- Развиће

Содержание
- 2. Онтогенетско (индивидуално) развиће (грч. онтос = биће; генесис = постанак, развиће) процеси прображаја оплођеног јајета или
- 3. Полно и бесполно размножавање
- 4. Основне фазе онтогенетског развића животиња Гаметогенеза Оплођење (фертилизација) Браздање Гаструлација Органогенеза Раст и хистолошка диференцијација Метаморфоза
- 5. Гаметогенеза Прва фаза оногенетског развића је фаза гаметогенезе – развиће гамета: јајних ћелија - оогенеза (овогенеза)
- 6. Оплођење (фертилизација) Оплођење које представља спајање женског и мушког гамета. Спајањем њихових једара настаје диплоидно једро
- 7. Браздање Браздање представља серију узастопних митотичких деоба оплођеног јајета чиме настаје вишећелијско тело, најчешће лоптастог облика
- 8. Гаструлација У фази гаструлације од једнослојне бластуле настаје прво двослојно, а затим и трослојно тело -
- 9. Органогенеза У фази органогенезе, петој фази развића, долази до формирања органа. У сваком од клициних листова
- 10. Раст и хистолошка диференцијација Развиће различитих типова ћелија Образовани зачеци органа расту, па тако животиња ступњевито
- 11. Метаморфоза и регенерација Последња, седма фаза обухвата све процесе који се јављају у каснијем животу јединке,
- 12. Диференцијација У процесу развића ћелије се диференцирају у различита ткива и органе При томе добијају различите
- 14. Диференцијација – на нивоу гена У основи диференцијације стоји смењивање активности различитих група гена које делују
- 15. Гаметогенеза Основни генетски потенцијал новог организма тј. нове генерације потиче из гамета које стварају родитељи Гамети
- 16. Мејоза У полним органима животиња и биљака настају полне ћелије – гамети. Гамети имају половину укупног
- 17. Профаза I
- 18. Лептонема (лептотен) постепена кондензација хромозома
- 19. Зигонема (зиготен) међусобно приближавање хомологих хромозома. Формирање хромозомских парова – БИВАЛЕНАТА. Хромозоми се пиљубљују у процесу
- 20. Пахинема (пахитен) оба приљубљена хромозома се деле уздужно на по две хроматиде везане заједничком центромером. Тако
- 21. Диплонема (диплотен) Хроматиде хомологих хромозома се међусобно раздвајају (одмичу) Остају везане преко хијазми које су у
- 22. Дијакинеза (дијакинезис) Спирализација хромозома и терминализација хијазми достиже највећи степен Тетраде добијају циклични облик Једарце нестаје
- 23. Метафаза I Тетраде се крећу према екваторијалној равни и постављају у њеној оси Свака тетрада има
- 24. Анфаза I Раздвајање хомологих хромозома, кидање терминалних хијазми Хомологи хромозоми полазе ка супротним половима (дијаде, половине
- 25. Број хромозома се тако своди на половину, а како се биваленти раздвајају независно, мајчински и очински
- 26. Телофаза I Два хаплоидна сета хромозома се групишу на супротним половима ћелије Појављује се једарце Ствара
- 27. Друга мејотичка деоба – Мејоза II Слична је митози Број хромозома који улази у митозу II
- 28. Гамети воде порекло од примарних герминативних ћелија које се јављају у ембриону човека крајем треће недеље.
- 29. Оогенеза Формирање женских гамета започиње око 15 недеље ембрионалмог живота митотском деобом примарних герминативних ћелија при
- 31. Оогенеза У време рођења, у оба јајника се налази око 2 милиона примарних фоликула, а затим
- 32. Оогенеза - мејоза Примарне герминативне ћелије оваријума митозама се деле и дају ООГОНИЈЕ Оогоније расту једно
- 33. Оогенеза - мејоза I ДЕОБА: ооцита I реда дели се на две неједнаке ћелије: ооциту II
- 34. Оогенеза - мејоза
- 35. Вителогенеза Ооците пролазе кроз процес акумулације резервног материјала - ВИТЕЛОГЕНЕЗУ Општи назив за резервни материјал јајета
- 36. Типови јајних ћелија Зависно од количине жуманцета и његове распрострањености у јајној ћелији могу се разликовати
- 37. Сперматогенеза Сперматогенеза је процес настанка сперматозоида. Почиње у пубертету Она се континуирано одвија у семеним каналићима
- 38. Улога сперматозоида Активација јајне ћелије да отпочне са развићем Да своју хаплоидну гарнитуру хромозома преда јајној
- 39. Сперматогенеза се одвија у 4 фазе: СПЕРМАТОЦИТОГЕНЕЗА: сперматогоније се деле митотском деобом и настају примарни сперматоцити,
- 41. Сперматозоиди Глава – код човека дужине 5-7μm, ширине 2.5 - 3.5μm. Акрозом – прекрива предњи део
- 42. Сертолијеве ћелије Исхрана исходних клициних ћелија Ослобађање сперматозоида у лумен семених каналића
- 44. ОПЛОЂЕЊЕ - Фертилизација - Три главне функције фертилизације су: Трансмисија гена са оба родитеља на потомке
- 45. TIPOVI OPLOĐENJA I RAZVIĆA U životinjskom svetu se , prema mestu odvijanja, razlikuju dva osnovna tipa
- 46. Oviparnost (ovum = jaje; parere = rađati): i oplođenje i razviće embriona vrše u spoljašnjoj sredini
- 47. Фертилизација пролази кроз следеће фазе: Контакт између сперматозоида и јајне ћелије (акрозомална реакција) Улазак сперматозоида у
- 48. Aktivacija spermatozoida počinje kada on dodirne omotač jajne ćelije što dovodi do tzv. akrozomske reakcije. Akrozomska
- 49. ФУЗИЈА НУКЛЕУСА Након продора сперматозоида у јајну ћелију, долази до значајних промена у јајној ћелији Прво
- 50. Ране деобе (браздање)
- 51. Након оплођења у зиготу долази до физиолошких промена. Прво у зиготу је, у односу на јајну
- 52. Оплођена јајна ћелија или зигот трансформише се у вишећелијиски организам веома брзо ћелијском деобом. Ћелијске деобе
- 53. Типови браздања Радијално браздање - на ступњу од 8 бластомера свака бластомера горњег (анималног) слоја лежи
- 54. Жуманце има велики утицај на браздање. Вителус смањује браздање на вегетативном полу ембриона. Браздање увек отпочиње
- 55. Бластулација Бластула настаје митотичким деобама зигота Има лоптаст облик Један слој ћелија БЛАСТОМЕРА формира БЛАСТОДЕРМ Унутрашњост
- 56. БЛАСТУЛА
- 57. Типови бластула Постоје четири типа бластула: целобластула – то је лопта на чијој поврчини се налази
- 58. Сисари Од изолецитног оплођеног јајета образује се морула, а потом бластодиск у коме се образује дупља
- 59. Гаструлација Гаструлација укључује серију миграција ћелија (морфогенетских покрета) на позиције где ће оне формирати три примарна
- 60. Начини формирања гаструле су: инвагинација – гаструла настаје из целобластуле, која се на једном делу угиба
- 61. Гаструлација Три примарна ћелијска слоја су: ЕКТОДЕРМ – спољашњи слој МЕЗОДЕРМ – средишњи слој ЕНДОДЕРМ -
- 62. Морфогенеза – нижи хордати Морфогенеза је прогресивно стицање телесног облика и развој одређених структура током ембр.
- 63. Развиће амфиоксуса Браздањем настаје бластула Гаструлација почиње на ступњу од око 800 ћел Долази до заравњивања
- 64. Почиње неурулација: зарављена нервна плоча се увија и образује нервну цев која има отвор неуропор Архентерон
- 65. Неурулација Крајем гаструлације, образује се НЕРВНА ПЛОЧА Нервна плоча Почиње неурулација, образовање НЕРВНЕ ЦЕВИ (6.)
- 66. Развиће водоземаца Крајем гаструлације код водоземаца се формира се нервна плоча – то је једна равна
- 67. Неурулација и ембрионална индукција На формирање организма битан утицај имају околна ткива, тј. средина у којој
- 68. Околна ткива индукују настанак одређеног ткива и органа. Тако настанак нервне цеви (неурулацију) могу да индукују
- 69. Органогенеза Током органогенезе, деобе ћелија су удружене са миграцијом и агрегацијом Сва ткива и органи настају
- 70. Сматра се да су за то одговорни градијенти информационих молекула унутар ембриона који обавештавају ћелије о
- 71. Овде су дати кључни моменти процеса развића код животиња Најдетаљније су проучени код бодљокожацa, нижих хордата,
- 72. Развиће код различитих група
- 73. Ембриогенеза код човека Уместо бластоцела, ингресијом се развија БЛАСТОДИСК Као и код гмизаваца и птица, развијају
- 74. Екстраембрионалне опне Код гмизаваца и птица Жуманцетна кеса – садржи жуманце, има крвне судове који је
- 75. ОВИПАРИТЕТ – целокупно развиће ембриона тек после полагања јаја ОВОВИВИПАРИТЕТ – доношење на свет живих младунаца
- 76. ПЛАЦЕНТАЦИЈА ПЛАЦЕНТА – органи изграђени од ембрионалних и материнских ткива ПО ГРАЂИ се деле на: ХОРИО-ВИТЕЛИНСКУ
- 77. Плацента омогућава пролаз супстанци између материнских и ембрионалних ткива ПЛАЦЕНТАЛНА БАРИЈЕРА (слој ткива који омогућава транспорт)
- 78. Повећањем површине контакта Развитак наставака на површини хориона, тзв. ресица (villi) ДИФУЗНА ПЛАЦЕНТА – вили по
- 79. Смањење дебљине плаценталне баријере Епитело-хоријална плацента: присутно свих 8 елемената баријере (торбари и копитари) Ендотелио-хоријална плацента:
- 80. Код порођаја сисара са епитело-хоријалном плацентом ресице хориона се извлаче из ткива материце и не долази
- 82. Скачать презентацию

Онтогенетско (индивидуално) развиће
(грч. онтос = биће; генесис = постанак, развиће)
процеси
Онтогенетско (индивидуално) развиће
(грч. онтос = биће; генесис = постанак, развиће)
процеси

Полно и бесполно размножавање
Полно и бесполно размножавање

Основне фазе онтогенетског развића животиња
Гаметогенеза
Оплођење (фертилизација)
Браздање
Гаструлација
Органогенеза
Раст и хистолошка диференцијација
Метаморфоза и регенерација
Основне фазе онтогенетског развића животиња
Гаметогенеза
Оплођење (фертилизација)
Браздање
Гаструлација
Органогенеза
Раст и хистолошка диференцијација
Метаморфоза и регенерација

Гаметогенеза
Прва фаза оногенетског развића је фаза гаметогенезе – развиће гамета:
јајних
Гаметогенеза
Прва фаза оногенетског развића је фаза гаметогенезе – развиће гамета:
јајних

Оплођење (фертилизација)
Оплођење које представља спајање женског и мушког гамета.
Спајањем њихових
Оплођење (фертилизација)
Оплођење које представља спајање женског и мушког гамета.
Спајањем њихових

Браздање
Браздање представља серију узастопних митотичких деоба оплођеног јајета чиме настаје вишећелијско
Браздање
Браздање представља серију узастопних митотичких деоба оплођеног јајета чиме настаје вишећелијско

Гаструлација
У фази гаструлације од једнослојне бластуле настаје прво двослојно, а затим
Гаструлација
У фази гаструлације од једнослојне бластуле настаје прво двослојно, а затим

Органогенеза
У фази органогенезе, петој фази развића, долази до формирања органа.
У
Органогенеза
У фази органогенезе, петој фази развића, долази до формирања органа.
У

Раст и хистолошка диференцијација
Развиће различитих типова ћелија
Образовани зачеци органа расту, па
Раст и хистолошка диференцијација
Развиће различитих типова ћелија
Образовани зачеци органа расту, па

Метаморфоза и регенерација
Последња, седма фаза обухвата све процесе који се
Метаморфоза и регенерација
Последња, седма фаза обухвата све процесе који се

Диференцијација
У процесу развића ћелије се диференцирају у различита ткива и
Диференцијација
У процесу развића ћелије се диференцирају у различита ткива и
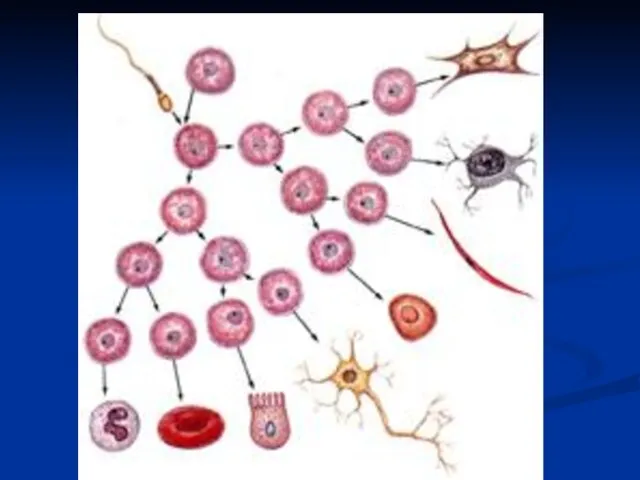

Диференцијација – на нивоу гена
У основи диференцијације стоји смењивање активности различитих
Диференцијација – на нивоу гена
У основи диференцијације стоји смењивање активности различитих

Гаметогенеза
Основни генетски потенцијал новог организма тј. нове генерације потиче из
Гаметогенеза
Основни генетски потенцијал новог организма тј. нове генерације потиче из

Мејоза
У полним органима животиња и биљака настају полне ћелије –
Мејоза
У полним органима животиња и биљака настају полне ћелије –

Профаза I
Профаза I

Лептонема (лептотен)
постепена кондензација хромозома
Лептонема (лептотен)
постепена кондензација хромозома

Зигонема (зиготен)
међусобно приближавање хомологих хромозома.
Формирање хромозомских парова – БИВАЛЕНАТА.
Хромозоми
Зигонема (зиготен)
међусобно приближавање хомологих хромозома.
Формирање хромозомских парова – БИВАЛЕНАТА.
Хромозоми

Пахинема (пахитен)
оба приљубљена хромозома се деле уздужно на по две хроматиде
Пахинема (пахитен)
оба приљубљена хромозома се деле уздужно на по две хроматиде

Диплонема (диплотен)
Хроматиде хомологих хромозома се међусобно раздвајају (одмичу)
Остају везане преко хијазми
Диплонема (диплотен)
Хроматиде хомологих хромозома се међусобно раздвајају (одмичу)
Остају везане преко хијазми

Дијакинеза (дијакинезис)
Спирализација хромозома и терминализација хијазми достиже највећи степен
Тетраде добијају циклични
Дијакинеза (дијакинезис)
Спирализација хромозома и терминализација хијазми достиже највећи степен
Тетраде добијају циклични

Метафаза I
Тетраде се крећу према екваторијалној равни и постављају у
Метафаза I
Тетраде се крећу према екваторијалној равни и постављају у

Анфаза I
Раздвајање хомологих хромозома, кидање терминалних хијазми
Хомологи хромозоми полазе ка супротним
Анфаза I
Раздвајање хомологих хромозома, кидање терминалних хијазми
Хомологи хромозоми полазе ка супротним

Број хромозома се тако своди на половину, а како се биваленти
Број хромозома се тако своди на половину, а како се биваленти

Телофаза I
Два хаплоидна сета хромозома се групишу на супротним половима ћелије
Појављује
Телофаза I
Два хаплоидна сета хромозома се групишу на супротним половима ћелије
Појављује

Друга мејотичка деоба – Мејоза II
Слична је митози
Број хромозома који улази
Друга мејотичка деоба – Мејоза II
Слична је митози
Број хромозома који улази

Гамети воде порекло од примарних герминативних ћелија које се јављају у
Гамети воде порекло од примарних герминативних ћелија које се јављају у

Оогенеза
Формирање женских гамета започиње око 15 недеље ембрионалмог живота митотском
Оогенеза
Формирање женских гамета започиње око 15 недеље ембрионалмог живота митотском
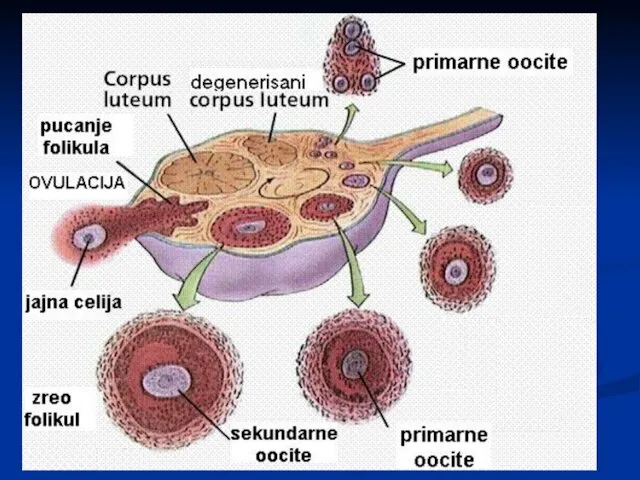

Оогенеза
У време рођења, у оба јајника се налази око 2 милиона
Оогенеза
У време рођења, у оба јајника се налази око 2 милиона

Оогенеза - мејоза
Примарне герминативне ћелије оваријума митозама се деле и дају
Оогенеза - мејоза
Примарне герминативне ћелије оваријума митозама се деле и дају

Оогенеза - мејоза
I ДЕОБА: ооцита I реда дели се на две
Оогенеза - мејоза
I ДЕОБА: ооцита I реда дели се на две
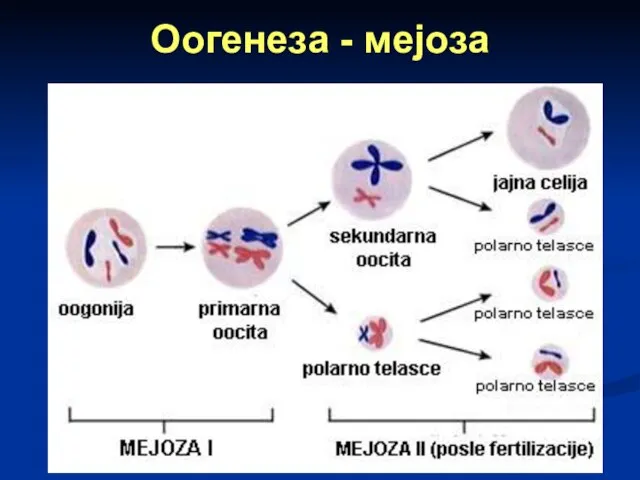
Оогенеза - мејоза
Оогенеза - мејоза

Вителогенеза
Ооците пролазе кроз процес акумулације резервног материјала - ВИТЕЛОГЕНЕЗУ
Општи назив за
Вителогенеза
Ооците пролазе кроз процес акумулације резервног материјала - ВИТЕЛОГЕНЕЗУ
Општи назив за

Типови јајних ћелија
Зависно од количине жуманцета и његове распрострањености у
Типови јајних ћелија
Зависно од количине жуманцета и његове распрострањености у

Сперматогенеза
Сперматогенеза је процес настанка сперматозоида.
Почиње у пубертету
Она се континуирано
Сперматогенеза
Сперматогенеза је процес настанка сперматозоида.
Почиње у пубертету
Она се континуирано

Улога сперматозоида
Активација јајне ћелије да отпочне са развићем
Да своју хаплоидну гарнитуру
Улога сперматозоида
Активација јајне ћелије да отпочне са развићем
Да своју хаплоидну гарнитуру

Сперматогенеза се одвија у 4 фазе:
СПЕРМАТОЦИТОГЕНЕЗА: сперматогоније се деле митотском деобом
Сперматогенеза се одвија у 4 фазе:
СПЕРМАТОЦИТОГЕНЕЗА: сперматогоније се деле митотском деобом
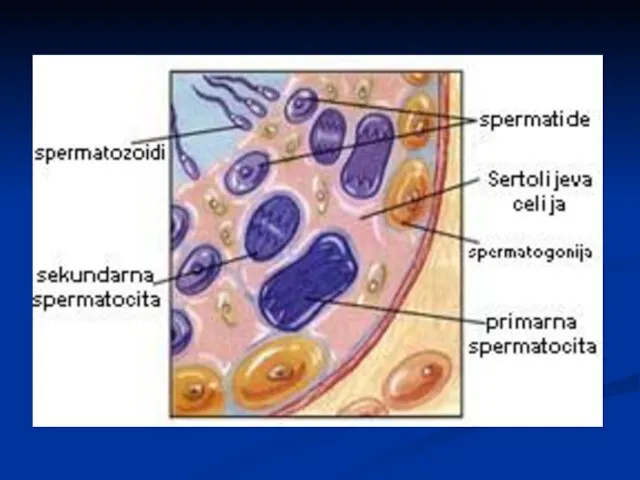

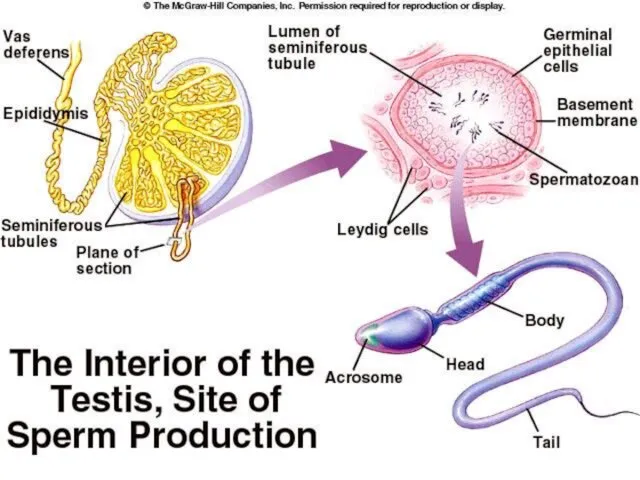
Сперматозоиди
Глава – код човека дужине 5-7μm, ширине 2.5 - 3.5μm.
Акрозом
Сперматозоиди
Глава – код човека дужине 5-7μm, ширине 2.5 - 3.5μm.
Акрозом

Сертолијеве ћелије
Исхрана исходних клициних ћелија
Ослобађање сперматозоида у лумен семених каналића
Сертолијеве ћелије
Исхрана исходних клициних ћелија
Ослобађање сперматозоида у лумен семених каналића

ОПЛОЂЕЊЕ
- Фертилизација -
Три главне функције фертилизације су:
Трансмисија гена са оба
ОПЛОЂЕЊЕ
- Фертилизација -
Три главне функције фертилизације су:
Трансмисија гена са оба

TIPOVI OPLOĐENJA I RAZVIĆA
U životinjskom svetu se , prema mestu odvijanja,
TIPOVI OPLOĐENJA I RAZVIĆA
U životinjskom svetu se , prema mestu odvijanja,

Oviparnost (ovum = jaje; parere = rađati): i oplođenje i razviće
Oviparnost (ovum = jaje; parere = rađati): i oplođenje i razviće

Фертилизација пролази кроз следеће фазе:
Контакт између сперматозоида и јајне ћелије (акрозомална
Фертилизација пролази кроз следеће фазе:
Контакт између сперматозоида и јајне ћелије (акрозомална

Aktivacija spermatozoida
počinje kada on dodirne omotač jajne ćelije što dovodi
Aktivacija spermatozoida
počinje kada on dodirne omotač jajne ćelije što dovodi

ФУЗИЈА НУКЛЕУСА
Након продора сперматозоида у јајну ћелију, долази до значајних промена
ФУЗИЈА НУКЛЕУСА
Након продора сперматозоида у јајну ћелију, долази до значајних промена
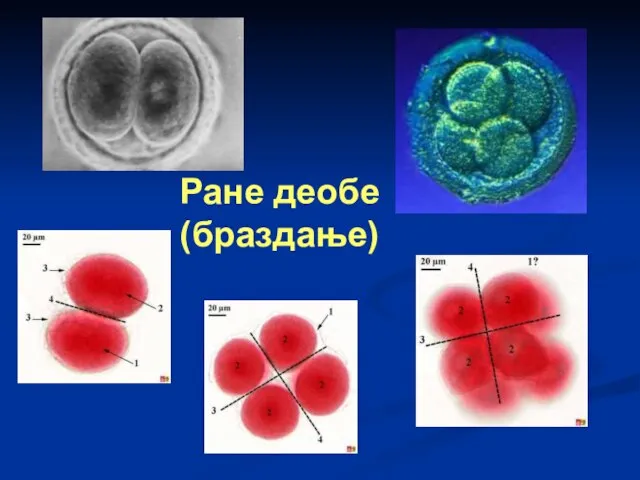
Ране деобе
(браздање)
Ране деобе
(браздање)

Након оплођења у зиготу долази до физиолошких промена. Прво у зиготу
Након оплођења у зиготу долази до физиолошких промена. Прво у зиготу

Оплођена јајна ћелија или зигот трансформише се у вишећелијиски организам веома
Оплођена јајна ћелија или зигот трансформише се у вишећелијиски организам веома

Типови браздања
Радијално браздање - на ступњу од 8 бластомера свака бластомера
Типови браздања
Радијално браздање - на ступњу од 8 бластомера свака бластомера

Жуманце има велики утицај на браздање.
Вителус смањује браздање на вегетативном полу
Жуманце има велики утицај на браздање.
Вителус смањује браздање на вегетативном полу
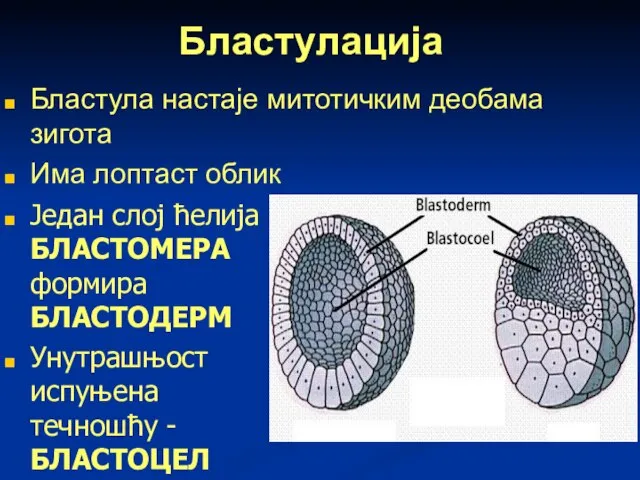
Бластулација
Бластула настаје митотичким деобама зигота
Има лоптаст облик
Један слој ћелија БЛАСТОМЕРА
Бластулација
Бластула настаје митотичким деобама зигота
Има лоптаст облик
Један слој ћелија БЛАСТОМЕРА
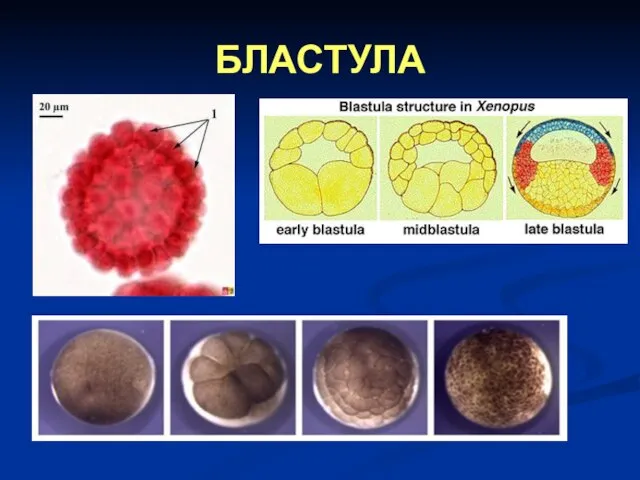
БЛАСТУЛА
БЛАСТУЛА

Типови бластула
Постоје четири типа бластула:
целобластула – то је лопта на чијој
Типови бластула
Постоје четири типа бластула:
целобластула – то је лопта на чијој

Сисари
Од изолецитног оплођеног јајета образује се морула, а потом бластодиск
Сисари
Од изолецитног оплођеног јајета образује се морула, а потом бластодиск
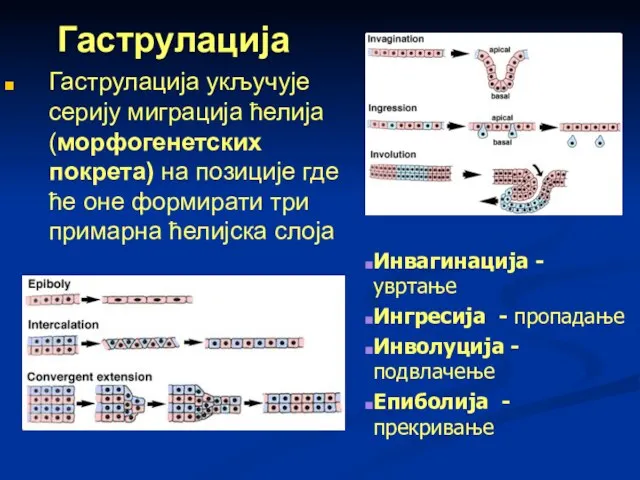
Гаструлација
Гаструлација укључује серију миграција ћелија (морфогенетских покрета) на позиције где
Гаструлација
Гаструлација укључује серију миграција ћелија (морфогенетских покрета) на позиције где

Начини формирања гаструле су:
инвагинација – гаструла настаје из целобластуле, која се
Начини формирања гаструле су:
инвагинација – гаструла настаје из целобластуле, која се
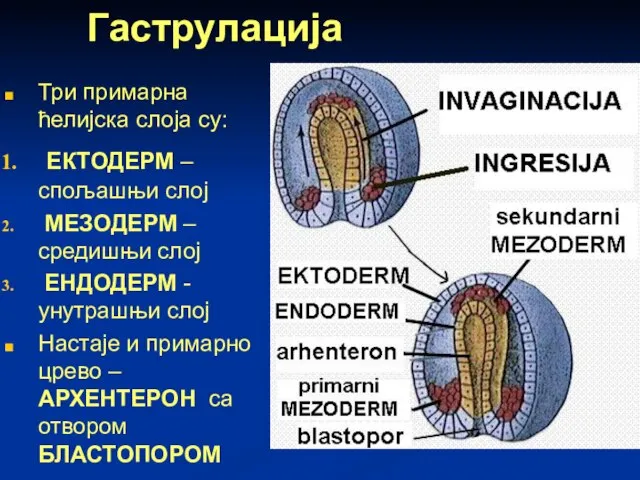
Гаструлација
Три примарна ћелијска слоја су:
ЕКТОДЕРМ – спољашњи слој
МЕЗОДЕРМ –
Гаструлација
Три примарна ћелијска слоја су:
ЕКТОДЕРМ – спољашњи слој
МЕЗОДЕРМ –

Морфогенеза – нижи хордати
Морфогенеза је прогресивно стицање телесног облика и развој
Морфогенеза – нижи хордати
Морфогенеза је прогресивно стицање телесног облика и развој

Развиће амфиоксуса
Браздањем настаје бластула
Гаструлација почиње на ступњу од око 800 ћел
Долази
Развиће амфиоксуса
Браздањем настаје бластула
Гаструлација почиње на ступњу од око 800 ћел
Долази

Почиње неурулација: зарављена нервна плоча се увија и образује нервну цев
Почиње неурулација: зарављена нервна плоча се увија и образује нервну цев
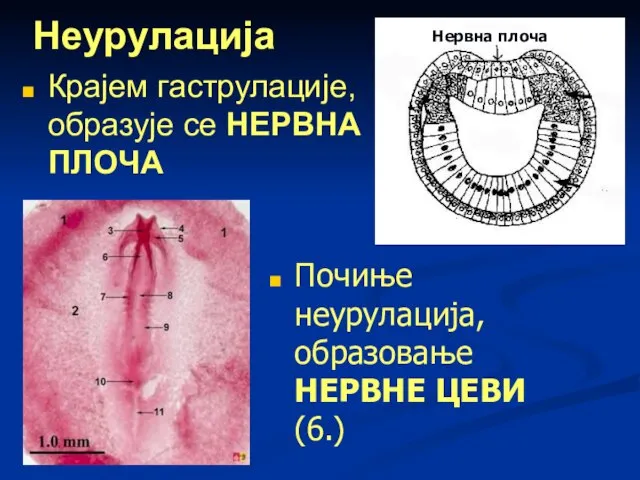
Неурулација
Крајем гаструлације, образује се НЕРВНА ПЛОЧА
Нервна плоча
Почиње неурулација, образовање НЕРВНЕ
Неурулација
Крајем гаструлације, образује се НЕРВНА ПЛОЧА
Нервна плоча
Почиње неурулација, образовање НЕРВНЕ

Развиће водоземаца
Крајем гаструлације код водоземаца се формира се нервна плоча –
Развиће водоземаца
Крајем гаструлације код водоземаца се формира се нервна плоча –

Неурулација и ембрионална индукција
На формирање организма битан утицај имају околна ткива,
Неурулација и ембрионална индукција
На формирање организма битан утицај имају околна ткива,

Околна ткива индукују настанак одређеног ткива и органа.
Тако настанак нервне
Околна ткива индукују настанак одређеног ткива и органа.
Тако настанак нервне

Органогенеза
Током органогенезе, деобе ћелија су удружене са миграцијом и агрегацијом
Органогенеза
Током органогенезе, деобе ћелија су удружене са миграцијом и агрегацијом

Сматра се да су за то одговорни градијенти информационих молекула унутар
Сматра се да су за то одговорни градијенти информационих молекула унутар
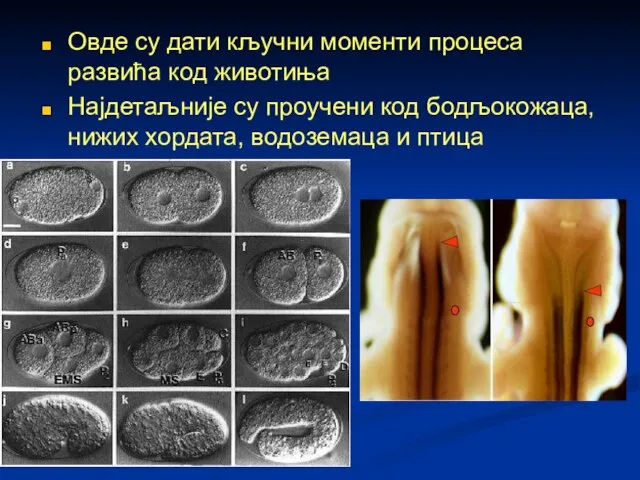
Овде су дати кључни моменти процеса развића код животиња
Најдетаљније су проучени
Овде су дати кључни моменти процеса развића код животиња
Најдетаљније су проучени
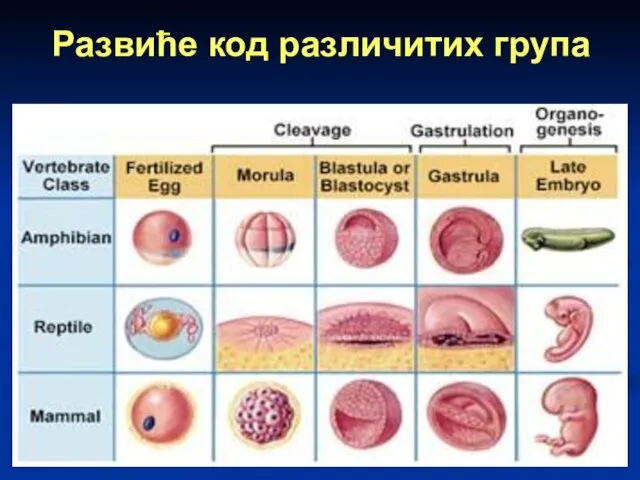
Развиће код различитих група
Развиће код различитих група
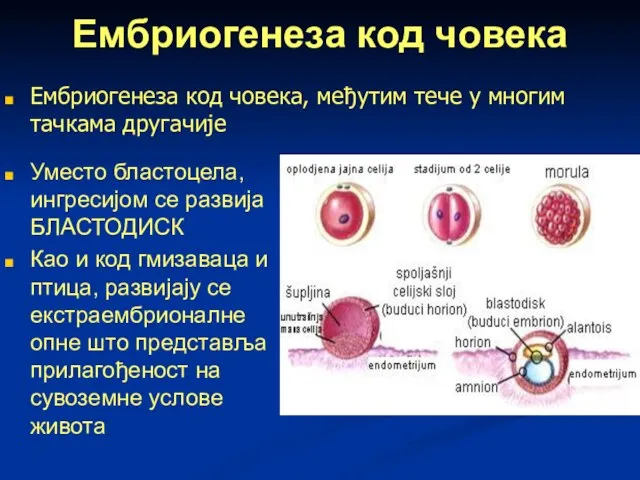
Ембриогенеза код човека
Уместо бластоцела, ингресијом се развија БЛАСТОДИСК
Као и код гмизаваца
Ембриогенеза код човека
Уместо бластоцела, ингресијом се развија БЛАСТОДИСК
Као и код гмизаваца

Екстраембрионалне опне
Код гмизаваца и птица
Жуманцетна кеса – садржи жуманце, има крвне
Екстраембрионалне опне
Код гмизаваца и птица
Жуманцетна кеса – садржи жуманце, има крвне

ОВИПАРИТЕТ – целокупно развиће ембриона тек после полагања јаја
ОВОВИВИПАРИТЕТ – доношење
ОВИПАРИТЕТ – целокупно развиће ембриона тек после полагања јаја
ОВОВИВИПАРИТЕТ – доношење

ПЛАЦЕНТАЦИЈА
ПЛАЦЕНТА – органи изграђени од ембрионалних и материнских ткива
ПО ГРАЂИ се
ПЛАЦЕНТАЦИЈА
ПЛАЦЕНТА – органи изграђени од ембрионалних и материнских ткива
ПО ГРАЂИ се

Плацента омогућава пролаз супстанци између материнских и ембрионалних ткива
ПЛАЦЕНТАЛНА БАРИЈЕРА (слој
Плацента омогућава пролаз супстанци између материнских и ембрионалних ткива
ПЛАЦЕНТАЛНА БАРИЈЕРА (слој

Повећањем површине контакта
Развитак наставака на површини хориона, тзв. ресица (villi)
ДИФУЗНА ПЛАЦЕНТА
Повећањем површине контакта
Развитак наставака на површини хориона, тзв. ресица (villi)
ДИФУЗНА ПЛАЦЕНТА

Смањење дебљине плаценталне баријере
Епитело-хоријална плацента: присутно свих 8 елемената баријере (торбари
Смањење дебљине плаценталне баријере
Епитело-хоријална плацента: присутно свих 8 елемената баријере (торбари

Код порођаја сисара са епитело-хоријалном плацентом ресице хориона се извлаче из
Код порођаја сисара са епитело-хоријалном плацентом ресице хориона се извлаче из
 Неразъёмные соединения. (Лекция 4)
Неразъёмные соединения. (Лекция 4) Political system of Azerbaijan
Political system of Azerbaijan Чувашская Республика _
Чувашская Республика _ Социальные сети в России
Социальные сети в России Формы общения
Формы общения Презентация "Ананьинская культура" - скачать презентации по МХК
Презентация "Ананьинская культура" - скачать презентации по МХК Лев Толстой в Ясной Поляне
Лев Толстой в Ясной Поляне Буксы. Общие сведения
Буксы. Общие сведения КОНТРОЛЬНАЯ РАБОТА ПО МХК В 10 КЛАССЕ (ПРОГРАММА ПОД РЕДАКЦИЕЙ Л.Г. ЕМОХОНОВОЙ) ХУДОЖЕСТВЕННАЯ КУЛЬТУРА СРЕДНИХ ВЕКОВ. ЗАПАДНАЯ ЕВ
КОНТРОЛЬНАЯ РАБОТА ПО МХК В 10 КЛАССЕ (ПРОГРАММА ПОД РЕДАКЦИЕЙ Л.Г. ЕМОХОНОВОЙ) ХУДОЖЕСТВЕННАЯ КУЛЬТУРА СРЕДНИХ ВЕКОВ. ЗАПАДНАЯ ЕВ Пироэлектрики
Пироэлектрики Школьная форма
Школьная форма Общая характеристика подсистемы радиоразведки автоматизированных станций помех
Общая характеристика подсистемы радиоразведки автоматизированных станций помех Політ через хмари Амазона Аудиторія: developers, project managers, sysadmins Anton Marfey, 2011 amarfey@softserveinc.com. - презентация
Політ через хмари Амазона Аудиторія: developers, project managers, sysadmins Anton Marfey, 2011 amarfey@softserveinc.com. - презентация Нарушение белкового обмена при патологии печени Выполнила: Студентка 3 курса
Нарушение белкового обмена при патологии печени Выполнила: Студентка 3 курса Центральный Федеральный округ
Центральный Федеральный округ Тип даних множина
Тип даних множина Строки. Инициализация строки
Строки. Инициализация строки Теории демократии
Теории демократии Геноцид в Мьянме
Геноцид в Мьянме Электромобиль
Электромобиль Wizard of Oz. Туда и обратно
Wizard of Oz. Туда и обратно Язычество восточных славян
Язычество восточных славян Программирование на алгоритмическом языке (§ 54 - § 61)
Программирование на алгоритмическом языке (§ 54 - § 61) Презентация Юрий Михайлович Лотман
Презентация Юрий Михайлович Лотман Электрические коммутационные аппараты, устройство, классификация и основные характеристики
Электрические коммутационные аппараты, устройство, классификация и основные характеристики Спорт. Страницы смерти
Спорт. Страницы смерти Закрепление внетабличного умножения и деления
Закрепление внетабличного умножения и деления Характеристика общения в спорте
Характеристика общения в спорте