- Окислительное фосфорилирование. Тканевое дыхание

Содержание
- 2. Общая схема катаболизма
- 3. Карта основных путей метаболизма
- 4. Катаболизм основных пищевых веществ 1-3 пищеварение; 4-8 специфические пути катаболизма; 9-10 заключительный (общий путь) катаболизма; 11-
- 5. Связь реакций общего пути катаболизма с ЦПЭ 1-5 первичные доноры водорода для ЦПЭ; 1, 2, 3,
- 7. Катаболизм органических веществ в тканях сопровождается потреблением О2 и выделением СО2. Этот процесс называют тканевым дыханием.
- 8. Окисляемые различные органические вещества (S – субстраты), представляют собой метаболиты катаболизма, их дегидрирование является экзорганическим процессом.
- 9. Окислительное фосфорилирование – основная часть биологического окисления или тканевого дыхания, т.е. процессов, идущих в организме с
- 10. Впервые механизм окислительного фосфорилирования был предложен П.Митчеллом. Согласно этой гипотезе перенос электронов, происходящий на внутренней митохондриальной
- 11. Митохондрии По современным представлениям внутренняя митохондриальная мембрана содержит ряд мультиферментных комплексов, включающих множество ферментов. Эти ферменты
- 12. По современным представлениям внутренняя митохондриальная мембрана содержит ряд мультиферментных комплексов, включающих множество ферментов. Эти ферменты называют
- 13. ПРИНЦИП РАБОТЫ ДЫХАТЕЛЬНОЙ ЦЕПИ В целом работа дыхательной цепи заключается в следующем: 1. Образующиеся в реакциях
- 14. Общая схема окислительного фосфорилирования Таким образом восстановленные формы НАДН и ФАДН2 окисляются ферментами дыхательной цепи, благодаря
- 15. ФЕРМЕНТНЫЕ КОМПЛЕКСЫ ДЫХАТЕЛЬНОЙ ЦЕПИ Всего цепь переноса электронов включает в себя около 40 разнообразных белков и
- 16. Комплексы дыхательной цепи I комплекс - НАДН-дегидрогеназа, II комплекс - включает в себя ФАД-зависимые ферменты (как
- 17. Митохондриальная цепь переноса электронов
- 18. Функции комплексов дыхательной цепи I комплекс – 1. Принимает электроны от НАДН и передает их на
- 19. IV комплекс – 1. Принимает электроны от цитохрома с и передает их на кислород с образованием
- 20. Пути поступления электронов и протонов в дыхательную цепь переносчиков (ферментов от первичных доноров)
- 21. Работа комплексов дыхательной цепи Ферменты дыхательной цепи расположены в строго определенной последовательности: каждый последующий белок обладает
- 22. Стандартные окислительно-восстановительные потенциалы некоторых сопряженных пар
- 23. 3. Здесь атомы водорода (от НАДН и ФАДН2) передают свои электроны в дыхательную ферментативную цепь, по
- 24. 5. Перенос ионов водорода через мембрану (выкачивание) происходит не случайно, а в строго определенных участках мембраны.
- 25. Энергетические соотношения в дыхательной цепи митохондрий и участки переноса ионов водорода через внутреннюю мембрану в межмембранное
- 26. Рис. Изменение свободной энергии (Ео) при переносе электронов по ЦПЭ Е-ФМН – комплекс I; Е-ФАД –
- 27. 6. Как завершение всех предыдущих событий и необходимый их результат происходит наработка АТФ. Ионы водорода теряют
- 28. Сопряжение тканевого дыхания и синтеза АТФ 1. Перенос электронов по ЦПЭ при участии комплексов I, III
- 29. Рис. Сопряжение дыхания и синтеза АТФ в митохондриях I – НАДН-дегидрогеназа; III – QН2- дегидрогеназа; IV-
- 30. 3. Каждый из трех комплексов ЦПЭ (I, III и IV ) обеспечивает необходимый протонный градиент для
- 31. о Витамине С Вит С
- 32. Сопряжение дыхания и синтеза АТФ в митохондриях
- 33. При участии АТФ-АДФ транслоказы, расположенной во внутренней мембране митохондрий, АТФ транспортируется в цитоплазму в обмен на
- 34. На всех этапах процесса часть энергии рассеивается в виде тепла
- 35. Аналоги АТФ: ГТФ ЦТФ УТФ
- 36. Дыхательный контроль Перенос электронов по ЦПЭ и синтез АТФ тесно сопряжены, т.е. могут происходить только одномоментно
- 37. Синтез АТФ регулируется Работа дыхательных ферментов регулируется с помощью эффекта, который получил название дыхательный контроль. Дыхательный
- 38. Возрастание протонного градиента возникает при исчерпании запасов АДФ и накоплении АТФ, т.е. когда АТФ-синтаза лишена своего
- 39. Таким образом скорость синтеза АТФ точно соответствует потребностям клетки в энергии. Ускорение тканевого дыхания и окислительного
- 40. Разобщение дыхания и синтеза АТФ Некоторые липофильные вещества (2,4-динитрофенол, некоторые жирные кислоты) могут переносить ионы водорода
- 41. Терморегуляторная функция дыхания При переносе электронов по ЦПЭ часть энергии рассеивается в виде теплоты, которая используется
- 42. Кроме того дополнительное образование теплоты может происходить путем разобщения дыхания и фосфорилирования в процессе адаптации к
- 43. У новорожденных и зимнеспящих животных разобщение связано с наличием разобщающего белка – термогенина (до 15% от
- 44. Рис. Роль термогенина в митохондрии Кроме динитрофенола, жирных кислот и термогенина протонофорами, к примеру, являются салицилаты,
- 45. Ингибиторы дыхания Некоторые лекарственные вещества, химические агенты и антибиотики вызывают ингибирование ферментов, либо собственно дыхательной цепи:
- 46. Рис. Ингибиторы ферментов дыхательной цепи
- 47. Гипоэнергетические состояния Причинами гипоэнергетических состояний может быть следующее: 1. гиповитаминозы экзогенные и/или эндогенные – снижается скорость
- 48. Состав компонентов дыхательной цепи НАД содержит витамин В3 или РР (никотиновая кислота) ФМН и ФАД –
- 49. Компоненты дыхательной цепи
- 50. Коферменты дегидрогеназ
- 51. Структурные формулы рабочей части коферментов НАД и НАДФ
- 52. Структурные формулы рабочей части коферментов ФАД и ФМН
- 53. Структура убихинона (кофермента Q)
- 54. Цитохромы - это гемопротеины - белки, содержащие в качестве прочно связанной простетической группы гем (окрашены в
- 55. Строение гема А
- 56. Атом железа в геме может менять валентность, присоединяя или отдавая электроны: В дыхательной цепи цитохромы служат
- 57. В цитохромах С и С1 дополнительные ковалентные связи формируются между тиогруппами цистеина и боковыми винильными группами
- 58. Цитохромоксидаза включает комплекс цитохромов а и а3 (комплекс IV). Цитохромоксидаза кроме гема (с Fe) содержит ионы
- 59. Структура гема цитохромов b, c, c1
- 60. Катаболизм основных пищевых веществ 1-3 пищеварение; 4-8 специфические пути катаболизма; 9-10 заключительный (общий путь) катаболизма; 11-
- 62. Скачать презентацию
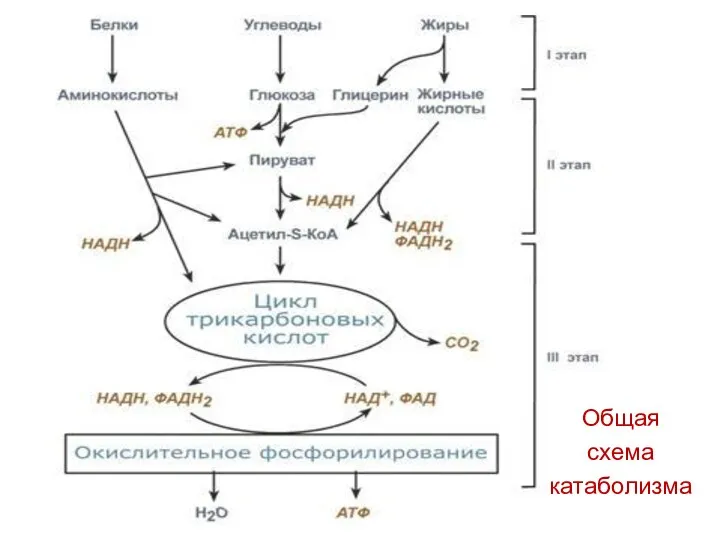
Общая
схема
катаболизма
Общая
схема
катаболизма
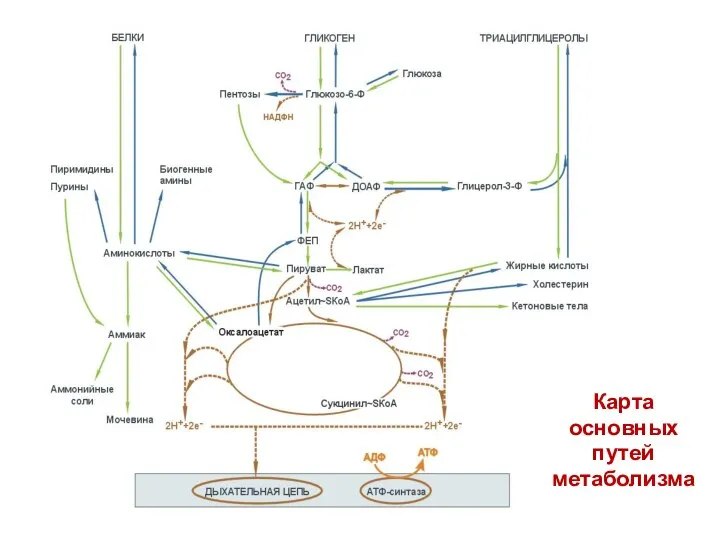
Карта
основных
путей
метаболизма
Карта
основных
путей
метаболизма
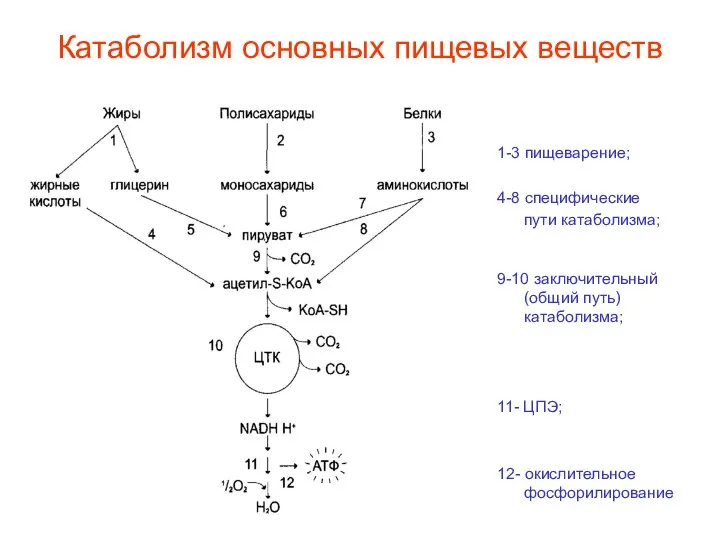
Катаболизм основных пищевых веществ
1-3 пищеварение;
4-8 специфические
пути катаболизма;
9-10 заключительный
Катаболизм основных пищевых веществ
1-3 пищеварение;
4-8 специфические
пути катаболизма;
9-10 заключительный
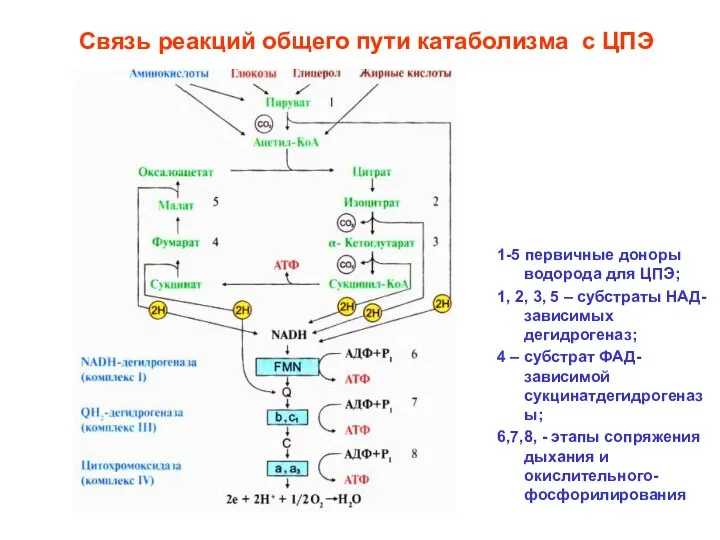
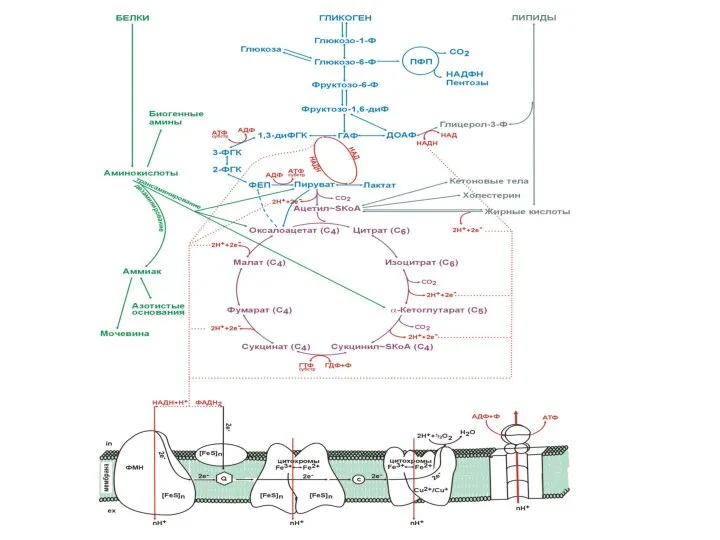
Связь реакций общего пути катаболизма с ЦПЭ
1-5 первичные доноры водорода для
Связь реакций общего пути катаболизма с ЦПЭ
1-5 первичные доноры водорода для

Катаболизм органических веществ в тканях
сопровождается потреблением О2 и
выделением СО2.
Этот
Катаболизм органических веществ в тканях
сопровождается потреблением О2 и
выделением СО2.
Этот

Окисляемые различные органические вещества
(S – субстраты), представляют собой метаболиты
катаболизма, их
Окисляемые различные органические вещества
(S – субстраты), представляют собой метаболиты
катаболизма, их

Окислительное фосфорилирование – основная часть
биологического окисления или тканевого дыхания, т.е.
процессов, идущих
Окислительное фосфорилирование – основная часть
биологического окисления или тканевого дыхания, т.е.
процессов, идущих

Впервые механизм окислительного фосфорилирования
был предложен П.Митчеллом. Согласно этой гипотезе
перенос электронов,
Впервые механизм окислительного фосфорилирования
был предложен П.Митчеллом. Согласно этой гипотезе
перенос электронов,
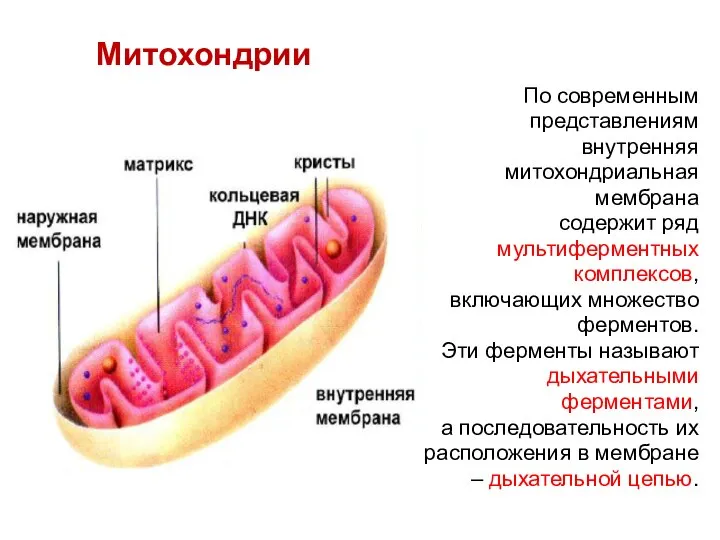
Митохондрии
По современным
представлениям
внутренняя
митохондриальная
мембрана
содержит ряд
мультиферментных
комплексов,
включающих множество
ферментов.
Митохондрии
По современным
представлениям
внутренняя
митохондриальная
мембрана
содержит ряд
мультиферментных
комплексов,
включающих множество
ферментов.

По современным представлениям внутренняя
митохондриальная мембрана содержит ряд
мультиферментных комплексов, включающих
По современным представлениям внутренняя
митохондриальная мембрана содержит ряд
мультиферментных комплексов, включающих

ПРИНЦИП РАБОТЫ ДЫХАТЕЛЬНОЙ ЦЕПИ
В целом работа дыхательной цепи заключается в
ПРИНЦИП РАБОТЫ ДЫХАТЕЛЬНОЙ ЦЕПИ
В целом работа дыхательной цепи заключается в
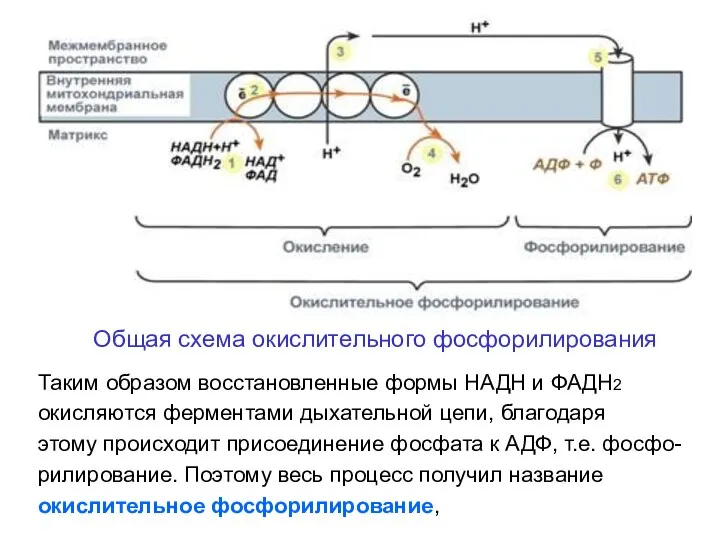
Общая схема окислительного фосфорилирования
Таким образом восстановленные формы НАДН и ФАДН2
окисляются
Общая схема окислительного фосфорилирования
Таким образом восстановленные формы НАДН и ФАДН2
окисляются
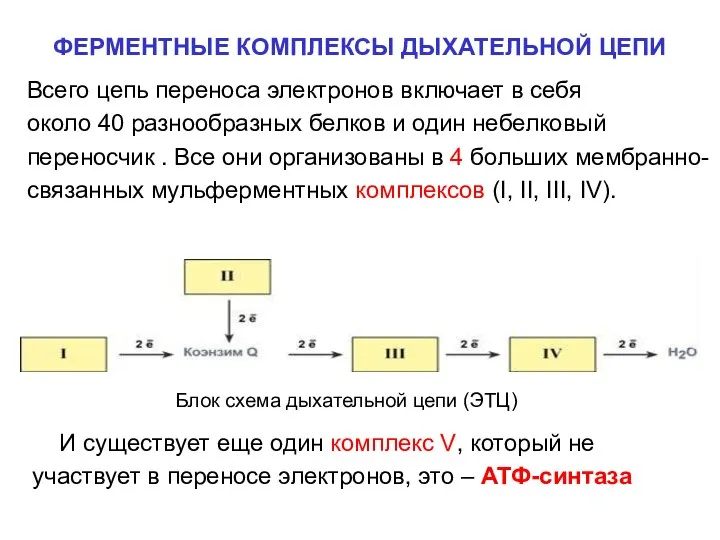
ФЕРМЕНТНЫЕ КОМПЛЕКСЫ ДЫХАТЕЛЬНОЙ ЦЕПИ
Всего цепь переноса электронов включает в себя
около
ФЕРМЕНТНЫЕ КОМПЛЕКСЫ ДЫХАТЕЛЬНОЙ ЦЕПИ
Всего цепь переноса электронов включает в себя
около

Комплексы дыхательной цепи
I комплекс - НАДН-дегидрогеназа,
II комплекс -
Комплексы дыхательной цепи
I комплекс - НАДН-дегидрогеназа,
II комплекс -
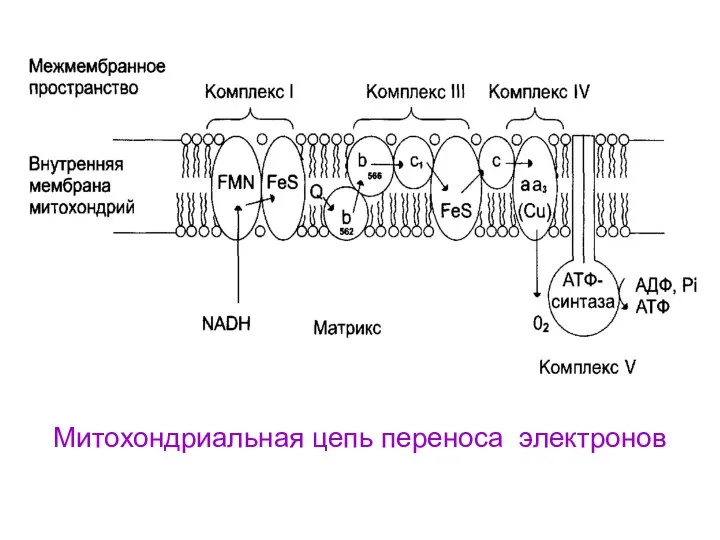
Митохондриальная цепь переноса электронов
Митохондриальная цепь переноса электронов

Функции комплексов дыхательной цепи
I комплекс –
1. Принимает электроны от НАДН
Функции комплексов дыхательной цепи
I комплекс –
1. Принимает электроны от НАДН

IV комплекс –
1. Принимает электроны от цитохрома с и передает
1. Принимает электроны от цитохрома с и передает
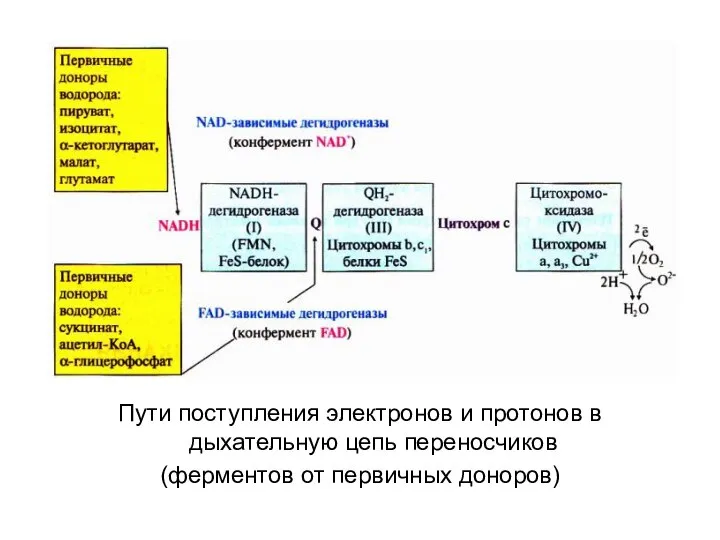
Пути поступления электронов и протонов в дыхательную цепь переносчиков
(ферментов
Пути поступления электронов и протонов в дыхательную цепь переносчиков
(ферментов

Работа комплексов дыхательной цепи
Ферменты дыхательной цепи расположены в строго
определенной последовательности:
Работа комплексов дыхательной цепи
Ферменты дыхательной цепи расположены в строго
определенной последовательности:

Стандартные окислительно-восстановительные потенциалы некоторых сопряженных пар
Стандартные окислительно-восстановительные потенциалы некоторых сопряженных пар

3. Здесь атомы водорода (от НАДН и ФАДН2)
передают свои электроны
3. Здесь атомы водорода (от НАДН и ФАДН2)
передают свои электроны

5. Перенос ионов водорода через мембрану (выкачивание)
происходит не случайно, а
происходит не случайно, а
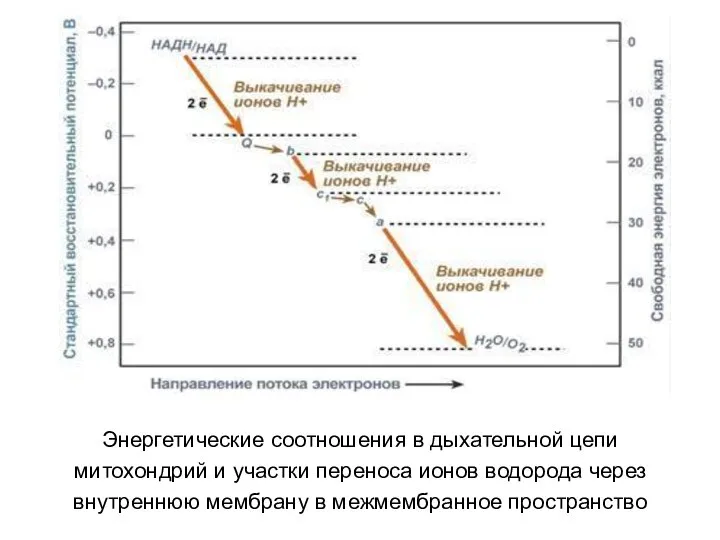
Энергетические соотношения в дыхательной цепи
митохондрий и участки переноса ионов водорода
Энергетические соотношения в дыхательной цепи
митохондрий и участки переноса ионов водорода
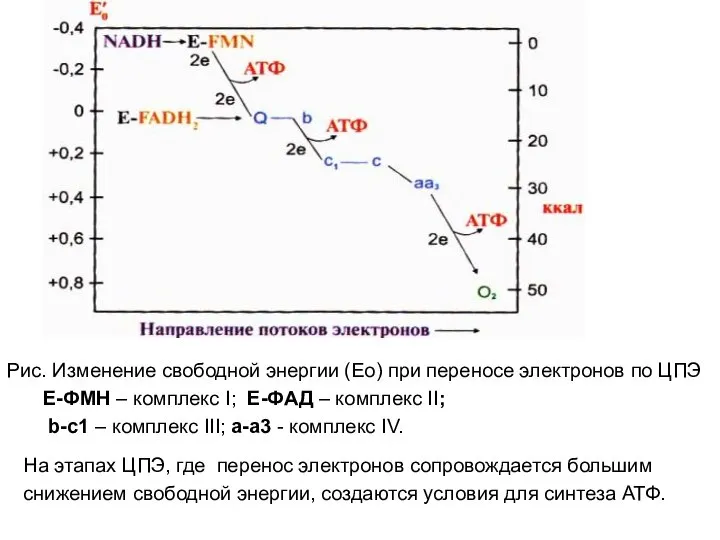
Рис. Изменение свободной энергии (Ео) при переносе электронов по ЦПЭ
Е-ФМН –
Рис. Изменение свободной энергии (Ео) при переносе электронов по ЦПЭ
Е-ФМН –

6. Как завершение всех предыдущих событий и
необходимый их результат происходит
необходимый их результат происходит

Сопряжение тканевого дыхания и
синтеза АТФ
1. Перенос электронов по ЦПЭ при
Сопряжение тканевого дыхания и
синтеза АТФ
1. Перенос электронов по ЦПЭ при
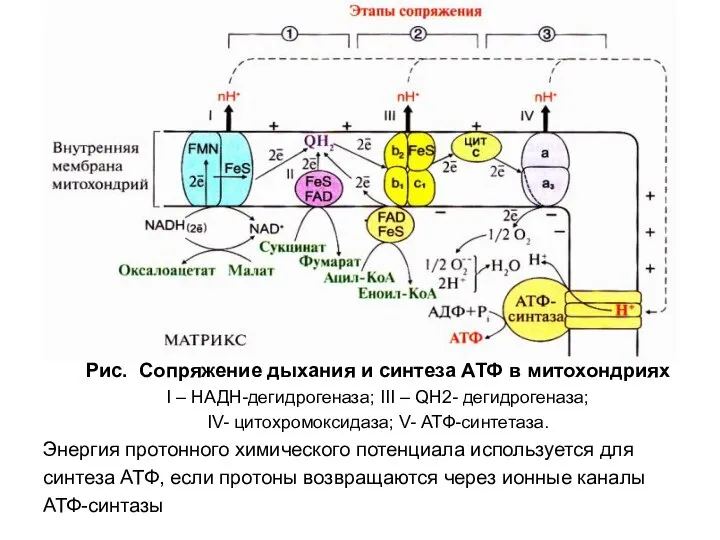
Рис. Сопряжение дыхания и синтеза АТФ в митохондриях
I – НАДН-дегидрогеназа; III
Рис. Сопряжение дыхания и синтеза АТФ в митохондриях
I – НАДН-дегидрогеназа; III

3. Каждый из трех комплексов ЦПЭ (I, III и IV )
3. Каждый из трех комплексов ЦПЭ (I, III и IV )

о Витамине С
Вит С
о Витамине С
Вит С
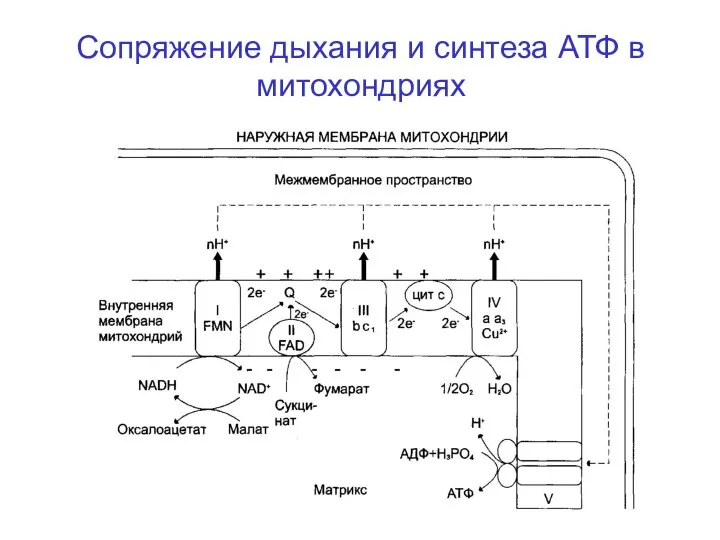
Сопряжение дыхания и синтеза АТФ в митохондриях
Сопряжение дыхания и синтеза АТФ в митохондриях

При участии АТФ-АДФ транслоказы, расположенной
во внутренней мембране митохондрий, АТФ
транспортируется
При участии АТФ-АДФ транслоказы, расположенной
во внутренней мембране митохондрий, АТФ
транспортируется
На всех этапах процесса часть энергии рассеивается в виде тепла

Аналоги АТФ:
ГТФ
ЦТФ
УТФ
Аналоги АТФ:
ГТФ
ЦТФ
УТФ

Дыхательный контроль
Перенос электронов по ЦПЭ и синтез АТФ тесно
сопряжены, т.е.
Дыхательный контроль
Перенос электронов по ЦПЭ и синтез АТФ тесно
сопряжены, т.е.

Синтез АТФ регулируется
Работа дыхательных ферментов регулируется с
помощью эффекта, который получил
Синтез АТФ регулируется
Работа дыхательных ферментов регулируется с
помощью эффекта, который получил

Возрастание протонного градиента возникает при
исчерпании запасов АДФ и накоплении АТФ,
Возрастание протонного градиента возникает при
исчерпании запасов АДФ и накоплении АТФ,

Таким образом скорость синтеза АТФ точно
соответствует потребностям клетки в энергии.
Ускорение
Таким образом скорость синтеза АТФ точно
соответствует потребностям клетки в энергии.
Ускорение

Разобщение дыхания и синтеза АТФ
Некоторые липофильные вещества (2,4-динитрофенол,
некоторые жирные кислоты) могут
Разобщение дыхания и синтеза АТФ
Некоторые липофильные вещества (2,4-динитрофенол,
некоторые жирные кислоты) могут

Терморегуляторная функция дыхания
При переносе электронов по ЦПЭ часть энергии
рассеивается в
Терморегуляторная функция дыхания
При переносе электронов по ЦПЭ часть энергии
рассеивается в

Кроме того дополнительное образование теплоты
может происходить путем разобщения дыхания и
фосфорилирования
Кроме того дополнительное образование теплоты
может происходить путем разобщения дыхания и
фосфорилирования

У новорожденных и зимнеспящих животных
разобщение связано с наличием разобщающего белка
У новорожденных и зимнеспящих животных
разобщение связано с наличием разобщающего белка
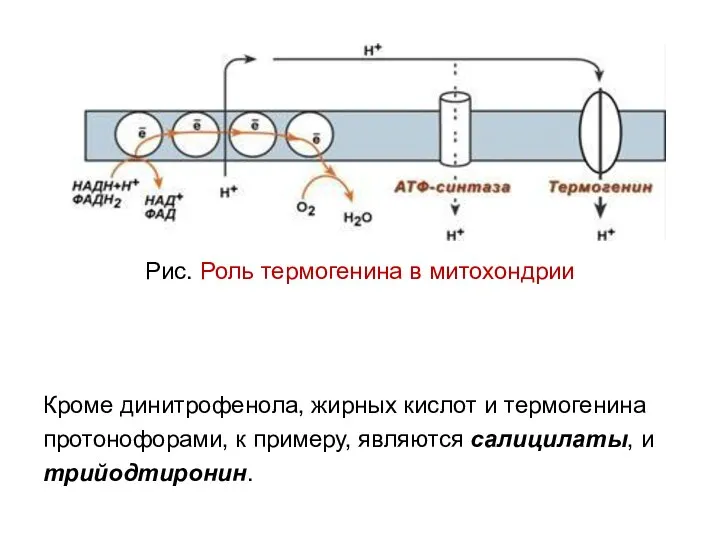
Рис. Роль термогенина в митохондрии
Кроме динитрофенола, жирных кислот и термогенина
протонофорами,
Рис. Роль термогенина в митохондрии
Кроме динитрофенола, жирных кислот и термогенина
протонофорами,

Ингибиторы дыхания
Некоторые лекарственные вещества, химические
агенты и антибиотики вызывают ингибирование
ферментов,
либо
Ингибиторы дыхания
Некоторые лекарственные вещества, химические
агенты и антибиотики вызывают ингибирование
ферментов,
либо
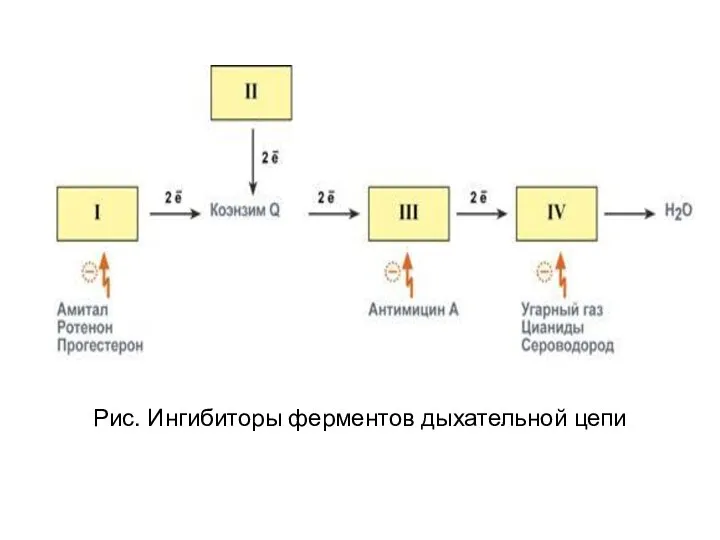
Рис. Ингибиторы ферментов дыхательной цепи
Рис. Ингибиторы ферментов дыхательной цепи

Гипоэнергетические состояния
Причинами гипоэнергетических состояний может быть следующее:
1. гиповитаминозы экзогенные
Гипоэнергетические состояния
Причинами гипоэнергетических состояний может быть следующее:
1. гиповитаминозы экзогенные

Состав компонентов дыхательной цепи
НАД содержит витамин В3 или РР (никотиновая кислота)
ФМН
Состав компонентов дыхательной цепи
НАД содержит витамин В3 или РР (никотиновая кислота)
ФМН
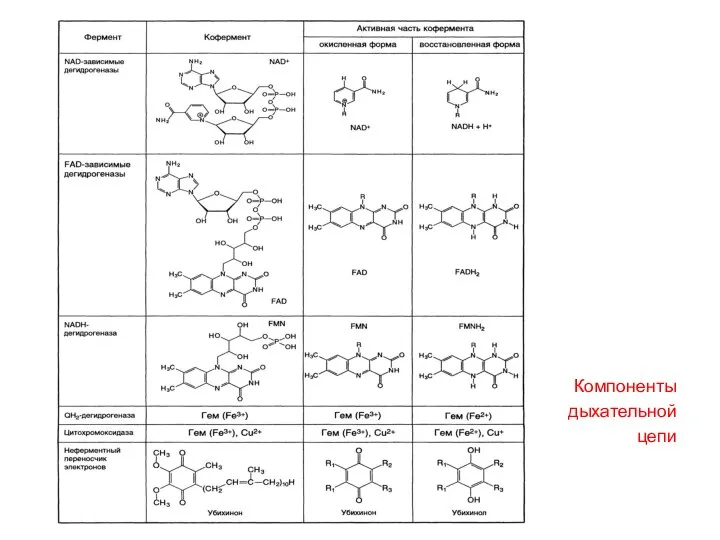
Компоненты
дыхательной
цепи
дыхательной
цепи
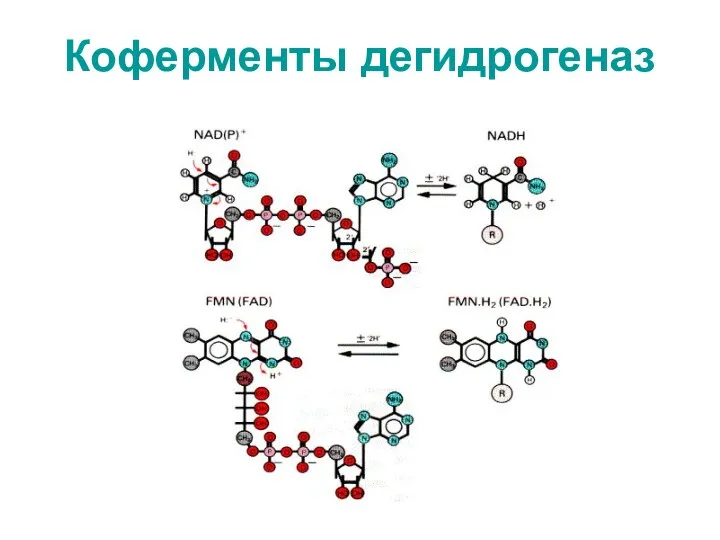
Коферменты дегидрогеназ
Коферменты дегидрогеназ
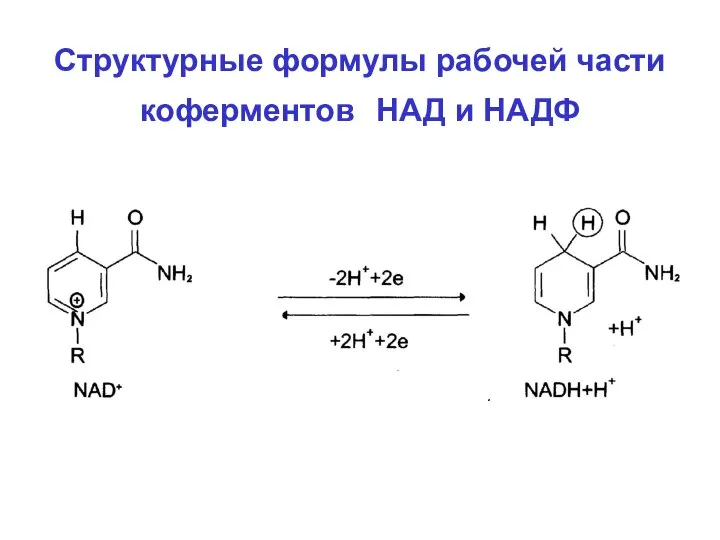
Структурные формулы рабочей части коферментов НАД и НАДФ
Структурные формулы рабочей части коферментов НАД и НАДФ
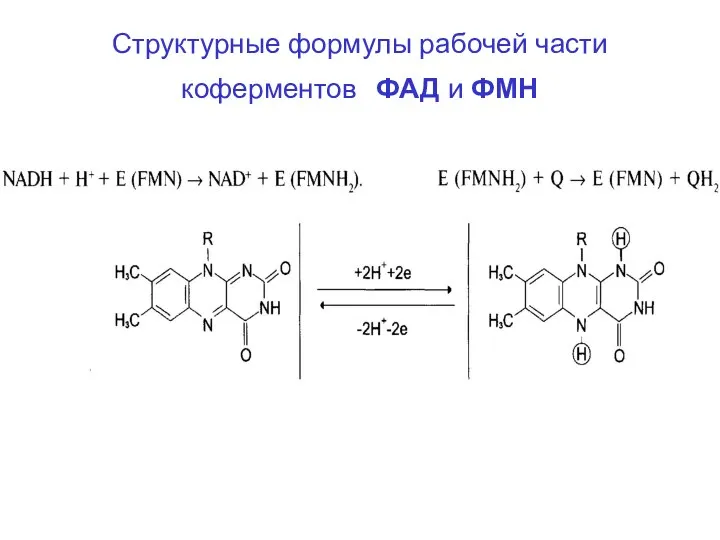
Структурные формулы рабочей части коферментов ФАД и ФМН
Структурные формулы рабочей части коферментов ФАД и ФМН
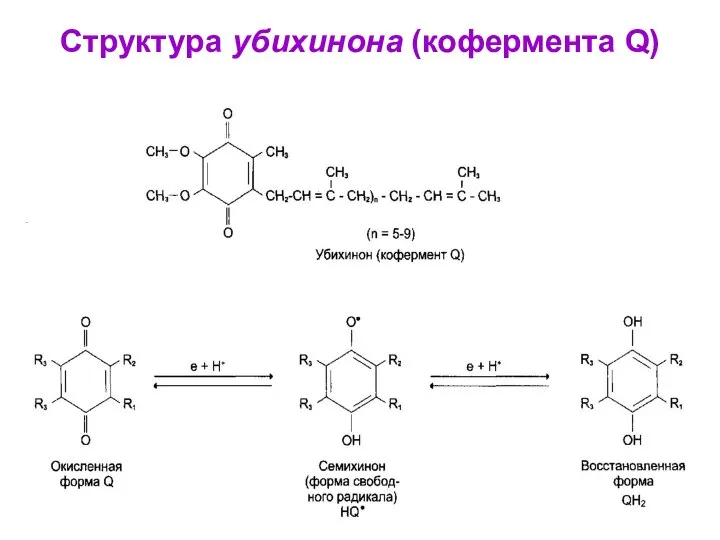
Структура убихинона (кофермента Q)
Структура убихинона (кофермента Q)
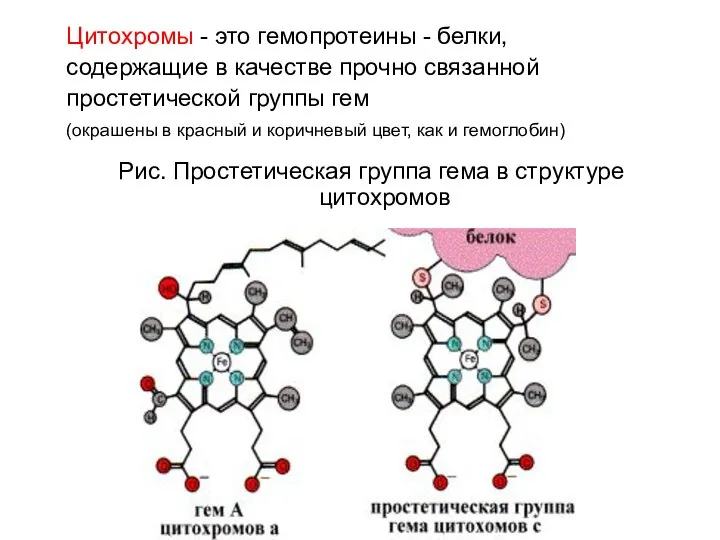
Цитохромы - это гемопротеины - белки,
содержащие в качестве прочно связанной
простетической группы
Цитохромы - это гемопротеины - белки,
содержащие в качестве прочно связанной
простетической группы
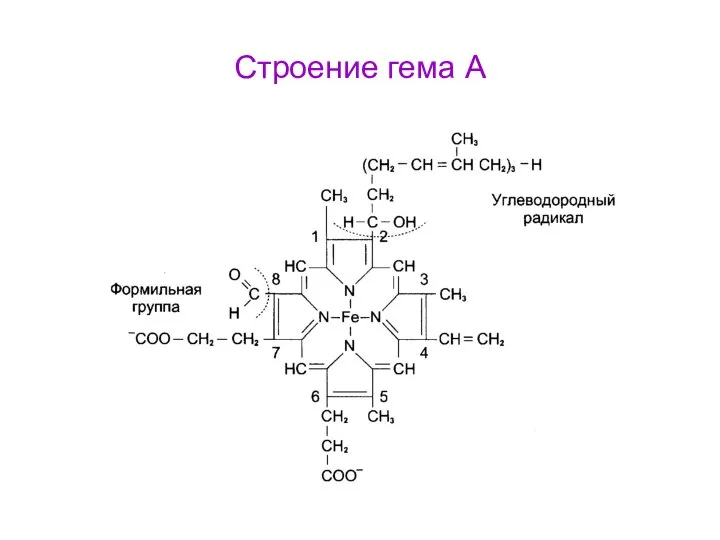
Строение гема А
Строение гема А

Атом железа в геме может менять
валентность, присоединяя или
отдавая электроны:
В
Атом железа в геме может менять
валентность, присоединяя или
отдавая электроны:
В

В цитохромах С и С1 дополнительные ковалентные
связи формируются между тиогруппами
В цитохромах С и С1 дополнительные ковалентные
связи формируются между тиогруппами

Цитохромоксидаза
включает комплекс цитохромов а и а3
(комплекс IV). Цитохромоксидаза кроме гема
Цитохромоксидаза
включает комплекс цитохромов а и а3
(комплекс IV). Цитохромоксидаза кроме гема
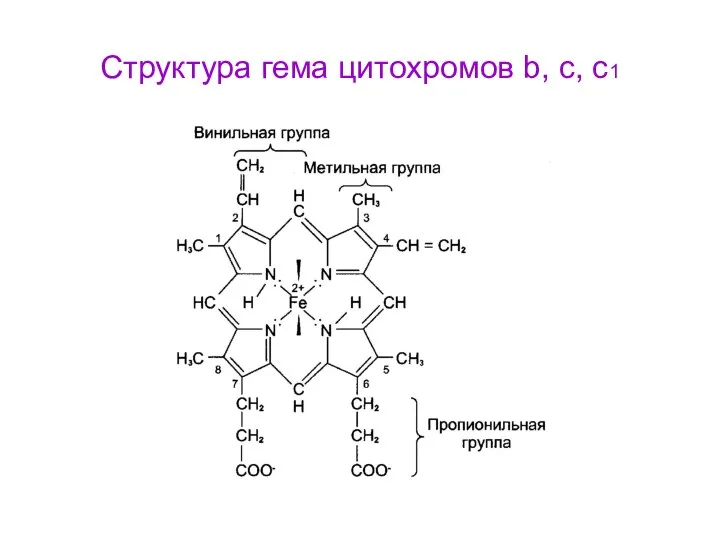
Структура гема цитохромов b, c, c1
Структура гема цитохромов b, c, c1
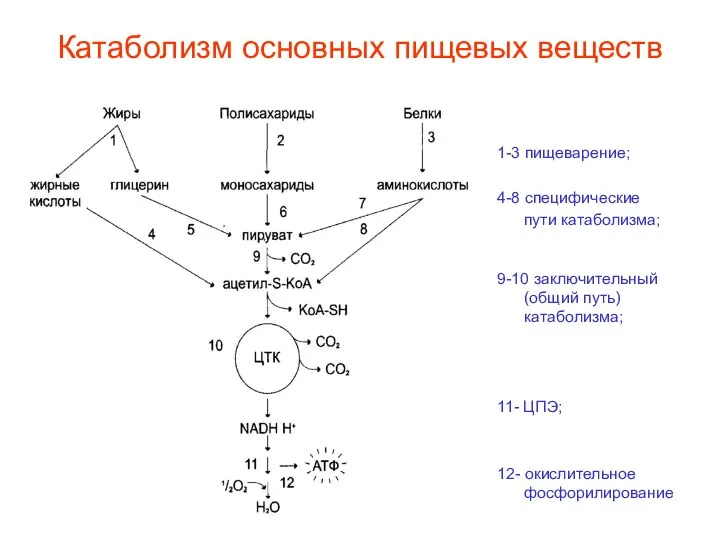
Катаболизм основных пищевых веществ
1-3 пищеварение;
4-8 специфические
пути катаболизма;
9-10 заключительный
Катаболизм основных пищевых веществ
1-3 пищеварение;
4-8 специфические
пути катаболизма;
9-10 заключительный
 Технология ремонта кузовов автомобиля
Технология ремонта кузовов автомобиля Вирусы гриппа Подготовила: Курганова Е., РЭ-2, 3 курс
Вирусы гриппа Подготовила: Курганова Е., РЭ-2, 3 курс Что делать в трудную минуту? Молодёжь Смоленской Центральной Церкви
Что делать в трудную минуту? Молодёжь Смоленской Центральной Церкви Домашняя мультимедиа-платформа с голосовым помощником Алисой
Домашняя мультимедиа-платформа с голосовым помощником Алисой Аналық тақша
Аналық тақша Basics of software development (continued)
Basics of software development (continued) Основы логики
Основы логики Электрические машины и электропривод. Введение
Электрические машины и электропривод. Введение Galileo galilei, February 15, 1564 - January 8, 1642
Galileo galilei, February 15, 1564 - January 8, 1642 Программирование. Экзамен
Программирование. Экзамен Устройства компьютера
Устройства компьютера Электрооборудование. Электроснабжение бортовой сети
Электрооборудование. Электроснабжение бортовой сети Весело колосится густая рожь.
Весело колосится густая рожь. Анималотерапия - презентация для начальной школы_
Анималотерапия - презентация для начальной школы_ Административное правонарушение Опрос
Административное правонарушение Опрос FORTRAN Название языка происходит от словосочетания FORmulae TRANslation — преобразование формул
FORTRAN Название языка происходит от словосочетания FORmulae TRANslation — преобразование формул Государственный Дарвиновский музей
Государственный Дарвиновский музей Типология и классификация политической культуры
Типология и классификация политической культуры Скульптурные шедевры Л.Бернини
Скульптурные шедевры Л.Бернини Ватто Антуан
Ватто Антуан Теория ландшафтной архитектуры и методология проектирования
Теория ландшафтной архитектуры и методология проектирования Презентацию выполнила студентка – заочница 2 курса филологического ф – та Купчихина Елена Николаевна Владимир, 2013 год
Презентацию выполнила студентка – заочница 2 курса филологического ф – та Купчихина Елена Николаевна Владимир, 2013 год Общая эндокринология
Общая эндокринология  Цивилизация и культура
Цивилизация и культура Political parties in Estonia
Political parties in Estonia Медный, бронзовый, железный века на Луганщине
Медный, бронзовый, железный века на Луганщине Методика работы Учителя физической культуры СОШ № 80 Чигилейчик Лады Юрьевны
Методика работы Учителя физической культуры СОШ № 80 Чигилейчик Лады Юрьевны Ноосфера . Эволюция представлений о месте человека в природе
Ноосфера . Эволюция представлений о месте человека в природе