- Липиды и обмен липидов

Содержание
- 4. ФУНКЦИИ ЛИПИДОВ 1. Энергетическая (1 г липидов при окислении дает 9,3 ккал; 1 моль пальмитиновой кислоты
- 5. Функции липидов Структурная: холестерол и фосфолипиды – структурные компоненты мембран. Метаболическая: холестерол образует большое число биоактивных
- 6. Функции липидов Регуляторная: стероидные гормоны, фосфатидилинозитол и его производные: диацилглицерол и инозитолфосфат (вторичные мессенджеры гормонов); производные
- 7. Патология липидного обмена Первичные нарушения: Гиперлипопротеинемия, дислипопротеинемия (гиперхолестеринемия: атеросклероз) Ожирение Желчнокаменная болезнь Метаболический ацидоз (кетонемия) Вторичные
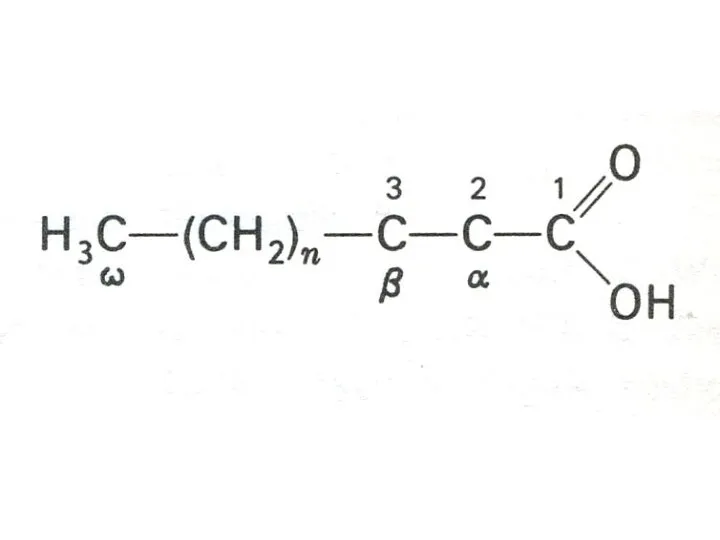
- 8. Жирные кислоты > 70 жирных кислот идентифицированы в организме. Свободные жирные кислоты находятся в основном в
- 9. ЖИРНЫЕ КИСЛОТЫ Галогеновое число определяет число двойных связей в жирной кислоте. У человека возможен синтез только
- 10. Производные ненасыщенных жирных кислот - ЭЙКОЗАНОИДЫ Семейства простагландинов, тромбоксанов, лейкотриенов др. - биологически активные вещества, действуют
- 11. ПРОСТАГЛАНДИНЫ Образуются при участии циклооксигеназы из арахидоновой кислоты (С20:4). Ингибируется аспирином по механизму ковалентной модификации фермента
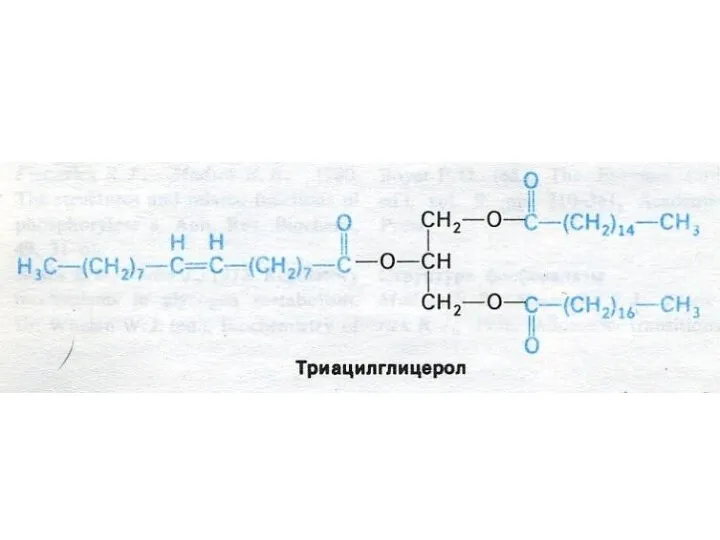
- 12. Внешний обмен липидов Потребность в жирах – 80 -100 г/сут (возрастает при необходимости перехода энергетического обмена
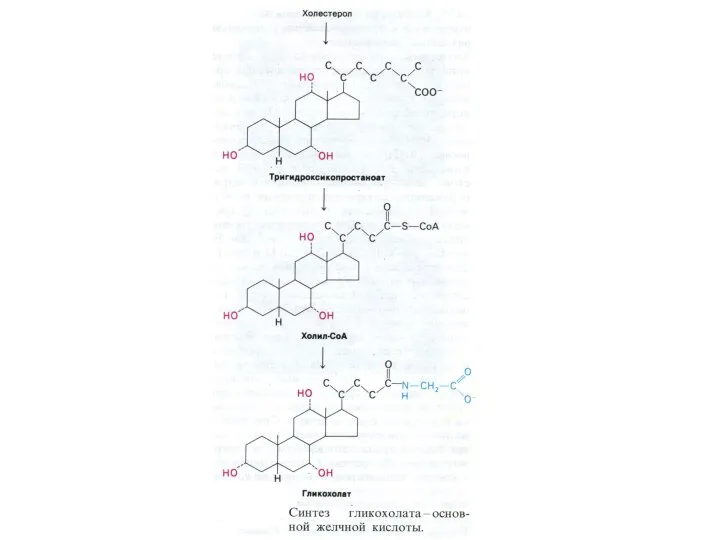
- 13. ВНЕШНИЙ ОБМЕН ЛИПИДОВ Основные эмульгаторы жиров в ЖКТ – желчные кислоты – производные холестерола, но и
- 17. ВНЕШНИЙ ОБМЕН ЛИПИДОВ Гидролиз нейтральных жиров в 12 –п кишечнике: моно- , ди- и триглицеридлипазы: продукты
- 18. ВСАСЫВАНИЕ ПРОДУКТОВ ГИДРОЛИЗА В ЖКТ Мелкие эмульгированные капли жира (до 0,5 мкм) проникают через кишечную стенку
- 19. Ресинтез жиров в энтероцитах На основе продуктов гидролиза экзогенных жиров в энтероцитах синтезируются видоспецифичные липиды. Чаще
- 20. Транспортные формы экзогенных липидов Продукты ресинтеза жиров (ТАГ, ЭХЛ. ФЛ) в энтероцитах образуют комплекс с Апо
- 21. Кругооборот эндогенных липидов Липиды, синтезирующиеся в печени (эндогенные) транспортируются в крови в составе ЛПОНП (Апо В-100).
- 22. Промежуточный обмен липидов Внутриклеточный липолиз: Адипоциты. Постабсорбтивный период, голодание, физическая нагрузка. ТАГ-липаза. Активна в фосфорилированном состоянии.
- 23. Промежуточный обмен липидов Жирные кислоты транспортируются в крови в комплексе с альбумином. Проникая в клетки, жирные
- 24. Метаболизм глицерина Глицерин образуется как продукт липолиза или возникает при восстановлении избытка диоксиацетонфосфата (метаболит гликолиза). Глицерин
- 26. Окисление жирных кислот Печень, миокард, скелетные мышцы более 50% энергии получают при окислении жирных кислот. Ленинджер,
- 28. Активация и транспорт жирных кислот На наружной мембране митохондрий ацил- Коа синтетаза : R- СООН +
- 31. Механизм реакций β −окисления жирных кислот. Ступенчатый процесс последовательно повторяющихся 4-х реакций. С участием ФАД- и
- 34. Особенности окисления жирных кислот: С нечетным числом атомов: 3-х углеродный остаток (пропионил-КоА) карбоксилируется до сукцинил-КоА, поступающий
- 37. Энергетика окисления жирных кислот Каждый этап β –окисления сопровождается образованием ФАДН2 и НАДН (реокисление их в
- 38. КЕТОГЕНЕЗ Избыточное образование ацетил-КоА или снижение его утилизации в ЦТК (причины!) приводит к активации кетогенеза в
- 40. Окисление кетоновых тел Печень не способна утилизировать кетоновые тела. В периферических тканях гидроксибутират окисляется до ацетоацетата.
- 42. Липогенез Абсорбтивный период. Инсулин. Ресинтез собственных жиров на основе продуктов гидролиза экзогенного жира в энтероцитах (этерификация
- 43. ЛИПОГЕНЕЗ Субстраты: глицерол-3-фосфат (образуется глицеролкиназой в энтероцитах и нефроцитах из глицерина; из диоксиацетонфосфата (НАДФН зависимая дегидрогеназа)
- 44. Липогенез Избыток глюкозы обеспечивает: 1. запас гликогена 2. ДАФ ?глицерол-3-фосфат 3. ПВК ? ацетил-КоА ? жирные
- 45. Биосинтез жирных кислот Цитозоль. Ацилсинтетаза (пальмитоил-синтетаза). Не обращение β – окисления! ацетил – КоА АТФ, НАДФН
- 47. Транспорт ацетил-КоА в цитоплазму В ситуации накопления АТФ и НАДН ингибируется изоцитрат ДГ и накапливающийся цитрат
- 48. Транспорт ацетил-КоА в цитоплазму Возвращение ОА в МХ: 1.ОА –> малат (НАД зависимая цитоплазматическая МДГ); Малат
- 50. Образование малонил-КоА ацетил-КоА- карбоксилаза (биотин-зависимая) – регуляторный фермент синтеза жирных кислот. активная форма –нефосфорилирована (в присутствии
- 51. Синтетаза жирных кислот мультиферментный комплекс (гомодимер, поэтому синтезируется две цепи одновременно). 6 ферментов и АПБ (2
- 53. АПБ – структура и функция Активный центр этого белка – фосфопантотеновая кислота и тиоэтиламин (аналогичные структуре
- 56. Пальмитоил- синтетаза Ацетил-КоА + 7 малонил-КоА + 14 НАДФН ?C16:0 + 7 СО2 +SH-АПБ + 14
- 57. Синтез триацилглицеролов Ацил-КоА- синтетаза активирует жирные кислоты с затратой АТФ через стадию образования ацил-аденилатов. Перенос ацилтрансферазой
- 60. Синтез фосфолипидов Синтез ТАГ и ФЛ конкурируют за общие субстраты для собственного синтеза (фосфатидная кислота). Синтез
- 65. Синтез сфинголипидов
- 67. Синтез холестерола 1.Конденсация 3-х ацетил-КоА ?гидроксиметилглутарил –КоА 2. восстановление НАДФН-редуктазой? мевалоновая кислота 3. фосфорилирование и декарбоксилирование?
- 68. Метаболизм холестерола 0,5 г эндогенного и 0,5 г экзогенного ХЛ 10% ХЛ находится в виде эфиров
- 75. Скачать презентацию

ФУНКЦИИ ЛИПИДОВ
1. Энергетическая (1 г липидов при окислении дает 9,3 ккал;
ФУНКЦИИ ЛИПИДОВ
1. Энергетическая (1 г липидов при окислении дает 9,3 ккал;

Функции липидов
Структурная: холестерол и фосфолипиды – структурные компоненты мембран.
Метаболическая: холестерол
Функции липидов
Структурная: холестерол и фосфолипиды – структурные компоненты мембран.
Метаболическая: холестерол

Функции липидов
Регуляторная: стероидные гормоны, фосфатидилинозитол и его производные: диацилглицерол и инозитолфосфат
Функции липидов
Регуляторная: стероидные гормоны, фосфатидилинозитол и его производные: диацилглицерол и инозитолфосфат

Патология липидного обмена
Первичные нарушения:
Гиперлипопротеинемия, дислипопротеинемия (гиперхолестеринемия: атеросклероз)
Ожирение
Желчнокаменная болезнь
Метаболический ацидоз (кетонемия)
Вторичные гиперлипопротеинемии
Патология липидного обмена
Первичные нарушения:
Гиперлипопротеинемия, дислипопротеинемия (гиперхолестеринемия: атеросклероз)
Ожирение
Желчнокаменная болезнь
Метаболический ацидоз (кетонемия)
Вторичные гиперлипопротеинемии

Жирные кислоты
> 70 жирных кислот идентифицированы в организме.
Свободные жирные кислоты находятся
Жирные кислоты
> 70 жирных кислот идентифицированы в организме.
Свободные жирные кислоты находятся

ЖИРНЫЕ КИСЛОТЫ
Галогеновое число определяет число двойных связей в жирной кислоте. У
ЖИРНЫЕ КИСЛОТЫ
Галогеновое число определяет число двойных связей в жирной кислоте. У

Производные ненасыщенных жирных кислот - ЭЙКОЗАНОИДЫ
Семейства простагландинов, тромбоксанов, лейкотриенов др. -
Производные ненасыщенных жирных кислот - ЭЙКОЗАНОИДЫ
Семейства простагландинов, тромбоксанов, лейкотриенов др. -

ПРОСТАГЛАНДИНЫ
Образуются при участии циклооксигеназы из арахидоновой кислоты (С20:4).
Ингибируется аспирином по механизму
ПРОСТАГЛАНДИНЫ
Образуются при участии циклооксигеназы из арахидоновой кислоты (С20:4).
Ингибируется аспирином по механизму

Внешний обмен липидов
Потребность в жирах – 80 -100 г/сут (возрастает при
Внешний обмен липидов
Потребность в жирах – 80 -100 г/сут (возрастает при

ВНЕШНИЙ ОБМЕН ЛИПИДОВ
Основные эмульгаторы жиров в ЖКТ – желчные кислоты –
ВНЕШНИЙ ОБМЕН ЛИПИДОВ
Основные эмульгаторы жиров в ЖКТ – желчные кислоты –
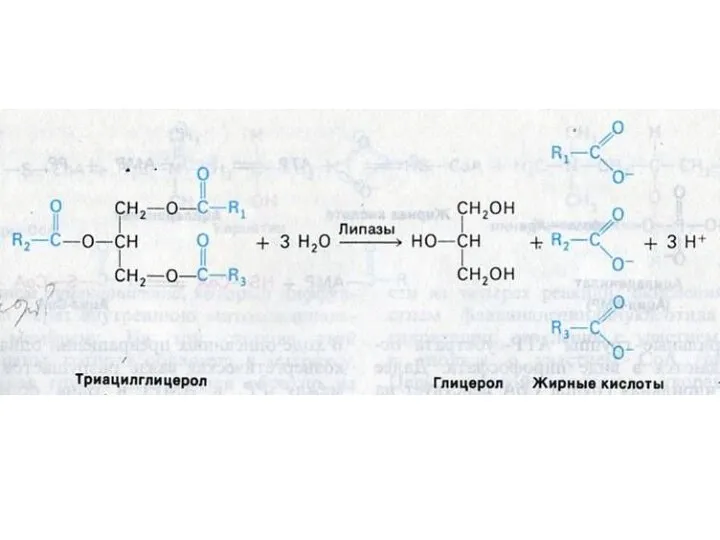
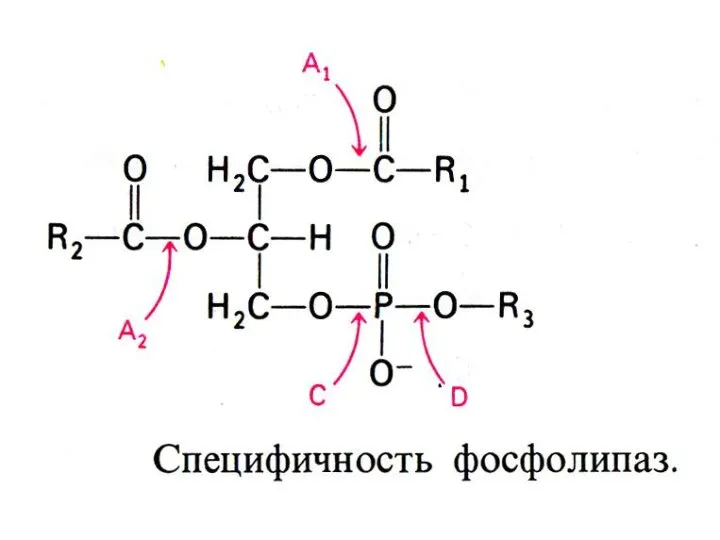

ВНЕШНИЙ ОБМЕН ЛИПИДОВ
Гидролиз нейтральных жиров в 12 –п кишечнике: моно- ,
ВНЕШНИЙ ОБМЕН ЛИПИДОВ
Гидролиз нейтральных жиров в 12 –п кишечнике: моно- ,

ВСАСЫВАНИЕ ПРОДУКТОВ ГИДРОЛИЗА В ЖКТ
Мелкие эмульгированные капли жира (до 0,5 мкм)
ВСАСЫВАНИЕ ПРОДУКТОВ ГИДРОЛИЗА В ЖКТ
Мелкие эмульгированные капли жира (до 0,5 мкм)

Ресинтез жиров в энтероцитах
На основе продуктов гидролиза экзогенных жиров в энтероцитах
Ресинтез жиров в энтероцитах
На основе продуктов гидролиза экзогенных жиров в энтероцитах

Транспортные формы экзогенных липидов
Продукты ресинтеза жиров (ТАГ, ЭХЛ. ФЛ) в энтероцитах
Транспортные формы экзогенных липидов
Продукты ресинтеза жиров (ТАГ, ЭХЛ. ФЛ) в энтероцитах

Кругооборот эндогенных липидов
Липиды, синтезирующиеся в печени (эндогенные) транспортируются в крови
Кругооборот эндогенных липидов
Липиды, синтезирующиеся в печени (эндогенные) транспортируются в крови

Промежуточный обмен липидов
Внутриклеточный липолиз:
Адипоциты. Постабсорбтивный период, голодание, физическая нагрузка.
ТАГ-липаза. Активна в
Промежуточный обмен липидов
Внутриклеточный липолиз:
Адипоциты. Постабсорбтивный период, голодание, физическая нагрузка.
ТАГ-липаза. Активна в

Промежуточный обмен липидов
Жирные кислоты транспортируются в крови в комплексе с альбумином.
Проникая
Промежуточный обмен липидов
Жирные кислоты транспортируются в крови в комплексе с альбумином.
Проникая

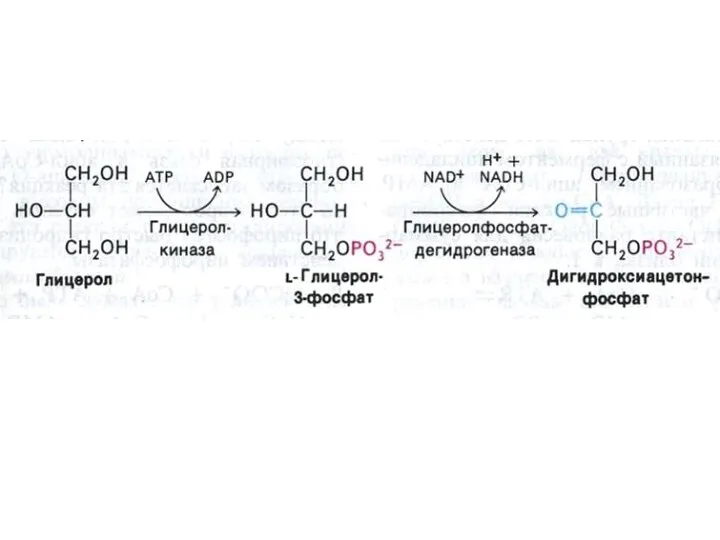
Метаболизм глицерина
Глицерин образуется как продукт липолиза или возникает при восстановлении избытка
Метаболизм глицерина
Глицерин образуется как продукт липолиза или возникает при восстановлении избытка

Окисление жирных кислот
Печень, миокард, скелетные мышцы более 50% энергии получают при
Окисление жирных кислот
Печень, миокард, скелетные мышцы более 50% энергии получают при

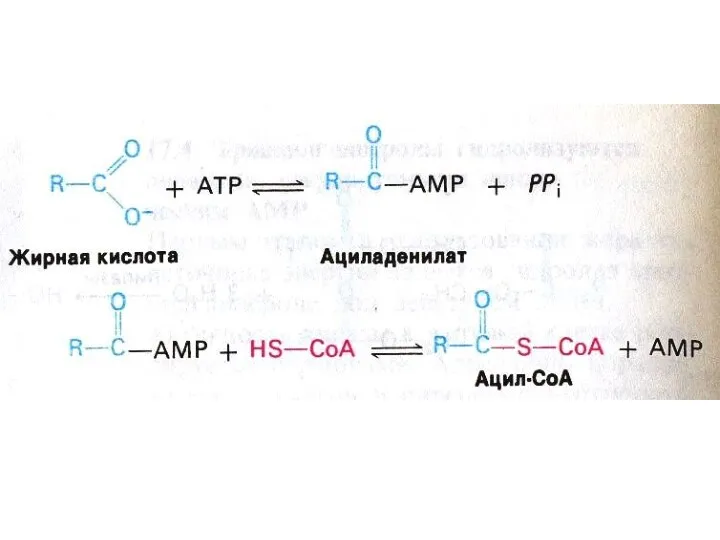
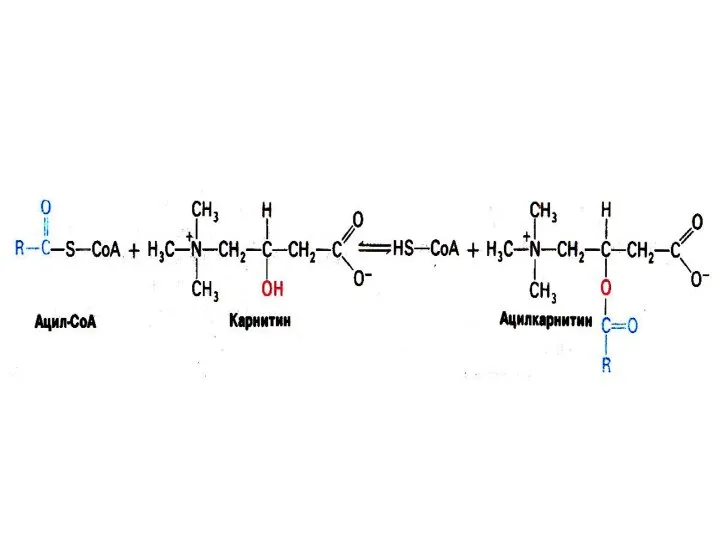
Активация и транспорт жирных кислот
На наружной мембране митохондрий ацил- Коа синтетаза
Активация и транспорт жирных кислот
На наружной мембране митохондрий ацил- Коа синтетаза

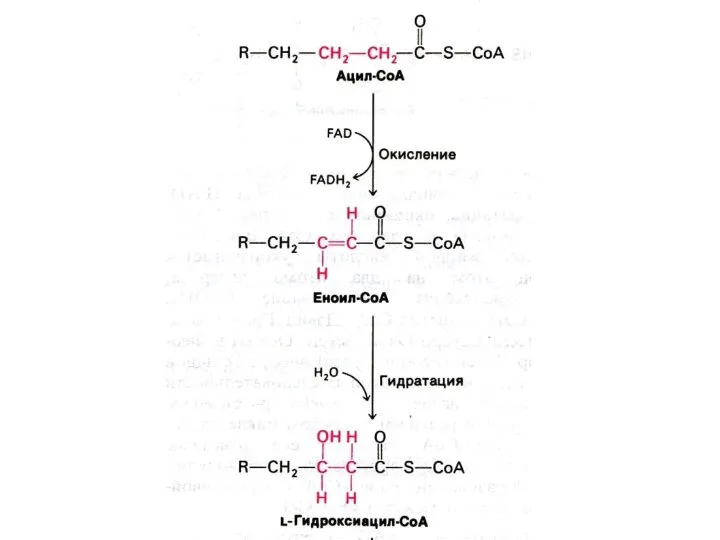
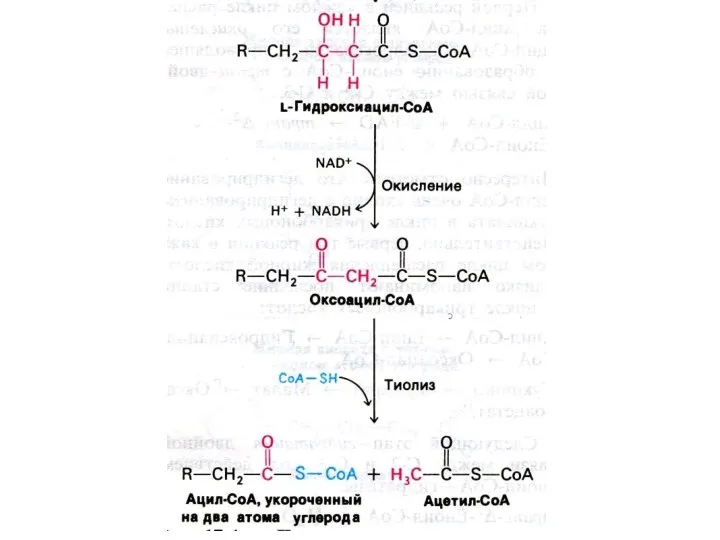
Механизм реакций β −окисления жирных кислот.
Ступенчатый процесс последовательно повторяющихся 4-х
Механизм реакций β −окисления жирных кислот.
Ступенчатый процесс последовательно повторяющихся 4-х

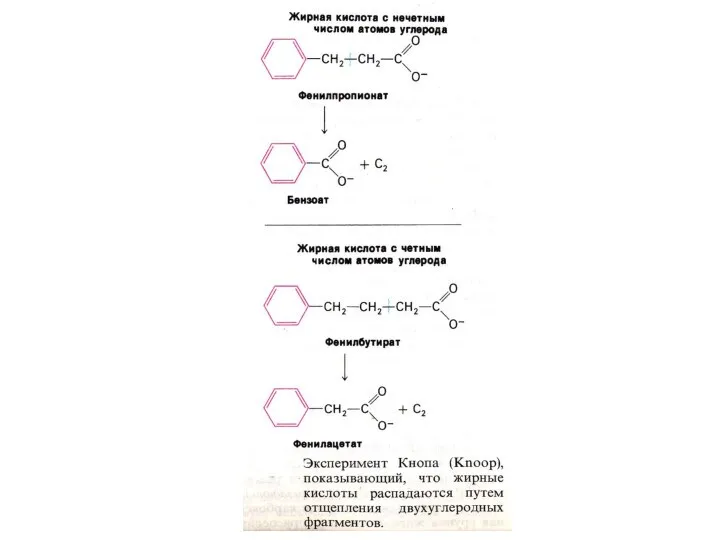
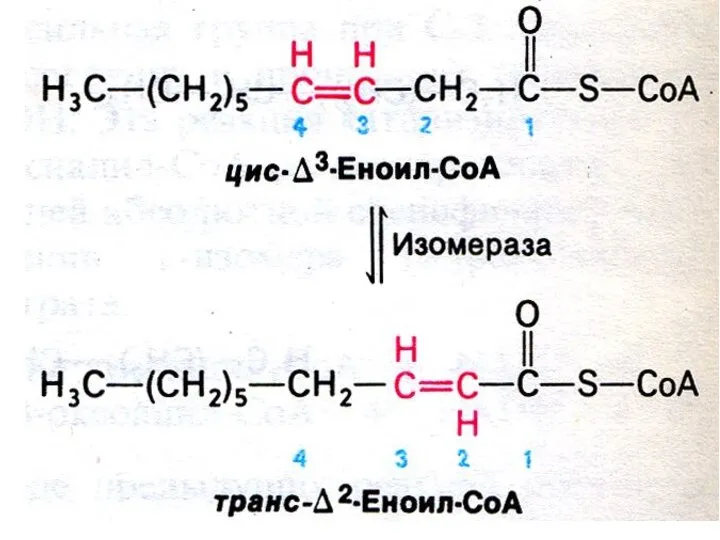
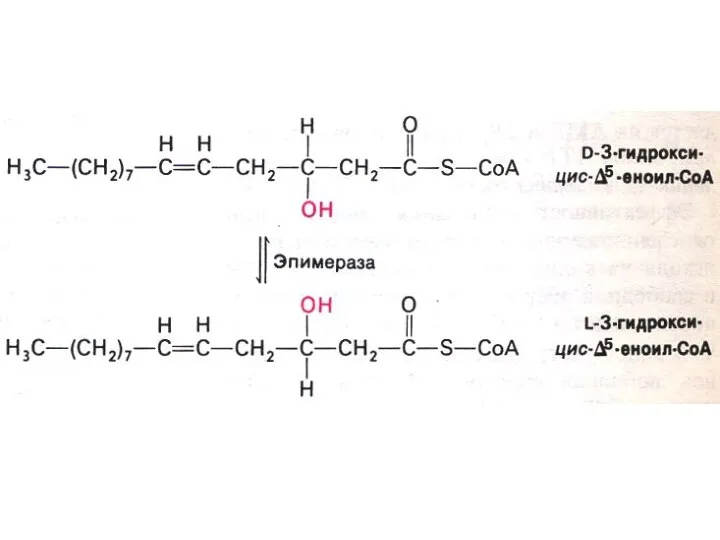
Особенности окисления жирных кислот:
С нечетным числом атомов:
3-х углеродный остаток (пропионил-КоА) карбоксилируется
Особенности окисления жирных кислот:
С нечетным числом атомов:
3-х углеродный остаток (пропионил-КоА) карбоксилируется

Энергетика окисления жирных кислот
Каждый этап β –окисления сопровождается образованием ФАДН2 и
Энергетика окисления жирных кислот
Каждый этап β –окисления сопровождается образованием ФАДН2 и

КЕТОГЕНЕЗ
Избыточное образование ацетил-КоА или снижение его утилизации в ЦТК (причины!) приводит
КЕТОГЕНЕЗ
Избыточное образование ацетил-КоА или снижение его утилизации в ЦТК (причины!) приводит
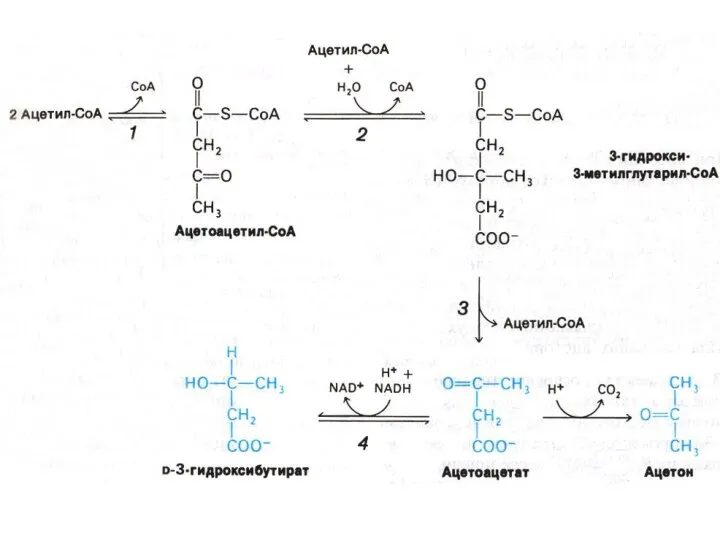

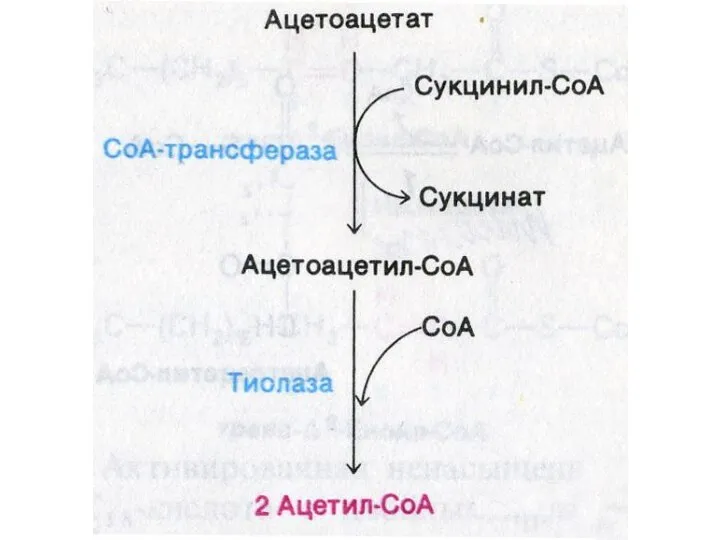
Окисление кетоновых тел
Печень не способна утилизировать кетоновые тела.
В периферических тканях
Окисление кетоновых тел
Печень не способна утилизировать кетоновые тела.
В периферических тканях

Липогенез
Абсорбтивный период. Инсулин.
Ресинтез собственных жиров на основе продуктов гидролиза экзогенного жира
Липогенез
Абсорбтивный период. Инсулин.
Ресинтез собственных жиров на основе продуктов гидролиза экзогенного жира

ЛИПОГЕНЕЗ
Субстраты:
глицерол-3-фосфат (образуется глицеролкиназой в энтероцитах и нефроцитах из глицерина; из диоксиацетонфосфата
ЛИПОГЕНЕЗ
Субстраты:
глицерол-3-фосфат (образуется глицеролкиназой в энтероцитах и нефроцитах из глицерина; из диоксиацетонфосфата

Липогенез
Избыток глюкозы обеспечивает:
1. запас гликогена
2. ДАФ ?глицерол-3-фосфат
3. ПВК ? ацетил-КоА ?
Липогенез
Избыток глюкозы обеспечивает:
1. запас гликогена
2. ДАФ ?глицерол-3-фосфат
3. ПВК ? ацетил-КоА ?

Биосинтез жирных кислот
Цитозоль. Ацилсинтетаза (пальмитоил-синтетаза). Не обращение β – окисления!
ацетил –
Биосинтез жирных кислот
Цитозоль. Ацилсинтетаза (пальмитоил-синтетаза). Не обращение β – окисления!
ацетил –
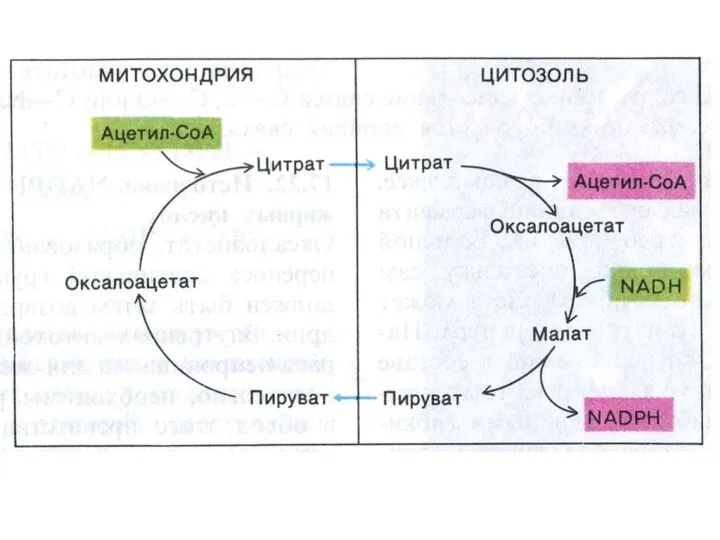

Транспорт ацетил-КоА в цитоплазму
В ситуации накопления АТФ и НАДН ингибируется изоцитрат
Транспорт ацетил-КоА в цитоплазму
В ситуации накопления АТФ и НАДН ингибируется изоцитрат

Транспорт ацетил-КоА в цитоплазму
Возвращение ОА в МХ:
1.ОА –> малат (НАД
Транспорт ацетил-КоА в цитоплазму
Возвращение ОА в МХ:
1.ОА –> малат (НАД
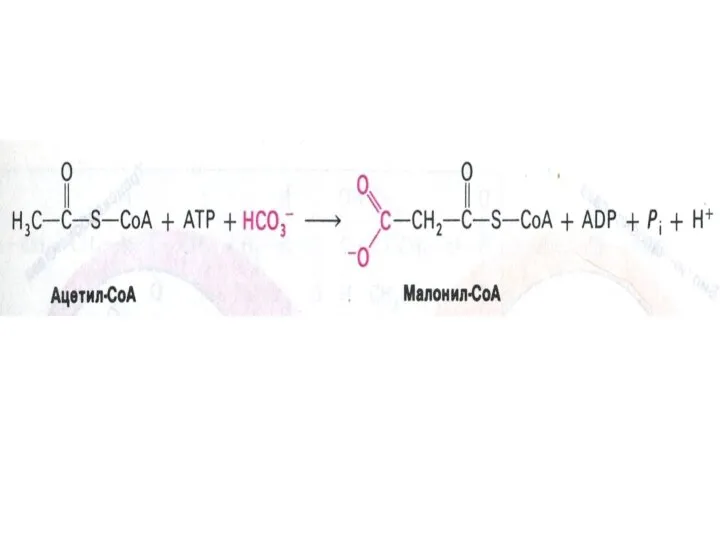

Образование малонил-КоА
ацетил-КоА- карбоксилаза (биотин-зависимая) – регуляторный фермент синтеза жирных кислот.
активная
Образование малонил-КоА
ацетил-КоА- карбоксилаза (биотин-зависимая) – регуляторный фермент синтеза жирных кислот.
активная

Синтетаза жирных кислот
мультиферментный комплекс (гомодимер, поэтому синтезируется две цепи одновременно).
6 ферментов
Синтетаза жирных кислот
мультиферментный комплекс (гомодимер, поэтому синтезируется две цепи одновременно).
6 ферментов
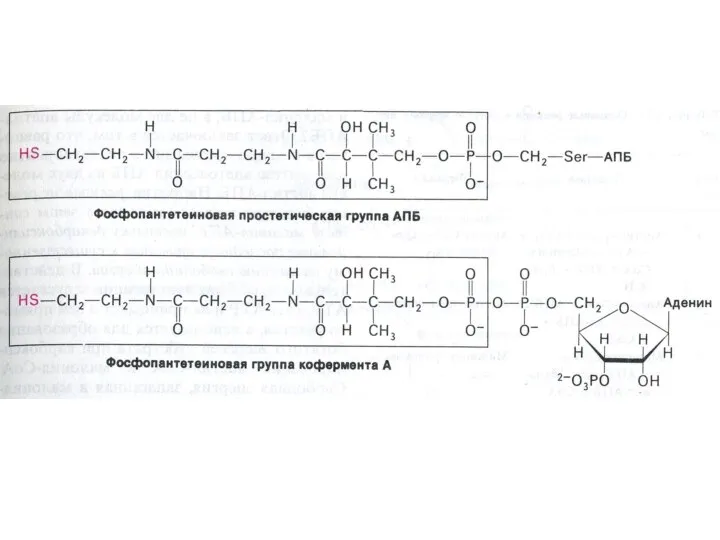

АПБ – структура и функция
Активный центр этого белка – фосфопантотеновая кислота
АПБ – структура и функция
Активный центр этого белка – фосфопантотеновая кислота
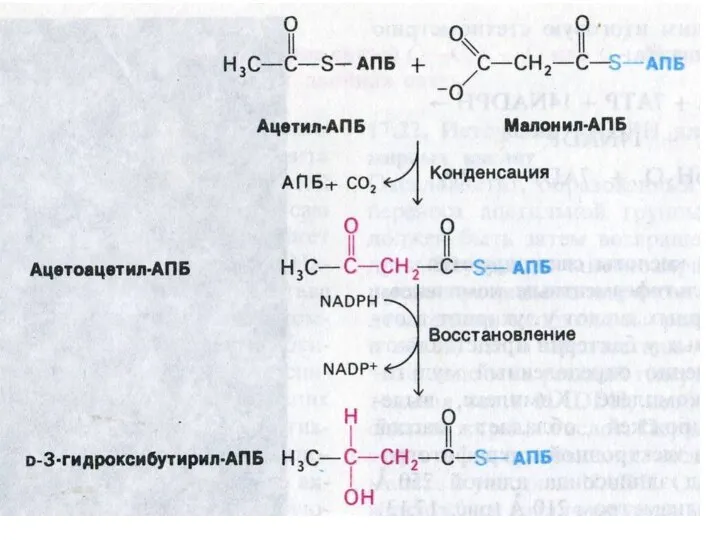
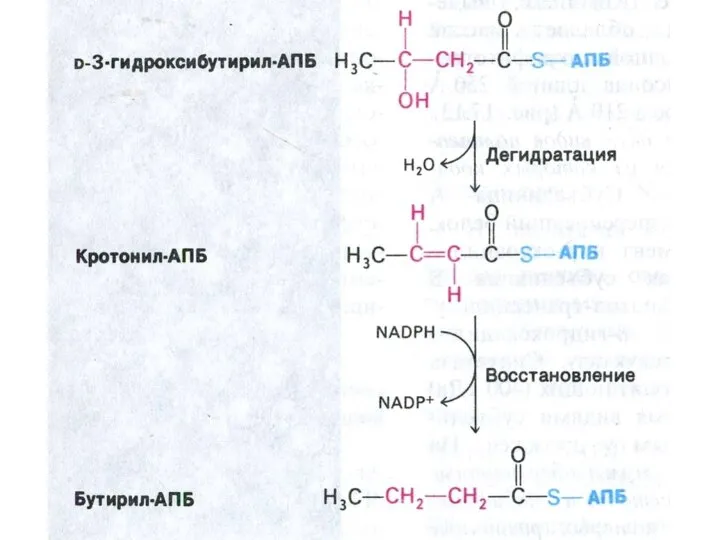

Пальмитоил- синтетаза
Ацетил-КоА + 7 малонил-КоА + 14 НАДФН ?C16:0 + 7
Пальмитоил- синтетаза
Ацетил-КоА + 7 малонил-КоА + 14 НАДФН ?C16:0 + 7

Синтез триацилглицеролов
Ацил-КоА- синтетаза активирует жирные кислоты с затратой АТФ через стадию
Синтез триацилглицеролов
Ацил-КоА- синтетаза активирует жирные кислоты с затратой АТФ через стадию
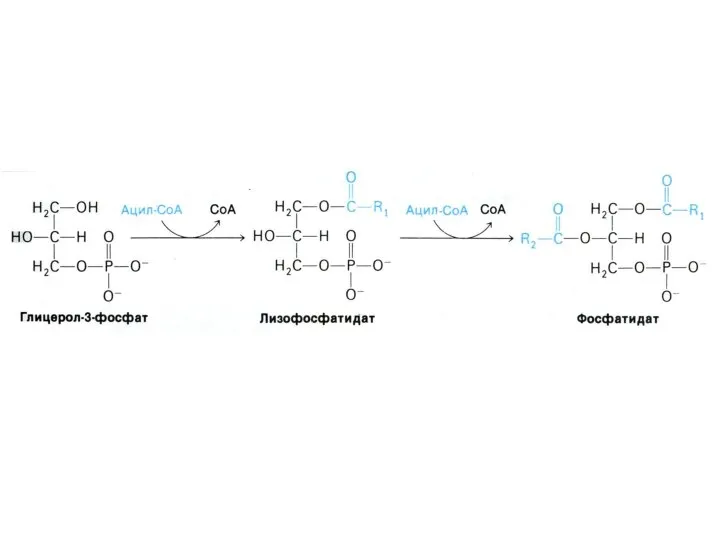
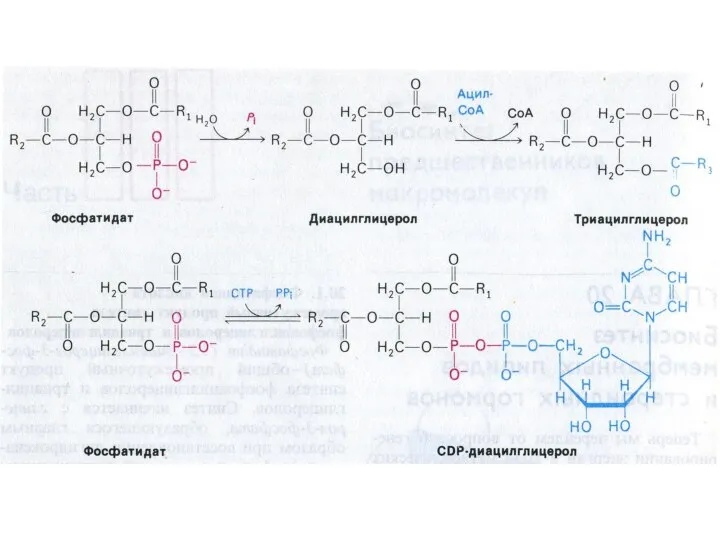

Синтез фосфолипидов
Синтез ТАГ и ФЛ конкурируют за общие субстраты для собственного
Синтез фосфолипидов
Синтез ТАГ и ФЛ конкурируют за общие субстраты для собственного
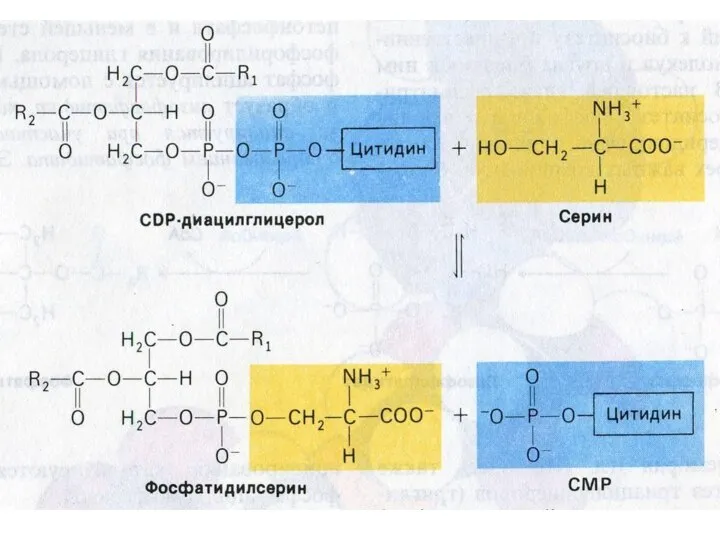
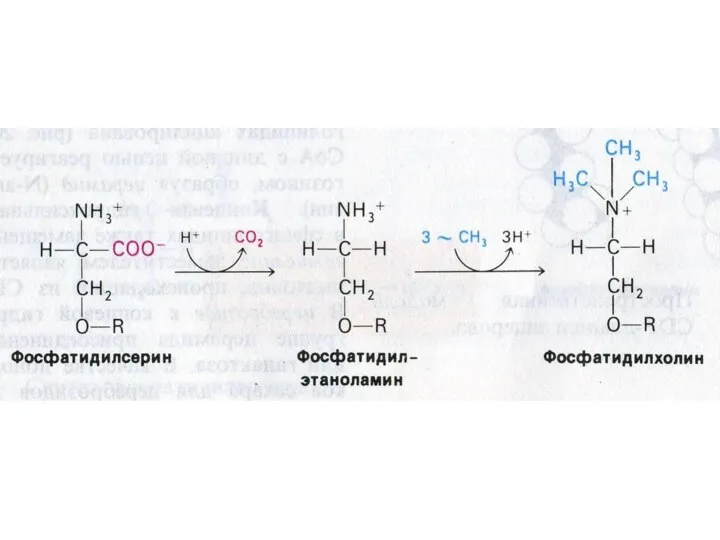
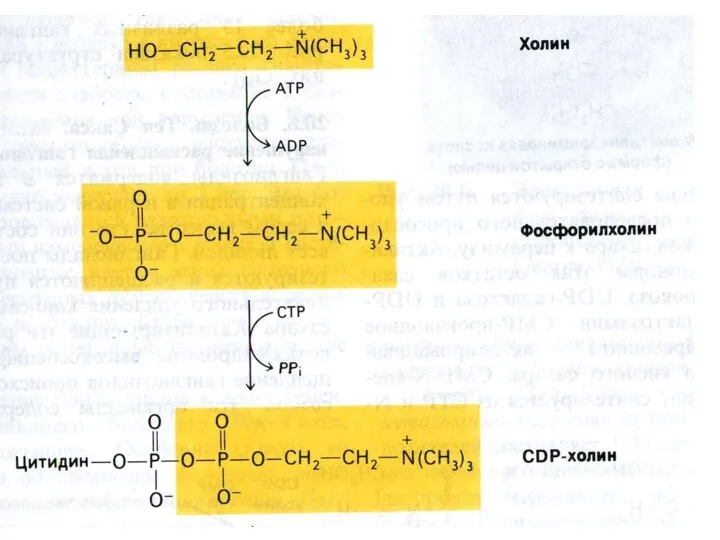
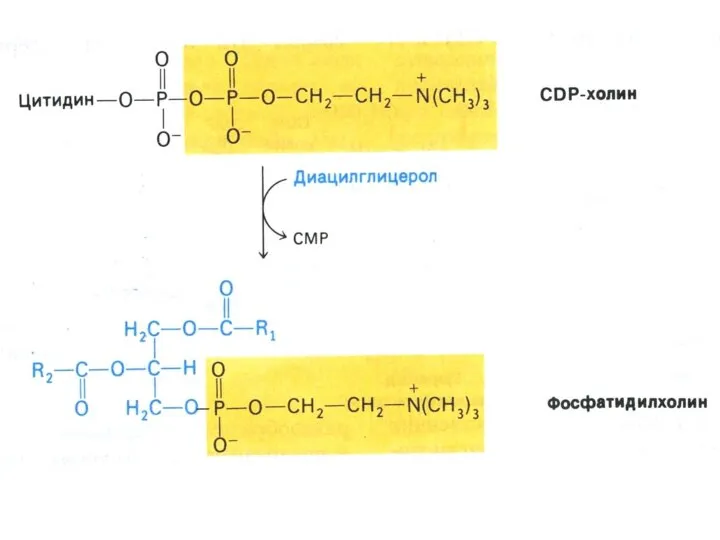

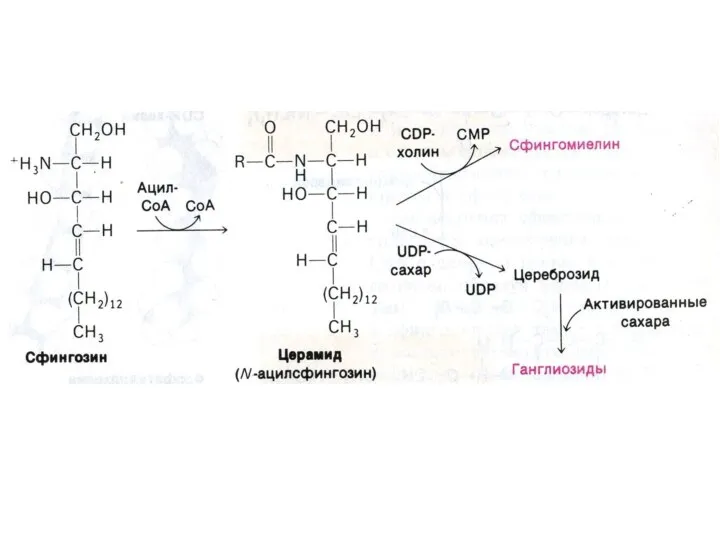
Синтез сфинголипидов
Синтез сфинголипидов

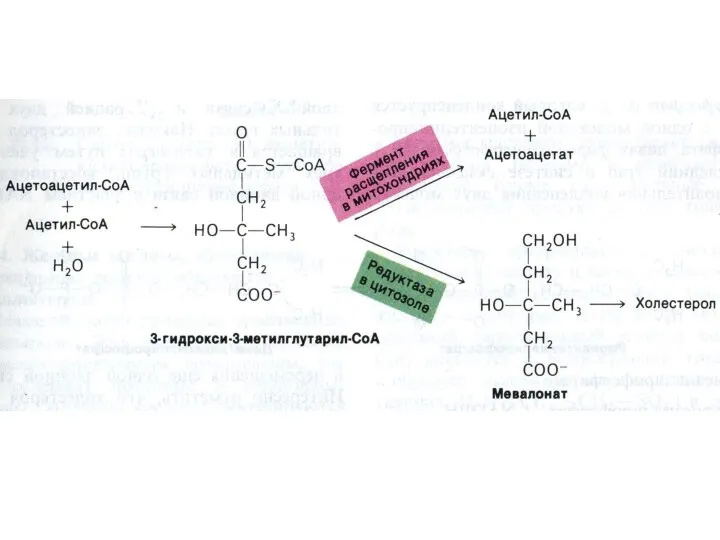
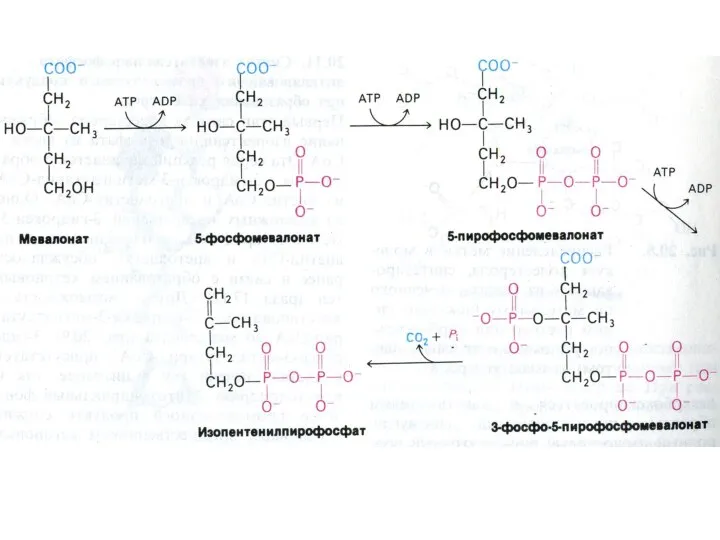
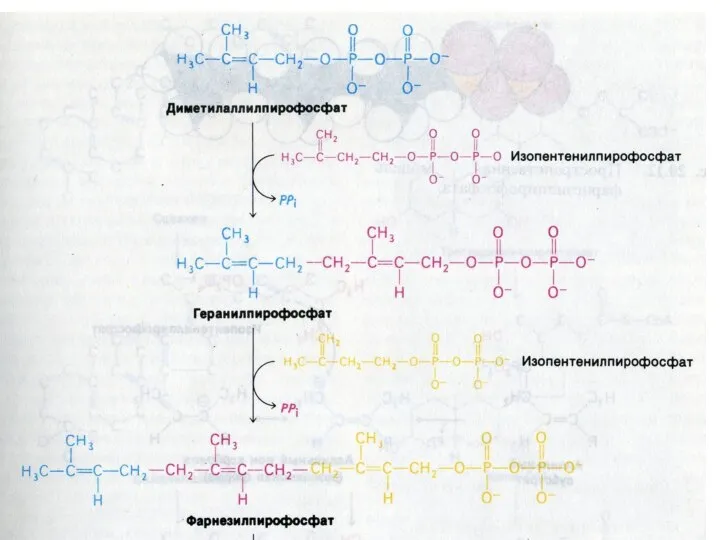
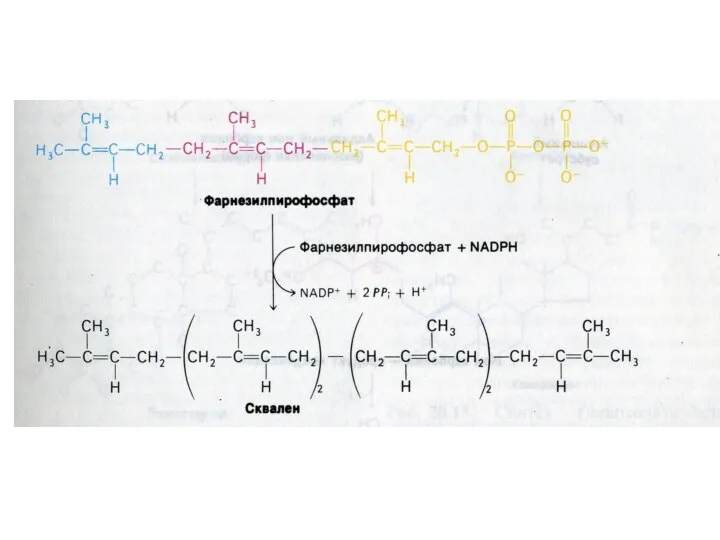
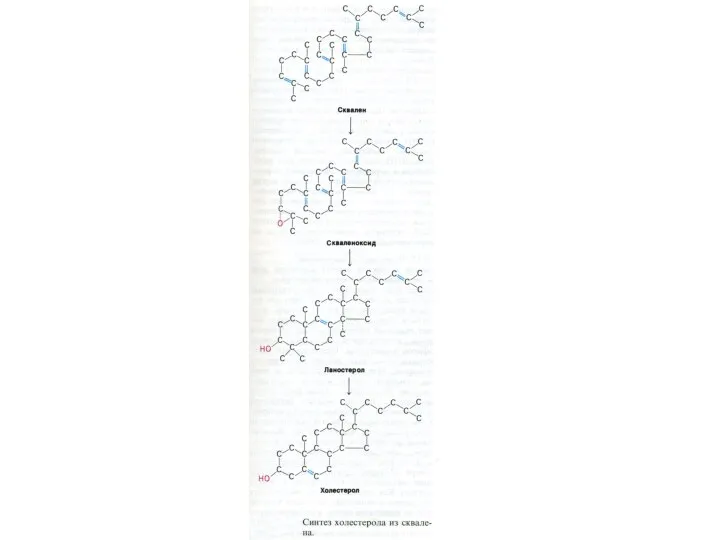
Синтез холестерола
1.Конденсация 3-х ацетил-КоА ?гидроксиметилглутарил –КоА
2. восстановление НАДФН-редуктазой? мевалоновая кислота
3.
Синтез холестерола
1.Конденсация 3-х ацетил-КоА ?гидроксиметилглутарил –КоА
2. восстановление НАДФН-редуктазой? мевалоновая кислота
3.

Метаболизм холестерола
0,5 г эндогенного и 0,5 г экзогенного ХЛ
10% ХЛ находится
Метаболизм холестерола
0,5 г эндогенного и 0,5 г экзогенного ХЛ
10% ХЛ находится
 Умственная работоспособность студентов и физическая культура
Умственная работоспособность студентов и физическая культура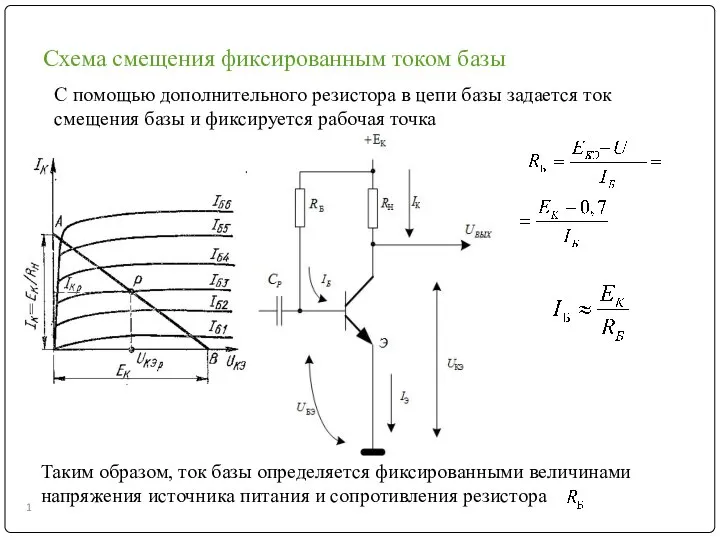 Схема смещения фиксированным током базы. (Лекция 7)
Схема смещения фиксированным током базы. (Лекция 7) Електоральні уподобання українців. Дослідження громадської думки. Березень 2019
Електоральні уподобання українців. Дослідження громадської думки. Березень 2019 Приемы запоминания материала к ЕГЭ по истории
Приемы запоминания материала к ЕГЭ по истории Инвестиционный проект «Комплексное развитие Нижнего Приангарья» О ХОДЕ РЕАЛИЗАЦИИ И ФИНАНСИРОВАНИИ
Инвестиционный проект «Комплексное развитие Нижнего Приангарья» О ХОДЕ РЕАЛИЗАЦИИ И ФИНАНСИРОВАНИИ Интересные факты о LINUX
Интересные факты о LINUX Разработка планировочного решения в реконструкции квартир
Разработка планировочного решения в реконструкции квартир Программирование ЖК дисплея
Программирование ЖК дисплея Мой папа - презентация для начальной школы
Мой папа - презентация для начальной школы Аренда. Договор аренды. Лизинг и его виды
Аренда. Договор аренды. Лизинг и его виды Модель машины
Модель машины Презентация на тему "РЕПРОДУКТИВНОЕ ЗДОРОВЬЕ" - скачать презентации по Медицине
Презентация на тему "РЕПРОДУКТИВНОЕ ЗДОРОВЬЕ" - скачать презентации по Медицине Функция График функции. 7 класс.
Функция График функции. 7 класс.  У истоков зодчества. Интеграционный проект учебных предметов биологии, географии, истории и химии.
У истоков зодчества. Интеграционный проект учебных предметов биологии, географии, истории и химии. Персональный компьютер
Персональный компьютер Термодинамика окислительно-восстановительных процессов
Термодинамика окислительно-восстановительных процессов  Проект на тему: Легко ли быть предпринимателем Выполнила: студентка 021 группы специальности «Коммерция» Созонтова Виктория Влади
Проект на тему: Легко ли быть предпринимателем Выполнила: студентка 021 группы специальности «Коммерция» Созонтова Виктория Влади Дорога без опасности - презентация для начальной школы
Дорога без опасности - презентация для начальной школы СНК
СНК Архитектоника
Архитектоника ВКР: Повышение производительности зерноуборочного комбайна «ACROS-595Plus»
ВКР: Повышение производительности зерноуборочного комбайна «ACROS-595Plus» УРОК РУССКОГО ЯЗЫКА в 1 классе
УРОК РУССКОГО ЯЗЫКА в 1 классе Достижение стратегических целей национального проекта «Образование»
Достижение стратегических целей национального проекта «Образование» Законы Хаммурапи 1792
Законы Хаммурапи 1792 Типовые конструкции изделия и узлы зданий и сооружений. Серия 1.020
Типовые конструкции изделия и узлы зданий и сооружений. Серия 1.020 Правосознание и правовая культура
Правосознание и правовая культура Письма Минстроя России о решениях общего собрания собственников помещений в многоквартирном доме
Письма Минстроя России о решениях общего собрания собственников помещений в многоквартирном доме Евлогий Александрийский, богослов, полемист
Евлогий Александрийский, богослов, полемист