- Recettori

Содержание
- 2. La segnalazione intercellulare L’evoluzione degli organismi multicellulari dipende dalla capacità delle cellule di comunicare una con
- 3. La segnalazione intercellulare Molte cellule secernono una o più molecole segnale che funzionano come mediatori chimici
- 4. MEDIATORI ENDOGENI Mediatori a basso peso molecolare: Aminoacidi: glutammato, glicina, acido gamma-amino-butirrico (GABA) Monoamine: catecolamine (dopamina,
- 5. Recettori Classificazione in base alla localizzazione: Recettori di membrana Recettori intracellulari I primi rispondono a mediatori
- 6. Recettori Classificazione in base alla localizzazione: Recettori di membrana Recettori intracellulari I secondi trasducono il segnale
- 7. Recettori intracellulari Divisi in due classi: I) citoplasmatici, formano omodimeri o dimeri con il ligando. Ne
- 8. Quattro (super)famiglie di recettori Recettori di membrana Recettori intracellulari
- 9. Aspetti quantitativi delle risposte ai farmaci: analisi delle curve La relazione fra CONCENTRAZIONE di un FARMACO
- 10. In molti modelli sperimentali la risposta che si misura è graduabile e direttamente correlabile con la
- 11. Riportando sulle ascisse le dosi impiegate e sulle ordinate le risposte misurate, l’andamento della curva dose-risposta
- 12. Si preferisce riportare le dosi in scala logaritmica , in questo modo la curva assume un
- 13. Esempi: letalità, convulsioni, sviluppo di un tumore, effetto ipnotico Ogni dose è somministrata ad un gruppo
- 14. Curve dose-risposta in una popolazione es. effetto letale di dosi crescenti di alcool etilico ISTOGRAMMA DELLE
- 15. Principali teorie recettoriali (1) Teoria occupazionale (Clark, 1933) I farmaci interagiscono con siti di binding indipendenti
- 16. Per la teoria di Clark KA = EC50 la costante di dissociazione di un agonista KA
- 17. Il contributo di Ariens (1954) Non tutti gli agonisti producono la stessa risposta massimale Principali teorie
- 18. Attività intrinseca E’ la capacità del farmaco di indurre una risposta biologica (α) La risposta biologica
- 19. gli agonisti si legano ai recettori producendo effetti di vario tipo gli antagonisti si legano ai
- 20. Farmaci agonisti Generalmente un agonista riproduce gli effetti dei ligandi endogeni. agonista pieno (α = 1)
- 21. Curve di farmaci agonisti con potenza diversa ed uguale efficacia Agonista, log M = efficacia potenza
- 22. Curve dose risposta per un agonista pieno e per agonisti parziali
- 23. Farmaci antagonisti Si legano con alta affinità ai recettori, ma non sono in grado di indurre
- 24. Antagonisti competitivi Agonista, log M ?Emax potenza apparente
- 25. Antagonisti competitivi Il blocco del recettore da parte dell’antagonista può essere superato aumentando la concentrazione dell’agonista
- 26. Antagonisti non competitivi ?Emax ≠ Emax IC50 Agonista, log M
- 27. Antagonisti non competitivi o irreversibili Un antagonista non competitivo sposta la curva dell’agonista verso destra in
- 28. Il contributo di Stephenson (1954) La risposta farmacologica non è linearmente proporzionale al numero di recettori
- 29. Il contributo di Furchgott (1966) Fornì l’evidenza sperimentale dell’esistenza degli spare receptors Principali teorie recettoriali (4)
- 30. Caratteristiche dell’interazione di un farmaco con il suo recettore AFFINITA’: esprime la forza con cui il
- 31. ………riassumendo
- 32. La tecnica del binding recettoriale Metodi semplici e rapidi per lo screening farmacologico di numerose molecole.
- 33. Si procede incubando un omogenato di tessuto opportunamente diluito e di cui si dovrà determinare la
- 34. Preparazione del tessuto Semplice omogenato (es. di cervello) Frazione subcellulare arricchita della componente rappresentata dalle membrane
- 35. I ligandi marcati sono disponibili commercialmente
- 36. Durante l’incubazione si forma il complesso ligando-recettore, che segue la legge d’azione di massa. Tale reazione
- 37. Si termina l’incubazione con una procedura che permetta di separare il ligando legato al recettore dal
- 38. Al termine dell’esperimento, la radioattività legata ai filtri o presente nel pellet viene quantificata mediante spettrofotometria
- 39. Binding misurato per concentrazioni crescenti di legante radioattivo fino alla saturazione dei siti di binding. Es.
- 40. membranes + hot ligand membranes + cold ligand + hot ligand Total bound Non specific bound
- 41. F = free B = bound Bmax = total receptor In ascisse la concentrazione di ligando
- 42. La concentrazione del legante radioattivo viene mantenuta fissa mentre sono i farmaci ad essere testati a
- 43. In questi esperimenti, il binding di un legante marcato è misurato in presenza di concentrazioni crescenti
- 44. Saturabilità – il numero di recettori è finito Reversibilità – il legame ligando-recettore non è covalente
- 45. Recettori accoppiati a proteine G Definiti anche come recettori metabotropici. Costituti da unico filamento che attraversa
- 46. Le proteine G Le proteine G sono eterotrimeri (composti da subunita α, β e γ). In
- 47. Il sistema dell’adenilato ciclasi L’AMP ciclico attiva la proteina chinasi A
- 48. Il sistema della fosfolipasi C La fosfolipasi C scinde il PIP3 a DAG ed inositolo tri
- 51. Скачать презентацию

La segnalazione intercellulare
L’evoluzione degli organismi multicellulari dipende dalla capacità delle cellule
La segnalazione intercellulare
L’evoluzione degli organismi multicellulari dipende dalla capacità delle cellule

La segnalazione intercellulare
Molte cellule secernono
una o più molecole
segnale che funzionano
come
La segnalazione intercellulare
Molte cellule secernono
una o più molecole
segnale che funzionano
come

MEDIATORI ENDOGENI
Mediatori a basso peso molecolare:
Aminoacidi: glutammato, glicina, acido gamma-amino-butirrico (GABA)
Monoamine:
MEDIATORI ENDOGENI
Mediatori a basso peso molecolare:
Aminoacidi: glutammato, glicina, acido gamma-amino-butirrico (GABA)
Monoamine:
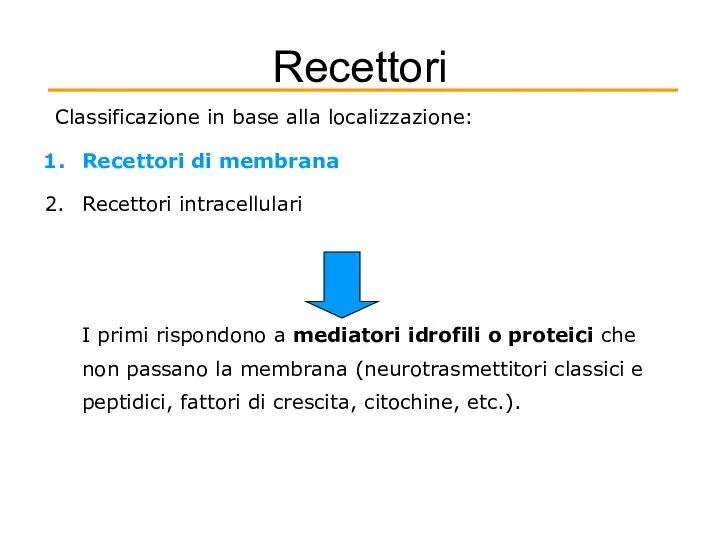
Recettori
Classificazione in base alla localizzazione:
Recettori di membrana
Recettori intracellulari
I primi rispondono a
Recettori
Classificazione in base alla localizzazione:
Recettori di membrana
Recettori intracellulari
I primi rispondono a

Recettori
Classificazione in base alla localizzazione:
Recettori di membrana
Recettori intracellulari
I secondi trasducono il
Recettori
Classificazione in base alla localizzazione:
Recettori di membrana
Recettori intracellulari
I secondi trasducono il

Recettori intracellulari
Divisi in due classi:
I) citoplasmatici, formano omodimeri o dimeri con
Recettori intracellulari
Divisi in due classi:
I) citoplasmatici, formano omodimeri o dimeri con
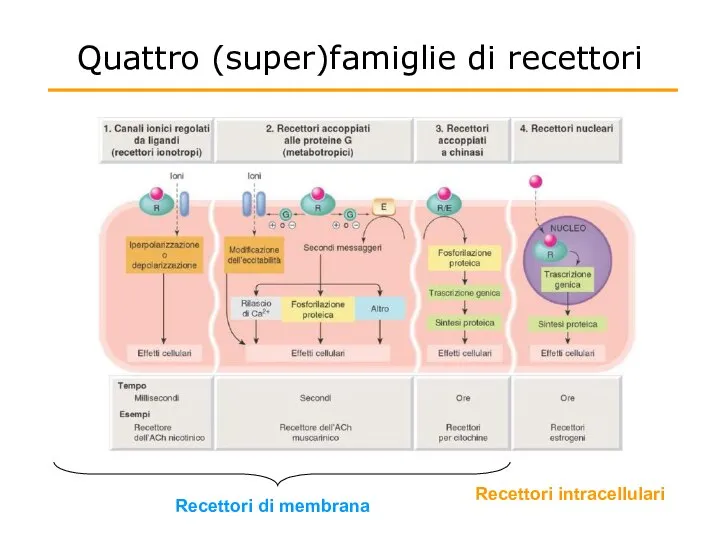
Quattro (super)famiglie di recettori
Recettori di membrana
Recettori intracellulari
Quattro (super)famiglie di recettori
Recettori di membrana
Recettori intracellulari

Aspetti quantitativi delle risposte ai farmaci:
analisi delle curve
La relazione
Aspetti quantitativi delle risposte ai farmaci:
analisi delle curve
La relazione

In molti modelli sperimentali la risposta che si misura è graduabile
In molti modelli sperimentali la risposta che si misura è graduabile
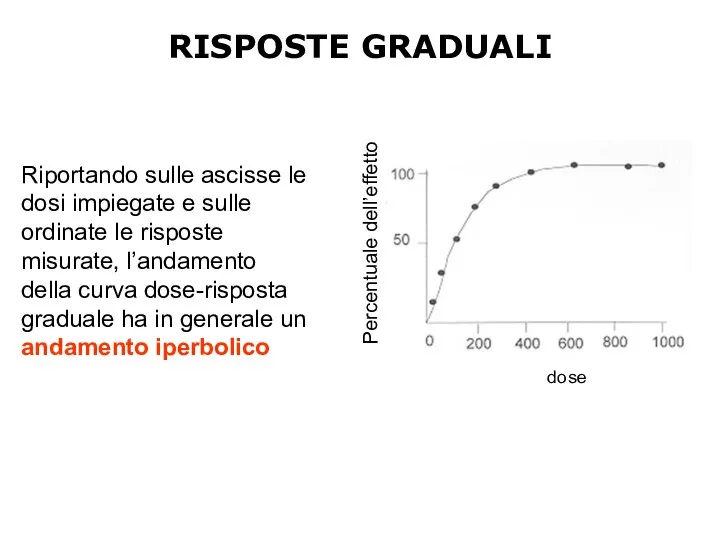
Riportando sulle ascisse le dosi impiegate e sulle ordinate le risposte
Riportando sulle ascisse le dosi impiegate e sulle ordinate le risposte
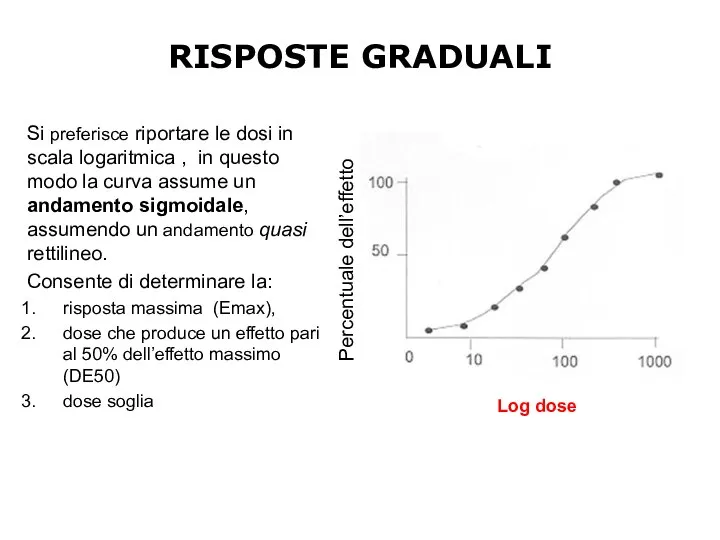
Si preferisce riportare le dosi in scala logaritmica , in questo
Si preferisce riportare le dosi in scala logaritmica , in questo
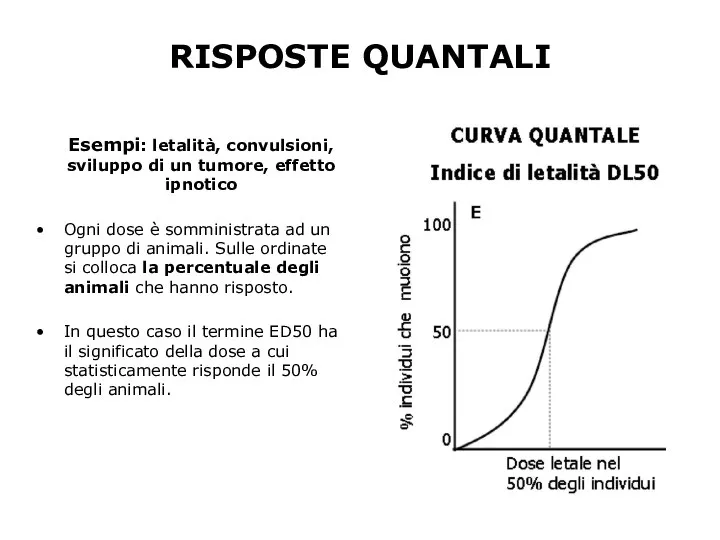
Esempi: letalità, convulsioni, sviluppo di un tumore, effetto ipnotico
Ogni dose
Esempi: letalità, convulsioni, sviluppo di un tumore, effetto ipnotico
Ogni dose
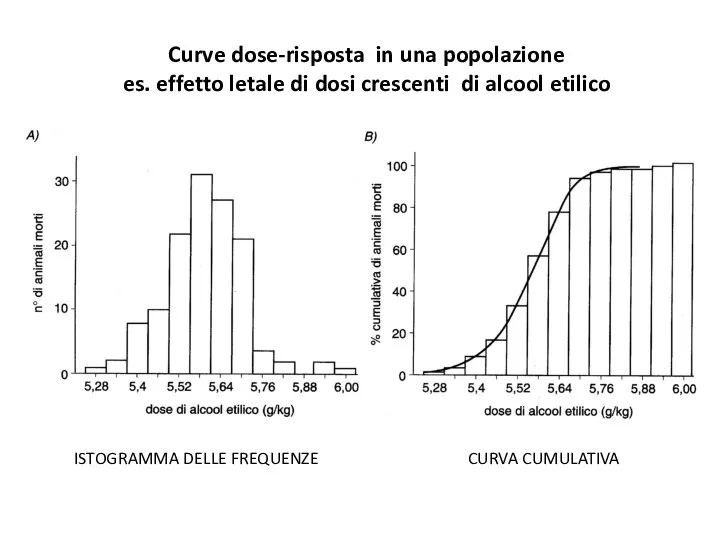
Curve dose-risposta in una popolazione
es. effetto letale di dosi crescenti di
Curve dose-risposta in una popolazione
es. effetto letale di dosi crescenti di
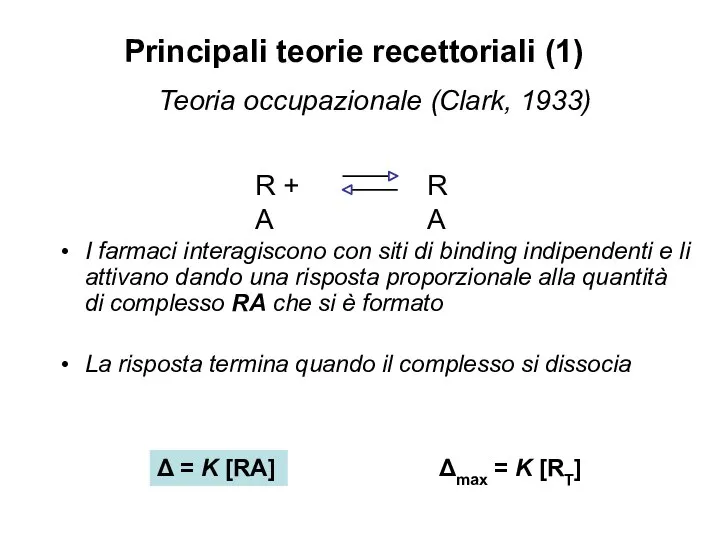
Principali teorie recettoriali (1)
Teoria occupazionale (Clark, 1933)
I farmaci interagiscono con
Principali teorie recettoriali (1)
Teoria occupazionale (Clark, 1933)
I farmaci interagiscono con
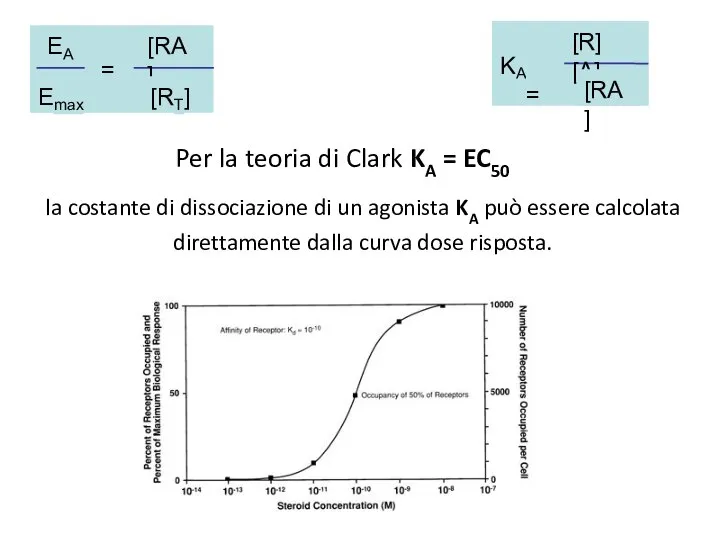
Per la teoria di Clark KA = EC50
la costante di dissociazione
Per la teoria di Clark KA = EC50
la costante di dissociazione

Il contributo di Ariens (1954)
Non tutti gli agonisti producono la stessa
Il contributo di Ariens (1954)
Non tutti gli agonisti producono la stessa

Attività intrinseca
E’ la capacità del farmaco di indurre
una risposta biologica
Attività intrinseca
E’ la capacità del farmaco di indurre
una risposta biologica

gli agonisti si legano ai recettori producendo effetti di vario tipo
gli agonisti si legano ai recettori producendo effetti di vario tipo

Farmaci agonisti
Generalmente un agonista riproduce gli effetti dei ligandi endogeni.
agonista
Farmaci agonisti
Generalmente un agonista riproduce gli effetti dei ligandi endogeni.
agonista
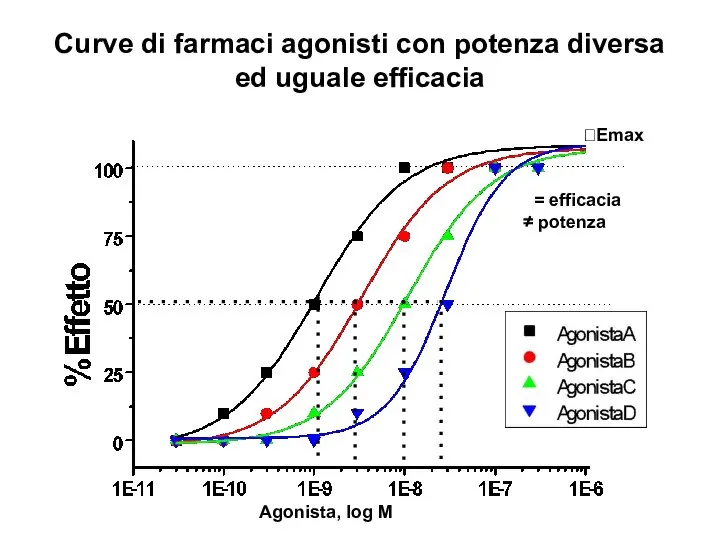
Curve di farmaci agonisti con potenza diversa ed uguale efficacia
Agonista, log
Curve di farmaci agonisti con potenza diversa ed uguale efficacia
Agonista, log
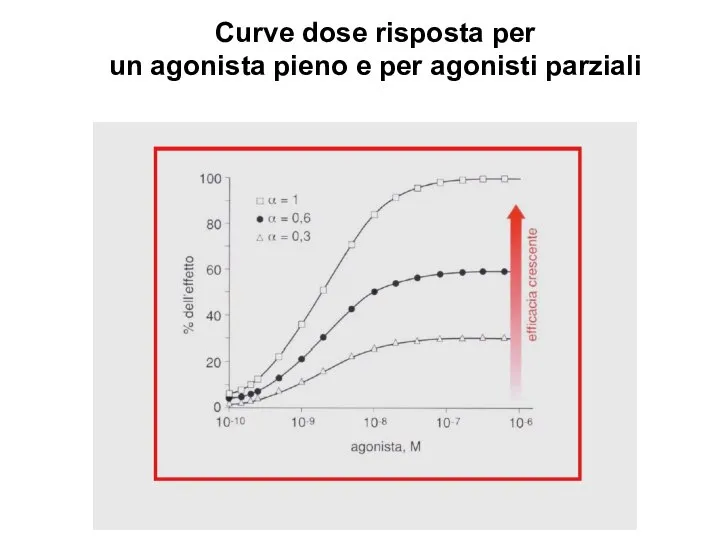
Curve dose risposta per
un agonista pieno e per agonisti parziali
Curve dose risposta per
un agonista pieno e per agonisti parziali

Farmaci antagonisti
Si legano con alta affinità ai recettori, ma non sono
Farmaci antagonisti
Si legano con alta affinità ai recettori, ma non sono
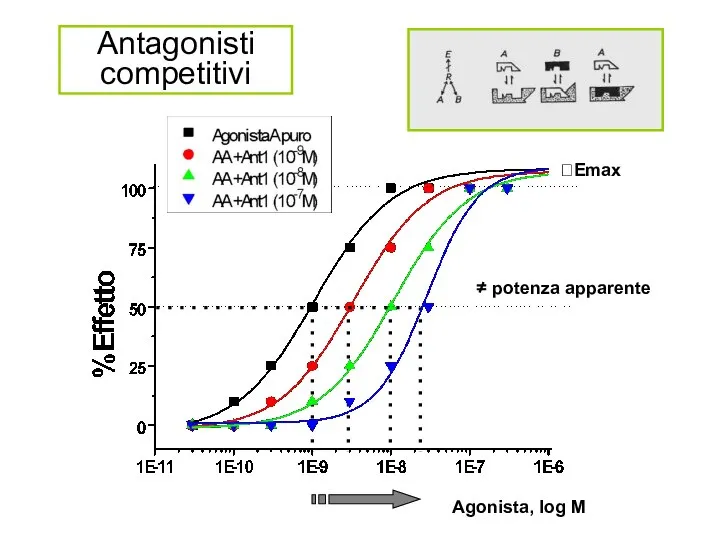
Antagonisti competitivi
Agonista, log M
?Emax
potenza apparente
Antagonisti competitivi
Agonista, log M
?Emax
potenza apparente
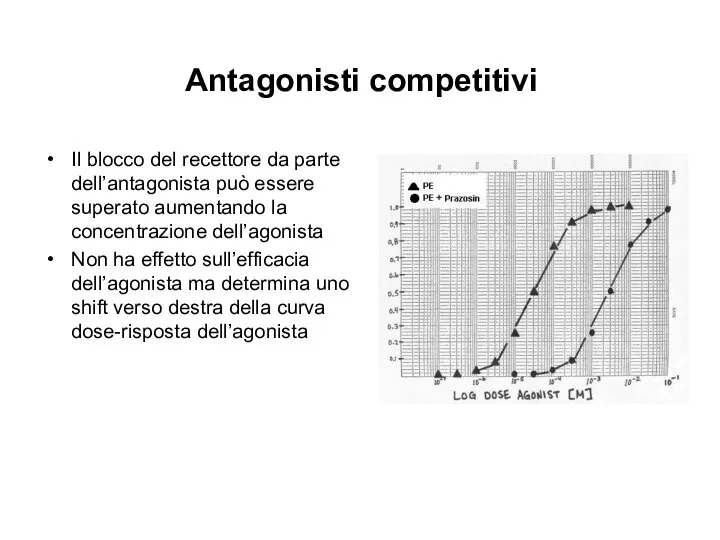
Antagonisti competitivi
Il blocco del recettore da parte dell’antagonista può essere superato
Antagonisti competitivi
Il blocco del recettore da parte dell’antagonista può essere superato
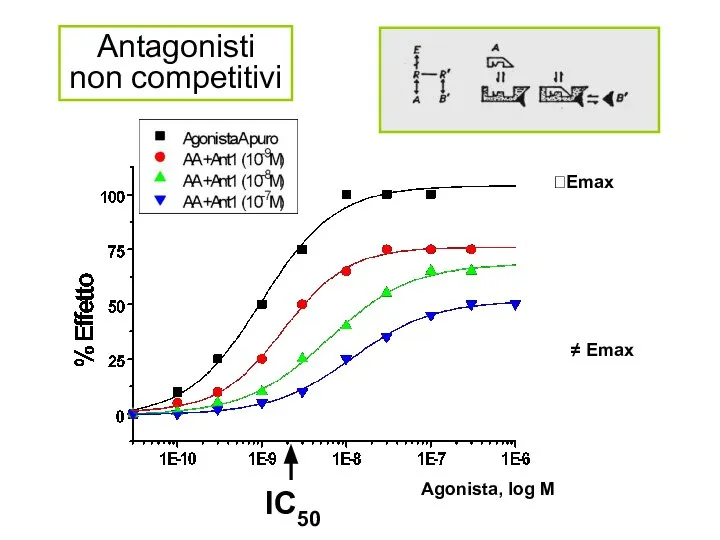
Antagonisti non competitivi
?Emax
≠ Emax
IC50
Agonista, log M
Antagonisti non competitivi
?Emax
≠ Emax
IC50
Agonista, log M

Antagonisti non competitivi o irreversibili
Un antagonista non competitivo sposta la curva
Antagonisti non competitivi o irreversibili
Un antagonista non competitivo sposta la curva

Il contributo di Stephenson (1954)
La risposta farmacologica non è linearmente proporzionale
Il contributo di Stephenson (1954)
La risposta farmacologica non è linearmente proporzionale

Il contributo di Furchgott (1966)
Fornì l’evidenza sperimentale dell’esistenza degli spare receptors
Principali
Il contributo di Furchgott (1966)
Fornì l’evidenza sperimentale dell’esistenza degli spare receptors
Principali

Caratteristiche dell’interazione
di un farmaco con il suo recettore
AFFINITA’: esprime la
Caratteristiche dell’interazione
di un farmaco con il suo recettore
AFFINITA’: esprime la
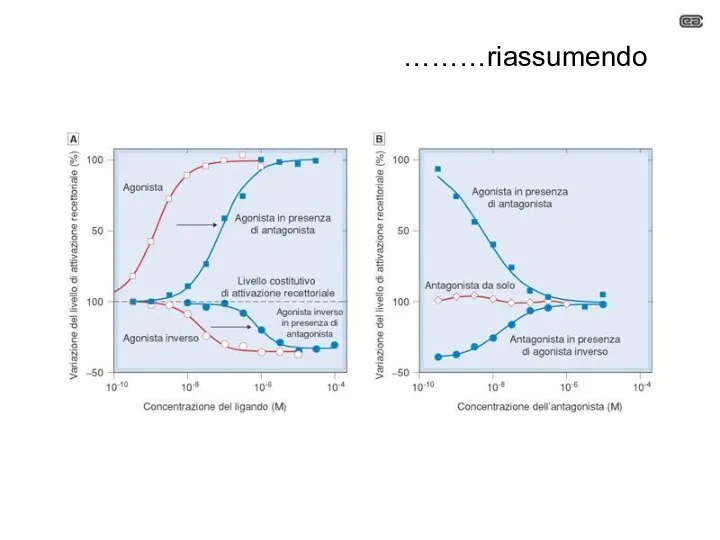
………riassumendo
………riassumendo

La tecnica del binding recettoriale
Metodi semplici e rapidi per lo screening
La tecnica del binding recettoriale
Metodi semplici e rapidi per lo screening

Si procede incubando un omogenato di tessuto opportunamente diluito e di
Si procede incubando un omogenato di tessuto opportunamente diluito e di

Preparazione del tessuto
Semplice omogenato (es. di cervello)
Frazione subcellulare arricchita della componente
Preparazione del tessuto
Semplice omogenato (es. di cervello)
Frazione subcellulare arricchita della componente
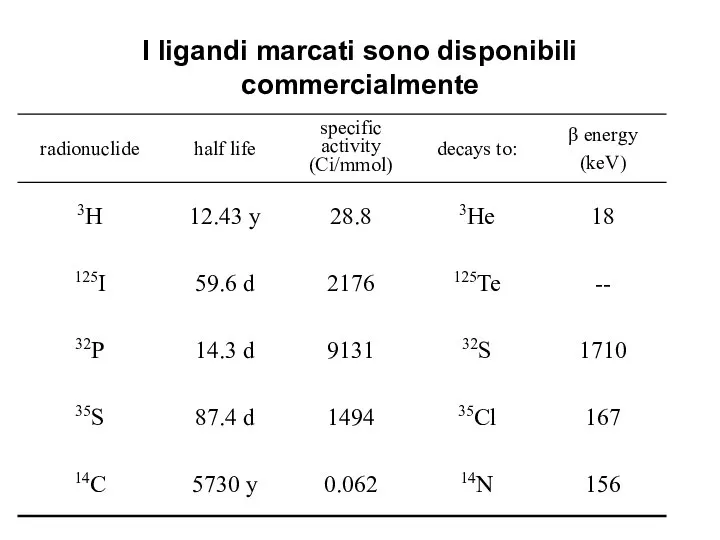
I ligandi marcati sono disponibili commercialmente
I ligandi marcati sono disponibili commercialmente

Durante l’incubazione si forma il complesso ligando-recettore, che segue la legge
Durante l’incubazione si forma il complesso ligando-recettore, che segue la legge

Si termina l’incubazione con una procedura che permetta di separare il
Si termina l’incubazione con una procedura che permetta di separare il

Al termine dell’esperimento, la radioattività legata ai filtri o presente nel
Al termine dell’esperimento, la radioattività legata ai filtri o presente nel

Binding misurato per concentrazioni crescenti di legante radioattivo fino alla saturazione
Binding misurato per concentrazioni crescenti di legante radioattivo fino alla saturazione
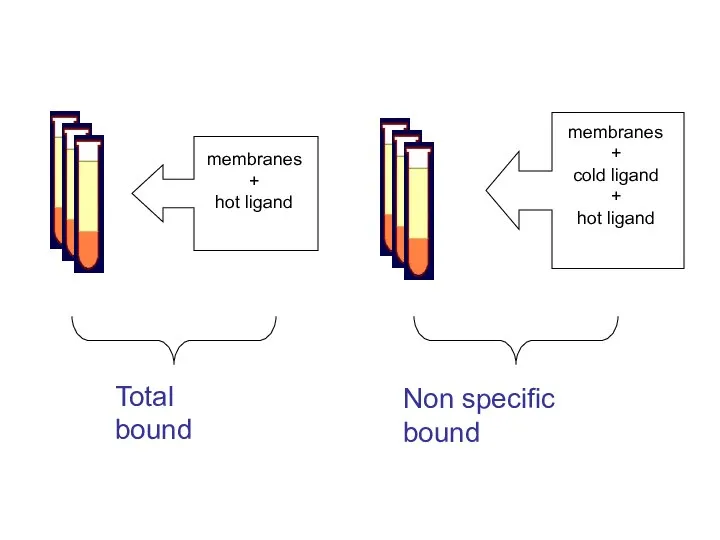
membranes
+
hot ligand
membranes
+
cold ligand
+
hot ligand
Total bound
Non specific bound
membranes
+
hot ligand
membranes
+
cold ligand
+
hot ligand
Total bound
Non specific bound
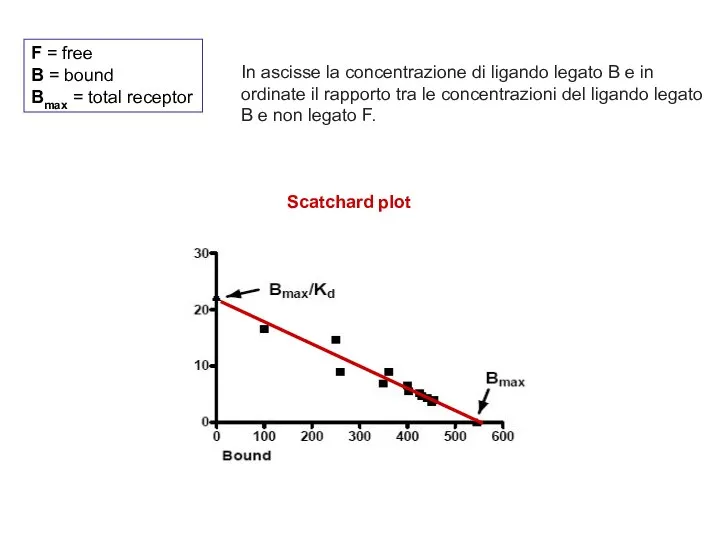
F = free
B = bound
Bmax = total receptor
In ascisse la concentrazione
F = free
B = bound
Bmax = total receptor
In ascisse la concentrazione
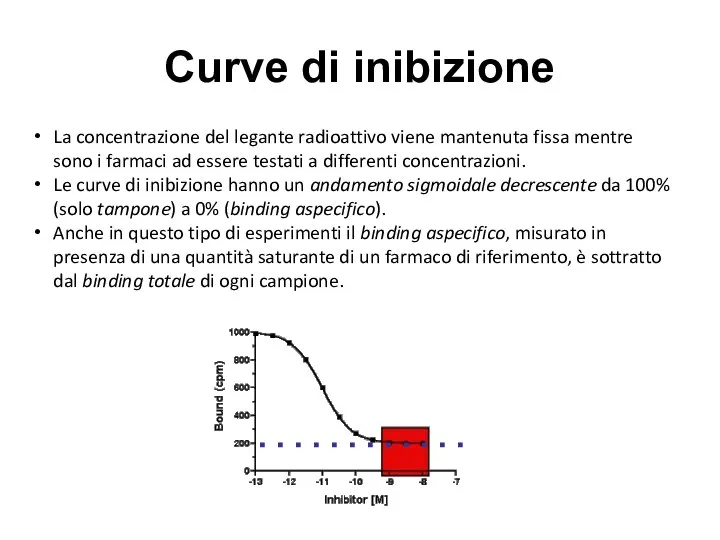
La concentrazione del legante radioattivo viene mantenuta fissa mentre sono i

In questi esperimenti, il binding di un legante marcato è misurato

Saturabilità – il numero di recettori è finito
Reversibilità – il
Saturabilità – il numero di recettori è finito
Reversibilità – il

Recettori accoppiati a proteine G
Definiti anche come recettori metabotropici.
Costituti da
Recettori accoppiati a proteine G
Definiti anche come recettori metabotropici.
Costituti da
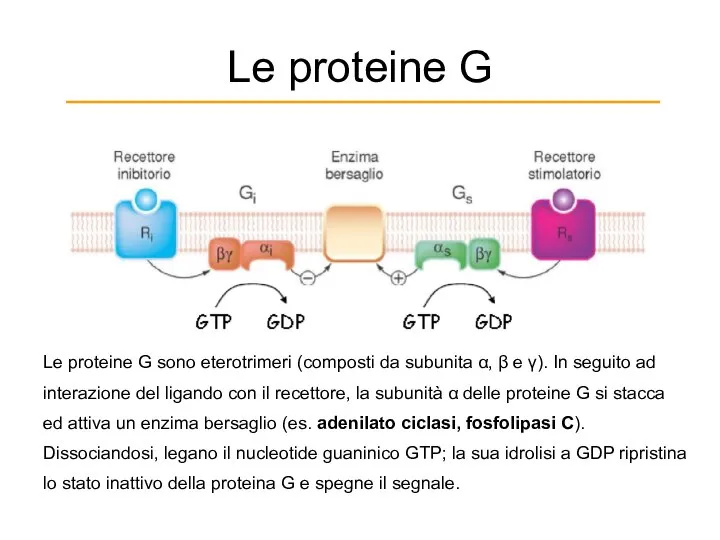
Le proteine G
Le proteine G sono eterotrimeri (composti da subunita α,
Le proteine G
Le proteine G sono eterotrimeri (composti da subunita α,
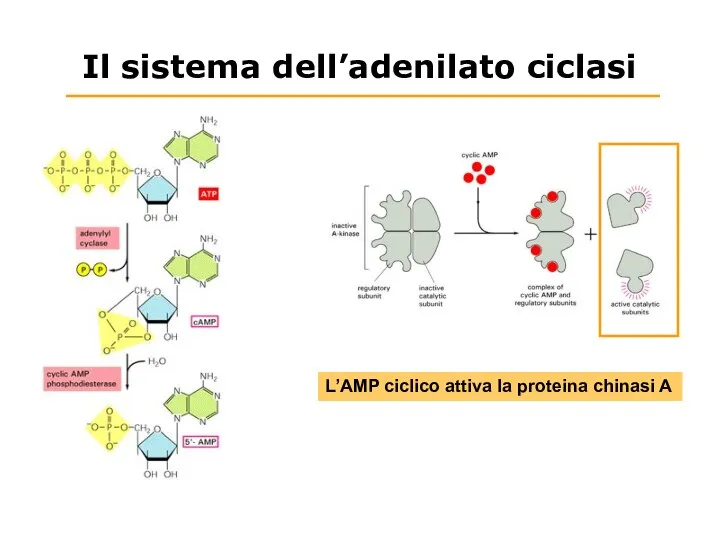
Il sistema dell’adenilato ciclasi
L’AMP ciclico attiva la proteina chinasi A
Il sistema dell’adenilato ciclasi
L’AMP ciclico attiva la proteina chinasi A

Il sistema della fosfolipasi C
La fosfolipasi C scinde il PIP3 a
Il sistema della fosfolipasi C
La fosfolipasi C scinde il PIP3 a

Похожие презентации
 Trouble Shooting Guide <Hardware Issues>
Trouble Shooting Guide <Hardware Issues> Ржев – город воинской славы
Ржев – город воинской славы День педагога. Зі святом. дорогі колеги
День педагога. Зі святом. дорогі колеги
 Wigilia bożego narodzeniaWigilia Bożego Narodzenia
Wigilia bożego narodzeniaWigilia Bożego Narodzenia 20151117_usloviya_prorastaniya_semyan
20151117_usloviya_prorastaniya_semyan Своя игра. Веков минувших достояние…
Своя игра. Веков минувших достояние… Foundmate помоги себе и другим
Foundmate помоги себе и другим Изготовление декоративной вазы для карандашей и кисточек (из подручных материалов)
Изготовление декоративной вазы для карандашей и кисточек (из подручных материалов) Конструкции гражданских зданий
Конструкции гражданских зданий Неорганические вяжущие вещества. Лекция №8
Неорганические вяжущие вещества. Лекция №8 бизнес план
бизнес план Індуїзм. Храм
Індуїзм. Храм Схема Медведевского ДК
Схема Медведевского ДК Going shopping
Going shopping Тесты Фармакология спорта
Тесты Фармакология спорта Определение содержания ионов меди и железа в различных фракциях крови в норме и при патологии(ССЗ)
Определение содержания ионов меди и железа в различных фракциях крови в норме и при патологии(ССЗ) Подготовка инструмента к работе
Подготовка инструмента к работе Рисунок, геометрические фигуры
Рисунок, геометрические фигуры Lead the way!
Lead the way! 20170211_prilozhenie_no2
20170211_prilozhenie_no2 Закрепление изученного. Что узнали. Чему научились
Закрепление изученного. Что узнали. Чему научились Продукция НЛМК
Продукция НЛМК Детский совет в ДОУ по ФГОС
Детский совет в ДОУ по ФГОС листание 8
листание 8 c794b18c6edbeb069a3764451eb9c0a0
c794b18c6edbeb069a3764451eb9c0a0 Шаблон проекта. Территория 2020
Шаблон проекта. Территория 2020 Проект размера рубок ухода в сосновых лесах ГЛХУ Щучинский лесхоз с использованием ГИС-технологий
Проект размера рубок ухода в сосновых лесах ГЛХУ Щучинский лесхоз с использованием ГИС-технологий