- Инсулин, глюкагон, лептин, метаболический синдром

Содержание
- 2. Введение Инсулин - секретируется в кровь b-клетками островков Лангерганса поджелудочной железы. Главная функция инсулина - регуляция
- 3. Биосинтез инсулин млекопитающих кодируется одним геном (у некоторых видов - двумя), определяющим образование одноцепочечного крупного белка
- 4. Осуществлен полный хим. синтез инсулин человека, разработаны способы трансформации инсулина животных в инсулин человека, а также
- 5. Секреция инсулина
- 7. Важнейшие пути регуляции панкреатической секреции
- 8. Секреция инсулина контролируется изменениями концентраций циркулирующих в крови нутриентов (глюкозы, аминокислот, жирных кислот), гормонами желудочно-кишечного тракта,
- 9. Эффекты нутриентов на секреторную активность b-клеток поджелудочной железы являются результатом их прямого взаимодействия с клеточными мембранами
- 10. В состоянии покоя мембранный потенциал (МПП) на внутренней поверхности мембраны b-клеток равен -50-70мВ. В мембранах b-клеток
- 11. Образовавшийся АТФ вызывает закрытие АТФ-чувствительных калиевых каналов. Это предотвращает выход К+ из клетки, что является результатом
- 12. Ионы Са2+ обеспечивают секрецию инсулина из секреторных гранул несколькими путями: 1) Положительно заряженные ионы Са2+ облегчают
- 13. Ингибиторами секреции инсулина является вещества, активирующиеся симпатической системой: соматостатин, гормоны гипофиза (АКТГ, ГР, ТТГ, пролактин, вазопрессин),
- 14. Действие инсулина на клетки Инсулин активирует транспортеры глюкозы, которые переносят глюкозу в клетку. Инсулин отвечает за
- 15. Путь трансмембранного проведения гормонального сигнала - тирозинкиназный. Инициация действия инсулина обуславливается взаимодействием гормон-рецептор. Рецепторы инсулина относятся
- 16. В связи с анаболической функцией инсулина стимуляция симпатической нервной системы угнетает секрецию инсулина, а парасимпатической усиливает
- 17. Роль инсулина в регуляции обмена веществ выходит за рамки регуляции уровня глюкозы в крови. В мышечных
- 18. Синтез инсулиновых рецепторов находится под контролем гена INSR. Существует множество мутаций этого гена. Кроме того, транспорт
- 19. Поэтому в человеческой популяции существует большая доля лиц с врожденной резистентностью мышечной и жировой тканей к
- 20. Регуляция уровня глюкозы в крови Запасание Инсулин Соматотропин Расход запасов Глюкагон Адреналин Кортизол Тироксин Соматотропин (
- 21. Гипоталамус способен стимулировать выделение инсулина из b-клеток панкреатических островков по нервнопроводниковому пути, который назван «паравентрикуло-вагусным».. Этот
- 22. Глюкагон Стимулирует распад гликогена в печени до глюкозы и повышает содержание глюкозы в крови. Глюкагон способствует
- 23. 19-16
- 24. Реципрокные отношения инсулина с глюкагоном
- 26. Инсулин и глюкоза в регуляции центральной симпатической активности
- 27. Обмен глюкозы
- 28. Лептин и его роль в регуляции метаболизма и физиологических функций
- 29. ЛЕПТИН Действует как липостат ↑ запасы жира в жировой ткани ↑ увеличивается выход лептина в кровь
- 30. Лептин - гормон, секретируемый жировыми клетками, циркулирующий в крови в свободной и связанной формах. Содержание лептина
- 31. Лептин частично связывается белками плазмы. Связанная фракция лептина м. достигать 80% у человека при мутациях рецепторов
- 32. Концентрация лептина тесно коррелирует с индексом массы тела (ИМТ). Лептин регулирует чувство насыщения на уровне дугообразного
- 33. На уровень лептина влияет уменьшение количества жировой ткани (снижение веса тела на 10% приводит к 53%-му
- 34. Уровень лептина регулируется и другими факторами. Пик уровня глюкокортикоида совпадает с самым низким уровнем лептина в
- 35. И именно падение уровня сывороточного лептина в результате болезни или радикальных диет запускает компенсаторный механизм, снижающий
- 36. Основные действия лептина - повышение печеночного глюкогенолиза и захвата глюкозы скелетными мышцами; - повышение скорости липолиза
- 37. Действие лептина на липидный обмен осуществляется через стимуляцию экспрессии гена ПОМКосуществляется через стимуляцию экспрессии гена ПОМК
- 38. Первые сообщения о том, что меланокортины вовлечены в регуляцию пищевого поведенияПервые сообщения о том, что меланокортины
- 39. Таким образом. Потребление пищи зависят от двух типов эффекторных систем в ЦНС и гипоталамусе. Первая -
- 40. В организме существует как долговремен-ная, так и кратковременная система контроля пищевого поведения и расхода энергии. Концентрация
- 41. Содержание лептина в крови может служить сигналом, указывающим на готовность организма к выполнению репродуктивной функции и
- 42. Обратная корреляция между пульсирующей секрецией лептина и кортизола и адренокортикотропина (АСТН) известна у человека. Препубертальное увеличение
- 43. Вывод: нормальное функционирование рецептора лептина необходимо не только для регуляции веса тела, но также для регуляции
- 44. По современным представлениям лептин подает сигнал в гипоталамус через активацию специфического лептинового рецептора, что проявляется уменьшением
- 45. лептиновый рецептор (Ob-R) был выделен из мышиного хороидного (сосудистого) сплетения и был идентифицирован как представитель цитокинового
- 46. была подтверждена экспрессия Ob-Rb в ядрах гипоталамуса: аркуатном (дугообразном), дорсомедиальном, паравентрикулярном, вентромедиальном, а также латеральном. Эти
- 47. Таблица 3. Гипоталамические регуляторы пищевого поведения [1]
- 49. Лептин и Инсулин регулируют активность рецепторов пептидэргических нейронов
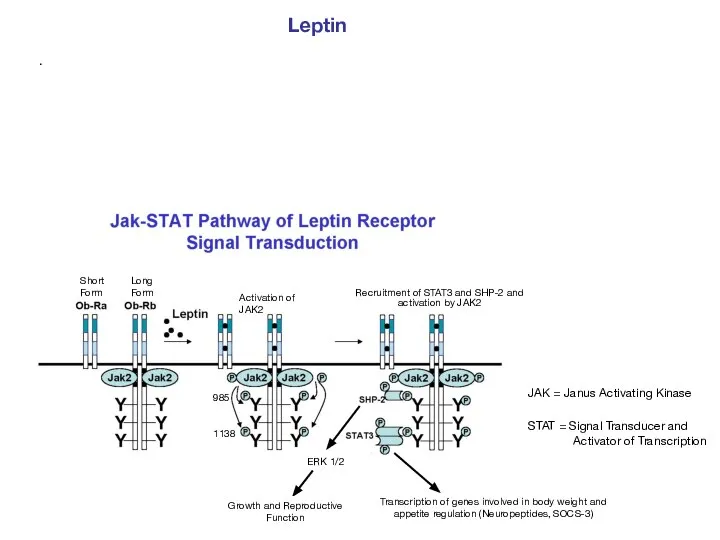
- 50. . Leptin JAK = Janus Activating Kinase STAT = Signal Transducer and Activator of Transcription Short
- 51. Intracellular Leptin Nucleus Extracellular DNA JAK Kinase JAK Kinase OB-R Leptin STAT STAT
- 52. Intracellular Leptin Lepn Extracellular STAT JAK Kinase JAK Kinase OB-R JAK Kinase JAK Kinase OB-R P
- 53. Intracellular Leptin Lepn Extracellular STAT STAT JAK Kinase JAK Kinase OB-R JAK Kinase JAK Kinase OB-R
- 54. Intracellular Leptin Lepn Extracellular STAT STAT JAK Kinase JAK Kinase OB-R JAK Kinase JAK Kinase OB-R
- 55. Intracellular Leptin Lepn Extracellular STAT STAT JAK Kinase JAK Kinase OB-R JAK Kinase JAK Kinase OB-R
- 56. Еще одним ярким примером нейроиммуноэндокринных взаимодействий является развитие сахарного диабета. Нередко у лиц среднего и пожилого
- 57. Метаболический синдром (основные дефиниции)
- 58. История вопроса В 1948 г. известный клиницист Е. М. Тареев писал: «Представление о гипертонике наиболее часто
- 59. Метаболический синдром (История вопроса) В 1988 г. G.Reaven высказал предположение об участии инсулинорезистентности и гиперинсулинемии в
- 60. МЕТАБОЛИЧЕСКИЙ X СИНДРОМ 1. ИНСУЛИНОРЕЗИСТЕНТНОСТЬ 2. ГИПЕРИНСУЛИНЕМИЯ 3. СНИЖЕНИЕ ТОЛЕРАНТНОСТИ К ГЛЮКОЗЕ 4. ДИСЛИПИДЕМИЯ 5. ОЖИРЕНИЕ
- 61. Метаболический синдром
- 62. Основные причины развития метаболического синдрома Метаболический синдром – причины: Снижение физической активности. Высокоуглеводный характер питания. Недостаток
- 63. Основными механизмами, приводящими к повышению АД при метаболическом синдроме, являются гиперволемия, обусловленная повышенной реабсорбцией натрия в
- 64. Патогенез МС (роль глюкозы) Повышение уровня глюкозы в крови сопровождается неферментативным образованием различных соединений глюкозы с
- 65. Прогрессирующее поражение почек при сахарном диабете CV Events Death Смерть
- 66. Упрощенные критерии инсулинорезистентности Национальным институтом здоровья США в 2001 году были сформулированы более упрощённые критерии, позволяющие
- 71. Скачать презентацию

Введение
Инсулин - секретируется в кровь b-клетками островков Лангерганса поджелудочной железы.
Главная
Введение
Инсулин - секретируется в кровь b-клетками островков Лангерганса поджелудочной железы.
Главная

Биосинтез инсулин млекопитающих кодируется одним геном (у некоторых видов - двумя),
Биосинтез инсулин млекопитающих кодируется одним геном (у некоторых видов - двумя),

Осуществлен полный хим. синтез инсулин человека, разработаны способы трансформации инсулина животных
Осуществлен полный хим. синтез инсулин человека, разработаны способы трансформации инсулина животных

Секреция инсулина
Секреция инсулина

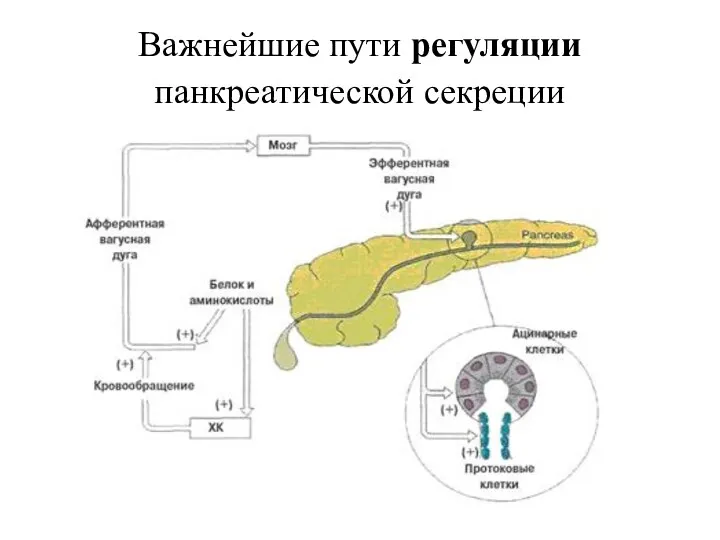
Важнейшие пути регуляции панкреатической секреции
Важнейшие пути регуляции панкреатической секреции

Секреция инсулина контролируется изменениями концентраций циркулирующих в крови нутриентов (глюкозы, аминокислот,
Секреция инсулина контролируется изменениями концентраций циркулирующих в крови нутриентов (глюкозы, аминокислот,

Эффекты нутриентов на секреторную активность b-клеток поджелудочной железы являются результатом их
Эффекты нутриентов на секреторную активность b-клеток поджелудочной железы являются результатом их

В состоянии покоя мембранный потенциал (МПП) на внутренней поверхности мембраны b-клеток
В состоянии покоя мембранный потенциал (МПП) на внутренней поверхности мембраны b-клеток

Образовавшийся АТФ вызывает закрытие АТФ-чувствительных калиевых каналов.
Это предотвращает выход К+
Образовавшийся АТФ вызывает закрытие АТФ-чувствительных калиевых каналов.
Это предотвращает выход К+

Ионы Са2+ обеспечивают секрецию инсулина из секреторных гранул несколькими путями:
1)
Ионы Са2+ обеспечивают секрецию инсулина из секреторных гранул несколькими путями:
1)

Ингибиторами секреции инсулина является вещества, активирующиеся симпатической системой: соматостатин, гормоны гипофиза
Ингибиторами секреции инсулина является вещества, активирующиеся симпатической системой: соматостатин, гормоны гипофиза

Действие инсулина на клетки
Инсулин активирует транспортеры глюкозы, которые переносят глюкозу в
Действие инсулина на клетки
Инсулин активирует транспортеры глюкозы, которые переносят глюкозу в

Путь трансмембранного проведения гормонального сигнала - тирозинкиназный.
Инициация действия инсулина обуславливается
Путь трансмембранного проведения гормонального сигнала - тирозинкиназный.
Инициация действия инсулина обуславливается

В связи с анаболической функцией инсулина стимуляция симпатической нервной системы угнетает
В связи с анаболической функцией инсулина стимуляция симпатической нервной системы угнетает

Роль инсулина в регуляции обмена веществ выходит за рамки регуляции уровня
Роль инсулина в регуляции обмена веществ выходит за рамки регуляции уровня

Синтез инсулиновых рецепторов находится под контролем гена INSR. Существует множество мутаций
Синтез инсулиновых рецепторов находится под контролем гена INSR. Существует множество мутаций

Поэтому в человеческой популяции существует большая доля лиц с врожденной резистентностью
Поэтому в человеческой популяции существует большая доля лиц с врожденной резистентностью

Регуляция уровня глюкозы в крови
Запасание
Инсулин
Соматотропин
Расход запасов
Глюкагон
Адреналин
Кортизол
Тироксин
Соматотропин
( длительная секреция)
Регуляция уровня глюкозы в крови
Запасание
Инсулин
Соматотропин
Расход запасов
Глюкагон
Адреналин
Кортизол
Тироксин
Соматотропин
( длительная секреция)

Гипоталамус способен стимулировать выделение инсулина из b-клеток панкреатических островков по нервнопроводниковому
Гипоталамус способен стимулировать выделение инсулина из b-клеток панкреатических островков по нервнопроводниковому

Глюкагон
Стимулирует распад гликогена в печени до глюкозы и повышает содержание глюкозы
Глюкагон
Стимулирует распад гликогена в печени до глюкозы и повышает содержание глюкозы
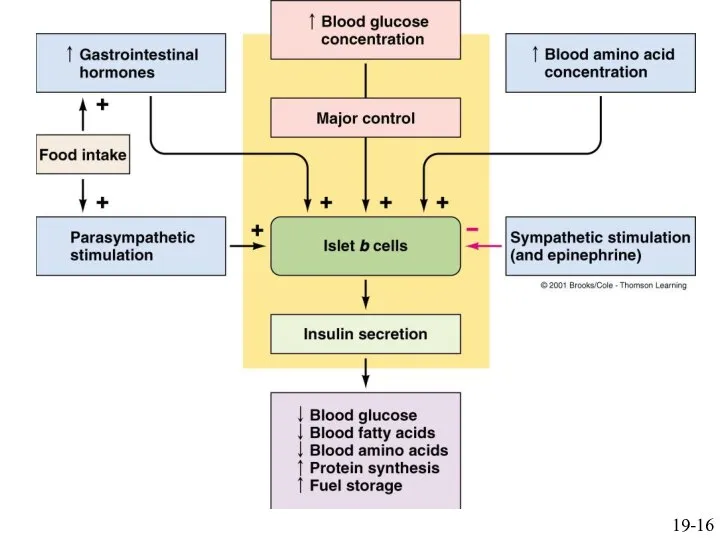
19-16
19-16

Реципрокные отношения инсулина с глюкагоном
Реципрокные отношения инсулина с глюкагоном

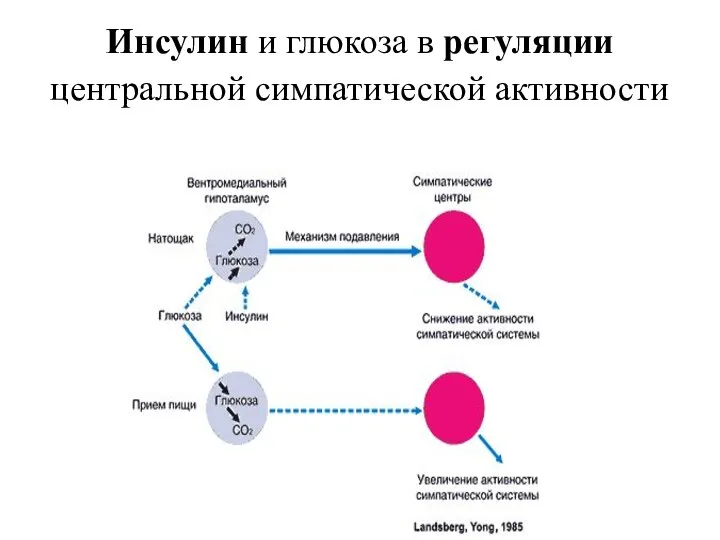
Инсулин и глюкоза в регуляции центральной симпатической активности
Инсулин и глюкоза в регуляции центральной симпатической активности

Обмен глюкозы
Обмен глюкозы

Лептин и его роль в регуляции метаболизма и физиологических функций
Лептин и его роль в регуляции метаболизма и физиологических функций
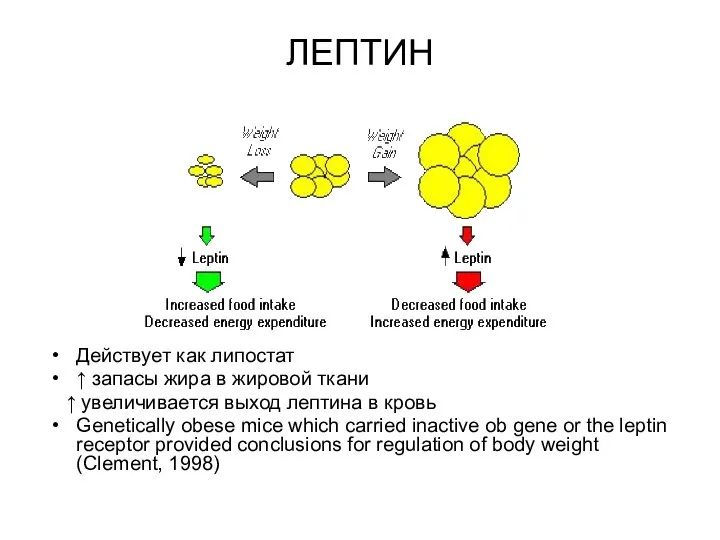
ЛЕПТИН
Действует как липостат
↑ запасы жира в жировой ткани
↑ увеличивается выход
ЛЕПТИН
Действует как липостат
↑ запасы жира в жировой ткани
↑ увеличивается выход

Лептин - гормон, секретируемый жировыми клетками, циркулирующий в крови в свободной
Лептин - гормон, секретируемый жировыми клетками, циркулирующий в крови в свободной

Лептин частично связывается белками плазмы. Связанная фракция лептина м. достигать 80%
Лептин частично связывается белками плазмы. Связанная фракция лептина м. достигать 80%

Концентрация лептина тесно коррелирует с индексом массы тела (ИМТ).
Лептин регулирует
Концентрация лептина тесно коррелирует с индексом массы тела (ИМТ).
Лептин регулирует

На уровень лептина влияет уменьшение количества жировой ткани (снижение веса тела
На уровень лептина влияет уменьшение количества жировой ткани (снижение веса тела

Уровень лептина регулируется и другими факторами.
Пик уровня глюкокортикоида совпадает с
Уровень лептина регулируется и другими факторами.
Пик уровня глюкокортикоида совпадает с

И именно падение уровня сывороточного лептина в результате болезни или радикальных
И именно падение уровня сывороточного лептина в результате болезни или радикальных

Основные действия лептина
- повышение печеночного глюкогенолиза и захвата глюкозы скелетными мышцами;
-
Основные действия лептина
- повышение печеночного глюкогенолиза и захвата глюкозы скелетными мышцами; -

Действие лептина на липидный обмен
осуществляется через стимуляцию экспрессии гена ПОМКосуществляется через
Действие лептина на липидный обмен
осуществляется через стимуляцию экспрессии гена ПОМКосуществляется через

Первые сообщения о том, что меланокортины вовлечены в регуляцию пищевого поведенияПервые
Первые сообщения о том, что меланокортины вовлечены в регуляцию пищевого поведенияПервые

Таким образом.
Потребление пищи зависят от двух типов эффекторных систем в ЦНС
Потребление пищи зависят от двух типов эффекторных систем в ЦНС

В организме существует как долговремен-ная, так и кратковременная система контроля пищевого
В организме существует как долговремен-ная, так и кратковременная система контроля пищевого

Содержание лептина в крови может служить сигналом, указывающим на готовность организма
Содержание лептина в крови может служить сигналом, указывающим на готовность организма

Обратная корреляция между пульсирующей секрецией лептина и кортизола и адренокортикотропина (АСТН)
Обратная корреляция между пульсирующей секрецией лептина и кортизола и адренокортикотропина (АСТН)

Вывод: нормальное функционирование рецептора лептина необходимо не только для регуляции веса
Вывод: нормальное функционирование рецептора лептина необходимо не только для регуляции веса

По современным представлениям лептин подает сигнал в гипоталамус через активацию специфического
По современным представлениям лептин подает сигнал в гипоталамус через активацию специфического

лептиновый рецептор (Ob-R) был выделен из мышиного хороидного (сосудистого) сплетения и
лептиновый рецептор (Ob-R) был выделен из мышиного хороидного (сосудистого) сплетения и

была подтверждена экспрессия Ob-Rb в ядрах гипоталамуса: аркуатном (дугообразном), дорсомедиальном, паравентрикулярном,
была подтверждена экспрессия Ob-Rb в ядрах гипоталамуса: аркуатном (дугообразном), дорсомедиальном, паравентрикулярном,
![Таблица 3. Гипоталамические регуляторы пищевого поведения [1]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1298554/slide-46.jpg)
Таблица 3. Гипоталамические регуляторы пищевого поведения [1]
Таблица 3. Гипоталамические регуляторы пищевого поведения [1]


Лептин и Инсулин регулируют активность рецепторов пептидэргических нейронов
Лептин и Инсулин регулируют активность рецепторов пептидэргических нейронов
.
Leptin
JAK = Janus Activating Kinase
STAT = Signal Transducer and Activator
.
Leptin
JAK = Janus Activating Kinase
STAT = Signal Transducer and Activator
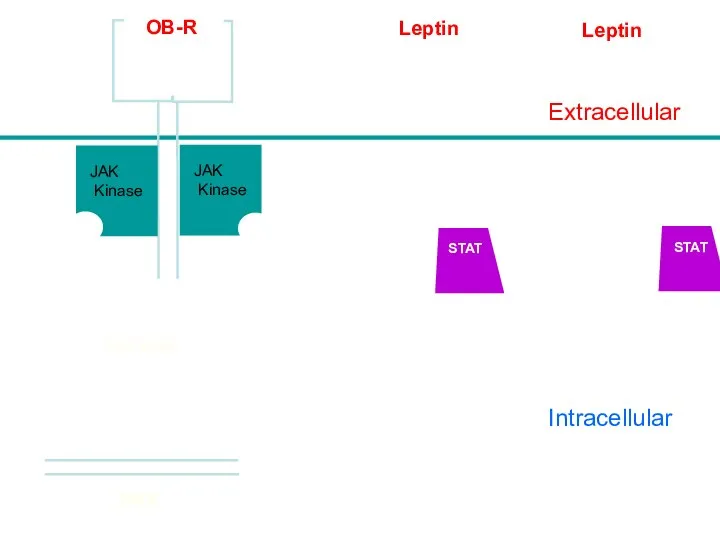
Intracellular
Leptin
Nucleus
Extracellular
DNA
JAK
Kinase
JAK
Kinase
OB-R
Leptin
STAT
STAT
Intracellular
Leptin
Nucleus
Extracellular
DNA
JAK
Kinase
JAK
Kinase
OB-R
Leptin
STAT
STAT

Intracellular
Leptin
Lepn
Extracellular
STAT
JAK
Kinase
JAK
Kinase
OB-R
JAK
Kinase
JAK
Kinase
OB-R
P
P
P
P
Leptin
Leptin
Leptin
STAT
Nucleus
DNA
Intracellular
Leptin
Lepn
Extracellular
STAT
JAK
Kinase
JAK
Kinase
OB-R
JAK
Kinase
JAK
Kinase
OB-R
P
P
P
P
Leptin
Leptin
Leptin
STAT
Nucleus
DNA
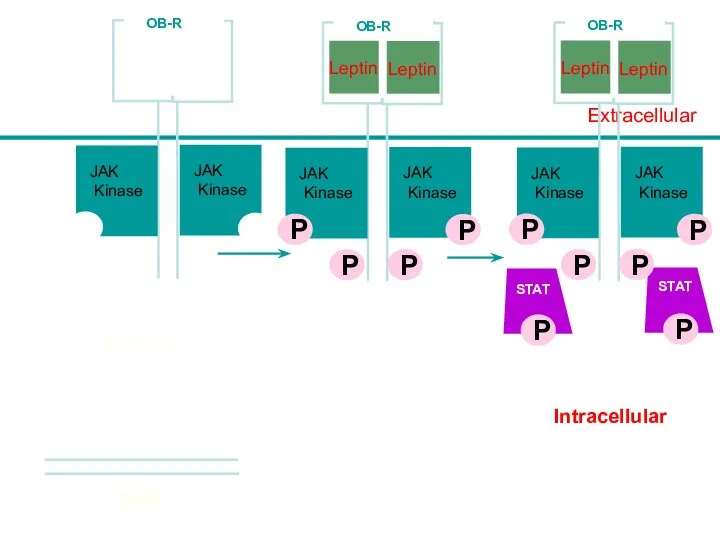
Intracellular
Leptin
Lepn
Extracellular
STAT
STAT
JAK
Kinase
JAK
Kinase
OB-R
JAK
Kinase
JAK
Kinase
OB-R
P
P
P
P
Leptin
Leptin
Leptin
Lepn
JAK
Kinase
JAK
Kinase
OB-R
P
P
P
P
Leptin
Leptin
P
P
Nucleus
DNA
Intracellular
Leptin
Lepn
Extracellular
STAT
STAT
JAK
Kinase
JAK
Kinase
OB-R
JAK
Kinase
JAK
Kinase
OB-R
P
P
P
P
Leptin
Leptin
Leptin
Lepn
JAK
Kinase
JAK
Kinase
OB-R
P
P
P
P
Leptin
Leptin
P
P
Nucleus
DNA

Intracellular
Leptin
Lepn
Extracellular
STAT
STAT
JAK
Kinase
JAK
Kinase
OB-R
JAK
Kinase
JAK
Kinase
OB-R
P
P
P
P
Leptin
Leptin
Leptin
Lepn
JAK
Kinase
JAK
Kinase
OB-R
P
P
P
P
Leptin
Leptin
P
P
STAT
P
STAT
P
Nucleus
DNA
Intracellular
Leptin
Lepn
Extracellular
STAT
STAT
JAK
Kinase
JAK
Kinase
OB-R
JAK
Kinase
JAK
Kinase
OB-R
P
P
P
P
Leptin
Leptin
Leptin
Lepn
JAK
Kinase
JAK
Kinase
OB-R
P
P
P
P
Leptin
Leptin
P
P
STAT
P
STAT
P
Nucleus
DNA
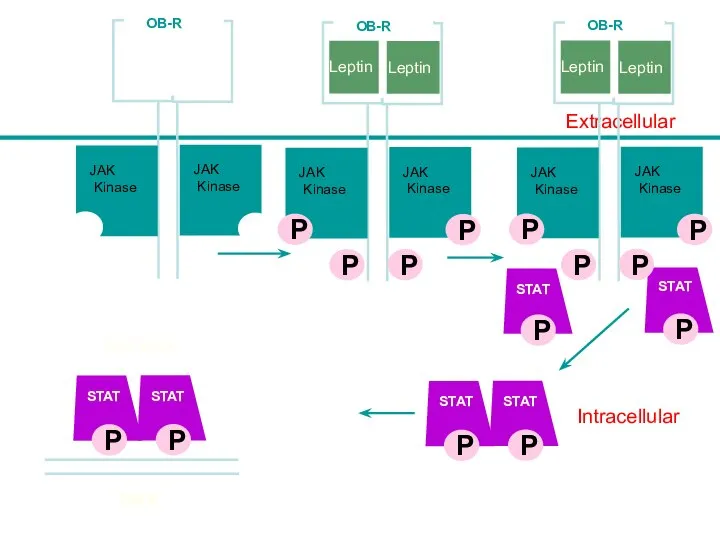
Intracellular
Leptin
Lepn
Extracellular
STAT
STAT
JAK
Kinase
JAK
Kinase
OB-R
JAK
Kinase
JAK
Kinase
OB-R
P
P
P
P
Leptin
Leptin
Leptin
Lepn
JAK
Kinase
JAK
Kinase
OB-R
P
P
P
P
Leptin
Leptin
P
P
STAT
P
STAT
P
Nucleus
DNA
STAT
P
STAT
P
Intracellular
Leptin
Lepn
Extracellular
STAT
STAT
JAK
Kinase
JAK
Kinase
OB-R
JAK
Kinase
JAK
Kinase
OB-R
P
P
P
P
Leptin
Leptin
Leptin
Lepn
JAK
Kinase
JAK
Kinase
OB-R
P
P
P
P
Leptin
Leptin
P
P
STAT
P
STAT
P
Nucleus
DNA
STAT
P
STAT
P

Еще одним ярким примером нейроиммуноэндокринных взаимодействий является развитие сахарного диабета. Нередко
Еще одним ярким примером нейроиммуноэндокринных взаимодействий является развитие сахарного диабета. Нередко

Метаболический синдром (основные дефиниции)
Метаболический синдром (основные дефиниции)

История вопроса
В 1948 г. известный клиницист Е. М. Тареев писал: «Представление
История вопроса
В 1948 г. известный клиницист Е. М. Тареев писал: «Представление

Метаболический синдром
(История вопроса)
В 1988 г. G.Reaven высказал предположение об участии инсулинорезистентности
Метаболический синдром
(История вопроса)
В 1988 г. G.Reaven высказал предположение об участии инсулинорезистентности

МЕТАБОЛИЧЕСКИЙ X СИНДРОМ
1. ИНСУЛИНОРЕЗИСТЕНТНОСТЬ
2. ГИПЕРИНСУЛИНЕМИЯ
3. СНИЖЕНИЕ ТОЛЕРАНТНОСТИ К ГЛЮКОЗЕ
МЕТАБОЛИЧЕСКИЙ X СИНДРОМ
1. ИНСУЛИНОРЕЗИСТЕНТНОСТЬ 2. ГИПЕРИНСУЛИНЕМИЯ 3. СНИЖЕНИЕ ТОЛЕРАНТНОСТИ К ГЛЮКОЗЕ

Метаболический синдром
Метаболический синдром

Основные причины развития метаболического синдрома
Метаболический синдром – причины:
Снижение физической активности.
Основные причины развития метаболического синдрома
Метаболический синдром – причины:
Снижение физической активности.

Основными механизмами, приводящими к повышению АД при метаболическом синдроме, являются гиперволемия,
Основными механизмами, приводящими к повышению АД при метаболическом синдроме, являются гиперволемия,

Патогенез МС (роль глюкозы)
Повышение уровня глюкозы в крови сопровождается неферментативным образованием
Патогенез МС (роль глюкозы)
Повышение уровня глюкозы в крови сопровождается неферментативным образованием
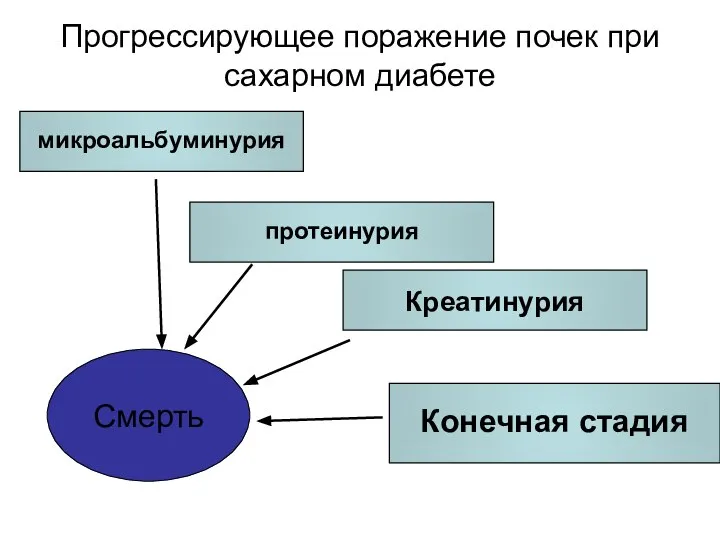
Прогрессирующее поражение почек при сахарном диабете
CV Events
Death
Смерть
Прогрессирующее поражение почек при сахарном диабете
CV Events
Death
Смерть

Упрощенные критерии инсулинорезистентности
Национальным институтом здоровья США в 2001 году были
Упрощенные критерии инсулинорезистентности
Национальным институтом здоровья США в 2001 году были


 Сани (для дошкольников)
Сани (для дошкольников) ИНФОРМАЦИОННЫЕ ТЕХНОЛОГИИ В ДОКАЗАТЕЛЬНОЙ МЕДИЦИНЕ ОСНОВНЫЕ ПОНЯТИЯ ДОКАЗАТЕЛЬНОЙ РАДИОЛОГИИ
ИНФОРМАЦИОННЫЕ ТЕХНОЛОГИИ В ДОКАЗАТЕЛЬНОЙ МЕДИЦИНЕ ОСНОВНЫЕ ПОНЯТИЯ ДОКАЗАТЕЛЬНОЙ РАДИОЛОГИИ Создание сайта самостоятельно (02)
Создание сайта самостоятельно (02) Легковые электромобили в США
Легковые электромобили в США  Акустические и огнезащитные материалы
Акустические и огнезащитные материалы Тема: Редкие животные, занесенные в красную книгу. Исполнитель: Прудникова Александра, ученица 3б класса МОУ СОШ № 31 Г. Новочерка
Тема: Редкие животные, занесенные в красную книгу. Исполнитель: Прудникова Александра, ученица 3б класса МОУ СОШ № 31 Г. Новочерка Аттестационная работа. Методическая разработка в рамках межшкольного взаимодействия «Интерактивный этномузей»
Аттестационная работа. Методическая разработка в рамках межшкольного взаимодействия «Интерактивный этномузей» Заимствованные слова
Заимствованные слова Биография Г.Х.Андерсена. Веб-квест,,В гостях у сказок’’
Биография Г.Х.Андерсена. Веб-квест,,В гостях у сказок’’ Электрические машины. Трёхфазные трансформаторы. (Лекция 7)
Электрические машины. Трёхфазные трансформаторы. (Лекция 7) Процессуальное право. Административное юрисдикция и конституционное судопроизводство
Процессуальное право. Административное юрисдикция и конституционное судопроизводство Машиностроительный комплекс: роль, значение, проблемы развития, факторы размещения. Тема урока
Машиностроительный комплекс: роль, значение, проблемы развития, факторы размещения. Тема урока Нормативно-правовые документы, обеспечивающие функционирование КСЗИ
Нормативно-правовые документы, обеспечивающие функционирование КСЗИ Россия и США. Игра престолов. Англосаксонский мир
Россия и США. Игра престолов. Англосаксонский мир Тема 4 Основы и социальная сущность бизнеса
Тема 4 Основы и социальная сущность бизнеса  ТОЛЕРАНТНОСТЬ ИНТОЛЕРАНТНОСТЬ социальный педагог ОПСОП по Куйбышевскому району ЦРО г.о. Самара Мищенкова Лариса Петровна
ТОЛЕРАНТНОСТЬ ИНТОЛЕРАНТНОСТЬ социальный педагог ОПСОП по Куйбышевскому району ЦРО г.о. Самара Мищенкова Лариса Петровна Презентация "О перспективах освоения газовых ресурсов Восточной Сибири и Дальнего Востока" - скачать презентации по Экономик
Презентация "О перспективах освоения газовых ресурсов Восточной Сибири и Дальнего Востока" - скачать презентации по Экономик Металлические пломбировочные материалы (амальгама)
Металлические пломбировочные материалы (амальгама)  ТОВАРНЫЙ АССОРТИМЕНТ И ТОВАРНЫЕ ЗНАКИ Выполнила работу: студент группы 52 н Мусихин Антон
ТОВАРНЫЙ АССОРТИМЕНТ И ТОВАРНЫЕ ЗНАКИ Выполнила работу: студент группы 52 н Мусихин Антон Технология игры
Технология игры Банки
Банки  Основные понятия и средства языка программирования
Основные понятия и средства языка программирования Тест дизайн
Тест дизайн Формирование пооперационного контроля у учащихся 2 класса при освоении обобщенного способа умножения и деления многозначного чис
Формирование пооперационного контроля у учащихся 2 класса при освоении обобщенного способа умножения и деления многозначного чис Подбор сечений деревянных элементов сплошного сечения
Подбор сечений деревянных элементов сплошного сечения Схема станции М
Схема станции М Мода 40-х годов ХХ века. Дресс-код танцевальной ретро-площадки
Мода 40-х годов ХХ века. Дресс-код танцевальной ретро-площадки Презентация Поворот во внешней политике Советского Союза – курс на прекращение «Холодной войны»
Презентация Поворот во внешней политике Советского Союза – курс на прекращение «Холодной войны»