- ИММУНОГЛОБУЛИНЫ

Содержание
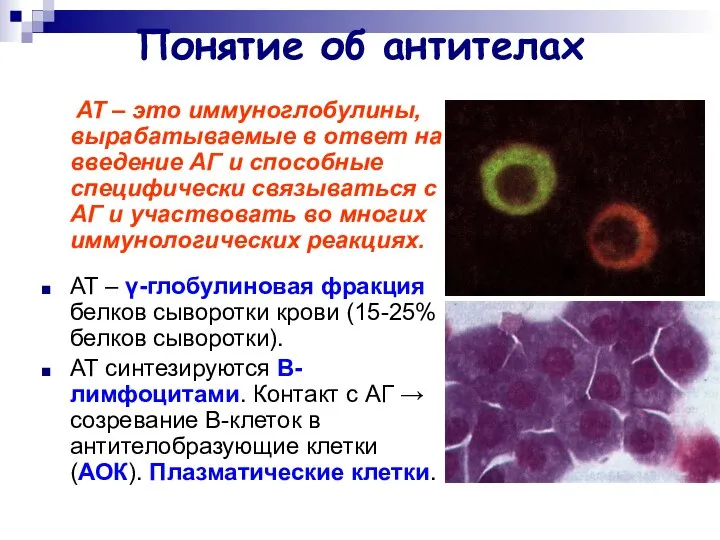
- 2. Понятие об антителах АТ – это иммуноглобулины, вырабатываемые в ответ на введение АГ и способные специфически
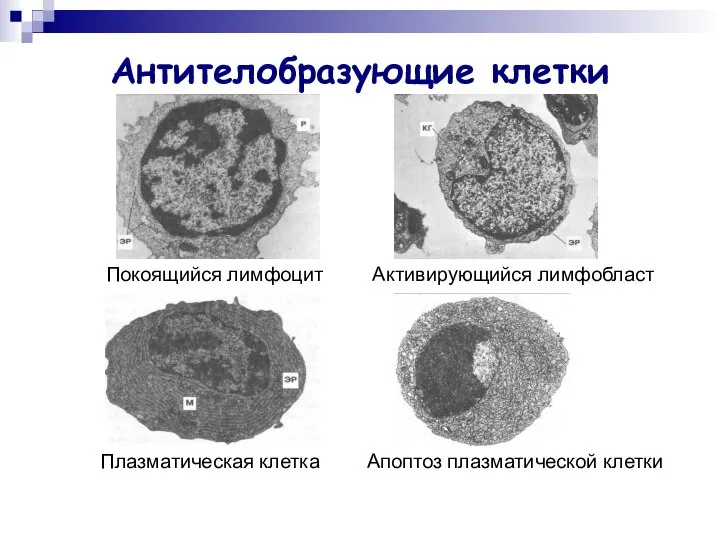
- 3. Антителобразующие клетки Покоящийся лимфоцит Активирующийся лимфобласт Плазматическая клетка Апоптоз плазматической клетки
- 4. Типы иммуноглобулинов Иммуноглобулины: ∙ циркулирующие АТ (сывороточные и секреторные); ∙ рецепторные молекулы на иммунных клетках; ∙
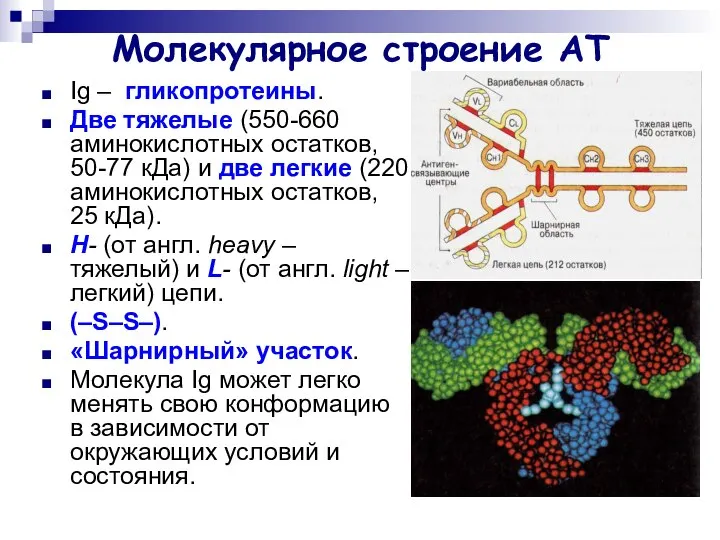
- 5. Молекулярное строение АТ Ig – гликопротеины. Две тяжелые (550-660 аминокислотных остатков, 50-77 кДа) и две легкие
- 6. Молекулярное строение АТ Легкие цепи: κ и λ. Тяжелые цепи: α, γ, μ, ε и δ.
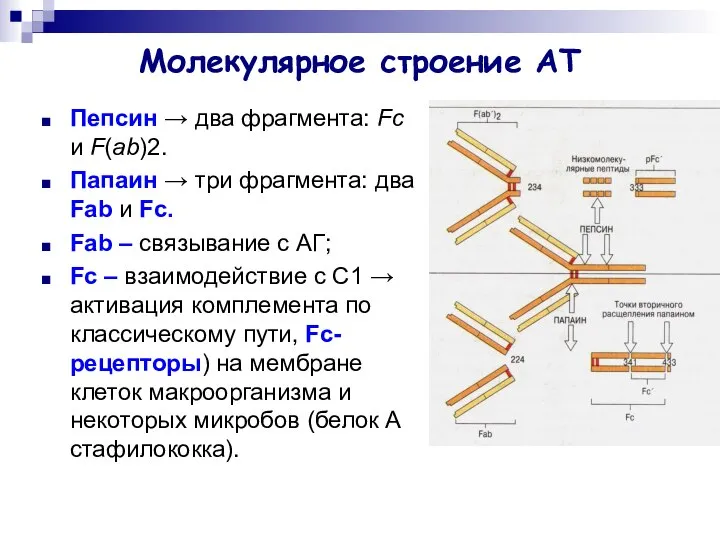
- 7. Молекулярное строение АТ Пепсин → два фрагмента: Fc и F(ab)2. Папаин → три фрагмента: два Fab
- 8. Дополнительные полипептидные цепи Ig IgM, IgA – J-пептид (от англ. join – соединяю). Секреторные IgA –
- 9. Механизм взаимодействия АТ с АГ В процессе взаимодействия с АГ участвует антигенсвязывающий центр (паратоп) Fab-фрагмента. АТ
- 10. Механизм взаимодействия АТ с АГ [АГ]+[АТ] ↔ [ИК] Сила нековалентной связи зависит прежде всего от расстояния
- 11. Механизм взаимодействия АТ с АГ
- 12. Аффинность АТ Аффинность – прочность связи одного антигенсвязывающего центра с индивидуальным эпитопом АГ. Зависит от степени
- 13. Авидность АТ Авидность – прочность связывания АТ и АГ (суммарная сила). Определяется аффинностью и числом антигенсвязывающих
- 14. Эффективность взаимодействия АГ и АТ Доступность эпитопа для антигенсвязывающего центра Ig, число эпитопов в составе молекулы
- 15. Защитные механизмы гуморального иммунитета Cвязывание с АГ : ∙ маркирование АГ, инактивация биологически активных молекул (токсинов),
- 16. Структурно-функциональные особенности иммуноглобулинов различных классов Молекулы, содержащие тяжелую цепь α-типа, относят к изотипу А (сокращенно IgА),
- 17. Иммуноглобулин класса G IgG составляет 70 – 75 % иммуноглобулинов сыворотки крови, 50 % содержится в
- 18. Иммуноглобулин класса G IgG синтезируется зрелыми B-лимфоцитами (Вγ) и плазматическими клетками, хорошо определяется в сыворотке крови
- 19. Иммуноглобулин класса М IgМ – пентамер, 10 антигенсвязывающих центров, молекулярная масса – около 970 кДа, константа
- 20. Иммуноглобулин класса М IgМ – высокая авидность, связывает комплемент, сывороточный и секреторный гуморальный иммунитет. Большая часть
- 21. Иммуноглобулин класса А Сывороточный IgА: около 15 – 20 % всех сывороточных Ig. Период полураспада IgА
- 22. Иммуноглобулин класса А Секреторный (sIgА): ди-, три- или тетрамеры, содержит J- и S-пептиды. Молекулярная масса IgА
- 23. Иммуноглобулин класса Е IgЕ – реагины – около 0,002 % всех циркулирующих Ig, молекулярная масса около
- 24. Иммуноглобулин класса D IgD – 0,2 % общего количества циркулирующих АТ, но обильно представлен на мембране
- 25. Рецепторные иммуноглобулины Рецепторные (мембранные) Ig локализуются на ЦПМ B-лимфоцитов. Антигенспецифические рецепторы. Имеют тот же изотип и
- 26. Нормальные антитела Нормальные (естественные) АТ – базальный уровень иммуноглобулинов. Изогемагглютинины – АТ, направленные против эритроцитарных АГ
- 27. Моноклональные антитела Каждый B-лимфоцит и его потомки (клон) → АТ строго определенной специфичности – моноклинальные. Д.
- 28. Полные и неполные антитела Полные АТ – способны образовывать в РА или РП хорошо различимую глазом
- 29. Генетика иммуноглобулинов Структура Ig контролируется большим числом генов, которые имеют фрагментарную организацию, располагаются на 3 различных
- 30. Динамика антителопродукции Первичный иммунный ответ: длительная латентная (3 – 5 сут) и логарифмическая (7 – 15
- 31. Формирование первичного и вторичного иммунного ответа (первичное воздействие) (повторное воздействие)
- 32. Теории разнообразия антител Теория «боковых цепей» П. Эрлиха (1898) заложила основные представления о гуморальном иммунитете и
- 33. Теории разнообразия антител «Клонально-селекционная» теория Ф. Бернета. Лимфоидная ткань состоит из огромного числа клонов лимфоцитов, которые
- 34. Теории разнообразия антител Молекулярно-генетическая теория С. Тонегавы. Теория сетевой регуляции иммунной системы. В основе Н. идея
- 35. Теории разнообразия антител В 60-е годы П.Ф. Здродовский → физиологическая концепция иммуногенеза – гипоталамо-адреналовая теория регуляции
- 36. Источники разнообразия АТ ∙ Множественность гаметных генов V-областей. ∙ Соматический мутагенез. ∙ Соматические рекомбинации между сегментами,
- 38. Скачать презентацию
Понятие об антителах
АТ – это иммуноглобулины, вырабатываемые в ответ на
Понятие об антителах
АТ – это иммуноглобулины, вырабатываемые в ответ на
Антителобразующие клетки
Покоящийся лимфоцит
Активирующийся лимфобласт
Плазматическая клетка
Апоптоз плазматической клетки
Антителобразующие клетки
Покоящийся лимфоцит
Активирующийся лимфобласт
Плазматическая клетка
Апоптоз плазматической клетки

Типы иммуноглобулинов
Иммуноглобулины:
∙ циркулирующие АТ (сывороточные и секреторные);
∙ рецепторные молекулы
Типы иммуноглобулинов
Иммуноглобулины:
∙ циркулирующие АТ (сывороточные и секреторные);
∙ рецепторные молекулы
Молекулярное строение АТ
Ig – гликопротеины.
Две тяжелые (550-660 аминокислотных остатков, 50-77
Молекулярное строение АТ
Ig – гликопротеины.
Две тяжелые (550-660 аминокислотных остатков, 50-77

Молекулярное строение АТ
Легкие цепи: κ и λ.
Тяжелые цепи: α, γ,
Молекулярное строение АТ
Легкие цепи: κ и λ.
Тяжелые цепи: α, γ,
Молекулярное строение АТ
Пепсин → два фрагмента: Fc и F(ab)2.
Папаин → три
Молекулярное строение АТ
Пепсин → два фрагмента: Fc и F(ab)2.
Папаин → три

Дополнительные полипептидные цепи Ig
IgM, IgA – J-пептид (от англ. join –
Дополнительные полипептидные цепи Ig
IgM, IgA – J-пептид (от англ. join –
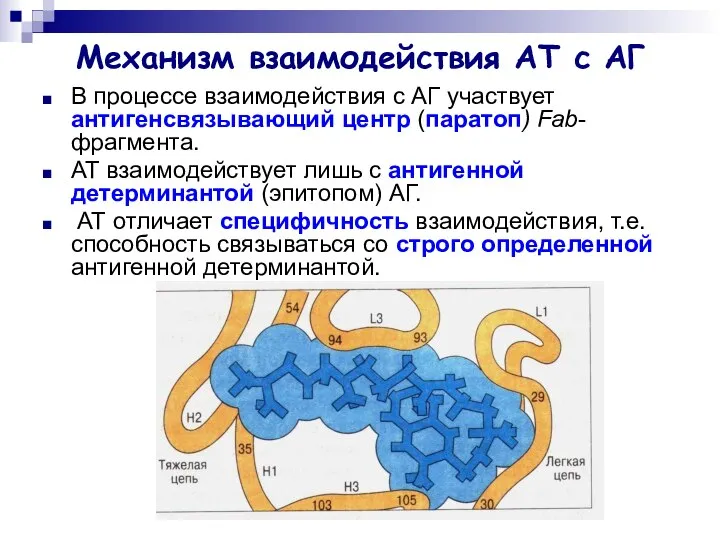
Механизм взаимодействия АТ с АГ
В процессе взаимодействия с АГ участвует антигенсвязывающий
Механизм взаимодействия АТ с АГ
В процессе взаимодействия с АГ участвует антигенсвязывающий
![Механизм взаимодействия АТ с АГ [АГ]+[АТ] ↔ [ИК] Сила нековалентной связи](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1303076/slide-9.jpg)
Механизм взаимодействия АТ с АГ
[АГ]+[АТ] ↔ [ИК]
Сила нековалентной связи зависит
Механизм взаимодействия АТ с АГ
[АГ]+[АТ] ↔ [ИК]
Сила нековалентной связи зависит
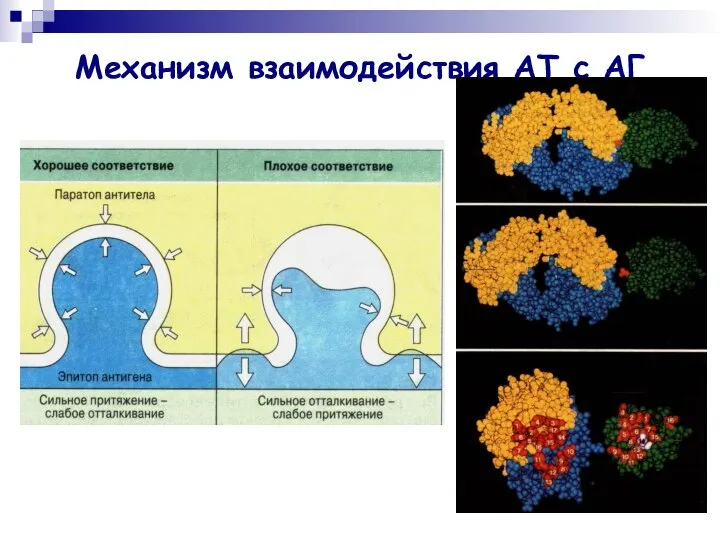
Механизм взаимодействия АТ с АГ
Механизм взаимодействия АТ с АГ
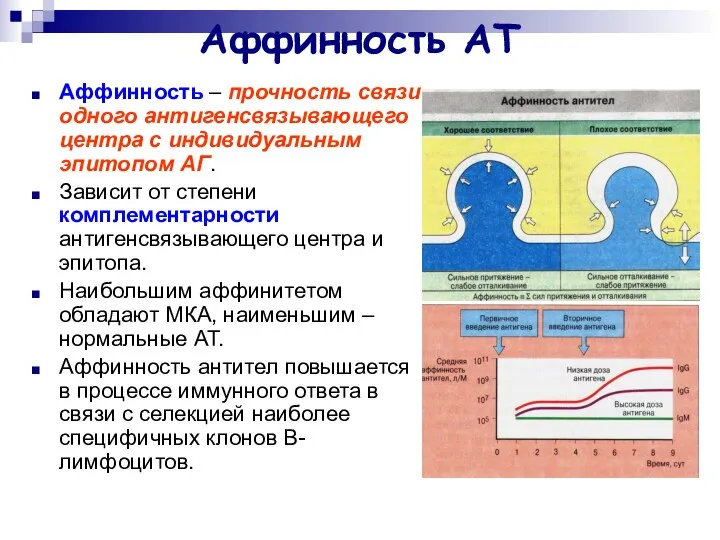
Аффинность АТ
Аффинность – прочность связи одного антигенсвязывающего центра с индивидуальным эпитопом
Аффинность АТ
Аффинность – прочность связи одного антигенсвязывающего центра с индивидуальным эпитопом

Авидность АТ
Авидность – прочность связывания АТ и АГ (суммарная сила).
Определяется
Авидность АТ
Авидность – прочность связывания АТ и АГ (суммарная сила).
Определяется

Эффективность взаимодействия АГ и АТ
Доступность эпитопа для антигенсвязывающего центра Ig, число
Эффективность взаимодействия АГ и АТ
Доступность эпитопа для антигенсвязывающего центра Ig, число

Защитные механизмы гуморального иммунитета
Cвязывание с АГ :
∙ маркирование АГ, инактивация
Защитные механизмы гуморального иммунитета
Cвязывание с АГ :
∙ маркирование АГ, инактивация

Структурно-функциональные особенности иммуноглобулинов различных классов
Молекулы, содержащие тяжелую цепь α-типа, относят к
Структурно-функциональные особенности иммуноглобулинов различных классов
Молекулы, содержащие тяжелую цепь α-типа, относят к
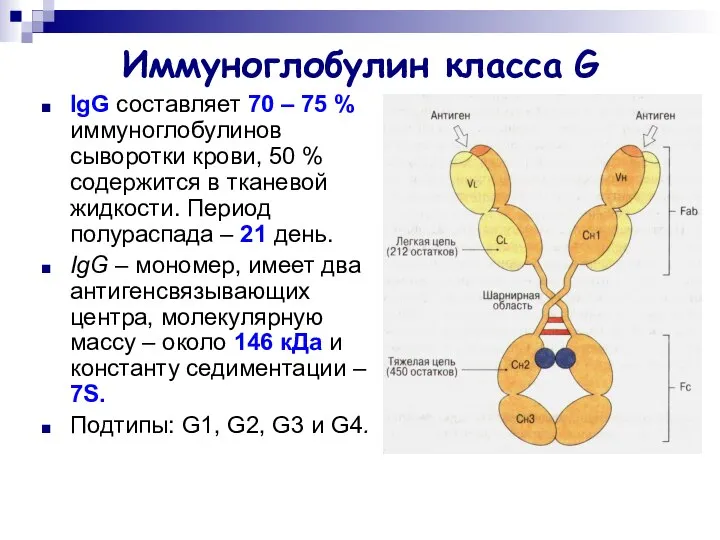
Иммуноглобулин класса G
IgG составляет 70 – 75 % иммуноглобулинов сыворотки крови,
Иммуноглобулин класса G
IgG составляет 70 – 75 % иммуноглобулинов сыворотки крови,

Иммуноглобулин класса G
IgG синтезируется зрелыми B-лимфоцитами (Вγ) и плазматическими клетками, хорошо
Иммуноглобулин класса G
IgG синтезируется зрелыми B-лимфоцитами (Вγ) и плазматическими клетками, хорошо
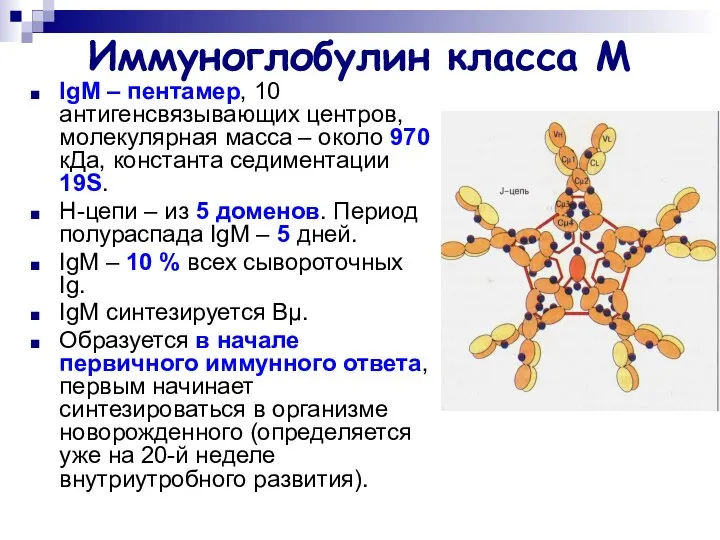
Иммуноглобулин класса М
IgМ – пентамер, 10 антигенсвязывающих центров, молекулярная масса –
Иммуноглобулин класса М
IgМ – пентамер, 10 антигенсвязывающих центров, молекулярная масса –
Иммуноглобулин класса М
IgМ – высокая авидность, связывает комплемент, сывороточный и секреторный
Иммуноглобулин класса М
IgМ – высокая авидность, связывает комплемент, сывороточный и секреторный

Иммуноглобулин класса А
Сывороточный IgА: около 15 – 20 % всех сывороточных
Иммуноглобулин класса А
Сывороточный IgА: около 15 – 20 % всех сывороточных

Иммуноглобулин класса А
Секреторный (sIgА): ди-, три- или тетрамеры, содержит J- и
Иммуноглобулин класса А
Секреторный (sIgА): ди-, три- или тетрамеры, содержит J- и

Иммуноглобулин класса Е
IgЕ – реагины – около 0,002 % всех циркулирующих
Иммуноглобулин класса Е
IgЕ – реагины – около 0,002 % всех циркулирующих
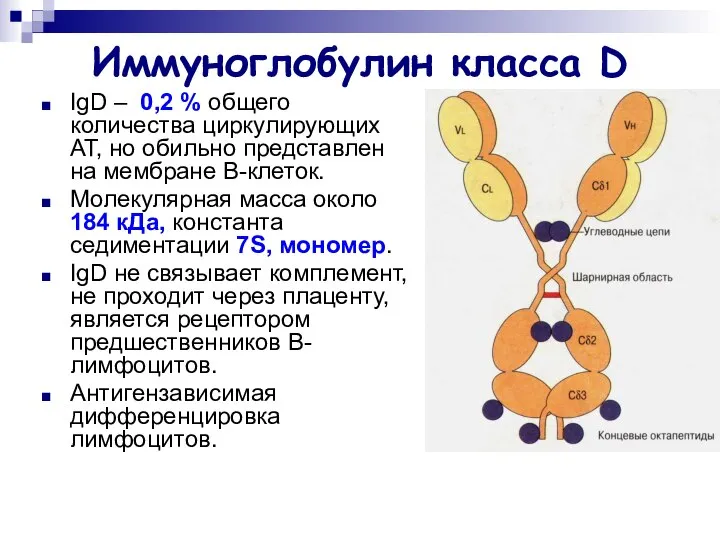
Иммуноглобулин класса D
IgD – 0,2 % общего количества циркулирующих АТ, но
Иммуноглобулин класса D
IgD – 0,2 % общего количества циркулирующих АТ, но
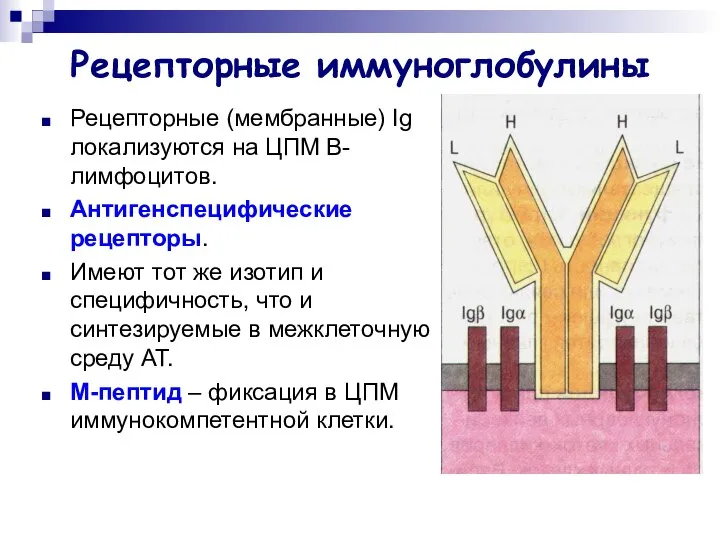
Рецепторные иммуноглобулины
Рецепторные (мембранные) Ig локализуются на ЦПМ B-лимфоцитов.
Антигенспецифические рецепторы.
Имеют
Рецепторные иммуноглобулины
Рецепторные (мембранные) Ig локализуются на ЦПМ B-лимфоцитов.
Антигенспецифические рецепторы.
Имеют

Нормальные антитела
Нормальные (естественные) АТ – базальный уровень иммуноглобулинов.
Изогемагглютинины – АТ,
Нормальные антитела
Нормальные (естественные) АТ – базальный уровень иммуноглобулинов.
Изогемагглютинины – АТ,
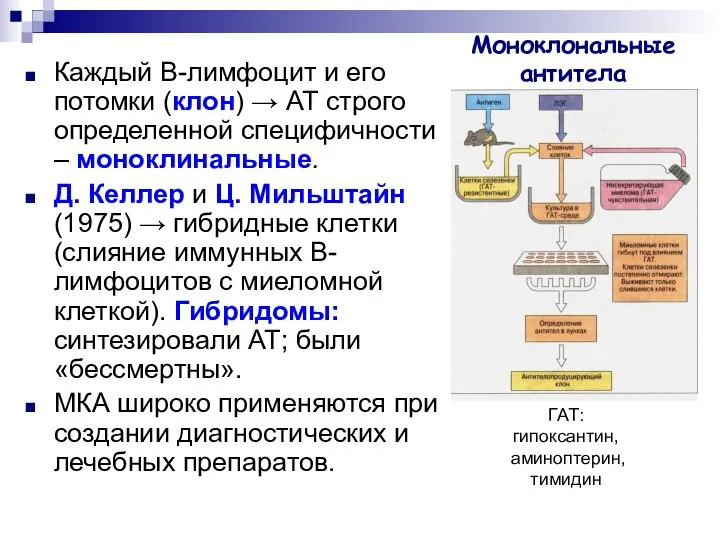
Моноклональные антитела
Каждый B-лимфоцит и его потомки (клон) → АТ строго определенной
Моноклональные антитела
Каждый B-лимфоцит и его потомки (клон) → АТ строго определенной

Полные и неполные антитела
Полные АТ – способны образовывать в РА или
Полные и неполные антитела
Полные АТ – способны образовывать в РА или
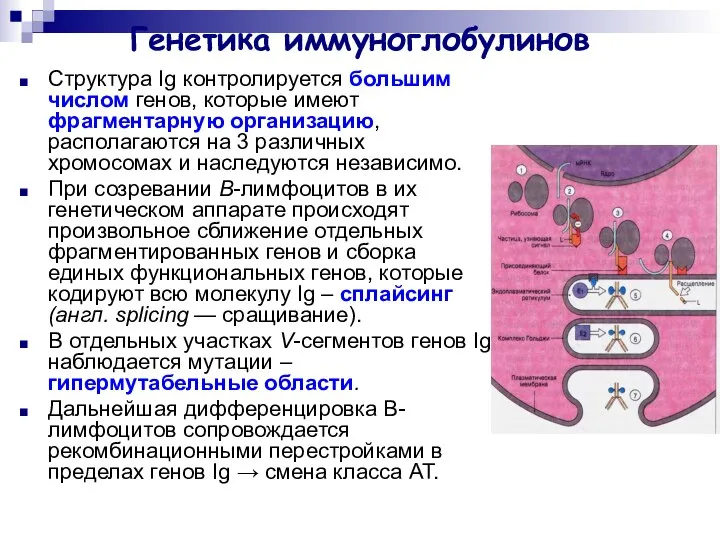
Генетика иммуноглобулинов
Структура Ig контролируется большим числом генов, которые имеют фрагментарную организацию,
Генетика иммуноглобулинов
Структура Ig контролируется большим числом генов, которые имеют фрагментарную организацию,
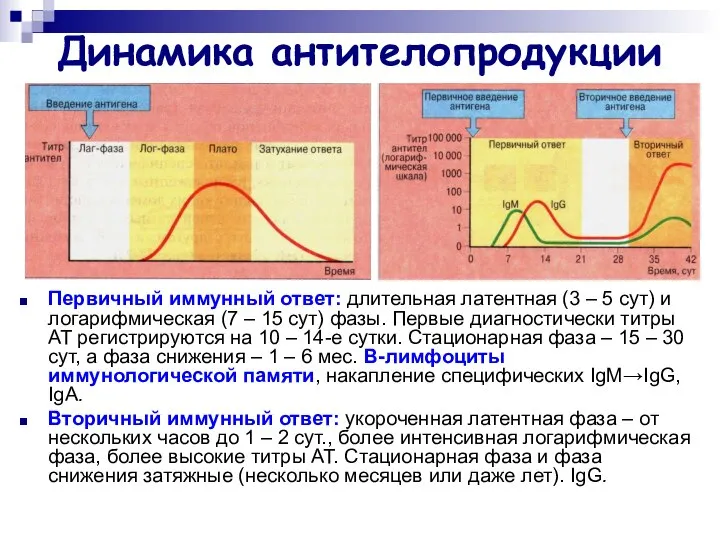
Динамика антителопродукции
Первичный иммунный ответ: длительная латентная (3 – 5 сут)
Динамика антителопродукции
Первичный иммунный ответ: длительная латентная (3 – 5 сут)
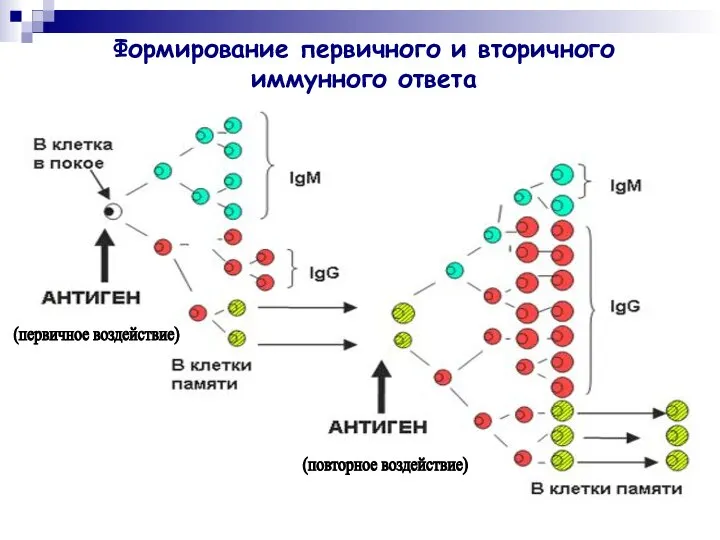
Формирование первичного и вторичного иммунного ответа
(первичное воздействие)
(повторное воздействие)
Формирование первичного и вторичного иммунного ответа
(первичное воздействие)
(повторное воздействие)
Теории разнообразия антител
Теория «боковых цепей» П. Эрлиха (1898) заложила основные представления
Теории разнообразия антител
Теория «боковых цепей» П. Эрлиха (1898) заложила основные представления

Теории разнообразия антител
«Клонально-селекционная» теория Ф. Бернета.
Лимфоидная ткань состоит из огромного
Теории разнообразия антител
«Клонально-селекционная» теория Ф. Бернета.
Лимфоидная ткань состоит из огромного

Теории разнообразия антител
Молекулярно-генетическая теория С. Тонегавы.
Теория сетевой регуляции иммунной системы. В
Теории разнообразия антител
Молекулярно-генетическая теория С. Тонегавы.
Теория сетевой регуляции иммунной системы. В

Теории разнообразия антител
В 60-е годы П.Ф. Здродовский → физиологическая концепция иммуногенеза
Теории разнообразия антител
В 60-е годы П.Ф. Здродовский → физиологическая концепция иммуногенеза
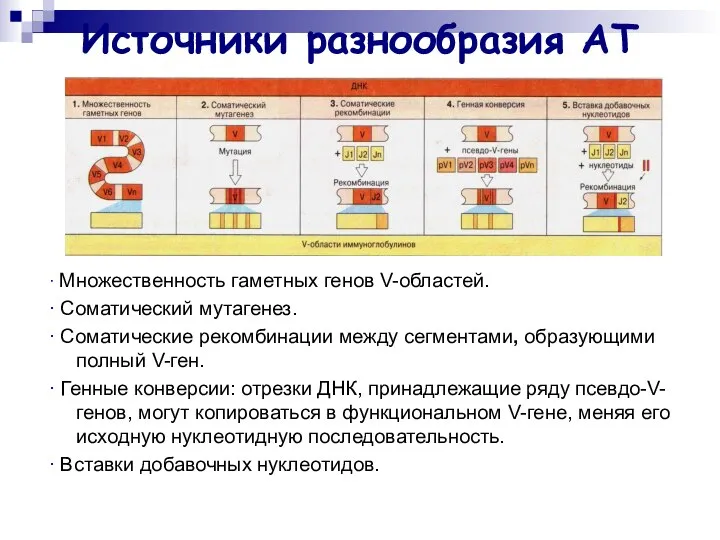
Источники разнообразия АТ
∙ Множественность гаметных генов V-областей.
∙ Соматический мутагенез.
∙ Соматические рекомбинации
Источники разнообразия АТ
∙ Множественность гаметных генов V-областей.
∙ Соматический мутагенез.
∙ Соматические рекомбинации
 ККО Прокопьевск-ПК-Угольный. Система видеонаблюдения
ККО Прокопьевск-ПК-Угольный. Система видеонаблюдения Как нарисовать рыбку? - презентация для начальной школы_
Как нарисовать рыбку? - презентация для начальной школы_ Государственное регулирование охраны здоровья граждан, здравоохранения и медицинской деятельности
Государственное регулирование охраны здоровья граждан, здравоохранения и медицинской деятельности Геополитические взаимоотношения России и Китая
Геополитические взаимоотношения России и Китая Проект о создание Туризма В Шегарском районе, области, стране, зарубежье
Проект о создание Туризма В Шегарском районе, области, стране, зарубежье Кондиционирование воздуха
Кондиционирование воздуха Социальные сети - современный инструмент для профессионального общения библиотекарей
Социальные сети - современный инструмент для профессионального общения библиотекарей древнерусская литература 1 ур
древнерусская литература 1 ур ТЕЛЕСКОПИ
ТЕЛЕСКОПИ Влияние алкоголя на организм человека Исполнители: Поротников Антон, Паклин Роман ученики 8 «б» класс
Влияние алкоголя на организм человека Исполнители: Поротников Антон, Паклин Роман ученики 8 «б» класс Люсьен Теньер
Люсьен Теньер Альбрехт Дюрер (1471-1528)
Альбрехт Дюрер (1471-1528) Основные способы плавания
Основные способы плавания Организация крупного фармпроизводства в соответствии со стандартом GMP
Организация крупного фармпроизводства в соответствии со стандартом GMP Травматизм. Первичная и вторичная профилактика. Первая помощь при травмах.
Травматизм. Первичная и вторичная профилактика. Первая помощь при травмах.  грибы - презентация для начальной школы
грибы - презентация для начальной школы Презентация Лизинг
Презентация Лизинг Эпиграфы
Эпиграфы Expression language
Expression language Мотивация деятельности в менеджменте
Мотивация деятельности в менеджменте  Практика привлечения к субсидиарной ответственности
Практика привлечения к субсидиарной ответственности Заглавная буква в именах существительных - презентация для начальной школы_
Заглавная буква в именах существительных - презентация для начальной школы_ Asynchronous Transfer Mode Обзор технологии
Asynchronous Transfer Mode Обзор технологии Класс Земноводные
Класс Земноводные «Путешествие по страницам школьной жизни» Составитель: Иванова Марина Васильевна, учитель начальн
«Путешествие по страницам школьной жизни» Составитель: Иванова Марина Васильевна, учитель начальн Відмінювання числівників
Відмінювання числівників Программирование на языке Си
Программирование на языке Си Презентация Исполнительная власть в механизме государственной власти
Презентация Исполнительная власть в механизме государственной власти