- Управление памятью (лекция 5)

Содержание
- 2. Перенос памяти - опыты Мак-Коннелла на планариях («химическая» теория памяти) Американский зоопсихолог Джеймс Мак-Коннел лизучал поведение
- 3. Перенос памяти Схема, иллюстрирующая передачу планариям-реципиентам условных рефлексов, выработанных на свет или вибрацию у планарий-доноров. Знаками
- 4. Павловское «проторение пути» («Электрическая» теория памяти)
- 5. Механизм Хебба В данной книге постулат Хебба звучит следующим образом: «When an axon of cell A
- 6. Синапсы Хебба Согласно Хеббу, следы памяти могут формироваться путем модификации синапсов. На этой схеме показано, как
- 7. Анатомия гиппокампа Сети гиппокампа: Гиппокамп образует главным образом однонаправленные сети, с входом от энторинальной коры (Entorinal
- 8. Циркуляция в гиппокампе Весь гиппокамп можно рассматривать как одну гигантскую корковую колонку. Нижний слой зернистых клеток
- 9. Гиппокампальное представительство физического пространства. Клетки места a. Участки зоны СА1 с центрами A-D, картирующими позицию крысы
- 10. Последовательное поведение клеток места воспроизводится как во время движения в прямом, так и в обратном направлении
- 11. Роль тэта-ритма Тэта-ритм - это очень точный способ задания временного хода активности клеток гиппокампа. Гиппокамп является
- 12. Прецессия фазы тэта-ритма Это выделение разных этапов обработки сигнала вдоль пути «ЭК-гиппокамп» согласуется с гипотезой, что
- 13. Рабочий цикл с переменой направления передачи Во время кодирования при бодрствовании, а также при консолидации памяти
- 14. Запоминание по механизму Хебба У грызунов гиппокампальные «клетки места» активны, когда животное проходит данное «место поля»
- 15. Вычислительная роль движущихся волн Из [Ermentrout GB, Kleinfeld D, 2001]: Волны обычно представлены во время периодов
- 16. Разности фаз тэта-волн вдоль оси гиппокампа a-d. Корональный MRI-срез по брегме - 3.8 (a), -4.3 (b),
- 17. Топография интраламинарного фазового профиля a-c. Серые кружки отмечают точки блока, где записывались LFPs в течении 45
- 18. Протез гиппокампа у крыс. Модель MIMO – «Много входов / много выходов» Два рычага (показаны зелёным
- 19. Рекуррентный интерфейс (R-BCI) A - схема передачи сигнала в рекуррентном интерфейсе (R-BCI) по Mavoori et al.,
- 20. Опыты со скотофобином, бродячие и домашние собаки; Опыты, о которых говорит К.Анохин (влияние пептидов на консолидацию
- 21. Голографическая гипотеза памяти
- 22. Оптическая голограмма Голография создана Деннисом Габором в 1947 г (Нобелевская премия по физике 1971 г)
- 23. Физический принцип голографии Рассеянные объектом волны характеризуются амплитудой и фазой. Регистрация амплитуды волн не представляет затруднений;
- 24. Дивергенция и конвергенция в нервной сети
- 25. Нейрональная голограмма
- 26. Спасибо за внимание
- 28. Скачать презентацию

Перенос памяти - опыты Мак-Коннелла на планариях
(«химическая» теория памяти)
Американский зоопсихолог Джеймс
Перенос памяти - опыты Мак-Коннелла на планариях
(«химическая» теория памяти)
Американский зоопсихолог Джеймс
Перенос памяти
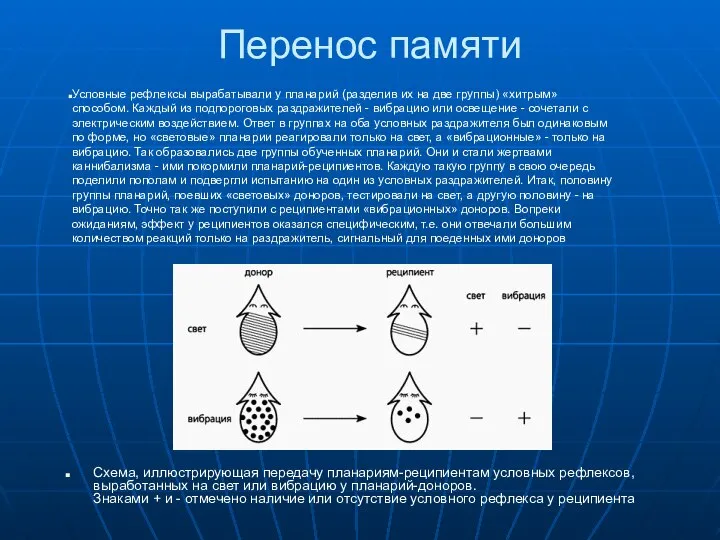
Схема, иллюстрирующая передачу планариям-реципиентам условных рефлексов,
выработанных на свет или
Перенос памяти
Схема, иллюстрирующая передачу планариям-реципиентам условных рефлексов, выработанных на свет или
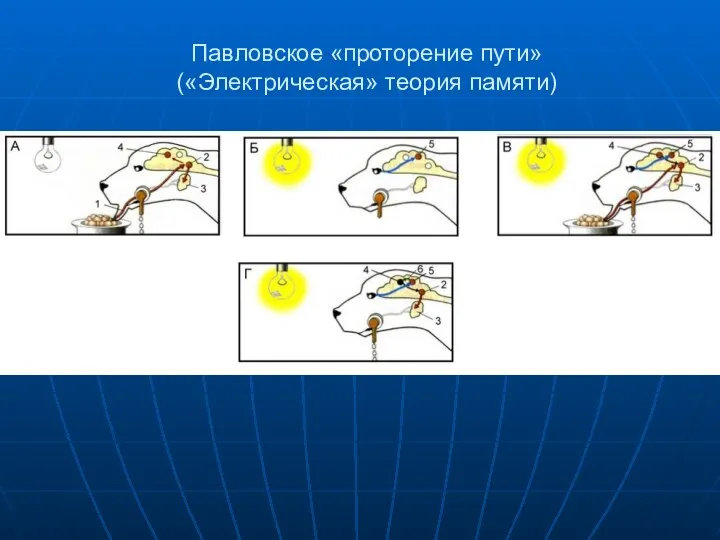
Павловское «проторение пути»
(«Электрическая» теория памяти)
Павловское «проторение пути»
(«Электрическая» теория памяти)

Механизм Хебба
В данной книге постулат Хебба звучит следующим образом: «When an
Механизм Хебба
В данной книге постулат Хебба звучит следующим образом: «When an

Синапсы Хебба
Согласно Хеббу, следы памяти могут формироваться путем модификации синапсов. На
Синапсы Хебба
Согласно Хеббу, следы памяти могут формироваться путем модификации синапсов. На
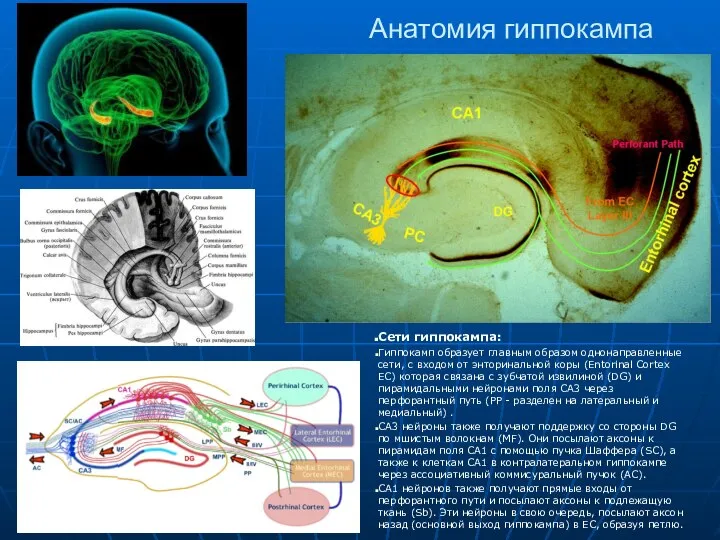
Анатомия гиппокампа
Сети гиппокампа:
Гиппокамп образует главным образом однонаправленные сети, с входом
Анатомия гиппокампа
Сети гиппокампа:
Гиппокамп образует главным образом однонаправленные сети, с входом
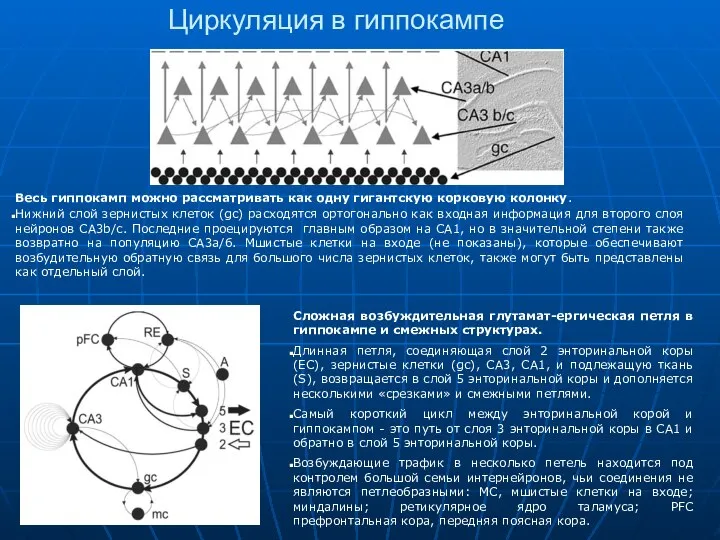
Циркуляция в гиппокампе
Весь гиппокамп можно рассматривать как одну гигантскую корковую колонку.
Циркуляция в гиппокампе
Весь гиппокамп можно рассматривать как одну гигантскую корковую колонку.

Гиппокампальное представительство физического пространства. Клетки места
a. Участки зоны СА1
Гиппокампальное представительство физического пространства. Клетки места
a. Участки зоны СА1
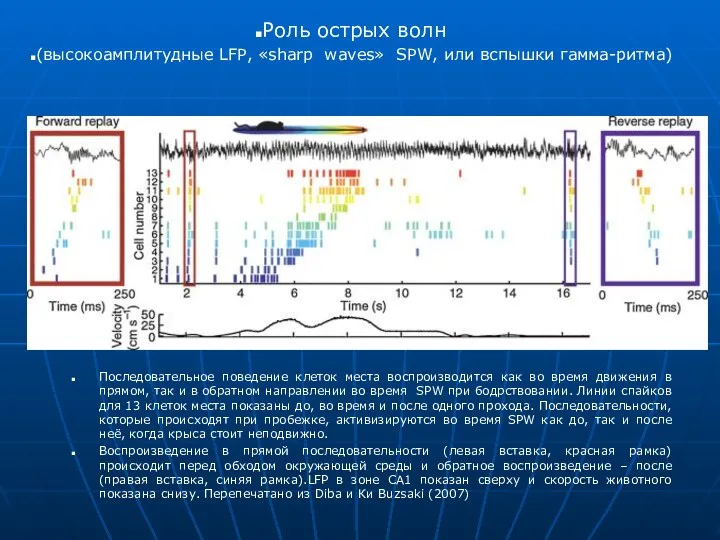
Последовательное поведение клеток места воспроизводится как во время движения в прямом,
Последовательное поведение клеток места воспроизводится как во время движения в прямом,

Роль тэта-ритма
Тэта-ритм - это очень точный способ задания временного хода активности
Роль тэта-ритма
Тэта-ритм - это очень точный способ задания временного хода активности

Прецессия фазы тэта-ритма
Это выделение разных этапов обработки сигнала вдоль пути «ЭК-гиппокамп»
Прецессия фазы тэта-ритма
Это выделение разных этапов обработки сигнала вдоль пути «ЭК-гиппокамп»
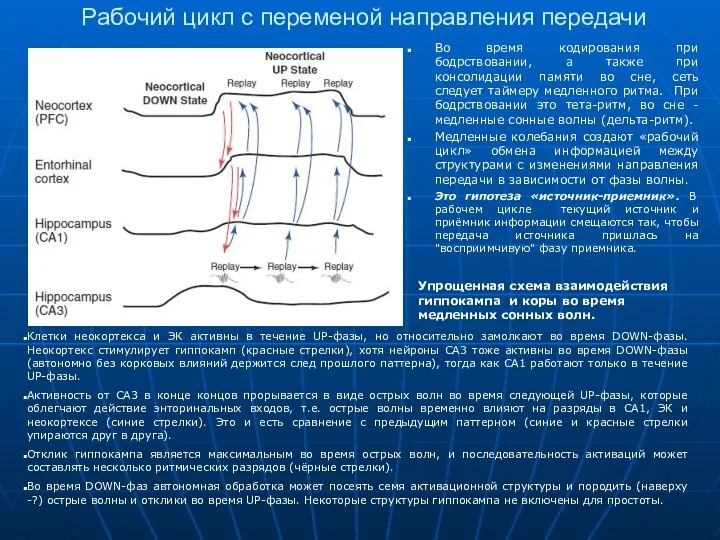
Рабочий цикл с переменой направления передачи
Во время кодирования при бодрствовании, а
Рабочий цикл с переменой направления передачи
Во время кодирования при бодрствовании, а

Запоминание по механизму Хебба
У грызунов гиппокампальные «клетки места» активны, когда животное
Запоминание по механизму Хебба
У грызунов гиппокампальные «клетки места» активны, когда животное
![Вычислительная роль движущихся волн Из [Ermentrout GB, Kleinfeld D, 2001]: Волны](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/667388/slide-14.jpg)
Вычислительная роль движущихся волн
Из [Ermentrout GB, Kleinfeld D, 2001]: Волны обычно
Вычислительная роль движущихся волн
Из [Ermentrout GB, Kleinfeld D, 2001]: Волны обычно
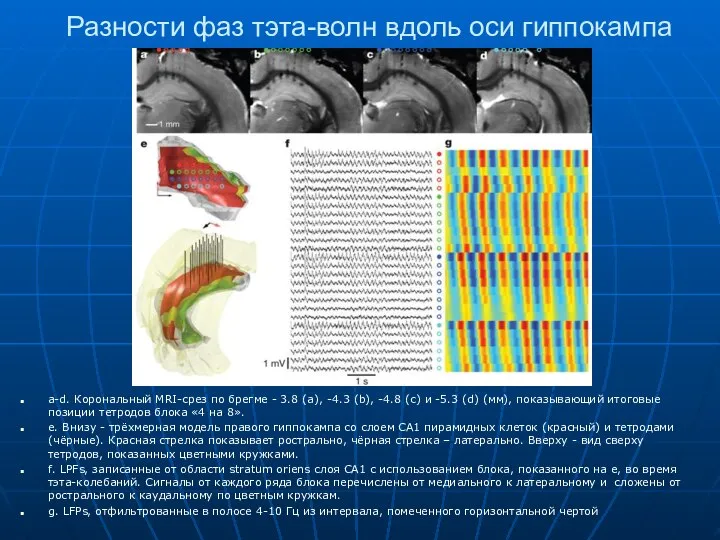
Разности фаз тэта-волн вдоль оси гиппокампа
a-d. Корональный MRI-срез по брегме -
Разности фаз тэта-волн вдоль оси гиппокампа
a-d. Корональный MRI-срез по брегме -
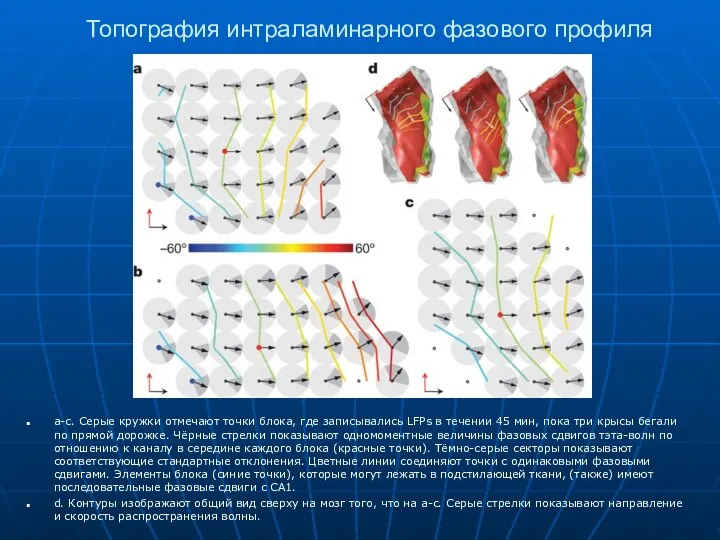
Топография интраламинарного фазового профиля
a-c. Серые кружки отмечают точки блока, где записывались
Топография интраламинарного фазового профиля
a-c. Серые кружки отмечают точки блока, где записывались
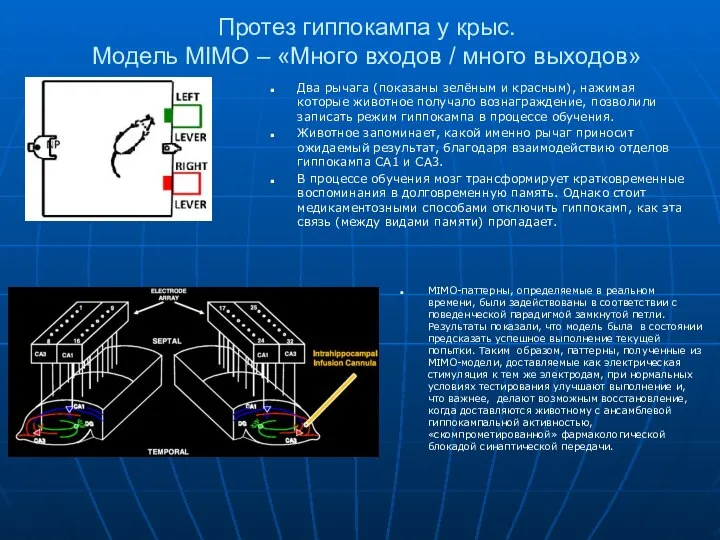
Протез гиппокампа у крыс.
Модель MIMO – «Много входов / много
Протез гиппокампа у крыс. Модель MIMO – «Много входов / много
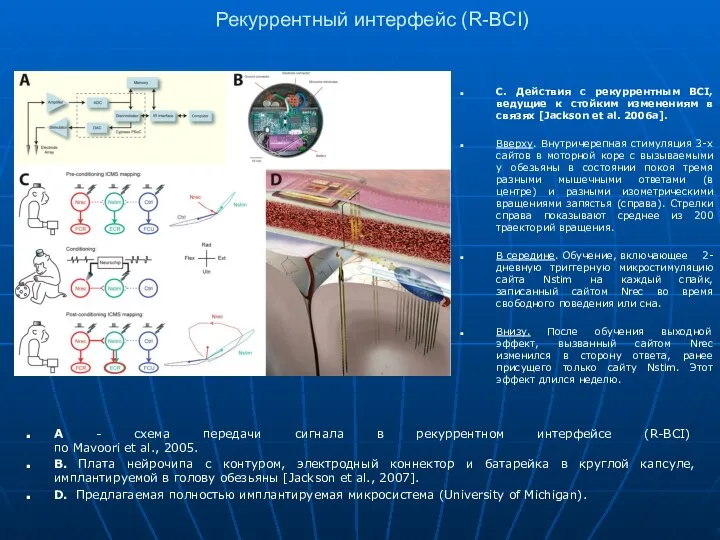
Рекуррентный интерфейс (R-BCI)
A - схема передачи сигнала в рекуррентном интерфейсе
Рекуррентный интерфейс (R-BCI)
A - схема передачи сигнала в рекуррентном интерфейсе

Опыты со скотофобином, бродячие и домашние собаки;
Опыты, о которых говорит К.Анохин
Опыты со скотофобином, бродячие и домашние собаки;
Опыты, о которых говорит К.Анохин

Голографическая гипотеза памяти
Голографическая гипотеза памяти
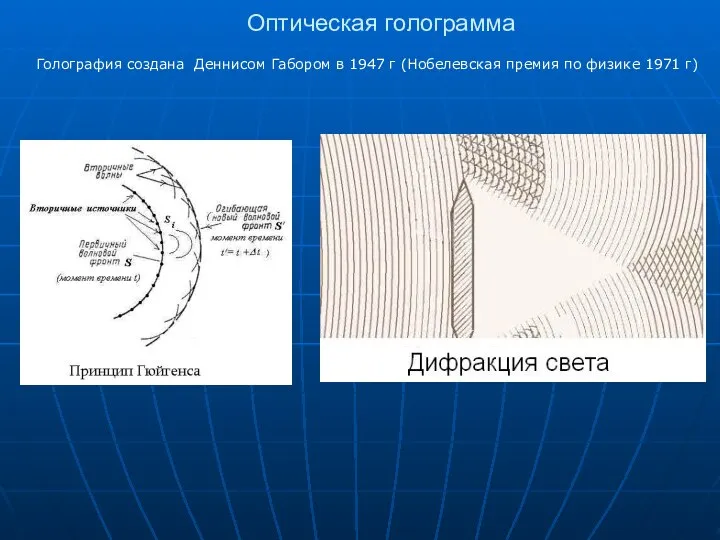
Оптическая голограмма
Голография создана Деннисом Габором в 1947 г (Нобелевская премия по
Оптическая голограмма
Голография создана Деннисом Габором в 1947 г (Нобелевская премия по

Физический принцип голографии
Рассеянные объектом волны характеризуются амплитудой и фазой. Регистрация амплитуды
Физический принцип голографии
Рассеянные объектом волны характеризуются амплитудой и фазой. Регистрация амплитуды
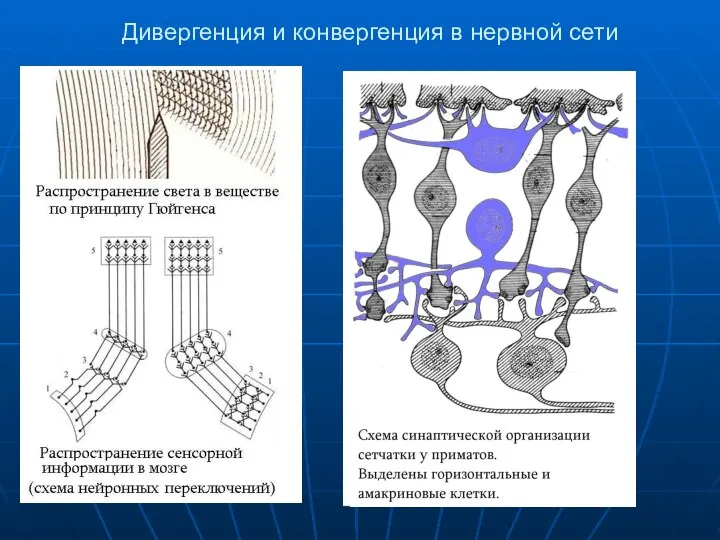
Дивергенция и конвергенция в нервной сети
Дивергенция и конвергенция в нервной сети

Нейрональная голограмма
Нейрональная голограмма

 Кроветворение и его регуляция. Эритропоэтины и лейкопоэтины
Кроветворение и его регуляция. Эритропоэтины и лейкопоэтины Врожденные аномалии развития зрительного нерва
Врожденные аномалии развития зрительного нерва Хронический гнойный средний отит
Хронический гнойный средний отит Гимнастика для глаз
Гимнастика для глаз Чувствительность и ее нарушения
Чувствительность и ее нарушения Сущность метода электрофореза
Сущность метода электрофореза 3D-печать в протезировании. Био-электрический протез кисти
3D-печать в протезировании. Био-электрический протез кисти Лечение синдромов инфекционных заболеваний. Инфекционная лихорадка у детей
Лечение синдромов инфекционных заболеваний. Инфекционная лихорадка у детей Генетика негіздері
Генетика негіздері Родовая и преимущественная стратегии
Родовая и преимущественная стратегии Курс косметолог-эстетист. Модуль 2.4 возрастные изменения кожи
Курс косметолог-эстетист. Модуль 2.4 возрастные изменения кожи Антенатальная охрана плода
Антенатальная охрана плода Неотложные состояния в офтальмологии. (клиника и современная терапия)
Неотложные состояния в офтальмологии. (клиника и современная терапия) Миокардит. Клинические признаки
Миокардит. Клинические признаки Теории личности в отечественной психологии. Современные подходы к личности. Тысячеликий герой Дж. Кемпбелла
Теории личности в отечественной психологии. Современные подходы к личности. Тысячеликий герой Дж. Кемпбелла О медработниках Пижмы
О медработниках Пижмы Тромбозы глубоких вен в системе нижней полой вены
Тромбозы глубоких вен в системе нижней полой вены Аурухана ішілік инфекцияны бақылауды ұйымдастыру
Аурухана ішілік инфекцияны бақылауды ұйымдастыру Клещевой энцефалит в Омской области
Клещевой энцефалит в Омской области ЖИТС-пен ауыратын науқастардағы комплаенс
ЖИТС-пен ауыратын науқастардағы комплаенс Стенотическое поражение сонных артерий
Стенотическое поражение сонных артерий Инфекционные поражения СОПР. Острый герпетический стоматит у детей
Инфекционные поражения СОПР. Острый герпетический стоматит у детей Дәрілік заттар әсерінің организмнің қасиеттеріне тәуелділігі
Дәрілік заттар әсерінің организмнің қасиеттеріне тәуелділігі ДНК – ның теломерлік бөлімдерінің репликациялануы
ДНК – ның теломерлік бөлімдерінің репликациялануы Рациональная фармакотерапия анемии беременных
Рациональная фармакотерапия анемии беременных Жергілікті сәулелік зақымдалулар
Жергілікті сәулелік зақымдалулар Вигодовування дітей раннього віку
Вигодовування дітей раннього віку Диабеттік ретинопатия
Диабеттік ретинопатия