- Селекция1

Содержание
- 2. Схема Гальтона Исторически статистический анализ биологических данных по наследуемости количественных признаков начался с работ Френсиса Гальтона
- 3. Одномерная линейная регрессия Ф.Гальтон
- 4. Одномерная линейная регрессия Ф.Гальтон
- 5. Одномерная линейная регрессия Уравнение линейной регресии. Метод наименьших квадратов
- 6. Наследственность -- свойство организмов передавать при размножении свои признаки и особенности развития своему потомству. Наследование --
- 7. Коэффициент наследуемости (гетерогенные родители и потомки) Lush. I. L. 1937. Animal Breeding Plans. Collegiate Press, Inc.,
- 8. Нас интересует прежде всего аддитивный эффект. Именно он отвечает за успешность отбора. Коэффициент наследуемости (гетерогенные родители
- 9. Коэффициент наследуемости (гетерогенные родители и потомки) Lush. I. L. 1937. Animal Breeding Plans. Collegiate Press, Inc.,
- 10. Коэффициент наследуемости (гетерогенные родители и потомки) в широком смысле: в узком смысле:
- 11. Коэффициент наследуемости через корреляции “родитель-потомок”: один родитель - потомок (в случае отсутствия ассортативности) один родитель -
- 12. Ассортативность скрещивания по коэффициенту умственного развития (IQ) выборке супружеских пар в Соединенных Штатах. ■ - муж;
- 13. Схема Менделя Г.И.Мендель 1822–1884 Иоганн Мендель родился 20 июля 1822 года в крестьянской семье. Проучившись два
- 14. Схема Менделя Г.И.Мендель 1822–1884 Когда-то в школьных учебниках Менделя изображали простым, но наблюдательным монахом-провинциалом, чьи открытия
- 15. Аддитивное наследование длины початка кукурузы при скрещивании длиннопочаткового сорта “Черный мексиканец” (справа) с короткопочатковым сортом “Мальчик-с-пальчик”
- 16. Коэффициент наследуемости Lush. I. L. 1937. Animal Breeding Plans. Collegiate Press, Inc., Ames, IA. в случае
- 17. d d/2 m P1 P2 F1 F2 Аддитивно-доминантное наследование количественного признака
- 18. Большинство хозяйственно ценных признаков сельскохозяйственных животных являются количественными (живая масса, высота в холке, удой, настриг шерсти,
- 19. Если развитие признака связано с действием одного гена, то его называют главным геном (олигогеном). В этом
- 20. Например, настриг шерсти у овец, зависит от целого ряда факторов, каждый из которых обусловлен своей группой
- 21. 1) Каждый из полигенов, рассматриваемый отдельно, оказывает незначительное влияние на изменчивость количественного признака. Чем больше пар
- 22. 3) Полигены могут модифицировать выраженность качественных признаков. В этом случае они составляют группу генов-модификаторов. Гены-модификаторы, действуя
- 23. Полимерия (полигения) – генетическая основа изменчивости количественных признаков
- 24. Полимерия (полигения) – генетическая основа изменчивости количественных признаков
- 25. Полимерия (полигения) – генетическая основа изменчивости количественных признаков Основные положения теории К. Мазера полигенного наследования количественных
- 26. Повторяемость Повторяемость имеет важное практическое значение. Она позволяет вести селекцию на большую устойчивость признаков животных, а
- 27. Повторяемость Повторяемость можно оценить по коэффициентам корреляции. Коэффициенты корреляции одних и тех же признаков (например, величина
- 28. Коэффициенты корреляции между лактациями по молочной продуктивности Повторяемость
- 29. Повторяемость Свойства коэффициента повторяемости, rw: 1) rw - это показатель генетического разнообразия; 2) rw является мерой
- 30. Одним из основных параметров, выражающих изменения количественных признаков в результате селекции, является селекционный эффект (Δμ), который
- 31. SD = μотобранных родителей - μ родителей Связь между селекционным эффектом (∆µ) и селекционным дифференциалом (SD)
- 32. Селекционный дифференциал
- 33. Интенсивность селекции (скорость отбора) Соотношение селекционного дифференциала (SD) и фенотипической изменчивости признака (σ2родителей) выражает интенсивность селекции
- 34. Интенсивность селекции (скорость отбора) При очень высокой интенсивности отбора по фенотипу с оставлением на племя 1-3%
- 36. Спасибо за внимание!
- 37. H = GP-1 Многомерный аналог коэффициента наследуемости Lande R (1979). Quantitative genetic analysis of multivariate evolution
- 38. Родители (X) Потомки (Y) P = RX/X G = RX/Y
- 39. Поиск осей с максимальной наследуемостью (в узком смысле) Ott J, Rabinowitz D (1999). A principal-components approach
- 40. Материал
- 42. Родственные связи
- 43. Родственные связи
- 44. Коэффициенты корреляции родителей с потомками по первым пяти компонентам с максимальной аддитивной наследуемостью (выделены достоверные при
- 45. Коэффициент наследуемости через корреляции “родитель-потомок”: один родитель - потомок (в случае отсутствия ассортативности) один родитель -
- 46. Расположение семей на плоскости первых двух компонент аддитивной наследуемости
- 47. Корреляция между родителями и детьми по первой компоненте аддитивной наследуемости
- 48. Корреляция между родителями (ассортативность) по первой компоненте аддитивной наследуемости
- 49. Корреляция между дедушками и бабушками по первой компоненте аддитивной наследуемости
- 50. Корреляция между родителями (два поколения)
- 51. После учета ассортативности выявилось, что четыре компоненты шкалирования имеют значимые коэффициенты наследуемости
- 52. Расположение центроидов родительских и гибридных выборок в многомерном пространстве признаков при аддитивно-доминантной модели наследования F1 -
- 54. Скачать презентацию

Схема Гальтона
Исторически статистический анализ биологических данных по наследуемости количественных признаков начался
Схема Гальтона
Исторически статистический анализ биологических данных по наследуемости количественных признаков начался
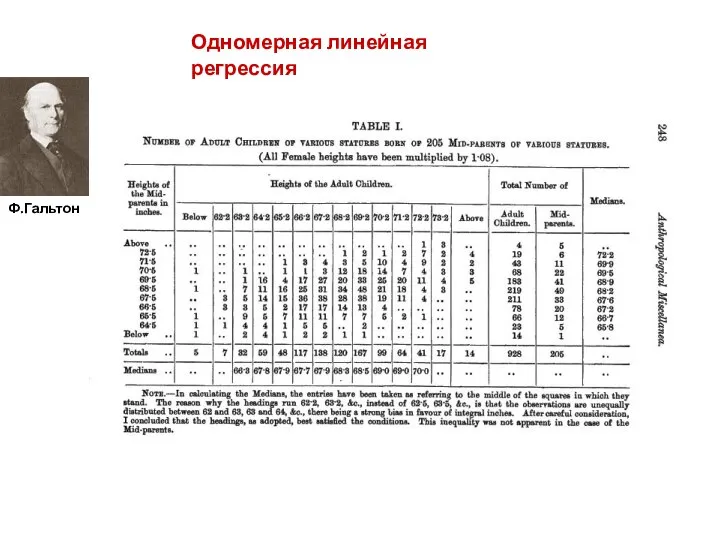
Одномерная линейная регрессия
Ф.Гальтон
Одномерная линейная регрессия
Ф.Гальтон
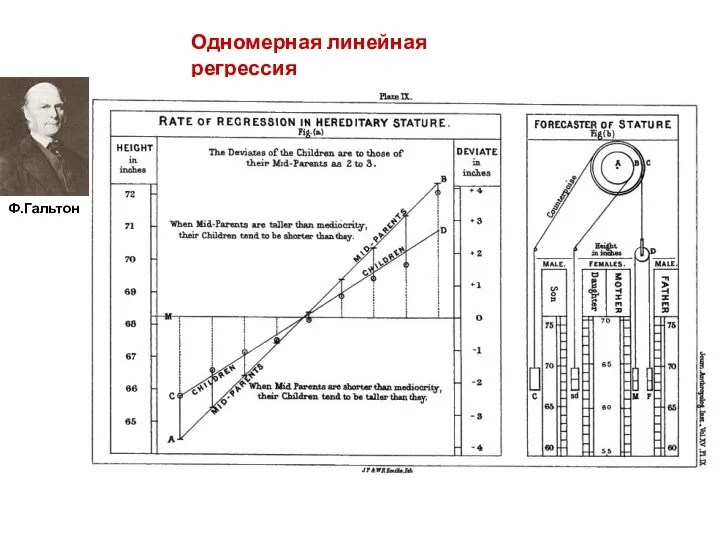
Одномерная линейная регрессия
Ф.Гальтон
Одномерная линейная регрессия
Ф.Гальтон

Одномерная линейная регрессия
Уравнение линейной регресии.
Метод наименьших квадратов
Одномерная линейная регрессия
Уравнение линейной регресии.
Метод наименьших квадратов

Наследственность -- свойство организмов передавать при размножении свои признаки и особенности
Наследственность -- свойство организмов передавать при размножении свои признаки и особенности

Коэффициент наследуемости
(гетерогенные родители и потомки)
Lush. I. L.
Коэффициент наследуемости
(гетерогенные родители и потомки)
Lush. I. L.

Нас интересует прежде всего аддитивный эффект. Именно он отвечает за успешность
Нас интересует прежде всего аддитивный эффект. Именно он отвечает за успешность

Коэффициент наследуемости
(гетерогенные родители и потомки)
Lush. I. L.
Коэффициент наследуемости
(гетерогенные родители и потомки)
Lush. I. L.

Коэффициент наследуемости
(гетерогенные родители и потомки)
в широком смысле:
в узком
Коэффициент наследуемости
(гетерогенные родители и потомки)
в широком смысле:
в узком

Коэффициент наследуемости
через корреляции “родитель-потомок”:
один родитель - потомок
(в случае отсутствия
Коэффициент наследуемости
через корреляции “родитель-потомок”:
один родитель - потомок
(в случае отсутствия
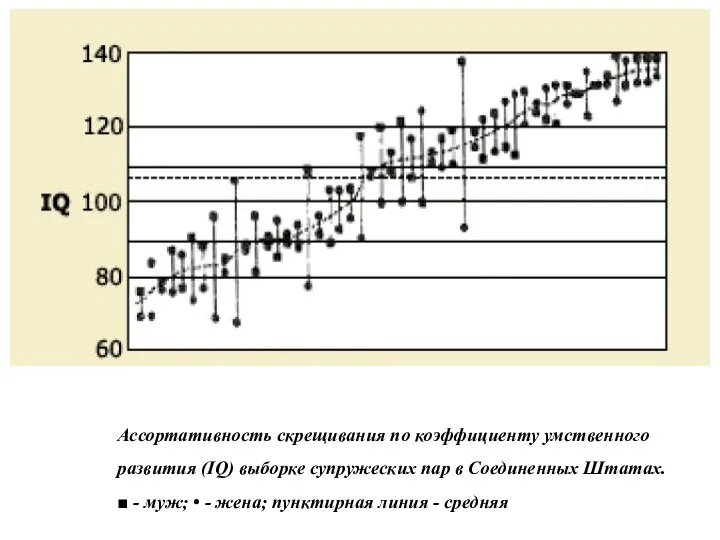
Ассортативность скрещивания по коэффициенту умственного развития (IQ) выборке супружеских пар в
Ассортативность скрещивания по коэффициенту умственного развития (IQ) выборке супружеских пар в

Схема Менделя
Г.И.Мендель
1822–1884
Иоганн Мендель родился 20 июля 1822 года в крестьянской семье. Проучившись два года
Схема Менделя
Г.И.Мендель
1822–1884
Иоганн Мендель родился 20 июля 1822 года в крестьянской семье. Проучившись два года

Схема Менделя
Г.И.Мендель
1822–1884
Когда-то в школьных учебниках Менделя изображали простым, но наблюдательным
Схема Менделя
Г.И.Мендель
1822–1884
Когда-то в школьных учебниках Менделя изображали простым, но наблюдательным
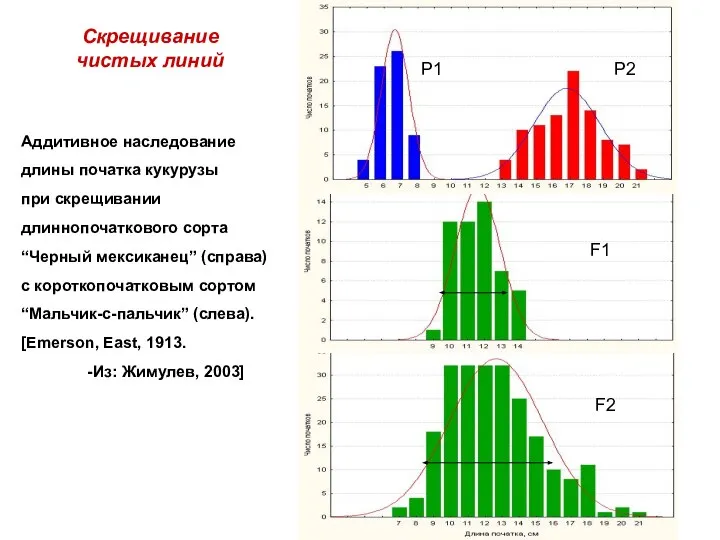
Аддитивное наследование длины початка кукурузы
при скрещивании длиннопочаткового сорта
“Черный мексиканец” (справа)
с
Аддитивное наследование длины початка кукурузы
при скрещивании длиннопочаткового сорта
“Черный мексиканец” (справа)
с

Коэффициент наследуемости
Lush. I. L. 1937. Animal Breeding Plans.
Коэффициент наследуемости
Lush. I. L. 1937. Animal Breeding Plans.
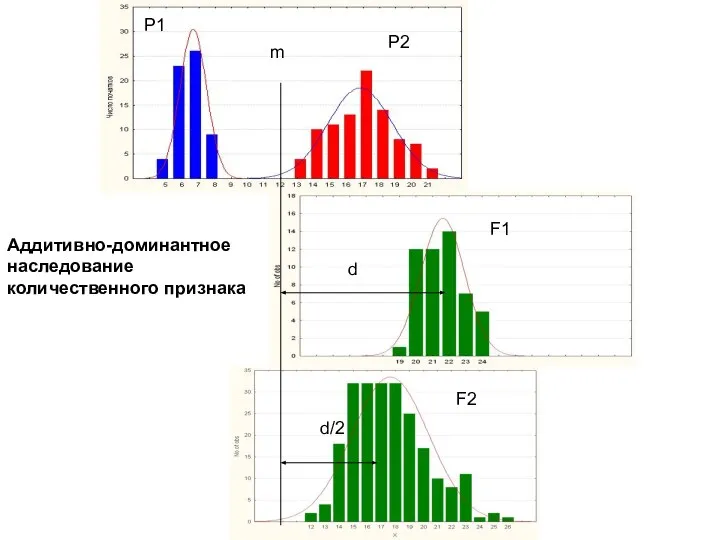
d
d/2
m
P1
P2
F1
F2
Аддитивно-доминантное
наследование
количественного признака
d
d/2
m
P1
P2
F1
F2
Аддитивно-доминантное
наследование
количественного признака

Большинство хозяйственно ценных признаков сельскохозяйственных животных являются количественными (живая масса, высота
Большинство хозяйственно ценных признаков сельскохозяйственных животных являются количественными (живая масса, высота

Если развитие признака связано с действием одного гена, то его называют
Если развитие признака связано с действием одного гена, то его называют
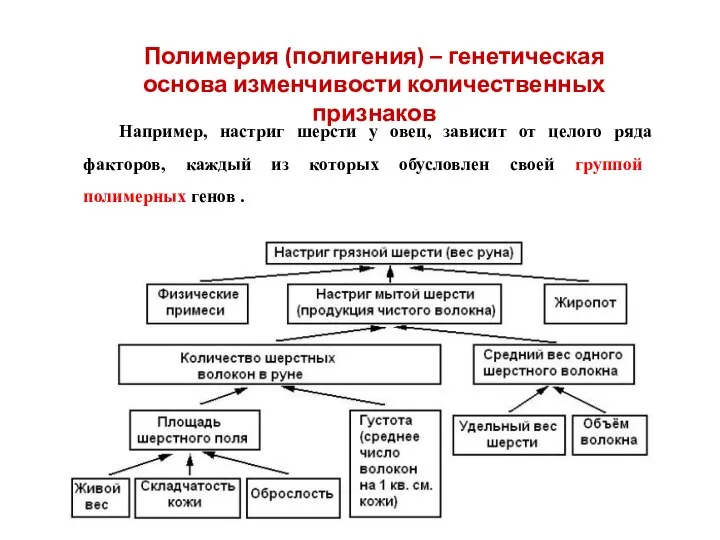
Например, настриг шерсти у овец, зависит от целого ряда факторов, каждый
Например, настриг шерсти у овец, зависит от целого ряда факторов, каждый

1) Каждый из полигенов, рассматриваемый отдельно, оказывает незначительное влияние на изменчивость
1) Каждый из полигенов, рассматриваемый отдельно, оказывает незначительное влияние на изменчивость

3) Полигены могут модифицировать выраженность качественных признаков. В этом случае они
3) Полигены могут модифицировать выраженность качественных признаков. В этом случае они

Полимерия (полигения) – генетическая основа изменчивости количественных признаков
Полимерия (полигения) – генетическая основа изменчивости количественных признаков

Полимерия (полигения) – генетическая основа изменчивости количественных признаков
Полимерия (полигения) – генетическая основа изменчивости количественных признаков

Полимерия (полигения) – генетическая основа изменчивости количественных признаков
Основные положения теории К.
Полимерия (полигения) – генетическая основа изменчивости количественных признаков
Основные положения теории К.

Повторяемость
Повторяемость имеет важное практическое значение. Она позволяет вести селекцию на большую
Повторяемость
Повторяемость имеет важное практическое значение. Она позволяет вести селекцию на большую

Повторяемость
Повторяемость можно оценить по коэффициентам корреляции. Коэффициенты корреляции одних и тех
Повторяемость
Повторяемость можно оценить по коэффициентам корреляции. Коэффициенты корреляции одних и тех
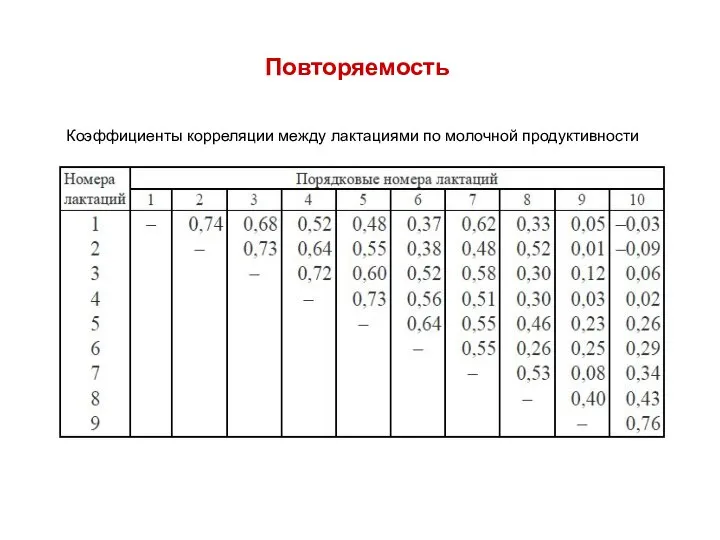
Коэффициенты корреляции между лактациями по молочной продуктивности
Повторяемость
Коэффициенты корреляции между лактациями по молочной продуктивности
Повторяемость

Повторяемость
Свойства коэффициента повторяемости, rw:
1) rw - это показатель генетического разнообразия;
2) rw
Повторяемость
Свойства коэффициента повторяемости, rw:
1) rw - это показатель генетического разнообразия;
2) rw

Одним из основных параметров, выражающих изменения количественных признаков в результате селекции,
Одним из основных параметров, выражающих изменения количественных признаков в результате селекции,

SD = μотобранных родителей - μ родителей
Связь между селекционным эффектом (∆µ)
SD = μотобранных родителей - μ родителей
Связь между селекционным эффектом (∆µ)
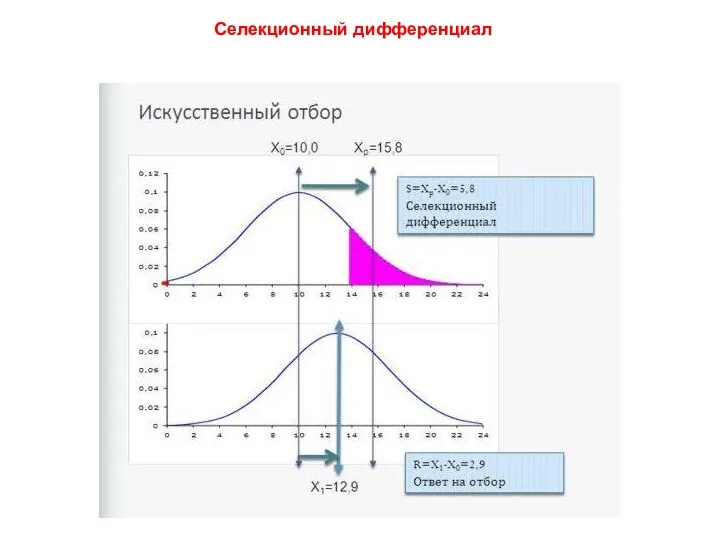
Селекционный дифференциал
Селекционный дифференциал

Интенсивность селекции (скорость отбора)
Соотношение селекционного дифференциала (SD) и фенотипической изменчивости признака
Интенсивность селекции (скорость отбора)
Соотношение селекционного дифференциала (SD) и фенотипической изменчивости признака

Интенсивность селекции (скорость отбора)
При очень высокой интенсивности отбора по фенотипу с
Интенсивность селекции (скорость отбора)
При очень высокой интенсивности отбора по фенотипу с


Спасибо за внимание!
Спасибо за внимание!
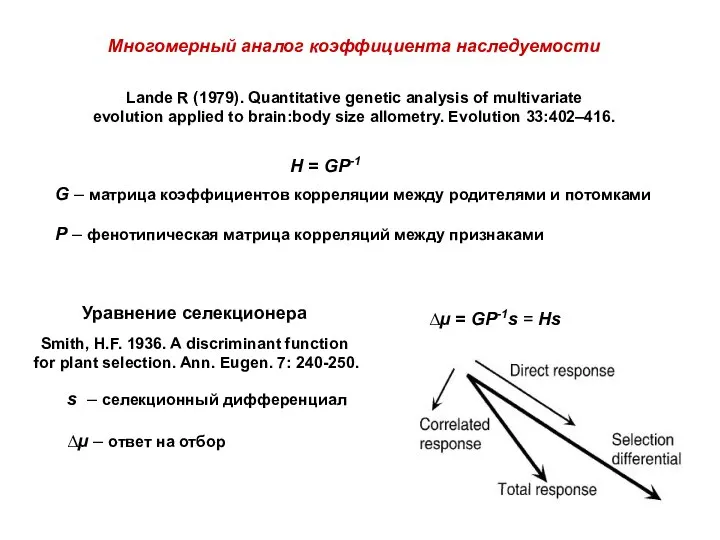
H = GP-1
Многомерный аналог коэффициента наследуемости
Lande R (1979). Quantitative genetic
H = GP-1
Многомерный аналог коэффициента наследуемости
Lande R (1979). Quantitative genetic
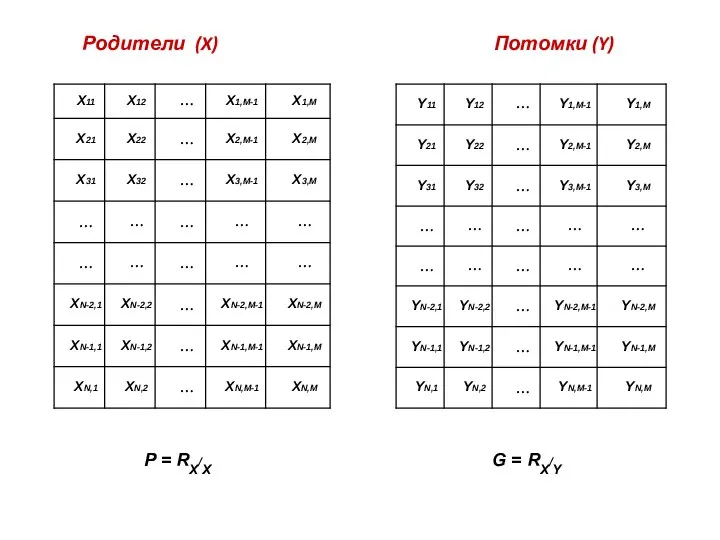
Родители (X) Потомки (Y)
P = RX/X
G = RX/Y
Родители (X) Потомки (Y)
P = RX/X
G = RX/Y

Поиск осей с максимальной наследуемостью
(в узком смысле)
Ott J, Rabinowitz D (1999).
Поиск осей с максимальной наследуемостью
(в узком смысле)
Ott J, Rabinowitz D (1999).
Материал
Материал
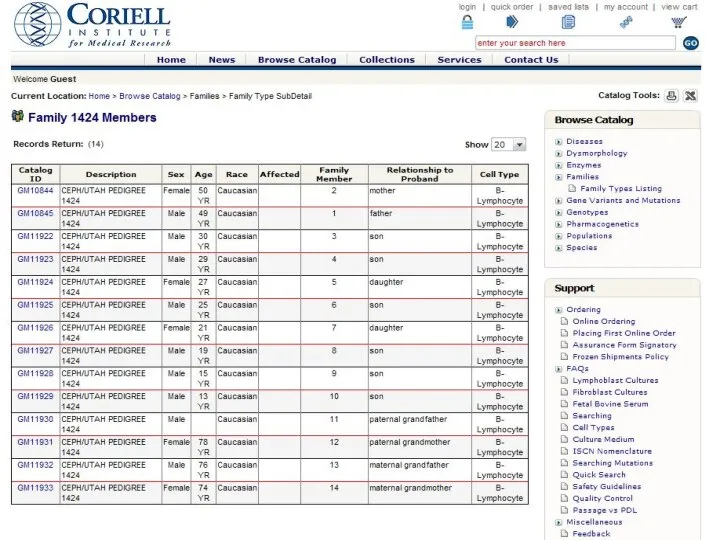
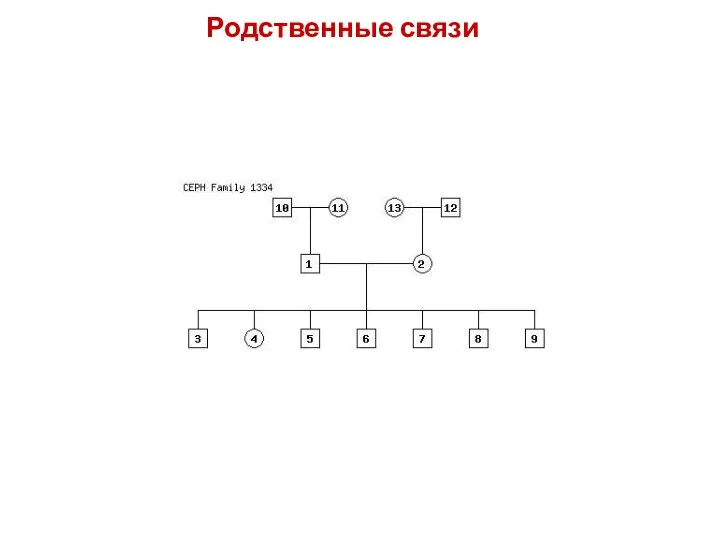
Родственные связи
Родственные связи
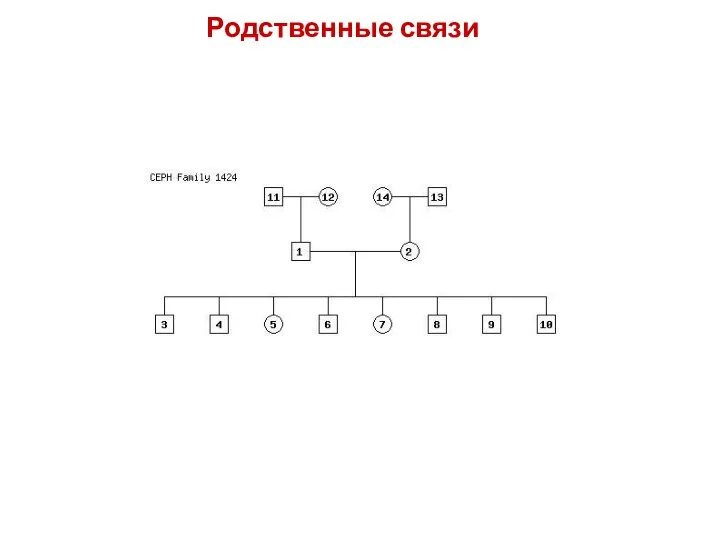
Родственные связи
Родственные связи
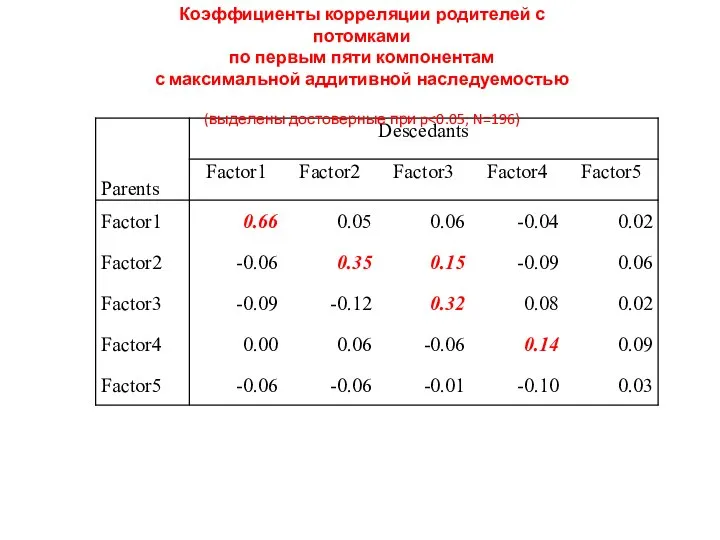
Коэффициенты корреляции родителей с потомками
по первым пяти компонентам
с максимальной аддитивной наследуемостью
(выделены
Коэффициенты корреляции родителей с потомками
по первым пяти компонентам
с максимальной аддитивной наследуемостью
(выделены

Коэффициент наследуемости
через корреляции “родитель-потомок”:
один родитель - потомок
(в случае отсутствия
Коэффициент наследуемости
через корреляции “родитель-потомок”:
один родитель - потомок
(в случае отсутствия
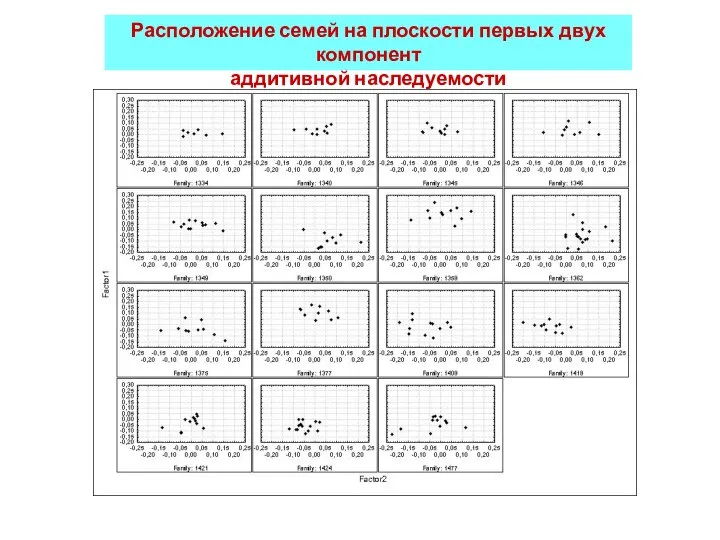
Расположение семей на плоскости первых двух компонент
аддитивной наследуемости
Расположение семей на плоскости первых двух компонент
аддитивной наследуемости
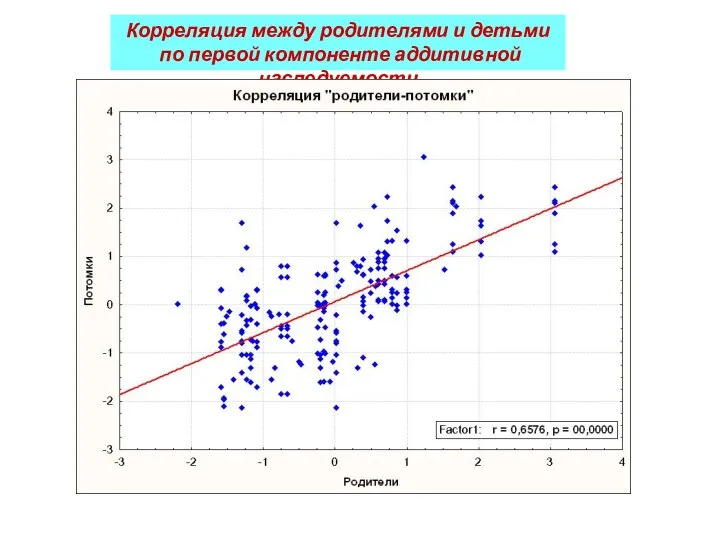
Корреляция между родителями и детьми
по первой компоненте аддитивной наследуемости
Корреляция между родителями и детьми
по первой компоненте аддитивной наследуемости
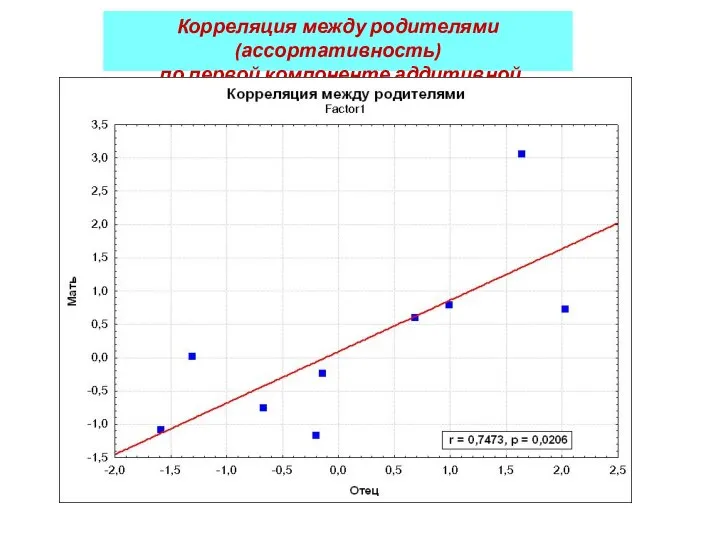
Корреляция между родителями (ассортативность)
по первой компоненте аддитивной наследуемости
Корреляция между родителями (ассортативность)
по первой компоненте аддитивной наследуемости
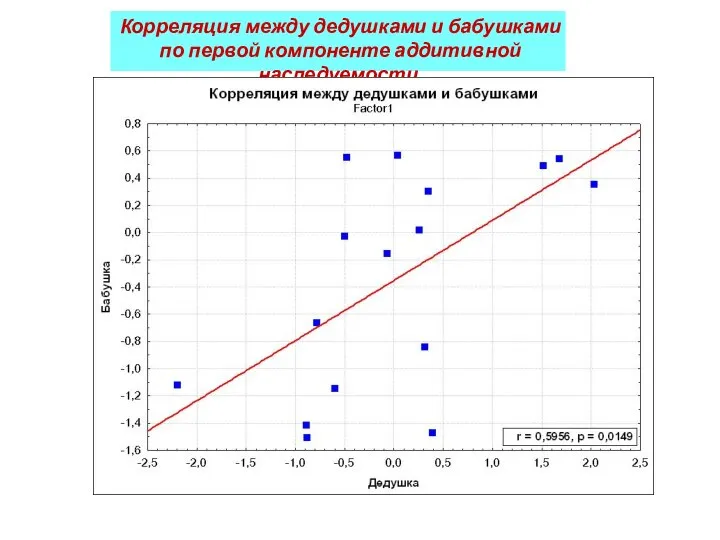
Корреляция между дедушками и бабушками
по первой компоненте аддитивной наследуемости
Корреляция между дедушками и бабушками
по первой компоненте аддитивной наследуемости
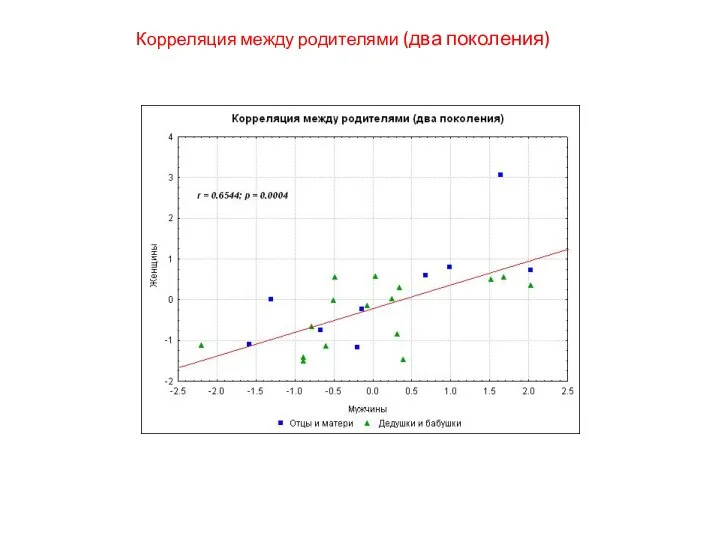
Корреляция между родителями (два поколения)
Корреляция между родителями (два поколения)
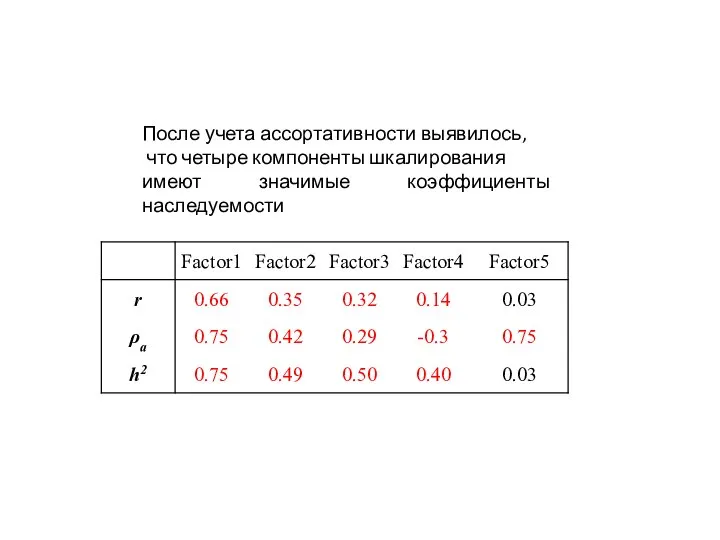
После учета ассортативности выявилось,
что четыре компоненты шкалирования
имеют значимые коэффициенты наследуемости
После учета ассортативности выявилось,
что четыре компоненты шкалирования
имеют значимые коэффициенты наследуемости
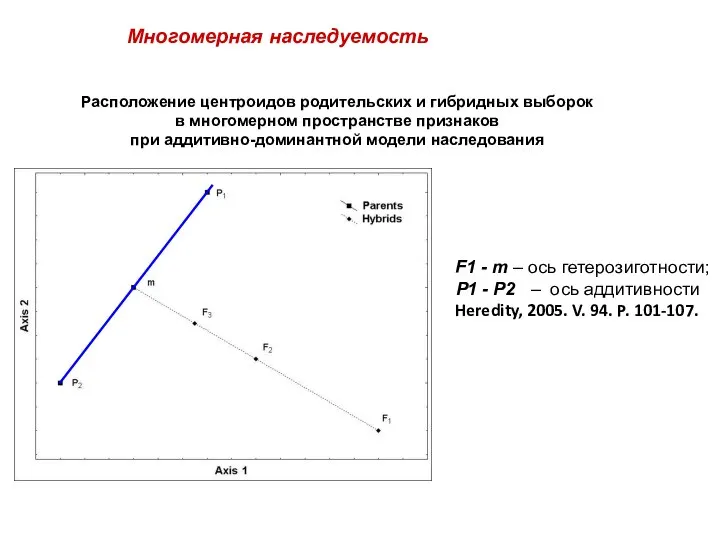
Расположение центроидов родительских и гибридных выборок
в многомерном пространстве признаков
при аддитивно-доминантной
Расположение центроидов родительских и гибридных выборок
в многомерном пространстве признаков
при аддитивно-доминантной
 20140316_trud_i_tvorchestvo
20140316_trud_i_tvorchestvo 20120305_10._povtori_chistogovorki
20120305_10._povtori_chistogovorki Разработка автоматической системы регулирования интервалов движения на железнодорожном перегоне
Разработка автоматической системы регулирования интервалов движения на железнодорожном перегоне Устройство патрона, выключателя
Устройство патрона, выключателя volkova_ebnacproekty_i_reshaem_vmeste_2022
volkova_ebnacproekty_i_reshaem_vmeste_2022 Презентация ЖК “Лефортово Парк”
Презентация ЖК “Лефортово Парк” 20170522_prezentatsiya_vesna_1
20170522_prezentatsiya_vesna_1 Рубский и скопцы
Рубский и скопцы Решение БП на 4 неделю августа
Решение БП на 4 неделю августа Улучшение эффективных показателей автомобилей в АО Гордормостстрой при эксплуатации в условиях г. Киров
Улучшение эффективных показателей автомобилей в АО Гордормостстрой при эксплуатации в условиях г. Киров Организация деятельности структурного подразделения машиностроительного предприятия по изготовлению детали Вал
Организация деятельности структурного подразделения машиностроительного предприятия по изготовлению детали Вал Техническое оснащение и технология работы грузовой станции
Техническое оснащение и технология работы грузовой станции Теплотехнический контроль и сигнализация основных параметров ЦВД турбины К-200-130
Теплотехнический контроль и сигнализация основных параметров ЦВД турбины К-200-130 Биосистема, объекты и методы в биотехнологии
Биосистема, объекты и методы в биотехнологии Запорізький Автомобільний Завод
Запорізький Автомобільний Завод 1881-1894 гг. – правление Александра III в России
1881-1894 гг. – правление Александра III в России Порівняння закладів
Порівняння закладів Тестовые задания для подготовки к ЕГЭ по теме Формы естественного отбора
Тестовые задания для подготовки к ЕГЭ по теме Формы естественного отбора 20130308_obrazovatelnoe_puteshestvie
20130308_obrazovatelnoe_puteshestvie Limitation periods
Limitation periods f6c-35c89872
f6c-35c89872 Святые земли Русской
Святые земли Русской Я, мы, Гитлер: 3 состояния
Я, мы, Гитлер: 3 состояния Мой гардероб. Павел Золотарев
Мой гардероб. Павел Золотарев Отделение непрерывной разливки в составе конвертерного цеха с производительностью 8,5 млн тонн стали в год
Отделение непрерывной разливки в составе конвертерного цеха с производительностью 8,5 млн тонн стали в год Elephone Products 2015
Elephone Products 2015 20130510_kategoriya_geroicheskogo_v_literature_grazhdanskoy_voyny
20130510_kategoriya_geroicheskogo_v_literature_grazhdanskoy_voyny Дизайн проект на тему Растительность на кухне
Дизайн проект на тему Растительность на кухне