапсы: 1 - аксо-соматический синапс; 2 - аксо-дендритный синапс; 3 - аксо-дендритный синапс
- 52. Основные медиаторы ЦНС 1. Амины (ацетилхолин, норадреналин, адреналин, дофамин, серотонин). 2. Аминокислоты (глицин, глутамин, аспарагиновая, ГАМК
- 53. Медиаторы - ионотропные и метаботропные. Ионотропные медиаторы после взаимодействия с рецепторами постсинаптической мембраны изменяют проницаемость ионных
- 54. Возбуждающий постсинаптический потенциал (ВПСП) а, б - деполяризация не достигает критического уровня, в - результат суммации
- 55. Виды суммации в ЦНС В ЦНС два вида суммации: Временная суммация – как в нервно-мышечном синапсе.
- 56. Разновидности торможения А – пресинаптическое торможение, Б – постсинатическое торможение: В – возбуждающий нейрон, Т -
- 57. Развитие гиперполяризации на постсинаптической мембране тормозного синапса А - Развитие гиперполяризации постсинаптической мембраны тормозного синапса. Б
- 58. Эфапс (нексус)
- 59. Электро энцефалограмма (ЭЭГ) А - при открытых глазах (видны по преимуществу β-волны); Б - при закрытых
- 60. Свойства нервных центров (нервный центр – скопление нейронов, выполняющих какую-либо функцию) А – конвергенция. Характерно для
- 61. Межцентральные взаимодействия Доминанта Один из основных принципов, обеспечения межцентральных функциональных взаимосвязей – ДОМИНАНТА. При наличии одновременного
- 62. Интегративные механизмы мозга Это системы нервных клеток (центров), которые не выполняют специфические функции (рефлексы), они регулируют
- 63. Влияния ретикулярной формации Восходящее влияние ретикулярной формации заключается во влиянии верхних отделов ретикулярной формации ствола головного
- 64. Аминергические системы ствола мозга По названию медиаторов различают: Норадренергическая система. ДОФАминергическая система. Серотонинергическая.
- 65. Аминоспецифические системы Нейроны, медиаторами которых являются моноамины (серотонин, норадреналин и дофамин), также участвуют в объединении различных
- 67. Скачать презентацию

Механизмы регуляции
1. Биологически активные соединения (гуморальная регуляция).
2. Нейрогенная регуляция.
Механизмы регуляции
1. Биологически активные соединения (гуморальная регуляция).
2. Нейрогенная регуляция.

Пути влияния механизмов регуляции
Все воздействия механизмов регуляции осуществляются через клеточную мембрану.
Пути влияния механизмов регуляции
Все воздействия механизмов регуляции осуществляются через клеточную мембрану.
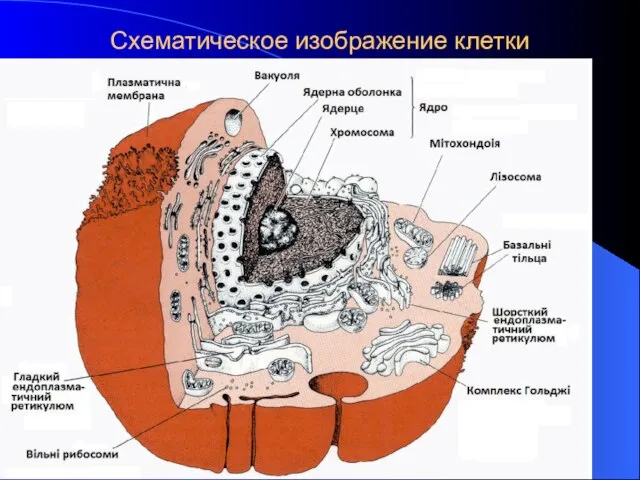
Схематическое изображение клетки
Схематическое изображение клетки

Схема клеточной мембраны
бислой липидов – основа мембраны.
интегративный белок,
периферический белок,
Схема клеточной мембраны
бислой липидов – основа мембраны.
интегративный белок,
периферический белок,
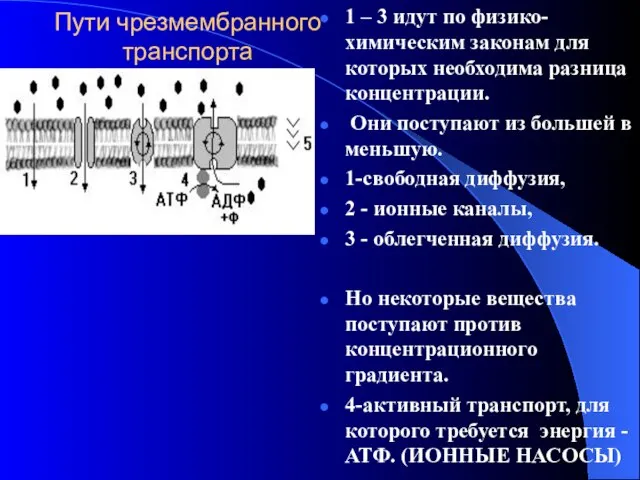
Пути чрезмембранного транспорта
1 – 3 идут по физико-химическим законам для которых
Пути чрезмембранного транспорта
1 – 3 идут по физико-химическим законам для которых
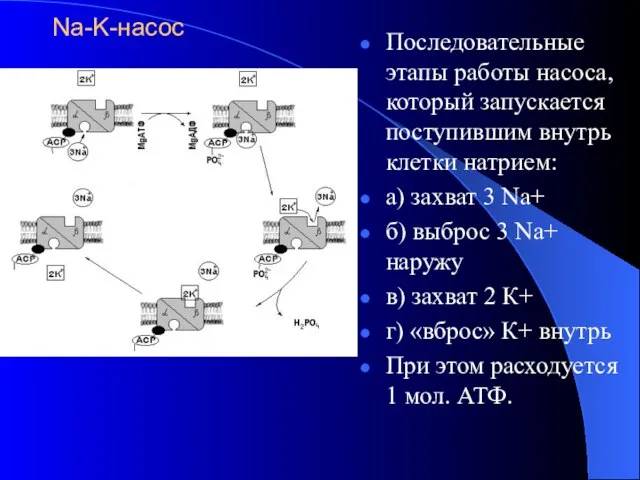
Na-K-насос
Последовательные этапы работы насоса, который запускается поступившим внутрь клетки натрием:
а) захват
Na-K-насос
Последовательные этапы работы насоса, который запускается поступившим внутрь клетки натрием:
а) захват

Мембранный потенциал (МП, ПП)
В покое проницаемость мембран клеток немного выше для
Мембранный потенциал (МП, ПП)
В покое проницаемость мембран клеток немного выше для
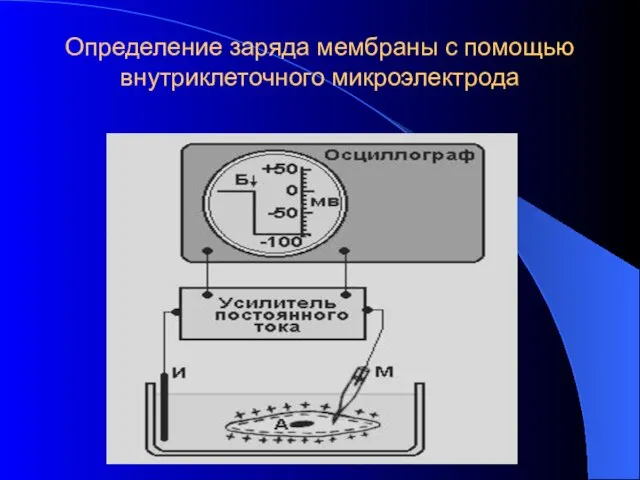
Определение заряда мембраны с помощью внутриклеточного микроэлектрода
Определение заряда мембраны с помощью внутриклеточного микроэлектрода

Возникновение потенциала действия (ПД)
А -Фазы развития ПД
1 – деполяризации,
2 – овершут,
3
Возникновение потенциала действия (ПД)
А -Фазы развития ПД
1 – деполяризации,
2 – овершут,
3
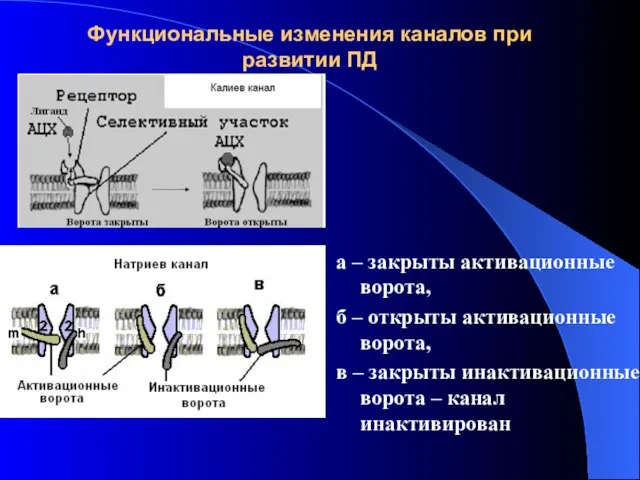
Функциональные изменения каналов при развитии ПД
а – закрыты активационные ворота,
б –
Функциональные изменения каналов при развитии ПД
а – закрыты активационные ворота,
б –
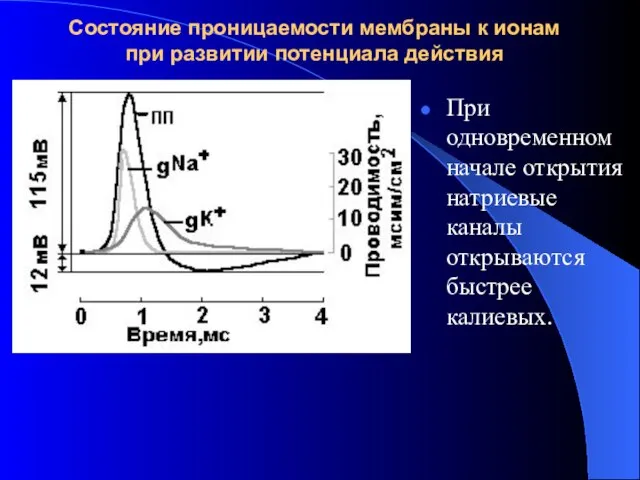
Состояние проницаемости мембраны к ионам при развитии потенциала действия
При одновременном начале
Состояние проницаемости мембраны к ионам при развитии потенциала действия
При одновременном начале
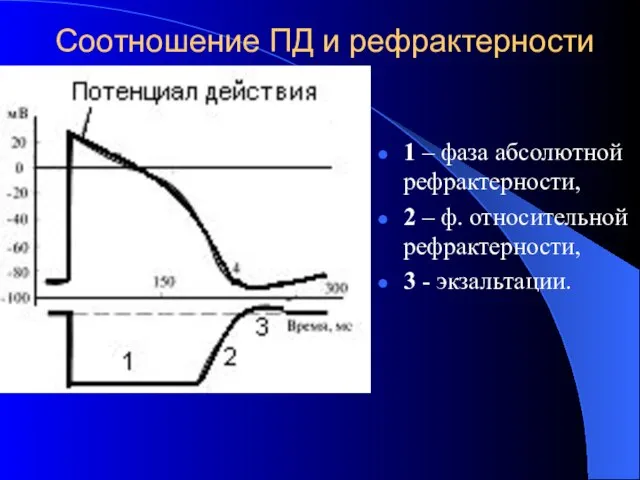
Соотношение ПД и рефрактерности
1 – фаза абсолютной рефрактерности,
2 – ф.
Соотношение ПД и рефрактерности
1 – фаза абсолютной рефрактерности,
2 – ф.
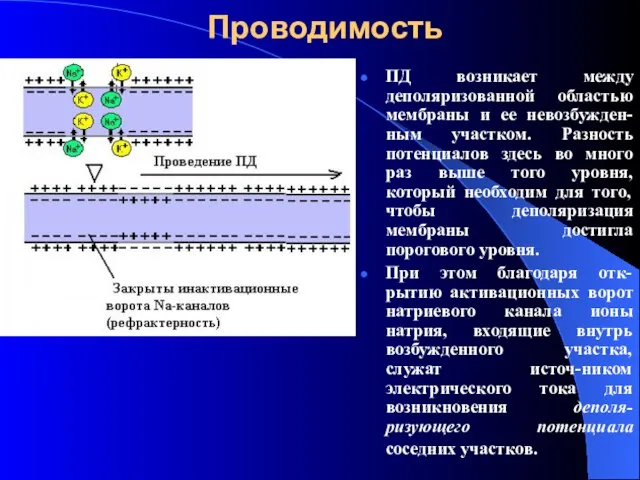
Проводимость
ПД возникает между деполяризованной областью мембраны и ее невозбужден-ным участком. Разность
Проводимость
ПД возникает между деполяризованной областью мембраны и ее невозбужден-ным участком. Разность
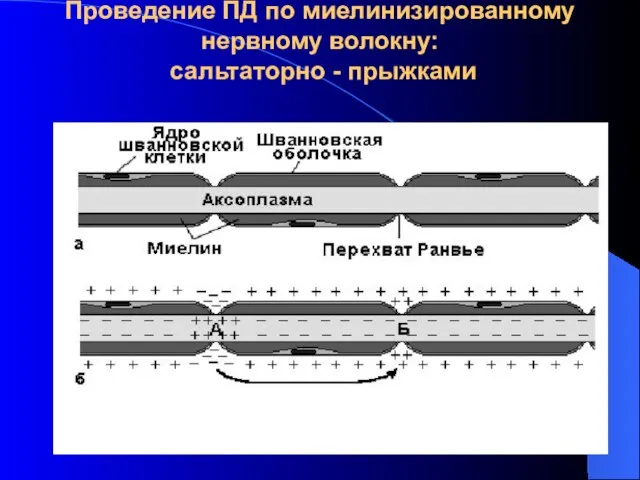
Проведение ПД по миелинизированному нервному волокну: сальтаторно - прыжками
Проведение ПД по миелинизированному нервному волокну: сальтаторно - прыжками

Физиология скелетных мышц
Физиология скелетных мышц
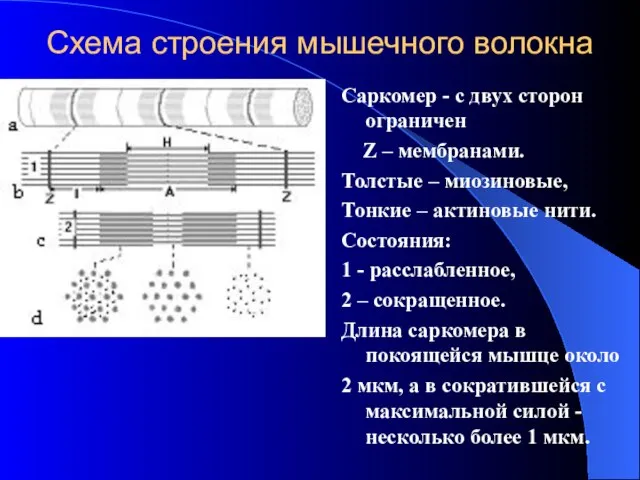
Схема строения мышечного волокна
Саркомер - с двух сторон ограничен
Z
Схема строения мышечного волокна
Саркомер - с двух сторон ограничен
Z

Саркоплазма
В саркоплазме находится весь набор типичных для любой клетки органоидов.
Особо следует
Саркоплазма
В саркоплазме находится весь набор типичных для любой клетки органоидов.
Особо следует

Актиновые миофиламенты
Актиновые филаменты, скомпанованы из двух актиновых нитей, представляющих собой как
Актиновые миофиламенты
Актиновые филаменты, скомпанованы из двух актиновых нитей, представляющих собой как

Схема строения и взаимосвязь актиновых и миозиновых филаментов
Миозиновые филаменты образуются более
Схема строения и взаимосвязь актиновых и миозиновых филаментов
Миозиновые филаменты образуются более

Двигательные единицы
К каждому мышечному волокну подходит отросток мотонейрона.
Как правило, 1 мотонейрон
Двигательные единицы
К каждому мышечному волокну подходит отросток мотонейрона.
Как правило, 1 мотонейрон
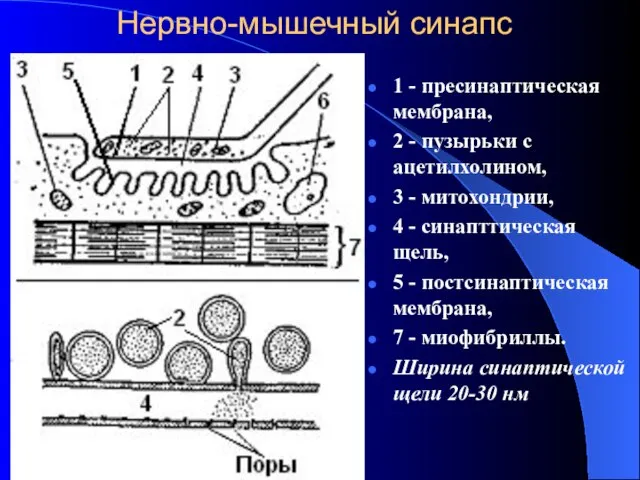
Нервно-мышечный синапс
1 - пресинаптическая мембрана,
2 - пузырьки с ацетилхолином,
3
Нервно-мышечный синапс
1 - пресинаптическая мембрана,
2 - пузырьки с ацетилхолином,
3
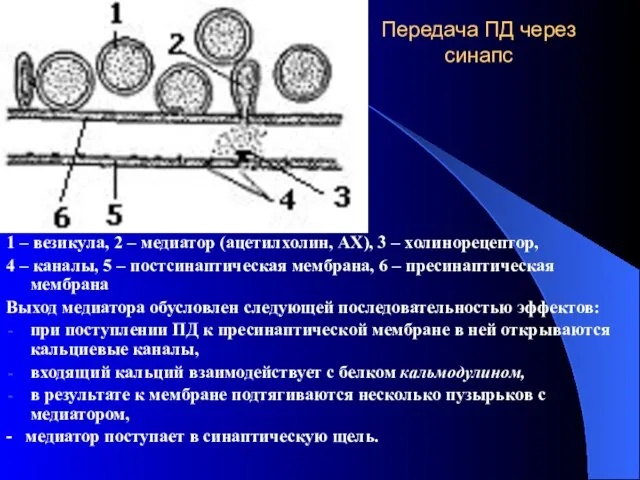
Передача ПД через синапс
1 – везикула, 2 – медиатор (ацетилхолин, АХ),
Передача ПД через синапс
1 – везикула, 2 – медиатор (ацетилхолин, АХ),

Взаимодействие медиатора с постсинаптической мебраной
Медиатор (АХ) диффундирует по синаптической жидкости и
Взаимодействие медиатора с постсинаптической мебраной
Медиатор (АХ) диффундирует по синаптической жидкости и

МП
Мышечное волокно имеет мембранный потенциал -80 - -90 мВ. Для того,
МП
Мышечное волокно имеет мембранный потенциал -80 - -90 мВ. Для того,
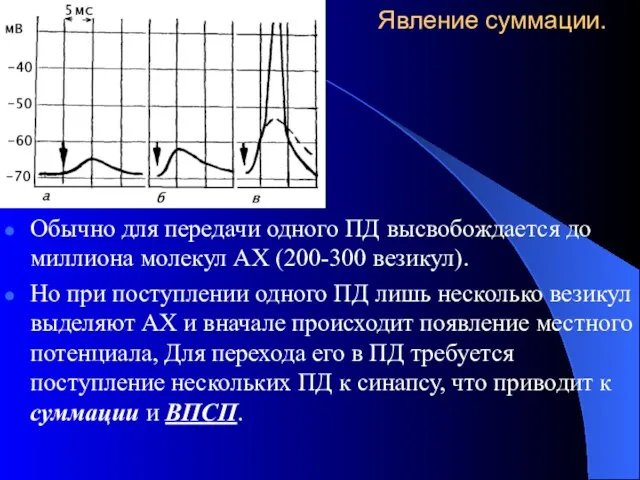
Явление суммации.
Обычно для передачи одного ПД высвобождается до миллиона молекул АХ
Явление суммации.
Обычно для передачи одного ПД высвобождается до миллиона молекул АХ

Для чего необходимо поступление ПД к мышце?
ВПСП распространяется по сарколемме.
Для чего необходимо поступление ПД к мышце?
ВПСП распространяется по сарколемме.
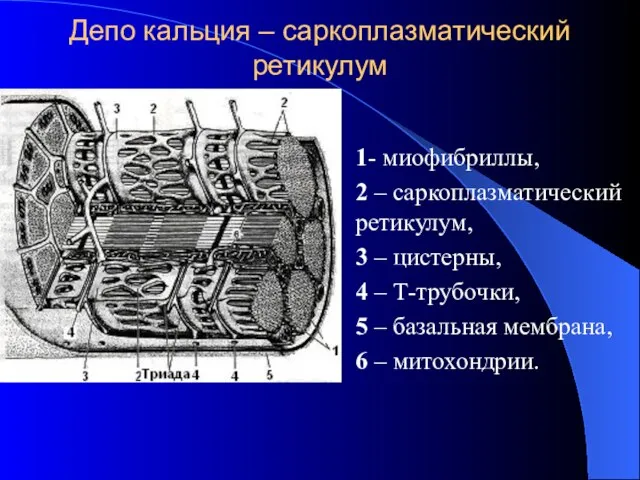
Депо кальция – саркоплазматический ретикулум
1- миофибриллы,
2 – саркоплазматический ретикулум,
Депо кальция – саркоплазматический ретикулум
1- миофибриллы,
2 – саркоплазматический ретикулум,

Роль кальция в мышечном сокращении
Последовательные этапы:
а – расслабление,
б – соединение миозиновых
Роль кальция в мышечном сокращении
Последовательные этапы:
а – расслабление,
б – соединение миозиновых
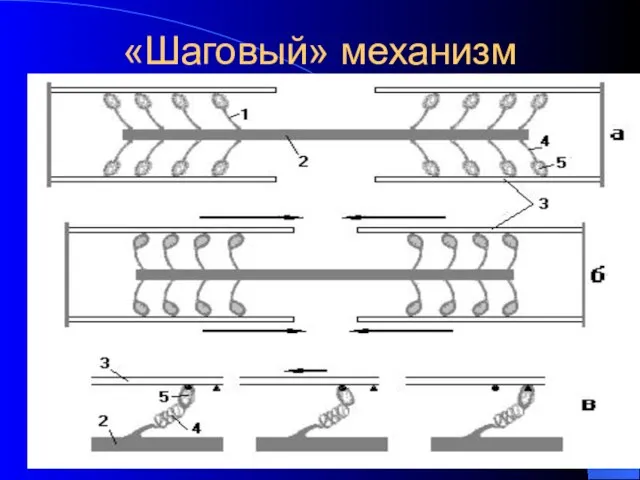
«Шаговый» механизм
«Шаговый» механизм
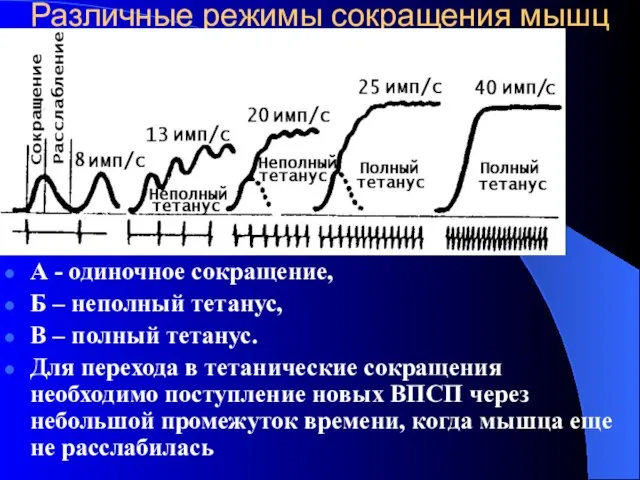
Различные режимы сокращения мышц
А - одиночное сокращение,
Б – неполный тетанус,
Различные режимы сокращения мышц
А - одиночное сокращение,
Б – неполный тетанус,

Анатомический и физиологический поперечники мышц
В естественных условиях на проявление силы мышцы
Анатомический и физиологический поперечники мышц
В естественных условиях на проявление силы мышцы

Роль АТФ в мышце
АТФ в мышце необходима для: а) сокращения (образования
Роль АТФ в мышце
АТФ в мышце необходима для: а) сокращения (образования

Максимальная мощность путей ресинтеза АТФ:
а) фосфагенный (КФ) - 3,6 моль АТФ/мин,
Максимальная мощность путей ресинтеза АТФ:
а) фосфагенный (КФ) - 3,6 моль АТФ/мин,

Типы ДЕ (двигательные единицы)
- процентное соотношение врожденное и у разных людей
Типы ДЕ (двигательные единицы) - процентное соотношение врожденное и у разных людей

Гладкие мышцы
Гладкие мышцы находятся в стенке внут-ренних органов, сосудов, коже. Структурной
Гладкие мышцы
Гладкие мышцы находятся в стенке внут-ренних органов, сосудов, коже. Структурной

Компановка сократимых миофиламентов внутри клетки.
Актиновые филаменты сгруппированы в пучки, которые время
Компановка сократимых миофиламентов внутри клетки.
Актиновые филаменты сгруппированы в пучки, которые время
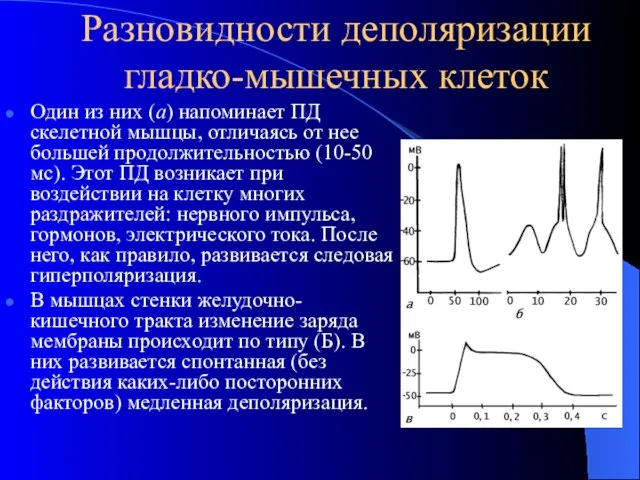
Разновидности деполяризации гладко-мышечных клеток
Один из них (а) напоминает ПД скелетной
Разновидности деполяризации гладко-мышечных клеток
Один из них (а) напоминает ПД скелетной

Пейсмекеры
Среди гладкомышечных клеток, образующих функциональный синцитий, имеются такие, которые обладают пейсмекерными
Пейсмекеры
Среди гладкомышечных клеток, образующих функциональный синцитий, имеются такие, которые обладают пейсмекерными

Нейронная регуляция
1. Отличие нейронной регуляции от гуморальной.
2. Рефлекторный принцип регуляции.
3. Физиологическая
Нейронная регуляция
1. Отличие нейронной регуляции от гуморальной.
2. Рефлекторный принцип регуляции.
3. Физиологическая

Отличие нейронной регуляции от гуморальной.
Точность «адресата».
Рефлекторный принцип регуляции.
Включение на конечном этапе
Отличие нейронной регуляции от гуморальной.
Точность «адресата».
Рефлекторный принцип регуляции.
Включение на конечном этапе
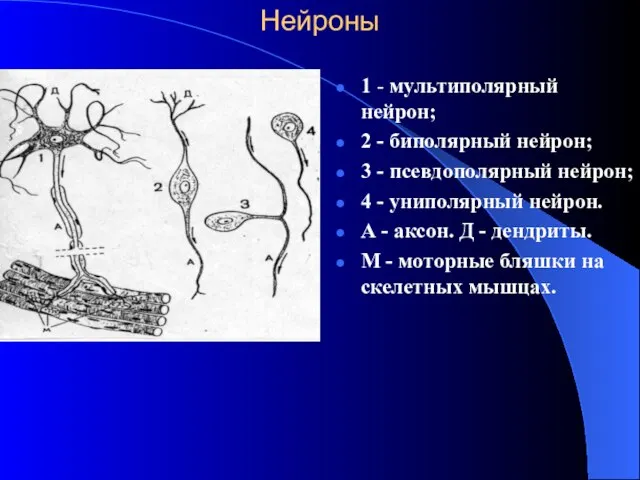
Нейроны
1 - мультиполярный нейрон;
2 - биполярный нейрон;
3 - псевдополярный
Нейроны
1 - мультиполярный нейрон;
2 - биполярный нейрон;
3 - псевдополярный

Рефлекторный принцип организации нейронной регуляции
Рефлексом называется стереотипная реакция организма или его
Рефлекторный принцип организации нейронной регуляции Рефлексом называется стереотипная реакция организма или его

Первично чувствующие рецепторы
Это нервные окончания чувствительных нейронов. При действии на них
Первично чувствующие рецепторы
Это нервные окончания чувствительных нейронов. При действии на них

Вторично чувствующие рецепторы
Это специализированные клетки, особо чувствительные к действия какого-либо раздражителя.
Вторично чувствующие рецепторы
Это специализированные клетки, особо чувствительные к действия какого-либо раздражителя.

Основной принцип рефлекторной регуляции
Обеспечивается точность регуляции, в основе которой лежит получение
Основной принцип рефлекторной регуляции
Обеспечивается точность регуляции, в основе которой лежит получение

В ЦНС нейронов лишь 10%, 90% - нейроглия
Глиальные клетки:
Астроциты
Резорбция
В ЦНС нейронов лишь 10%, 90% - нейроглия
Глиальные клетки:
Астроциты
Резорбция

Астроцит и схема гематоэнцефалического барьера
Астроцит создает преграду между нервом и кровеносным
Астроцит и схема гематоэнцефалического барьера
Астроцит создает преграду между нервом и кровеносным

Функциональные показатели нейронов
ПП – от –60 мВ до –90 мВ
Аксонный холмик
Функциональные показатели нейронов
ПП – от –60 мВ до –90 мВ
Аксонный холмик

Рефрактерность и лабильность
Абсолютный рефрактерный период примерно такой же, как и длительность
Рефрактерность и лабильность
Абсолютный рефрактерный период примерно такой же, как и длительность

Синапсы ЦНС
Межнейронные синапсы:
1 - аксо-соматический синапс;
2 - аксо-дендритный синапс;
Синапсы ЦНС
Межнейронные синапсы:
1 - аксо-соматический синапс;
2 - аксо-дендритный синапс;

Основные медиаторы ЦНС
1. Амины (ацетилхолин, норадреналин, адреналин, дофамин, серотонин).
2. Аминокислоты (глицин,
Основные медиаторы ЦНС
1. Амины (ацетилхолин, норадреналин, адреналин, дофамин, серотонин).
2. Аминокислоты (глицин,

Медиаторы - ионотропные и метаботропные.
Ионотропные медиаторы после взаимодействия с рецепторами постсинаптической
Медиаторы - ионотропные и метаботропные.
Ионотропные медиаторы после взаимодействия с рецепторами постсинаптической
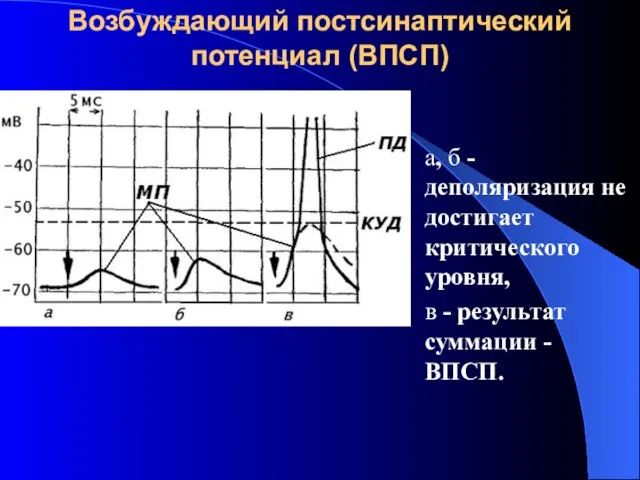
Возбуждающий постсинаптический потенциал (ВПСП)
а, б - деполяризация не достигает критического
Возбуждающий постсинаптический потенциал (ВПСП)
а, б - деполяризация не достигает критического

Виды суммации в ЦНС
В ЦНС два вида суммации:
Временная суммация – как
Виды суммации в ЦНС
В ЦНС два вида суммации:
Временная суммация – как
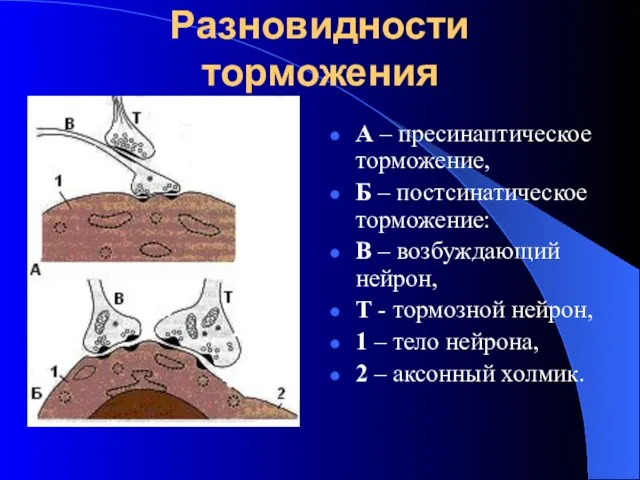
Разновидности торможения
А – пресинаптическое торможение,
Б – постсинатическое торможение:
В
Разновидности торможения
А – пресинаптическое торможение,
Б – постсинатическое торможение:
В

Развитие гиперполяризации на постсинаптической мембране тормозного синапса
А - Развитие гиперполяризации
Развитие гиперполяризации на постсинаптической мембране тормозного синапса
А - Развитие гиперполяризации
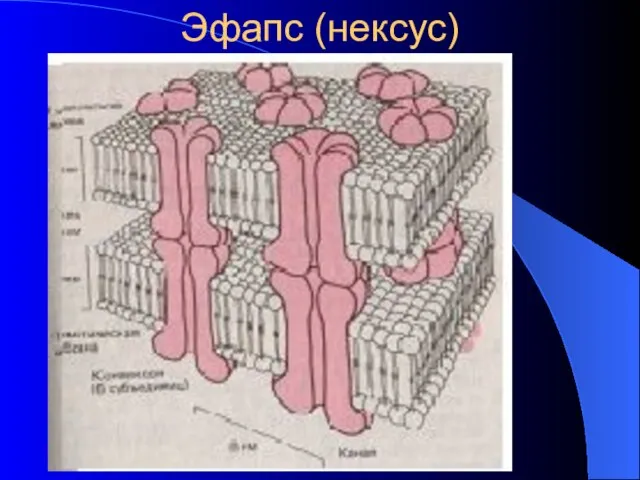
Эфапс (нексус)
Эфапс (нексус)

Электро энцефалограмма (ЭЭГ)
А - при открытых глазах (видны по преимуществу β-волны);
Электро энцефалограмма (ЭЭГ)
А - при открытых глазах (видны по преимуществу β-волны);
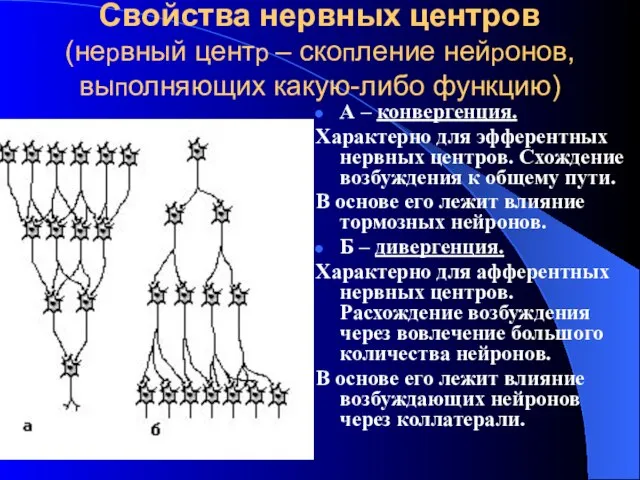
Свойства нервных центров
(нервный центр – скопление нейронов, выполняющих какую-либо функцию)
А –
Свойства нервных центров
(нервный центр – скопление нейронов, выполняющих какую-либо функцию)
А –

Межцентральные взаимодействия
Доминанта
Один из основных принципов, обеспечения межцентральных функциональных взаимосвязей – ДОМИНАНТА.
При
Межцентральные взаимодействия
Доминанта
Один из основных принципов, обеспечения межцентральных функциональных взаимосвязей – ДОМИНАНТА.
При

Интегративные механизмы мозга
Это системы нервных клеток (центров), которые не выполняют
Интегративные механизмы мозга
Это системы нервных клеток (центров), которые не выполняют

Влияния ретикулярной формации
Восходящее влияние ретикулярной формации заключается во влиянии верхних отделов
Влияния ретикулярной формации
Восходящее влияние ретикулярной формации заключается во влиянии верхних отделов

Аминергические системы ствола мозга
По названию медиаторов различают:
Норадренергическая система.
ДОФАминергическая система.
Серотонинергическая.
Аминергические системы ствола мозга
По названию медиаторов различают:
Норадренергическая система.
ДОФАминергическая система.
Серотонинергическая.

Аминоспецифические системы
Нейроны, медиаторами которых являются моноамины (серотонин, норадреналин и дофамин), также
Аминоспецифические системы
Нейроны, медиаторами которых являются моноамины (серотонин, норадреналин и дофамин), также
 Регуляция внутренних органов: роль отделов ЦНС, вегетативной нервной системы
Регуляция внутренних органов: роль отделов ЦНС, вегетативной нервной системы Вирусы: строение, виды, значение. Прионы. Вироиды
Вирусы: строение, виды, значение. Прионы. Вироиды Исчезнувшие животные
Исчезнувшие животные Презентация на тему "Влияние ионизирующих излучений на живые организмы" - скачать презентации по Биологии
Презентация на тему "Влияние ионизирующих излучений на живые организмы" - скачать презентации по Биологии Взаимодействие между животными
Взаимодействие между животными Взаимодействие генов
Взаимодействие генов Распространение плодов и семян
Распространение плодов и семян Характеристика царства растения
Характеристика царства растения Водоросли
Водоросли Презентация на тему "Гормоны и их роль в обменных процессах" - скачать презентации по Биологии
Презентация на тему "Гормоны и их роль в обменных процессах" - скачать презентации по Биологии МОУ СОШ п. АЛЯБЬЕВСКИЙ КЛАСС ПАУКООБРАЗНЫЕ – ХЕЛИЦЕРОВЫЕ, особенности строения и жизнедеятельности Презентацию подготовил:
МОУ СОШ п. АЛЯБЬЕВСКИЙ КЛАСС ПАУКООБРАЗНЫЕ – ХЕЛИЦЕРОВЫЕ, особенности строения и жизнедеятельности Презентацию подготовил:  Презентация на тему "Пищеварительная система" - скачать презентации по Биологии
Презентация на тему "Пищеварительная система" - скачать презентации по Биологии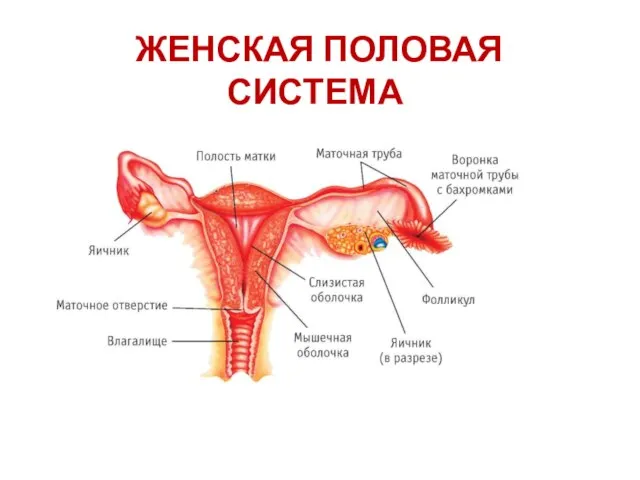 Женская половая система
Женская половая система Строение цветка
Строение цветка Презентация на тему Человек – часть живой природы
Презентация на тему Человек – часть живой природы Яблочный спас познавательная игра по биологии, 6 класс Автор: Лаврентьева Снежана Павловна, учитель химии и биологии
Яблочный спас познавательная игра по биологии, 6 класс Автор: Лаврентьева Снежана Павловна, учитель химии и биологии Обмен веществ в организме
Обмен веществ в организме  Изучение настоящих тюленей как потенциальных объектов промысла и использования в биотехнических системах
Изучение настоящих тюленей как потенциальных объектов промысла и использования в биотехнических системах Семейство кошачьих Презентация к уроку. Выполнила : учитель высшей категории МОУ «СОШ п. Новопушкинское» Энгельсского района Са
Семейство кошачьих Презентация к уроку. Выполнила : учитель высшей категории МОУ «СОШ п. Новопушкинское» Энгельсского района Са Презентация на тему "Вид- эволюционная единица, его критерии и структура" - скачать презентации по Биологии
Презентация на тему "Вид- эволюционная единица, его критерии и структура" - скачать презентации по Биологии Кто такой павлин?
Кто такой павлин? СӨЖ Көмірсулар
СӨЖ Көмірсулар Водный и минеральный обмен
Водный и минеральный обмен Основные законы наследования признаков
Основные законы наследования признаков Головной мозг и его тайны!
Головной мозг и его тайны! Функциональная анатомия мышц головы, шеи и туловища
Функциональная анатомия мышц головы, шеи и туловища Тема: Световая фаза фотосинтеза
Тема: Световая фаза фотосинтеза Сільськогосподарські рослини
Сільськогосподарські рослини