- Обмен нуклеотидов Матричные биосинтезы

Содержание
- 2. Актуальность темы Нуклеотиды и их производные выполняют многообразные функции в организме человека: участвуют в синтезе нуклеиновых
- 3. План лекции Образование фосфорибозилдифосфата (ФРДФ) – ключевой момент в синтезе нуклеотидов Синтез и катаболизм пуриновых нуклеотидов:
- 4. Цель лекции Знать: Основные метаболические пути превращения пуриновых и пиримидиновых нуклеотидов Химико-биологическую сущность процессов репликации, транскрипции,
- 5. Вспомните самостоятельно из курса химии, используя слайды 6-14 Пуриновые и пиримидиновые азотистые основания Структура пуриновых и
- 6. Строение нуклеиновых кислот Функция: хранение, передача, реализация наследственной информации Нуклеиновые кислоты (НК) - биополимеры Мономер –
- 7. Пентозы в структуре нуклеиновых кислот
- 8. Первичная структура НК: последовательность нуклеотидов Химические связи: 1 - 5′-фосфоэфирная 2 – N-гликозидная 3 - 3′,5′
- 9. Вторичная структура ДНК: двойная спираль Правозакрученная спираль (виток = 10 н.п.) Цепи антипараллельны: 5′→3′ и 3′→
- 10. Третичная структура ДНК: нуклеопротеидные комплексы (хромосомы) Гистоновые белки: белки с высоким содержанием лиз и арг 5
- 11. Структура нуклеосом
- 12. Пространственная структура РНК Одноцепочечная Шпильки – спирализованные участки (водородные связи) Не соблюдается правило Чаргаффа Виды РНК:
- 13. тРНК Структура тРНК: 1 – шпильки 2 - петли молекулы-адапторы: переводят информацию мРНК в последовательность аминокислот
- 14. рРНК структурный компонент рибосом 80% от общего количества РНК в клетке 4 типа у эукариот: 5S,
- 15. Образование фосфорибозилдифосфата (ФРДФ) Продукты расщепления нуклеиновых кислот тканей и пищи используются повторно в незначительной степени. Почти
- 16. Синтез пуриновых нуклеотидов: основные этапы (см. схему реакций на слайде 17) Сборка пуринового гетероциклического основания осуществляется
- 18. Происхождение атомов С и N в пуриновом основании
- 19. Образование АМФ и ГМФ из ИМФ Образование АДФ, ГДФ, ГТФ В образовании АМФ из ИМФ участвует
- 21. Ферменты синтеза АМФ И ГМФ: подписи к схеме слайда 20. В синтезе АМФ из ИМФ участвуют
- 22. Регуляция синтеза пуриновых нуклеотидов Аллостерические ферменты: ФРДФ-синтетаза амидофосфорибозилтрансфераза ИМФ-дегидрогеназа Аденилосукцинатсинтетаза Отрицательные эффекторы: АМФ, ГМФ
- 23. Запасные пути синтеза пуриновых нуклеотидов : роль «пути спасения» В период активного роста тканей синтез пуриновых
- 24. Пути спасения в синтезе пуриновых нуклеотидов
- 25. Ферменты «пути спасения» в синтезе пуриновых нуклеотидов К слайду 24: 1 – гипоксантин-гуанинфосфорибозилтрансфераза 2 – аденинфосфорибозилтрансфераза
- 26. Катаболизм пуриновых нуклеотидов Отщепление фосфата, аминогруппы, рибозы с образованием азотистых оснований гипоксантина и ксантина (см. схему
- 27. Схема реакций катаболизма пуриновых нуклеотидов
- 28. Ферменты катаболизма пуриновых нуклеотидов К слайду 27: 1 – фосфатаза (нуклеотидаза) 2 – аденозиндезаминаза 3 –
- 29. Нарушения обмена пуриновых нуклеотидов Дефект генов ферментов гиперактивация или устойчивость ФРДФ-синтетазы к аллостерическим ингибиторам снижение активности
- 30. Синтез пиримидиновых нуклеотидов Основные этапы синтеза: Формирование пиримидинового кольца (оротата) из глутамина, аспартата, СО2 Взаимодействие оротата
- 31. Образование оротата и УМФ глутамин + СО2 + 2 АТФ + Н2О → карбамоилфосфат + 2
- 32. Нарушения образования оротата Мутация в гене УМФ-синтазы приводит к нарушению образования УМФ их оротата и вызывает
- 33. Фосфорилирование УМФ и образование ЦТФ Фосфорилирование УМФ: образование УТФ УМФ + АТФ → УДФ + АДФ
- 34. Регуляция синтеза пиримидиновых нуклеотидов Аллостерическая регуляция по механизму отрицательной обратной связи: УТФ ингибирует КФС II в
- 35. Катаболизм пиримидиновых нуклеотидов Отщепление остатков фосфорной кислоты и рибозы (аналогично катаболизму пуриновых нуклеотидов) Пиримидиновые основания разрушаются
- 36. Образование дезоксирибонуклеотидов Образование дНДФ (А, Г, Ц, У) из НДФ Образование дТМФ из дУМФ Внутриклеточная концентрация
- 37. «Работа» рибонуклеотидредуктазы
- 38. Регуляция активности рибонуклеотидредуктазного комлпекса Аллостерический фермент Отрицательные эффекторы: дНТФ дАТФ – ингибитор восстановления всех рибонуклеотидов Иммунодефициты:
- 39. Синтез тимидиловых нуклеотидов Тимидилсинтазный комплекс ферментов и его ингибирование фторурацилом и метотрексатом
- 40. Тимидилсинтазный комплекс ферментов 1- Тимидилсинтаза (включение одноуглеродного радикала в дУМФ) 2- Дигидрофолатредуктаза 3- Сериноксиметилтрансфераза (перенос оксиметильной
- 41. Задание для самостоятельной работы Изучить информацию по теме: «Ферменты синтеза нуклеотидов – мишени действия противоопухолевых и
- 42. Заключение «Обмен нуклеотидов» Большая часть используемых в клетках нуклеотидов синтезируется de novo из простых предшественников (с
- 43. Литература по теме «Обмен нуклеотидов» 1. Биохимия с упражнениями и задачами: учебник для студентов ВУЗов /
- 44. Матричные биосинтезы Репликация Транскрипция Трансляция
- 45. РЕПЛИКАЦИЯ: синтез ДНК Протекает в ядре в S-фазу клеточного цикла перед митозом Стимулы: гормоны, ростовые факторы,
- 46. 1 этап репликации: инициация Формирование репликативной вилки: ДНК-топоизомераза гидролизует 3′,5′-фосфодиэфирную связь в одной из цепей ДНК
- 47. Схема инициации репликации
- 48. 2 этап репликации: элонгация Синтез новых цепей ДНК Лидирующая цепь: 3′ - 5′ (синтез непрерывный по
- 49. 3 этап репликации: терминация Исключение праймеров Завершение формирования отстающей цепи ДНК Эндонуклеаза (РНКаза) удаляет РНК-праймер ДНК-полимераза
- 50. Схема репликативной вилки
- 51. Репарация ошибок и повреждений ДНК Причина повреждений ДНК: действие факторов окружающей и внутренней среды Повреждение ДНК
- 52. Схема работы системы репарации ДНК
- 53. Роль системы репарации Репарация необходима для сохранения генома и возможна благодаря существованию 2-х цепей ДНК Снижение
- 54. ТРАНСКРИПЦИЯ: синтез РНК Протекает в ядре вне зависимости от фаз клеточного цикла Матрица: нить ДНК 3′
- 55. 1 этап транскрипции: инициация Промотор – последовательность ДНК (ТАТА), с которой связывается РНК-полимераза Сайт терминации –
- 56. 2 этап транскрипции: элонгация и терминация Элонгация: рост нити пре-РНК Факторы элонгации (E, H, F) повышают
- 57. Схема транскрипции
- 58. Посттранскрипционные модификации пре-РНК «Созревание» пре-мРНК «Кэпирование» на стадии элонгации Образование поли(А)- «хвоста» после транскрипции Сплайсинг –
- 59. Схема «созревания» пре-мРНК
- 60. «Созревание» пре-тРНК Удаление интронов Модификация азотистых оснований (10-15%) Формирование акцепторного участка и антикодона 3. Выход зрелых
- 61. «Созревание» пре-рРНК
- 62. ТРАНСЛЯЦИЯ: синтез белка Место синтеза: рибосомы Матрица: мРНК Субстраты: аминокислоты (АК) Адапторы: тРНК Источники энергии: АТФ,
- 63. Свойства биологического кода Триплетность Наличие терминирующих кодонов (UAA, UAG, UGA) Специфичность Вырожденность Универсальность Однонаправленность Колинеарность
- 64. Активация аминокислот
- 65. 1 этап трансляции: инициация К мРНК присоединяется малая субъединица рибосомы, фактор инициации IF, мет-тРНК и ГТФ.
- 66. 2 этап трансляции: элонгация (рост пептидной цепи) Стадии элонгации: Связывание аа-тРНК в А-центре при участии фактора
- 67. 3 этап трансляции: терминация Высвобождение пептида из связи с тРНК и рибосомой: Стоп-кодоны UAA, UAG, UGA
- 68. Посттрансляционные модификации белков – образование функционально активных белков Частичный протеолиз Фолдинг – формирование пространственной структуры (II,
- 69. Регуляция матричных биосинтезов Экспрессия генов — процесс, в ходе которого наследственная информация от гена (последовательности нуклеотидов
- 70. Адаптивная регуляция обеспечивает изменение скорости экспрессии генов в ответ на меняющиеся условия среды (индуцибельная экспрессия). Осуществляется
- 71. Примеры ингибиторов матричных биосинтезов Токсин белой поганки аманитин ингибирует РНК-полимеразу II (синтез мРНК) Энтеротоксин возбудителя дифтерии
- 72. Задание для самостоятельной работы Изучить информацию по теме: «Лекарственные препараты - ингибиторы синтеза нуклеиновых кислот и
- 73. Заключение «Матричные биосинтезы» Процессы репликации, транскрипции, трансляции (матричные биосинтезы) лежат в основе «производства» белков и ферментов,
- 75. Скачать презентацию

Актуальность темы
Нуклеотиды и их производные выполняют многообразные функции в организме человека:
Актуальность темы
Нуклеотиды и их производные выполняют многообразные функции в организме человека:

План лекции
Образование фосфорибозилдифосфата (ФРДФ) – ключевой момент в синтезе нуклеотидов
Синтез и
План лекции
Образование фосфорибозилдифосфата (ФРДФ) – ключевой момент в синтезе нуклеотидов
Синтез и

Цель лекции
Знать:
Основные метаболические пути превращения пуриновых и пиримидиновых нуклеотидов
Химико-биологическую сущность процессов
Цель лекции
Знать:
Основные метаболические пути превращения пуриновых и пиримидиновых нуклеотидов
Химико-биологическую сущность процессов

Вспомните самостоятельно
из курса химии, используя слайды 6-14
Пуриновые и пиримидиновые азотистые
Вспомните самостоятельно
из курса химии, используя слайды 6-14
Пуриновые и пиримидиновые азотистые
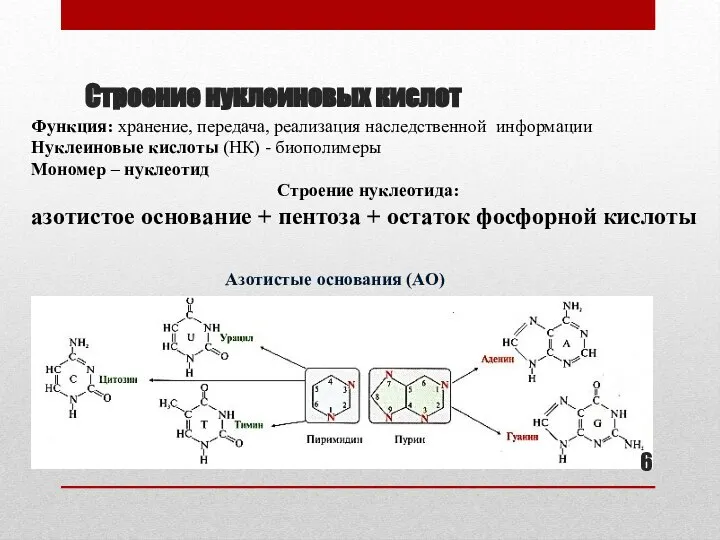
Строение нуклеиновых кислот
Функция: хранение, передача, реализация наследственной информации
Нуклеиновые кислоты (НК) -
Строение нуклеиновых кислот
Функция: хранение, передача, реализация наследственной информации
Нуклеиновые кислоты (НК) -
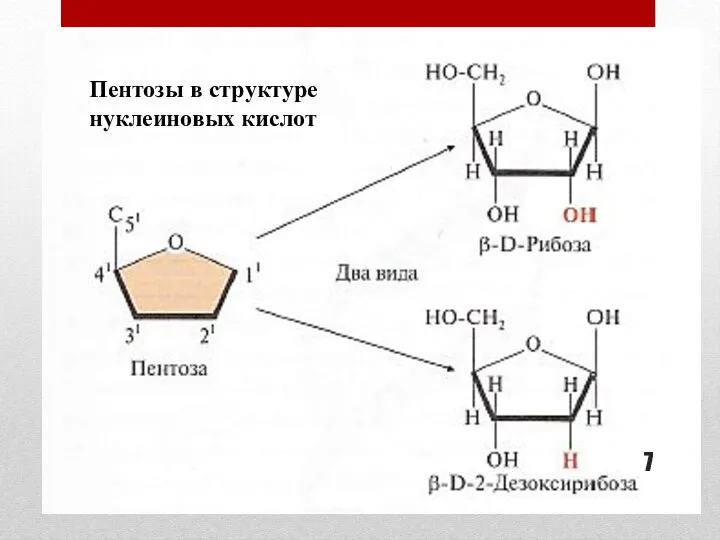
Пентозы в структуре нуклеиновых кислот
Пентозы в структуре нуклеиновых кислот
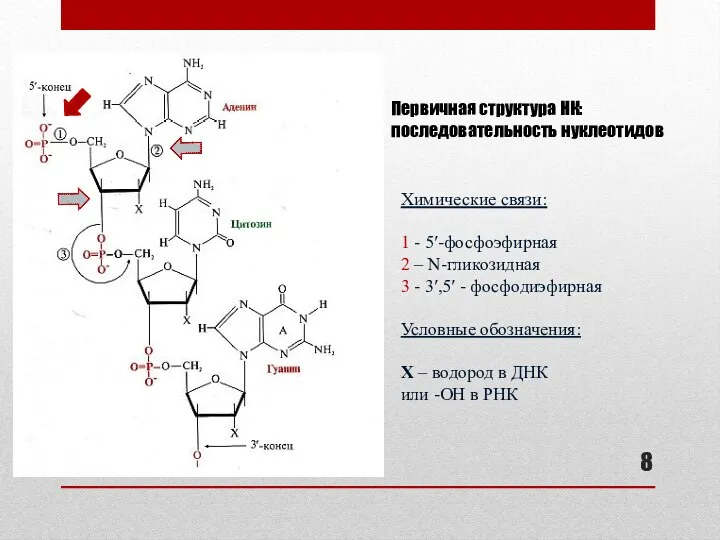
Первичная структура НК: последовательность нуклеотидов
Химические связи:
1 - 5′-фосфоэфирная
2 – N-гликозидная
3 -
Первичная структура НК: последовательность нуклеотидов
Химические связи:
1 - 5′-фосфоэфирная
2 – N-гликозидная
3 -
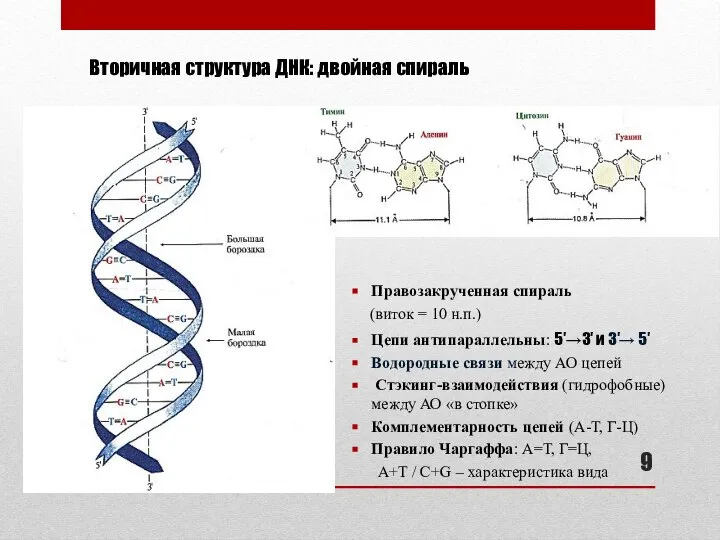
Вторичная структура ДНК: двойная спираль
Правозакрученная спираль
(виток = 10 н.п.)
Цепи антипараллельны:
Вторичная структура ДНК: двойная спираль
Правозакрученная спираль
(виток = 10 н.п.)
Цепи антипараллельны:

Третичная структура ДНК: нуклеопротеидные комплексы (хромосомы)
Гистоновые белки: белки с высоким содержанием
Третичная структура ДНК: нуклеопротеидные комплексы (хромосомы)
Гистоновые белки: белки с высоким содержанием
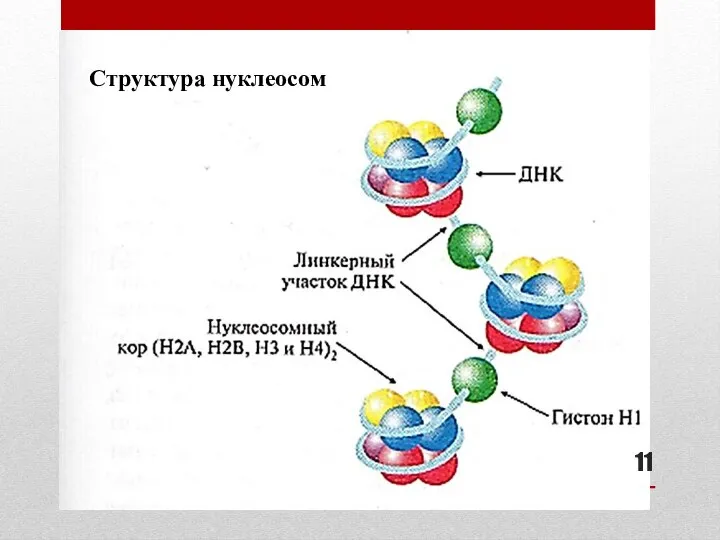
Структура нуклеосом
Структура нуклеосом

Пространственная структура РНК
Одноцепочечная
Шпильки – спирализованные участки (водородные связи)
Не соблюдается правило Чаргаффа
Виды
Пространственная структура РНК
Одноцепочечная
Шпильки – спирализованные участки (водородные связи)
Не соблюдается правило Чаргаффа
Виды
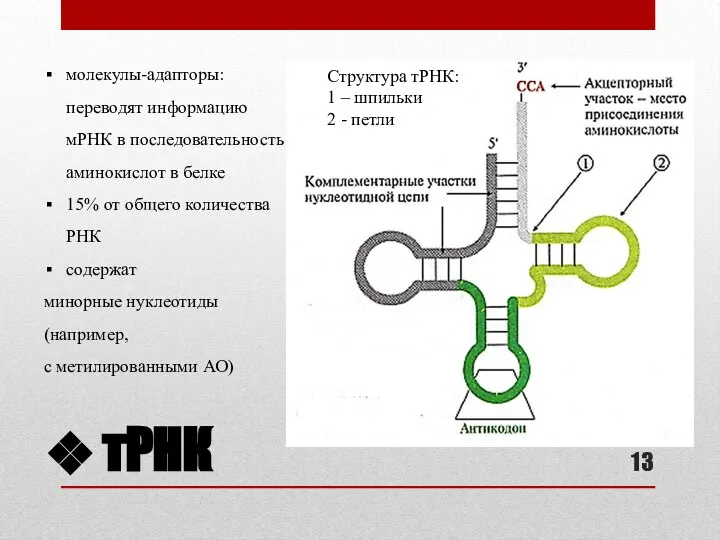
тРНК
Структура тРНК:
1 – шпильки
2 - петли
молекулы-адапторы: переводят информацию мРНК в последовательность
тРНК
Структура тРНК:
1 – шпильки
2 - петли
молекулы-адапторы: переводят информацию мРНК в последовательность
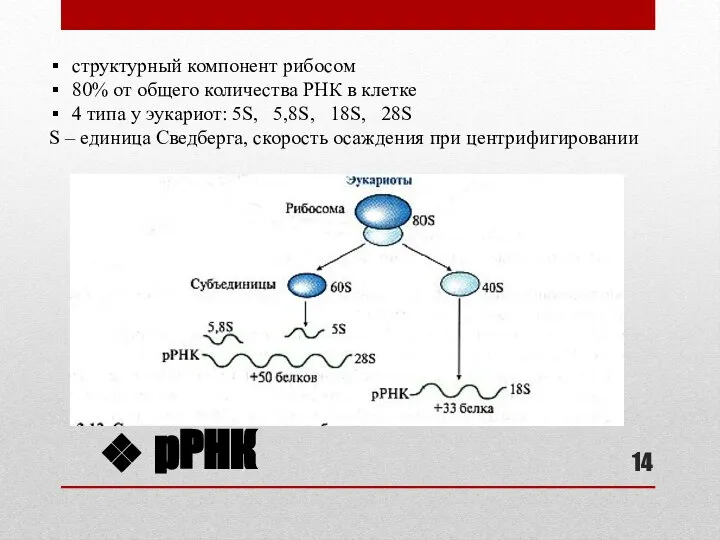
рРНК
структурный компонент рибосом
80% от общего количества РНК в клетке
4 типа у
рРНК
структурный компонент рибосом
80% от общего количества РНК в клетке
4 типа у

Образование фосфорибозилдифосфата (ФРДФ)
Продукты расщепления нуклеиновых кислот тканей и пищи используются повторно
Образование фосфорибозилдифосфата (ФРДФ)
Продукты расщепления нуклеиновых кислот тканей и пищи используются повторно

Синтез пуриновых нуклеотидов:
основные этапы (см. схему реакций на слайде 17)
Сборка
Синтез пуриновых нуклеотидов:
основные этапы (см. схему реакций на слайде 17)
Сборка
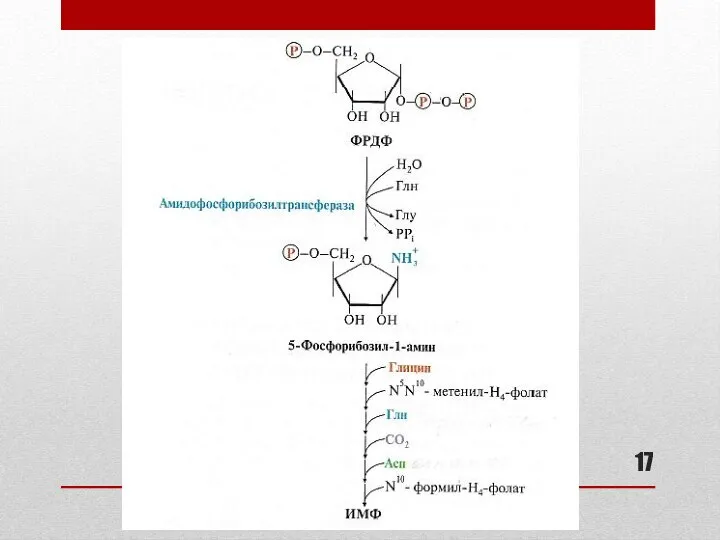
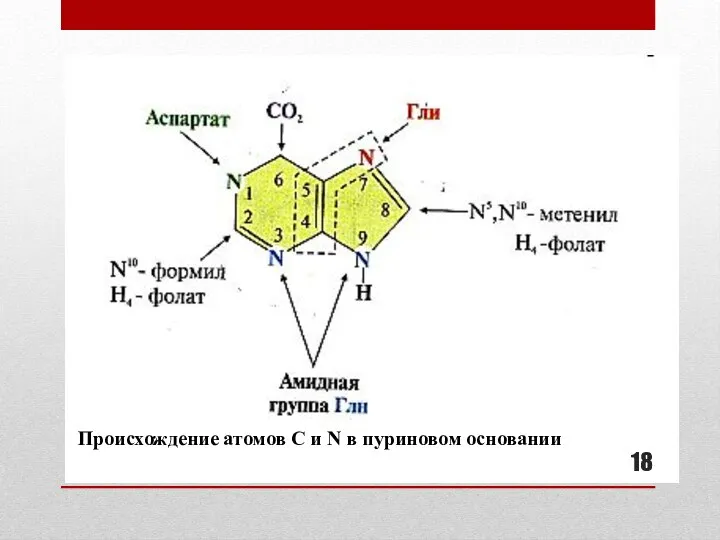
Происхождение атомов С и N в пуриновом основании
Происхождение атомов С и N в пуриновом основании

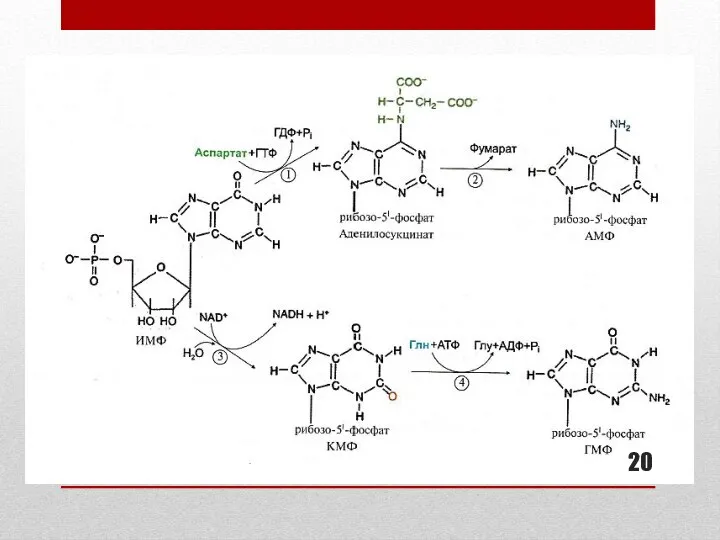
Образование АМФ и ГМФ из ИМФ
Образование АДФ, ГДФ, ГТФ
В образовании АМФ
Образование АМФ и ГМФ из ИМФ
Образование АДФ, ГДФ, ГТФ
В образовании АМФ

Ферменты синтеза АМФ И ГМФ:
подписи к схеме слайда 20.
В синтезе АМФ
Ферменты синтеза АМФ И ГМФ:
подписи к схеме слайда 20.
В синтезе АМФ

Регуляция синтеза пуриновых нуклеотидов
Аллостерические ферменты:
ФРДФ-синтетаза
амидофосфорибозилтрансфераза
ИМФ-дегидрогеназа
Аденилосукцинатсинтетаза
Отрицательные эффекторы: АМФ, ГМФ
Регуляция синтеза пуриновых нуклеотидов
Аллостерические ферменты:
ФРДФ-синтетаза
амидофосфорибозилтрансфераза
ИМФ-дегидрогеназа
Аденилосукцинатсинтетаза
Отрицательные эффекторы: АМФ, ГМФ

Запасные пути синтеза пуриновых нуклеотидов : роль «пути спасения»
В период активного
Запасные пути синтеза пуриновых нуклеотидов : роль «пути спасения»
В период активного
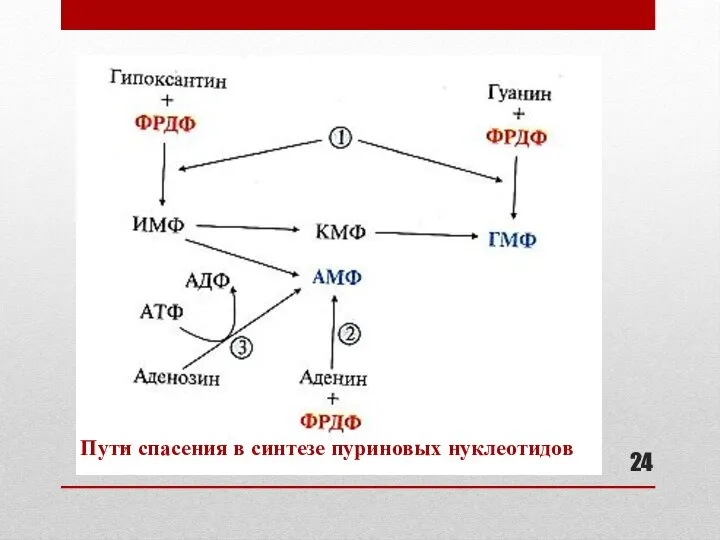
Пути спасения в синтезе пуриновых нуклеотидов
Пути спасения в синтезе пуриновых нуклеотидов

Ферменты «пути спасения» в синтезе пуриновых нуклеотидов
К слайду 24:
1 –
Ферменты «пути спасения» в синтезе пуриновых нуклеотидов
К слайду 24:
1 –

Катаболизм пуриновых нуклеотидов
Отщепление фосфата, аминогруппы, рибозы с образованием азотистых оснований гипоксантина
Катаболизм пуриновых нуклеотидов
Отщепление фосфата, аминогруппы, рибозы с образованием азотистых оснований гипоксантина
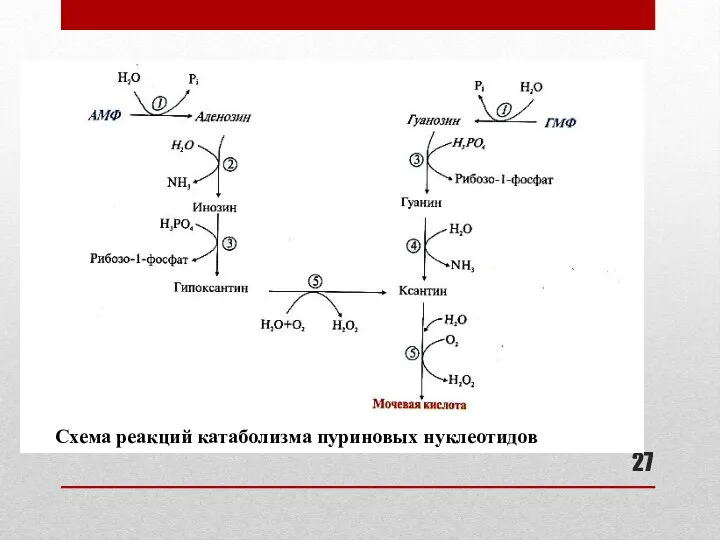
Схема реакций катаболизма пуриновых нуклеотидов
Схема реакций катаболизма пуриновых нуклеотидов

Ферменты катаболизма пуриновых нуклеотидов
К слайду 27:
1 – фосфатаза (нуклеотидаза)
2 – аденозиндезаминаза
3
Ферменты катаболизма пуриновых нуклеотидов
К слайду 27:
1 – фосфатаза (нуклеотидаза)
2 – аденозиндезаминаза
3

Нарушения обмена пуриновых нуклеотидов
Дефект генов ферментов
гиперактивация или устойчивость ФРДФ-синтетазы к аллостерическим
Нарушения обмена пуриновых нуклеотидов
Дефект генов ферментов
гиперактивация или устойчивость ФРДФ-синтетазы к аллостерическим

Синтез пиримидиновых нуклеотидов
Основные этапы синтеза:
Формирование пиримидинового кольца (оротата) из глутамина, аспартата,
Синтез пиримидиновых нуклеотидов
Основные этапы синтеза:
Формирование пиримидинового кольца (оротата) из глутамина, аспартата,

Образование оротата и УМФ
глутамин + СО2 + 2 АТФ +
Образование оротата и УМФ
глутамин + СО2 + 2 АТФ +

Нарушения образования оротата
Мутация в гене УМФ-синтазы приводит к нарушению образования УМФ
Нарушения образования оротата
Мутация в гене УМФ-синтазы приводит к нарушению образования УМФ

Фосфорилирование УМФ и
образование ЦТФ
Фосфорилирование УМФ: образование УТФ
УМФ + АТФ → УДФ
Фосфорилирование УМФ и
образование ЦТФ
Фосфорилирование УМФ: образование УТФ
УМФ + АТФ → УДФ

Регуляция синтеза пиримидиновых нуклеотидов
Аллостерическая регуляция по механизму отрицательной обратной связи:
УТФ ингибирует
Регуляция синтеза пиримидиновых нуклеотидов
Аллостерическая регуляция по механизму отрицательной обратной связи:
УТФ ингибирует

Катаболизм пиримидиновых нуклеотидов
Отщепление остатков фосфорной кислоты и рибозы (аналогично катаболизму пуриновых
Катаболизм пиримидиновых нуклеотидов
Отщепление остатков фосфорной кислоты и рибозы (аналогично катаболизму пуриновых

Образование дезоксирибонуклеотидов
Образование дНДФ (А, Г, Ц, У) из НДФ
Образование дТМФ из
Образование дезоксирибонуклеотидов
Образование дНДФ (А, Г, Ц, У) из НДФ
Образование дТМФ из
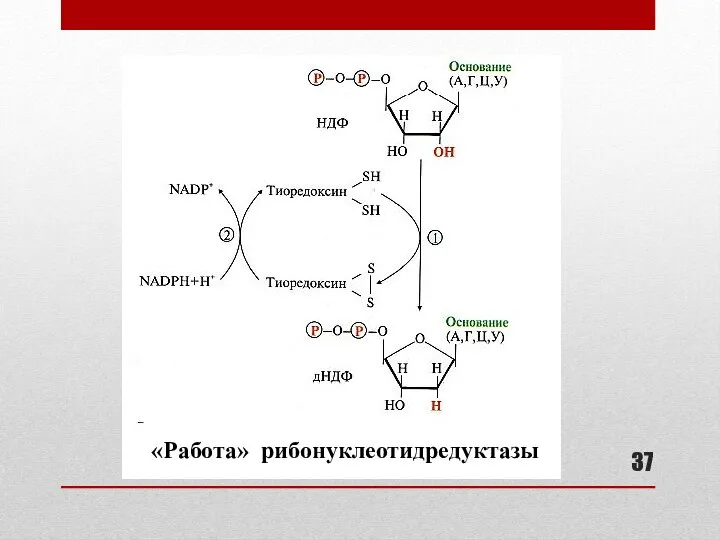
«Работа» рибонуклеотидредуктазы
«Работа» рибонуклеотидредуктазы

Регуляция активности рибонуклеотидредуктазного комлпекса
Аллостерический фермент
Отрицательные эффекторы: дНТФ
дАТФ – ингибитор восстановления всех
Регуляция активности рибонуклеотидредуктазного комлпекса
Аллостерический фермент
Отрицательные эффекторы: дНТФ
дАТФ – ингибитор восстановления всех
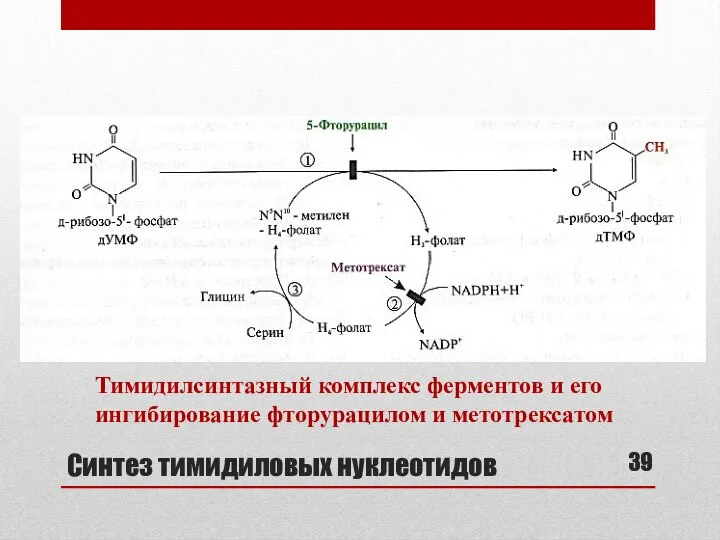
Синтез тимидиловых нуклеотидов
Тимидилсинтазный комплекс ферментов и его ингибирование фторурацилом и метотрексатом
Синтез тимидиловых нуклеотидов
Тимидилсинтазный комплекс ферментов и его ингибирование фторурацилом и метотрексатом

Тимидилсинтазный комплекс ферментов
1- Тимидилсинтаза (включение одноуглеродного радикала в дУМФ)
2- Дигидрофолатредуктаза
3- Сериноксиметилтрансфераза
Тимидилсинтазный комплекс ферментов
1- Тимидилсинтаза (включение одноуглеродного радикала в дУМФ)
2- Дигидрофолатредуктаза
3- Сериноксиметилтрансфераза

Задание для самостоятельной работы
Изучить информацию по теме: «Ферменты синтеза нуклеотидов –
Задание для самостоятельной работы
Изучить информацию по теме: «Ферменты синтеза нуклеотидов –

Заключение «Обмен нуклеотидов»
Большая часть используемых в клетках нуклеотидов синтезируется de novo
Заключение «Обмен нуклеотидов»
Большая часть используемых в клетках нуклеотидов синтезируется de novo

Литература по теме «Обмен нуклеотидов»
1. Биохимия с упражнениями и задачами: учебник для
Литература по теме «Обмен нуклеотидов»
1. Биохимия с упражнениями и задачами: учебник для

Матричные биосинтезы
Репликация
Транскрипция
Трансляция
Матричные биосинтезы
Репликация
Транскрипция
Трансляция

РЕПЛИКАЦИЯ: синтез ДНК
Протекает в ядре в S-фазу клеточного цикла перед митозом
Стимулы:
РЕПЛИКАЦИЯ: синтез ДНК
Протекает в ядре в S-фазу клеточного цикла перед митозом
Стимулы:

1 этап репликации: инициация
Формирование репликативной вилки:
ДНК-топоизомераза гидролизует 3′,5′-фосфодиэфирную связь в одной
1 этап репликации: инициация
Формирование репликативной вилки:
ДНК-топоизомераза гидролизует 3′,5′-фосфодиэфирную связь в одной
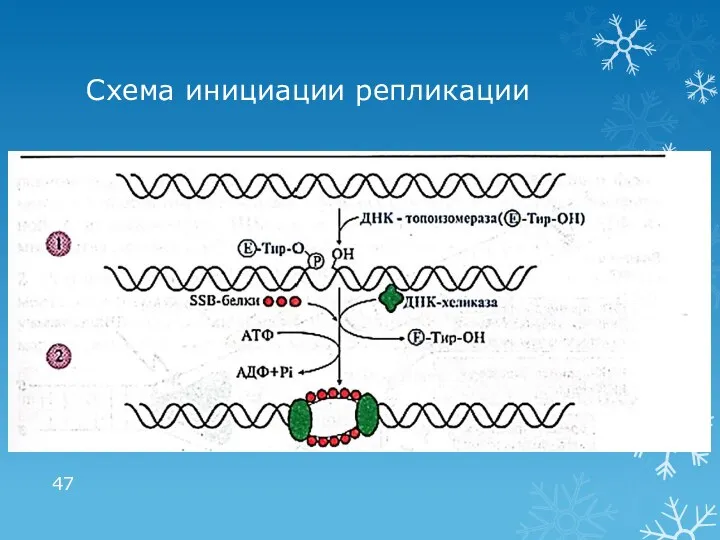
Схема инициации репликации
Схема инициации репликации

2 этап репликации: элонгация
Синтез новых цепей ДНК
Лидирующая цепь: 3′ - 5′
2 этап репликации: элонгация
Синтез новых цепей ДНК
Лидирующая цепь: 3′ - 5′

3 этап репликации: терминация
Исключение праймеров
Завершение формирования отстающей цепи ДНК
Эндонуклеаза (РНКаза) удаляет
3 этап репликации: терминация
Исключение праймеров
Завершение формирования отстающей цепи ДНК
Эндонуклеаза (РНКаза) удаляет
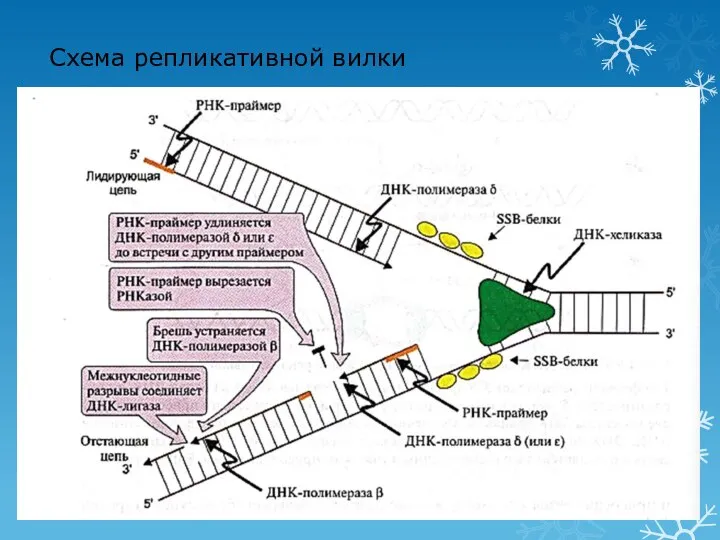
Схема репликативной вилки
Схема репликативной вилки

Репарация ошибок и повреждений ДНК
Причина повреждений ДНК:
действие факторов окружающей и внутренней
Репарация ошибок и повреждений ДНК
Причина повреждений ДНК:
действие факторов окружающей и внутренней
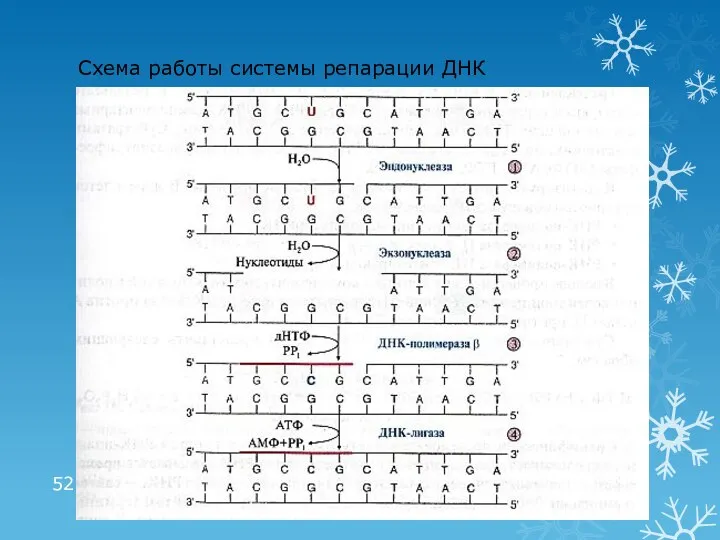
Схема работы системы репарации ДНК
Схема работы системы репарации ДНК

Роль системы репарации
Репарация необходима для сохранения генома и возможна благодаря существованию
Роль системы репарации
Репарация необходима для сохранения генома и возможна благодаря существованию

ТРАНСКРИПЦИЯ: синтез РНК
Протекает в ядре вне зависимости от фаз клеточного цикла
Матрица:
ТРАНСКРИПЦИЯ: синтез РНК
Протекает в ядре вне зависимости от фаз клеточного цикла
Матрица:

1 этап транскрипции: инициация
Промотор – последовательность ДНК (ТАТА), с которой связывается
1 этап транскрипции: инициация
Промотор – последовательность ДНК (ТАТА), с которой связывается

2 этап транскрипции:
элонгация и терминация
Элонгация: рост нити пре-РНК
Факторы элонгации (E, H,
2 этап транскрипции:
элонгация и терминация
Элонгация: рост нити пре-РНК
Факторы элонгации (E, H,
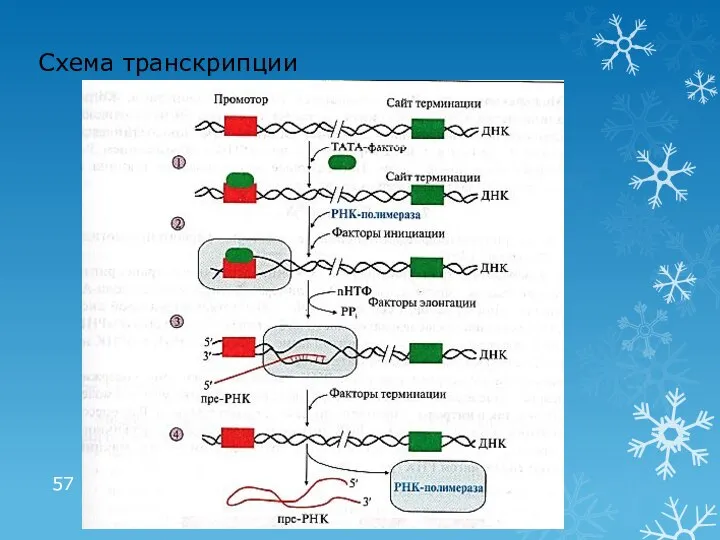
Схема транскрипции
Схема транскрипции

Посттранскрипционные модификации пре-РНК
«Созревание» пре-мРНК
«Кэпирование» на стадии элонгации
Образование поли(А)- «хвоста» после транскрипции
Сплайсинг
Посттранскрипционные модификации пре-РНК
«Созревание» пре-мРНК
«Кэпирование» на стадии элонгации
Образование поли(А)- «хвоста» после транскрипции
Сплайсинг
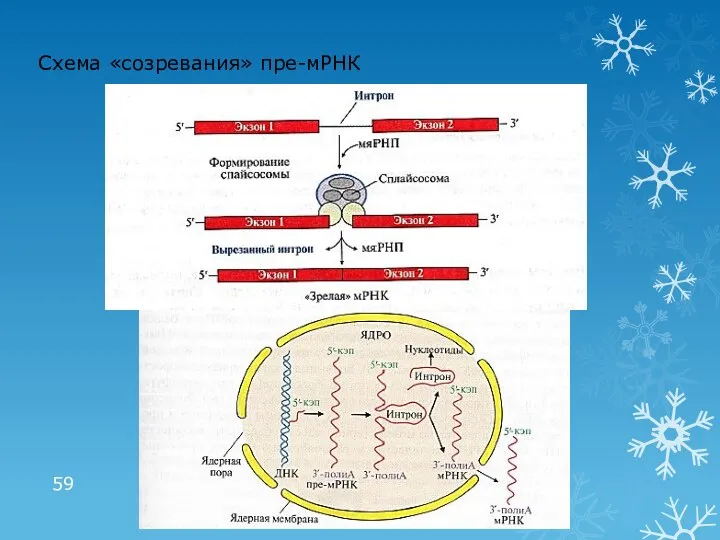
Схема «созревания» пре-мРНК
Схема «созревания» пре-мРНК
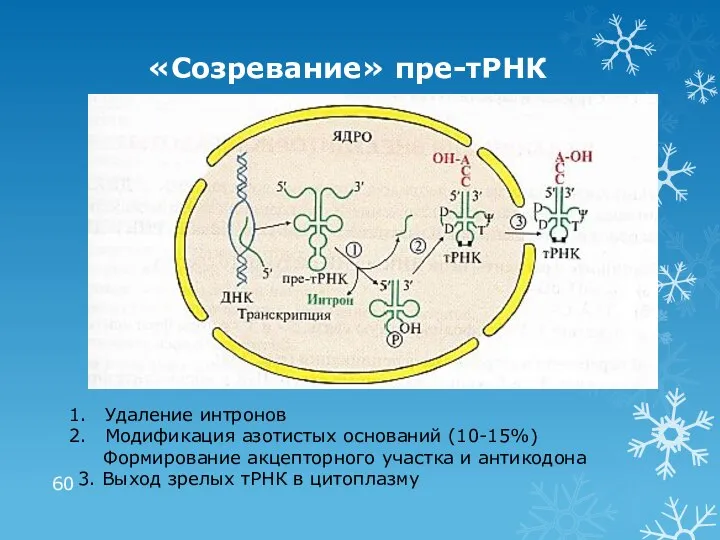
«Созревание» пре-тРНК
Удаление интронов
Модификация азотистых оснований (10-15%)
Формирование акцепторного участка и антикодона
3.
«Созревание» пре-тРНК
Удаление интронов
Модификация азотистых оснований (10-15%)
Формирование акцепторного участка и антикодона
3.
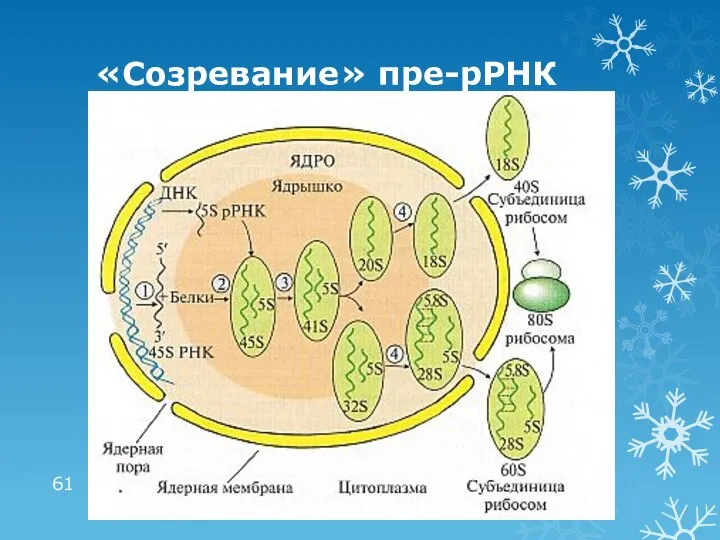
«Созревание» пре-рРНК
«Созревание» пре-рРНК

ТРАНСЛЯЦИЯ: синтез белка
Место синтеза: рибосомы Матрица: мРНК
Субстраты: аминокислоты (АК) Адапторы: тРНК
Источники
ТРАНСЛЯЦИЯ: синтез белка
Место синтеза: рибосомы Матрица: мРНК
Субстраты: аминокислоты (АК) Адапторы: тРНК
Источники

Свойства биологического кода
Триплетность
Наличие терминирующих кодонов (UAA, UAG, UGA)
Специфичность
Вырожденность
Универсальность
Однонаправленность
Колинеарность
Свойства биологического кода
Триплетность
Наличие терминирующих кодонов (UAA, UAG, UGA)
Специфичность
Вырожденность
Универсальность
Однонаправленность
Колинеарность
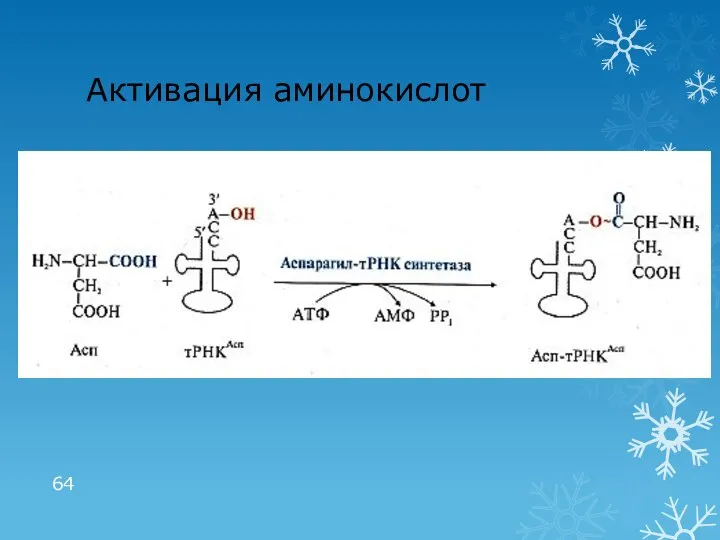
Активация аминокислот
Активация аминокислот
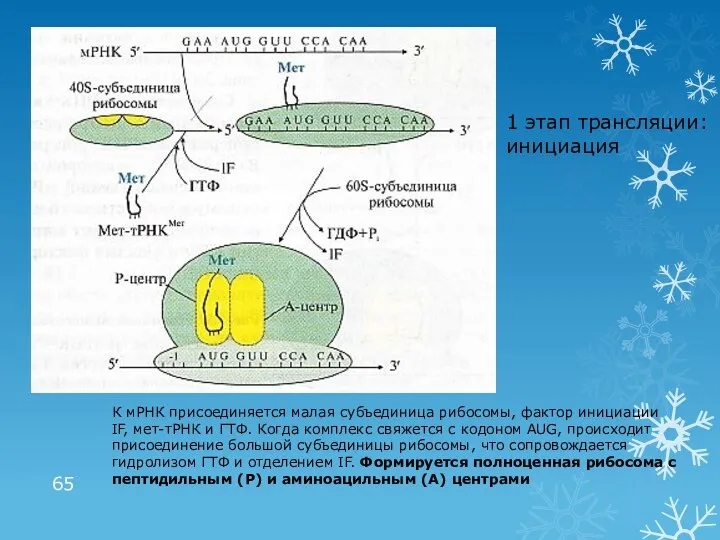
1 этап трансляции: инициация
К мРНК присоединяется малая субъединица рибосомы, фактор инициации
1 этап трансляции: инициация
К мРНК присоединяется малая субъединица рибосомы, фактор инициации

2 этап трансляции: элонгация (рост пептидной цепи)
Стадии элонгации:
Связывание аа-тРНК в А-центре
2 этап трансляции: элонгация (рост пептидной цепи)
Стадии элонгации:
Связывание аа-тРНК в А-центре
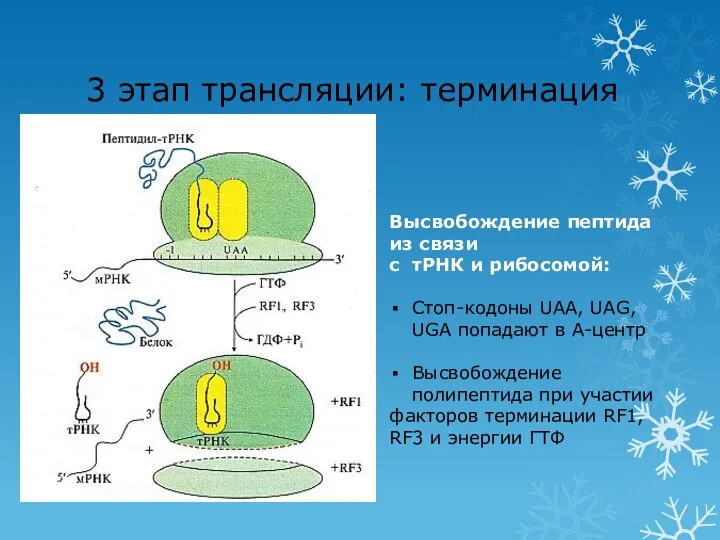
3 этап трансляции: терминация
Высвобождение пептида из связи
с тРНК и рибосомой:
Стоп-кодоны UAA,
3 этап трансляции: терминация
Высвобождение пептида из связи
с тРНК и рибосомой:
Стоп-кодоны UAA,

Посттрансляционные модификации белков – образование функционально активных белков
Частичный протеолиз
Фолдинг – формирование
Посттрансляционные модификации белков – образование функционально активных белков
Частичный протеолиз
Фолдинг – формирование

Регуляция матричных биосинтезов
Экспрессия генов — процесс, в ходе которого наследственная информация
Регуляция матричных биосинтезов
Экспрессия генов — процесс, в ходе которого наследственная информация

Адаптивная регуляция обеспечивает изменение скорости экспрессии генов в ответ на меняющиеся

Примеры ингибиторов матричных биосинтезов
Токсин белой поганки аманитин ингибирует РНК-полимеразу II (синтез
Примеры ингибиторов матричных биосинтезов
Токсин белой поганки аманитин ингибирует РНК-полимеразу II (синтез

Задание для самостоятельной работы
Изучить информацию по теме: «Лекарственные препараты - ингибиторы
Задание для самостоятельной работы
Изучить информацию по теме: «Лекарственные препараты - ингибиторы

Заключение «Матричные биосинтезы»
Процессы репликации, транскрипции, трансляции (матричные биосинтезы) лежат в основе
Заключение «Матричные биосинтезы»
Процессы репликации, транскрипции, трансляции (матричные биосинтезы) лежат в основе
 Безработица инфляция
Безработица инфляция  Презентация «КОМИССИЯ»
Презентация «КОМИССИЯ» Т Е М А «ОСНОВАНИЯ ВОЗНИКНОВЕНИЯ, ИЗМЕНЕНИЯ И ПРЕКРАЩЕНИЯ ГРАЖДАНСКИХ ПРАВООТНОШЕНИЙ»
Т Е М А «ОСНОВАНИЯ ВОЗНИКНОВЕНИЯ, ИЗМЕНЕНИЯ И ПРЕКРАЩЕНИЯ ГРАЖДАНСКИХ ПРАВООТНОШЕНИЙ» Поход 50 км. Преодолей себя
Поход 50 км. Преодолей себя Роль исламского образования и духовного воспитания в формировании религиозной идентичности детей
Роль исламского образования и духовного воспитания в формировании религиозной идентичности детей Инженерная графика. Основы начертальной геометрии
Инженерная графика. Основы начертальной геометрии Стимулирование качества труда учителя и критерии оценки качества Методика расчета
Стимулирование качества труда учителя и критерии оценки качества Методика расчета КАЗИМИР СЕВЕРИНОВИЧ МАЛЕВИЧ (1878-1935)
КАЗИМИР СЕВЕРИНОВИЧ МАЛЕВИЧ (1878-1935) The British isles
The British isles Гибкие методологии разработки программного обеспечения
Гибкие методологии разработки программного обеспечения Топ 5 горячих точек мира
Топ 5 горячих точек мира Программирование. Практическое занятие
Программирование. Практическое занятие Музыкальные инструменты народов Урала
Музыкальные инструменты народов Урала Переключение задач
Переключение задач Презентация Жалоба и представление на приговор, определение, постановление суда
Презентация Жалоба и представление на приговор, определение, постановление суда Презентация Психология толпы. Гюстав Лебон
Презентация Психология толпы. Гюстав Лебон American pronunciation
American pronunciation Основы технологии текущего и капитального ремонта скважин
Основы технологии текущего и капитального ремонта скважин Внутрибольничные инфекции
Внутрибольничные инфекции Экономическое обоснование бизнес-проекта GreenLeaf
Экономическое обоснование бизнес-проекта GreenLeaf Камешковая декоративная штукатурка
Камешковая декоративная штукатурка Политические партии и движения
Политические партии и движения Лекция №26 Гнойные заболевания костей и суставов.
Лекция №26 Гнойные заболевания костей и суставов. Вера Мухина
Вера Мухина Широкая Масленица
Широкая Масленица Банковская система Фатеева Т.А., учитель истории МБОУ-гимназии №20 г.Тулы
Банковская система Фатеева Т.А., учитель истории МБОУ-гимназии №20 г.Тулы  Выполнила студентка 41 группы: Лукахина Яна
Выполнила студентка 41 группы: Лукахина Яна Тема: «Числа 1-5» Составила: учитель начальных классов МОУ «Бичуринская ООШ» Мариинско-Посадского района Чувашской Респуб
Тема: «Числа 1-5» Составила: учитель начальных классов МОУ «Бичуринская ООШ» Мариинско-Посадского района Чувашской Респуб