- Биотехнология растений. Трансгенные растения (часть 4)

Содержание
- 2. Лекция 6: Трансгенные растения-4 Получение инсерционных мутантов растений на основе агробактериальной трансформации Клонирование генов Изучение с
- 3. Т-ДНК инсерционный мутагенез интеграция Т-ДНК изменение структуры одного из генов за счет встраивания Т-ДНК… ….отбор по
- 4. Схемы используемых плазмид для получения: рецессивных мутаций доминантных мутаций X - усилители транскрипции из промотора 35S
- 5. Клонирование генов
- 6. Клонирование гена agamous с помощью Т-ДНК инсерционного мутагенеза ЭМС-мутагенез 10 лепестков, 4 чашелистика агробактериальная трансформация семян
- 7. Ген, маркированный транспозоном (tagged gene), можно клонировать с помощью «вытягивания за транспозон»
- 8. Транспозонный мутагенез интеграция изменение структуры ….изменение транспозона одного из генов…. фенотипа Возможность клонировать ген, «вытянув» его
- 9. При встраивании транспозона в кодирующую последовательность гена, наблюдается мутантный фенотип. При вырезании фенотип восстанавливается
- 10. Клонирование генов на основе транспозонного (инсерционного) мутагенеза
- 11. Клонирование генов на основе транспозонного (инсерционного) мутагенеза
- 12. Получение мутации flo613 и клонирование гена FLORICAULA львиного зева (Coen et al., 1990) Выращивание 26 000
- 13. Рестрикционный анализ ДНК из выделенных клонов, необходимый для выявления участков растительной ДНК, фланкирующих транспозон Субклонирование растительной
- 14. Создание геномной библиотеки из мутанта flo 613, выращиваемого при tº=25ºC (Тam3 не перемещается)
- 15. Скрининг библиотеки с меченой пробой из Tam3 для выявления клонов, содержащих транспозон Получение отпечатков клонов библиотеки
- 16. Д.т. flo 613 9.0 5.5 1.5 Сравнение рестрикционных карт клонов, содержащих ген FLO, из геномной библиотеки
- 17. Ген FLORICAULA – уникальный ген, присутствующий в геноме покрытосеменных в 1 копии. Он кодирует белок, содержащий
- 18. Изучение механизмов регуляции экспрессии генов растений с использованием репортерных (индикаторных) генов Возможности метода: Выявление факторов (внешних
- 19. Слитые гены-репортеры К промотору исследуемого гена присоединяют репортерный ген
- 20. Green fluorescent protein (GFP) Luciferase (LUC) ß-glucuronidase (GUS) Promoter-reporter fusion: Использование гибридных конструкций, содержащих репортерный ген,
- 21. В трансформированных растениях проводят анализ экспрессии репротерного гена, отбирают трансформантов с искомым характером экспрессии
- 22. Анализ распределение ауксина в растениях с помощью генетической конструкции DR5:GUS DR5 – синтетический ауксин-регулируемый промотор
- 23. Анализ активности промоторов ключевых регуляторных генов в меристемах растений
- 24. Получение растений со сверхэкспрессией гена с целью изучения функции гена
- 25. Фенотип трансгенных растений 35S::LFY У растений 35S::LFY ген LFY экспрессируется и в АМ, что приводит к
- 26. Характер экспрессии генов-ортологов LFY/FLO может определять тип соцветия (открытое или закрытое)
- 27. Метод позволяет изучить регуляторную функцию любого участка гена Направленный мутагенез промоторной области гена LFY позволил обнаружить
- 28. Мутант арабидопсиса по гену LFY
- 29. Основные семейства транскрипционных факторов растений Консервативные для эукариот: Уникальные для растений: ТФ с MADS-доменом ТФ с
- 30. Гомеодомен-ДНК - комплекс вариабельный уч. гомеодомен N С Структура транскрипционного фактора с ДНК-связывающим гомеодоменом
- 31. Гены семейств KNOX (STM) и WOX (WUS) кодируют гомеодомен-содержащие транскрипционные факторы Гены семейства KNOX арабидопсиса Гены
- 32. ТФ с гомеодоменами (семейство TALE) KNOX мишени: гены биосинтеза ЦК и ГК программы развития: 1). Развитие
- 33. Ген KNOTTED1 был идентифи-цирован на основе доминантных мутаций kn1, вызывающих отсутствие лигулы и появление узелков (knots)
- 34. Зная о важной роли гомеобоксных генов в развитии животных, приступили к клонированию и изучению KNOTTED1-подобных генов
- 35. Сверхэкспрессия гена KNAT1 у арабидопсиса На листьях возникали дополнительные меристематические очаги, из которых могли возникать почки
- 36. Сверхэкспрессия гена KNAT в листьях томатов дикий тип 35S::KNAT
- 37. KNOTTED-подобные гены – возможные участники морфологической эволюции растений
- 38. Обнаружение связи между уровнем экспрессии KNOX-генов и усложнением структуры листа позволило предположить, что эти гены могли
- 39. MCM1 – регулятор транскрипции дрожжей AG – гомеозисный ген арабидопсиса (класс С) DEF –гомеозисный ген львиного
- 40. ТФ с MADS доменом вышележащие регуляторы: ТФ LFY мишени: ??? программы развития: развитие органов цветка AP1,
- 41. Исследования трансгенных растений с конститутивной экспрессией гомеозисных генов также доказывают верность АВС-модели 35S::AP3 35S::PI 35S::AP3 35S::PI
- 42. Цветки с двойным венчиком (лилейные) Цветки с двойной чашечкой Мутации в АВС- генах приводят к формирова-нию
- 43. Использование методов «генетической хирургии» для изучения взаимодействия клеток и тканей в процессах развития
- 44. Day, Galgoci, Irish, 1995. Генетическое удаление лепестков и тычинок для выяснения роли клеточных взаимодействий в развитии
- 45. «Пространственная» модель Holder, 1979. Под влиянием позиционных сигналов происходит разметка ФМ, после которой идет независимая дифференцировка
- 46. В основе «генетической хирургии» лежит метод трансформации растений химерными генами следующей конструкции: Кодирующая часть гена –
- 47. гены, кодирующие рибонуклеазы: Т1 рибонуклеазу Aspergillus oryzae и барназу Bacillus amyloliquefaciens, вызывающие внутриклеточную деградацию РНК; ген
- 48. Целью работы Day, Galgoci, Irish (1995) была проверка существующих моделей с использованием метода генетической хирургии Объекты:
- 49. Этапы работы Слитые гены включали в Т-ДНК область (фрагмент Ti-плазмиды,который передается растению) бинарного вектора pBIADN и
- 50. У резушки получено 5 трансгенных растений, в геноме которых присутствовали от 1 до 5 копий трансгенов
- 51. Использование трансформации для изучения роли гормонов в развитиии растений
- 53. Корончатый галл на растении табака как результат трансформации агробактерией (штамм дикого типа)
- 54. Трансгенные рстения с геном tmr
- 55. Ген tmr в трансгенных растениях, экспрессирующийся в плодах (промотор 2А11)
- 56. Опухолеобразование у редиса – генетический признак
- 57. Фенокопии опухолеобразования у трансгенных растений с геном ipt
- 58. Прямая генетика: фенотип ген Обратная генетика: ген фенотип Прямая и обратная генетика в изучении функции гена
- 59. - гомологичная рекомбинация/замещение гена -замещение гена: альтернативные подходы (Zn-fingers nucleases, TALEN) - РНК-интерференция/ генный сайленсинг -
- 60. TALE: TAL (transcription activator-like) effectors Эффекторы TALE – регуляторные белки, секретируются бактериями Xanthomonas (патогены растений) с
- 61. Основные типы замолкания гена (сайленсинга) пост-транскрипционный сайленсинг (регуляция экспресии гена на уровне трансляции) PTGS - Post-transcriptional
- 62. Белки, участвующие в метаболизме «малых» РНК у растений DCL1 (DICER-like) – рибонулеказа, участвующая в процессинге miRNA
- 63. Пример: выключение (сайленсинг) гена «икс» конститутивный (35S) регулируемый (индуцибельный) Интроны промотор икс ски dsRNA RISC –
- 64. Регулируемое замолкание гена PDS (PHYTOENE DEASATURASE) LB RB RB LB 35S LhGR GUS op PDS PDS
- 65. Регулируемое замолкание (индуцибельный сайленсинг) гена PDS (PHYTOENE DEASATURASE) No Dex Dex
- 66. Enhancer TATA En En En En Enhancer TATA Enhancer TATA 3. Enhancer trap (ловушка для энхансера)
- 67. Инсерционный мутагенез с использованием «ловушек» промоторов Репортерный ген без промотора встраивают в Т-ДНК векторной Ti-плазмиды. Растения
- 68. Enhancer TATA 4. Ловушка для промотора (promoter trap) -1 reporter gene гибридный белок Инсерция в экзон
- 69. Принцип работы «ловушки для энхансеров» GAL4:GFP Примеры использования Т-ДНК в качестве «зондов» для выявления тканеспецифичных энхансеров
- 70. Методы редактирования генома растений
- 71. Модификации генома: разновидности Изменение случайного места в геноме Редактирование генома
- 72. Редактирование генома Инактивация конкретного локуса Замена одного локуса на другой с помощью гомологичной рекомбинации (HR) Этот
- 73. Программируемые нуклеазы ZFN (Zink-Finger Nucleases) TALENs (Transcription Activator-Like Effector Nucleases) CRISPR-Cas система (Clustered regularly Interspaced Palindromic
- 74. Нуклеазы с цинковыми пальцами В качестве ДНК-связывающего домена используется универсальный домен типа «Цинковые пальцы»
- 76. Как программируют ZFN? Wolfe et al., 1999 Каждый цинковый палец распознаёт 3 (4) нуклеотида. Обычно используют
- 77. TALEN (Transcription Activator-Like Effectors Nucleases)
- 78. TALEN Kim, Kim, 2014 33-35 аминокислотные повторы 12ый -13ый аминокислотные остатки (repeat variable diresidues) связываются с
- 79. Применение TALEN Потеря функции гена LOX3 (кодирует липоксигеназу, создающую перекись водорода) увеличивает сроки хранения семян Этот
- 80. Система CRISPR/Cas9 (Clustered regularly Interspaced Palindromic Repeats- CRISPR associated protein)
- 81. Система CRISPR/Cas9 В 2005 обнаружили, что спейсеры гомологичны последовательностям фагов и конъюгативных плазмид (Mojica et al.,
- 82. Работа системы CRISPR-Cas в бактериях Horvath, Barrangou, 2010
- 83. Čermák et al., 2015
- 84. Защита от вирусов с помощью CRISPR-Cas9 Ji et al., 2015
- 85. Рецептор ауксина Auxin binding protein 1 (ABP1) Chen et al., 2001
- 86. Gao et al., 2015
- 88. Скачать презентацию

Лекция 6: Трансгенные растения-4
Получение инсерционных мутантов растений на основе агробактериальной трансформации
Клонирование
Лекция 6: Трансгенные растения-4
Получение инсерционных мутантов растений на основе агробактериальной трансформации
Клонирование
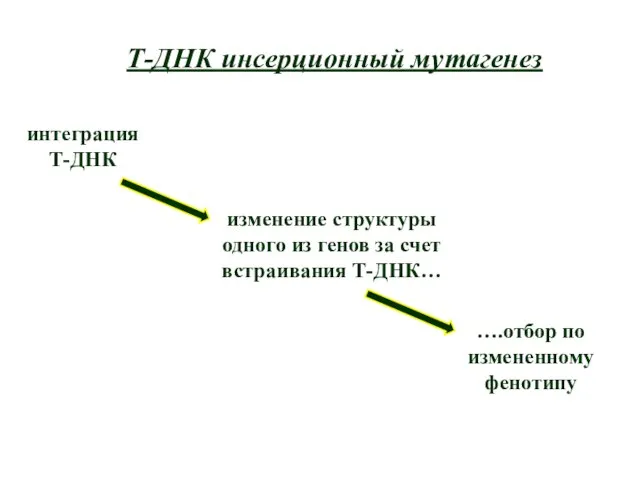
Т-ДНК инсерционный мутагенез
интеграция Т-ДНК
изменение структуры одного из генов за счет встраивания
Т-ДНК инсерционный мутагенез
интеграция Т-ДНК
изменение структуры одного из генов за счет встраивания
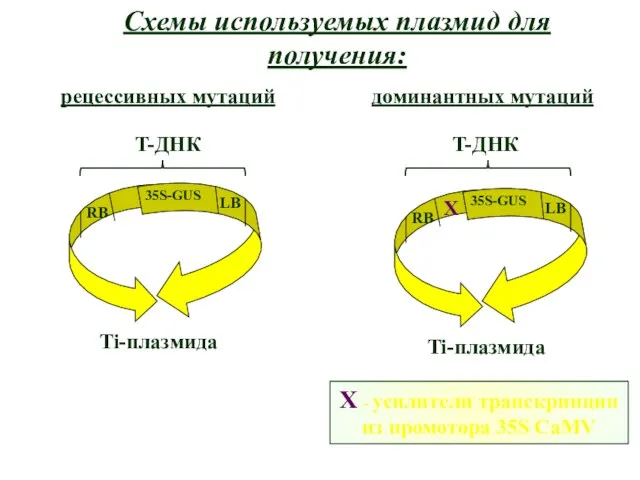
Схемы используемых плазмид для получения:
рецессивных мутаций
доминантных мутаций
X - усилители транскрипции из
Схемы используемых плазмид для получения:
рецессивных мутаций
доминантных мутаций
X - усилители транскрипции из

Клонирование генов
Клонирование генов
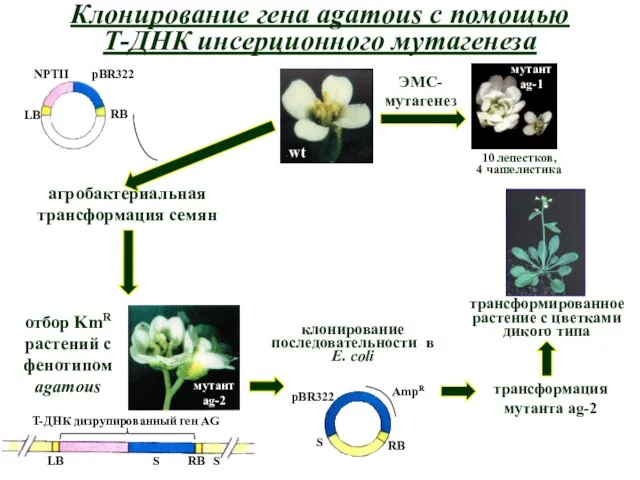
Клонирование гена agamous с помощью Т-ДНК инсерционного мутагенеза
ЭМС-мутагенез
10 лепестков,
4 чашелистика
агробактериальная
Клонирование гена agamous с помощью Т-ДНК инсерционного мутагенеза
ЭМС-мутагенез
10 лепестков,
4 чашелистика
агробактериальная

Ген, маркированный транспозоном (tagged gene), можно клонировать с помощью «вытягивания за
Ген, маркированный транспозоном (tagged gene), можно клонировать с помощью «вытягивания за
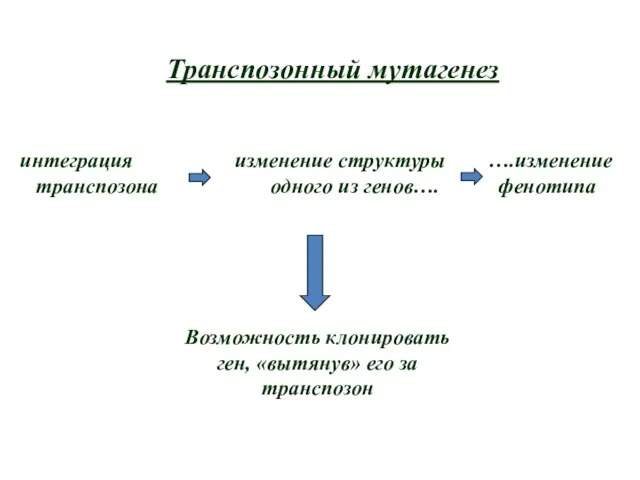
Транспозонный мутагенез
интеграция изменение структуры ….изменение транспозона одного из генов…. фенотипа
Возможность клонировать
ген,
Транспозонный мутагенез
интеграция изменение структуры ….изменение транспозона одного из генов…. фенотипа
Возможность клонировать
ген,

При встраивании транспозона в кодирующую последовательность гена, наблюдается мутантный фенотип.
При
При встраивании транспозона в кодирующую последовательность гена, наблюдается мутантный фенотип.
При
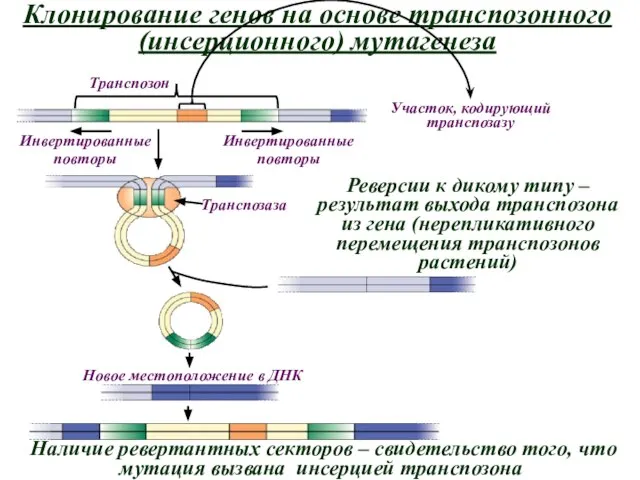
Клонирование генов на основе транспозонного (инсерционного) мутагенеза
Клонирование генов на основе транспозонного (инсерционного) мутагенеза

Клонирование генов на основе транспозонного (инсерционного) мутагенеза
Клонирование генов на основе транспозонного (инсерционного) мутагенеза

Получение мутации flo613 и клонирование гена FLORICAULA львиного зева
(Coen et
Получение мутации flo613 и клонирование гена FLORICAULA львиного зева (Coen et

Рестрикционный анализ ДНК из выделенных клонов, необходимый для выявления участков
Рестрикционный анализ ДНК из выделенных клонов, необходимый для выявления участков
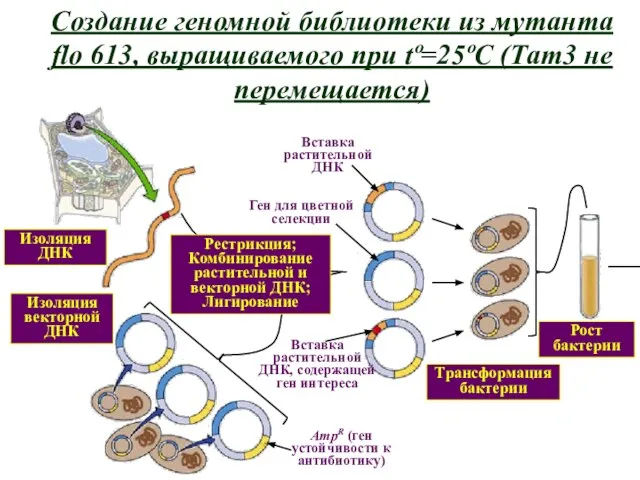
Создание геномной библиотеки из мутанта flo 613, выращиваемого при tº=25ºC (Тam3
Создание геномной библиотеки из мутанта flo 613, выращиваемого при tº=25ºC (Тam3
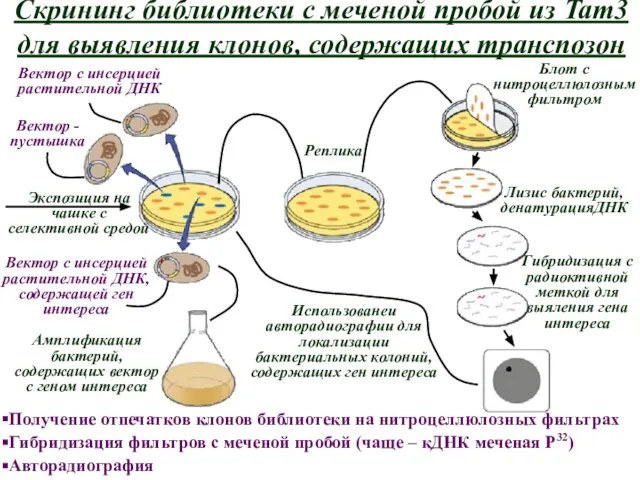
Скрининг библиотеки с меченой пробой из Tam3 для выявления клонов, содержащих
Скрининг библиотеки с меченой пробой из Tam3 для выявления клонов, содержащих
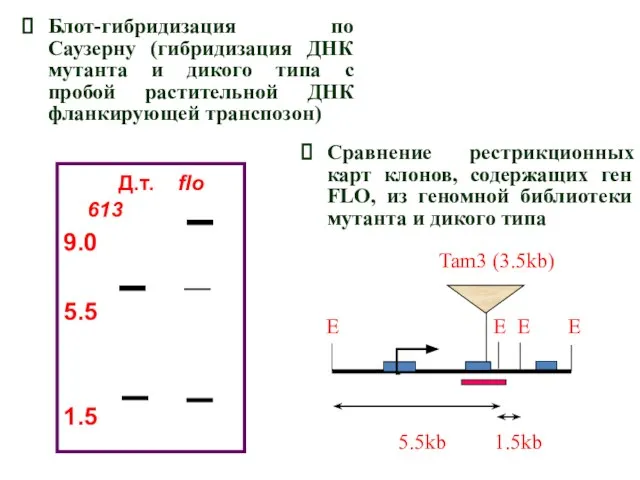
Д.т. flo 613
9.0
5.5
1.5
Сравнение рестрикционных карт клонов, содержащих ген FLO, из
Д.т. flo 613
9.0
5.5
1.5
Сравнение рестрикционных карт клонов, содержащих ген FLO, из

Ген FLORICAULA – уникальный ген, присутствующий в геноме покрытосеменных в 1
Ген FLORICAULA – уникальный ген, присутствующий в геноме покрытосеменных в 1

Изучение механизмов регуляции экспрессии генов растений с использованием репортерных (индикаторных)
Изучение механизмов регуляции экспрессии генов растений с использованием репортерных (индикаторных)
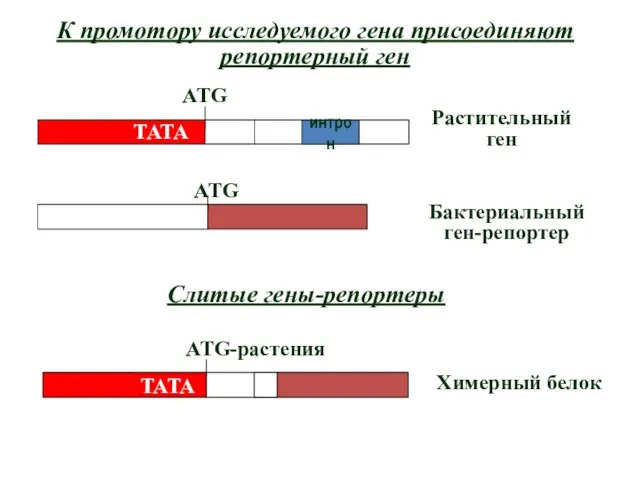
Слитые гены-репортеры
К промотору исследуемого гена присоединяют репортерный ген
Слитые гены-репортеры
К промотору исследуемого гена присоединяют репортерный ген

Green fluorescent protein (GFP)
Luciferase (LUC)
ß-glucuronidase (GUS)
Promoter-reporter fusion:
Использование гибридных конструкций, содержащих
репортерный
Green fluorescent protein (GFP)
Luciferase (LUC)
ß-glucuronidase (GUS)
Promoter-reporter fusion:
Использование гибридных конструкций, содержащих
репортерный
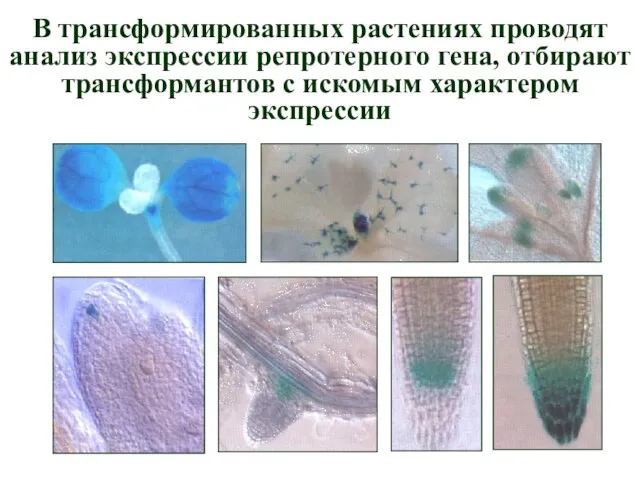
В трансформированных растениях проводят анализ экспрессии репротерного гена, отбирают трансформантов с
В трансформированных растениях проводят анализ экспрессии репротерного гена, отбирают трансформантов с
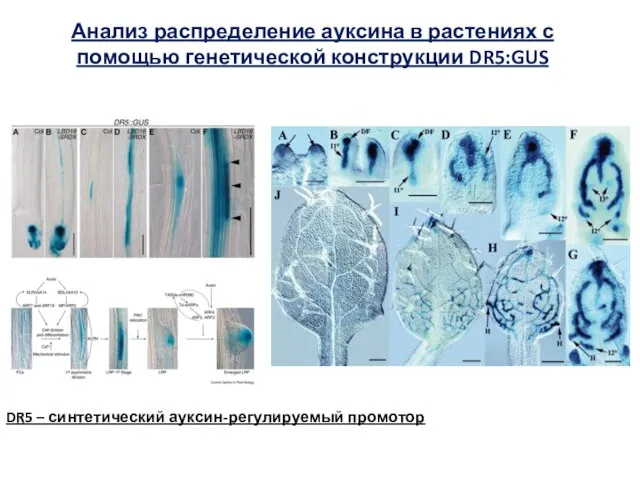
Анализ распределение ауксина в растениях с помощью генетической конструкции DR5:GUS
DR5 –
Анализ распределение ауксина в растениях с помощью генетической конструкции DR5:GUS
DR5 –
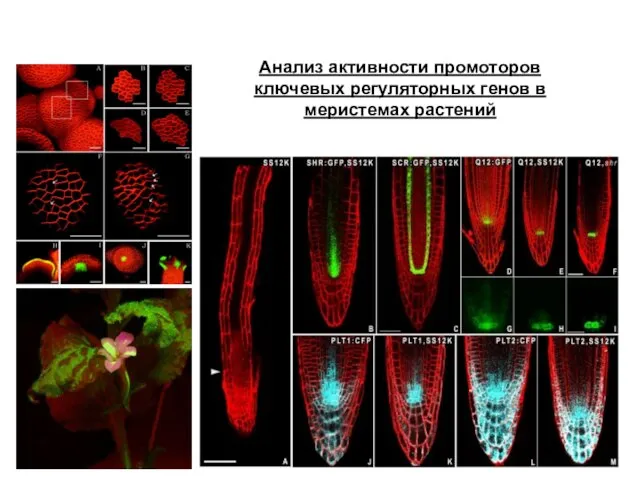
Анализ активности промоторов ключевых регуляторных генов в меристемах растений
Анализ активности промоторов ключевых регуляторных генов в меристемах растений
Получение растений со сверхэкспрессией гена с целью изучения функции гена
Получение растений со сверхэкспрессией гена с целью изучения функции гена
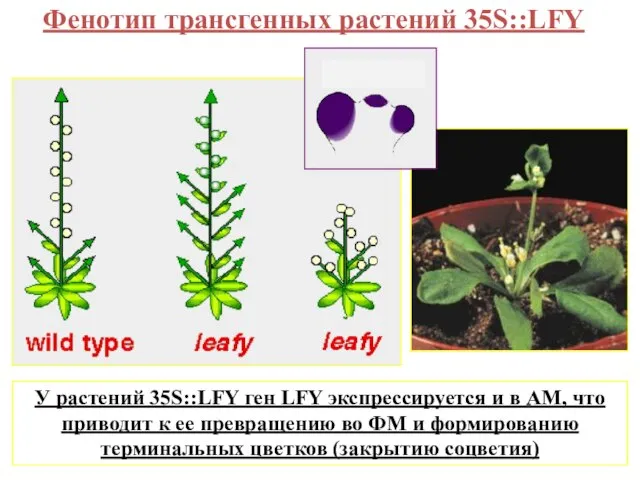
Фенотип трансгенных растений 35S::LFY
У растений 35S::LFY ген LFY экспрессируется и в
Фенотип трансгенных растений 35S::LFY
У растений 35S::LFY ген LFY экспрессируется и в
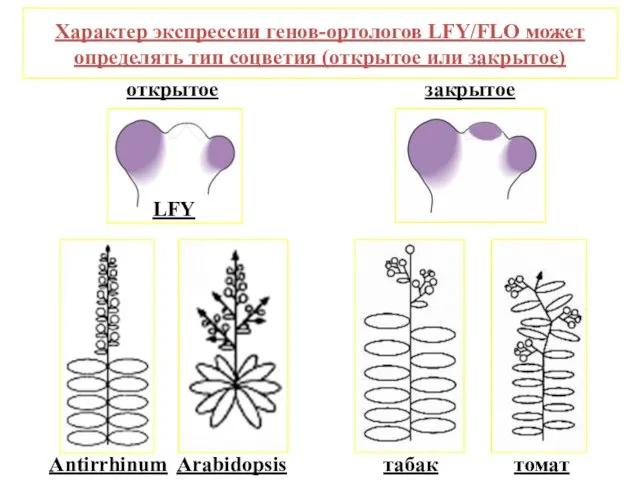
Характер экспрессии генов-ортологов LFY/FLO может определять тип соцветия (открытое или закрытое)
Характер экспрессии генов-ортологов LFY/FLO может определять тип соцветия (открытое или закрытое)
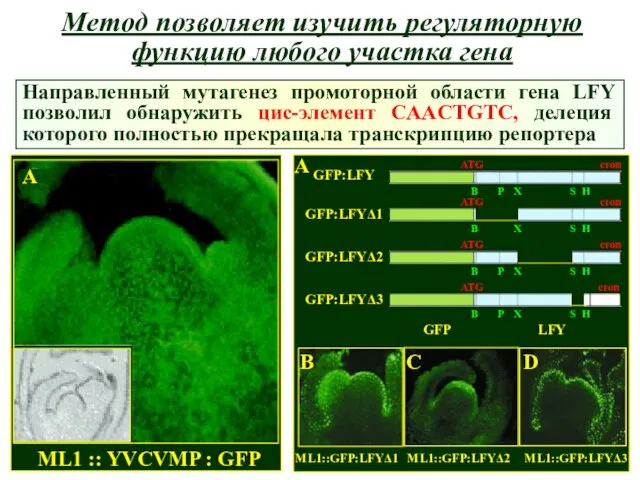
Метод позволяет изучить регуляторную функцию любого участка гена
Направленный мутагенез промоторной области
Метод позволяет изучить регуляторную функцию любого участка гена
Направленный мутагенез промоторной области

Мутант арабидопсиса по гену LFY
Мутант арабидопсиса по гену LFY

Основные семейства транскрипционных факторов растений
Консервативные для эукариот:
Уникальные для растений:
ТФ с MADS-доменом
ТФ
Основные семейства транскрипционных факторов растений
Консервативные для эукариот:
Уникальные для растений:
ТФ с MADS-доменом
ТФ
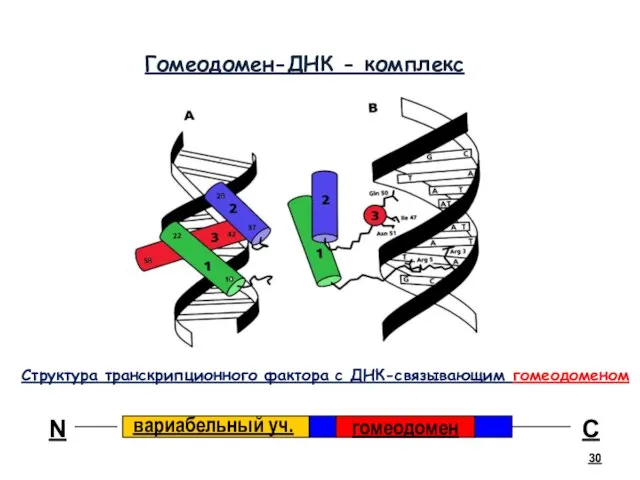
Гомеодомен-ДНК - комплекс
вариабельный уч.
гомеодомен
N
С
Структура транскрипционного фактора с ДНК-связывающим гомеодоменом
Гомеодомен-ДНК - комплекс
вариабельный уч.
гомеодомен
N
С
Структура транскрипционного фактора с ДНК-связывающим гомеодоменом
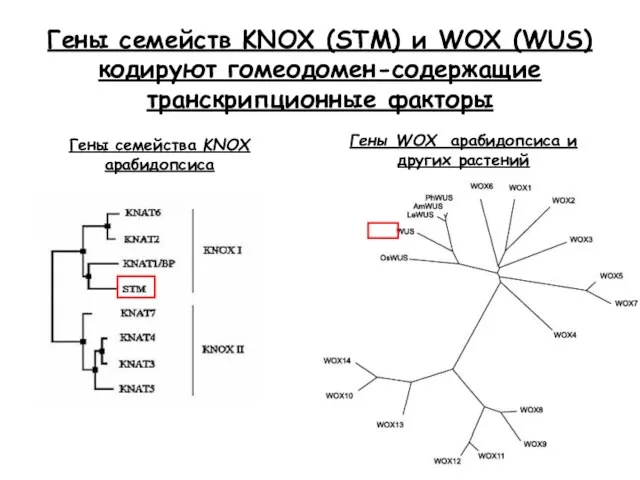
Гены семейств KNOX (STM) и WOX (WUS) кодируют гомеодомен-содержащие транскрипционные факторы
Гены
Гены семейств KNOX (STM) и WOX (WUS) кодируют гомеодомен-содержащие транскрипционные факторы
Гены
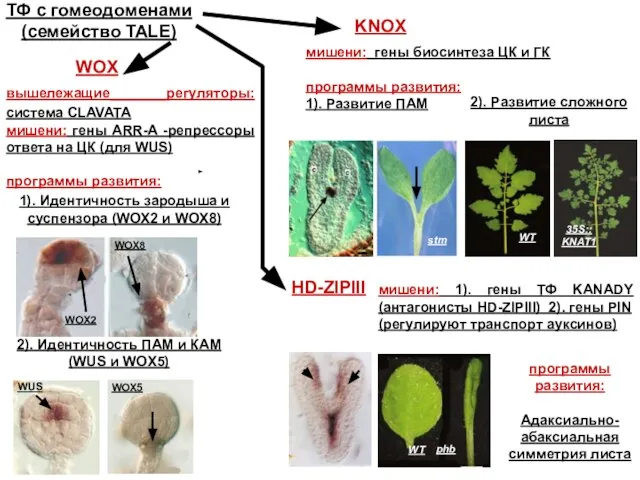
ТФ с гомеодоменами
(семейство TALE)
KNOX
мишени: гены биосинтеза ЦК и ГК
программы развития:
1). Развитие
ТФ с гомеодоменами
(семейство TALE)
KNOX
мишени: гены биосинтеза ЦК и ГК
программы развития:
1). Развитие

Ген KNOTTED1 был идентифи-цирован на основе доминантных мутаций kn1, вызывающих отсутствие
Ген KNOTTED1 был идентифи-цирован на основе доминантных мутаций kn1, вызывающих отсутствие

Зная о важной роли гомеобоксных генов в развитии животных, приступили к
Зная о важной роли гомеобоксных генов в развитии животных, приступили к

Сверхэкспрессия гена KNAT1 у арабидопсиса
На листьях возникали дополнительные меристематические очаги, из
Сверхэкспрессия гена KNAT1 у арабидопсиса
На листьях возникали дополнительные меристематические очаги, из

Сверхэкспрессия гена KNAT в листьях томатов
дикий тип
35S::KNAT
Сверхэкспрессия гена KNAT в листьях томатов
дикий тип
35S::KNAT

KNOTTED-подобные гены – возможные участники морфологической эволюции растений
KNOTTED-подобные гены – возможные участники морфологической эволюции растений

Обнаружение связи между уровнем экспрессии KNOX-генов и усложнением структуры листа позволило
Обнаружение связи между уровнем экспрессии KNOX-генов и усложнением структуры листа позволило
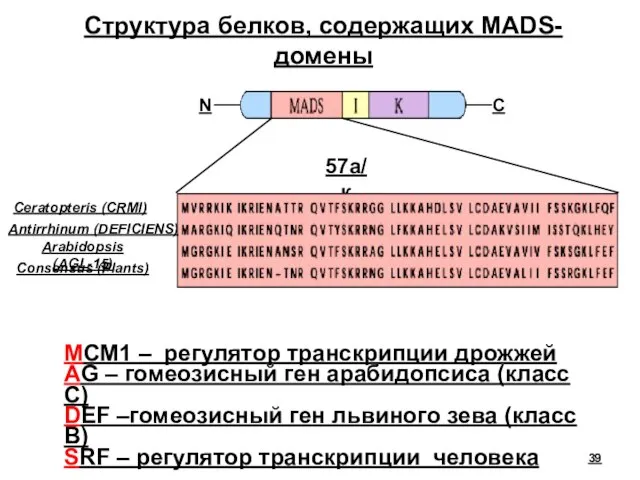
MCM1 – регулятор транскрипции дрожжей
AG – гомеозисный ген арабидопсиса (класс С)
DEF
MCM1 – регулятор транскрипции дрожжей
AG – гомеозисный ген арабидопсиса (класс С)
DEF
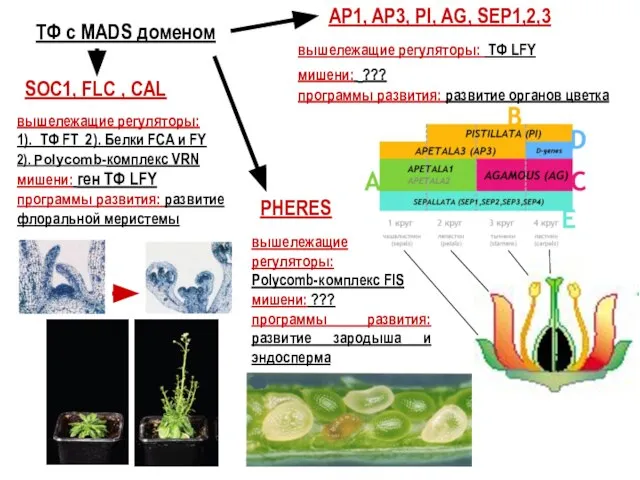
ТФ с MADS доменом
вышележащие регуляторы: ТФ LFY
мишени: ???
программы развития: развитие
ТФ с MADS доменом
вышележащие регуляторы: ТФ LFY
мишени: ???
программы развития: развитие
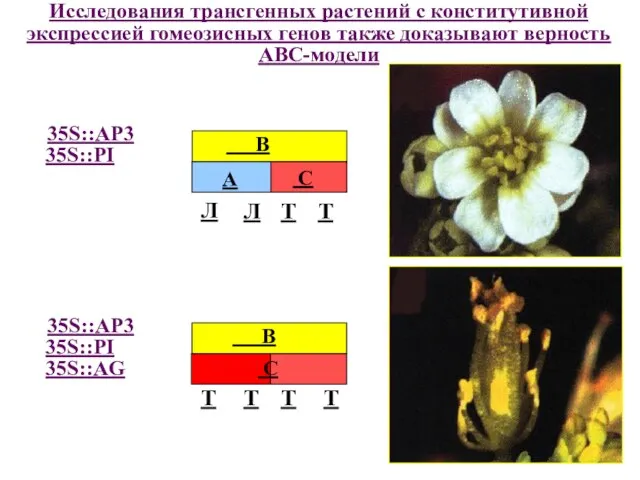
Исследования трансгенных растений с конститутивной экспрессией гомеозисных генов также доказывают верность
Исследования трансгенных растений с конститутивной экспрессией гомеозисных генов также доказывают верность
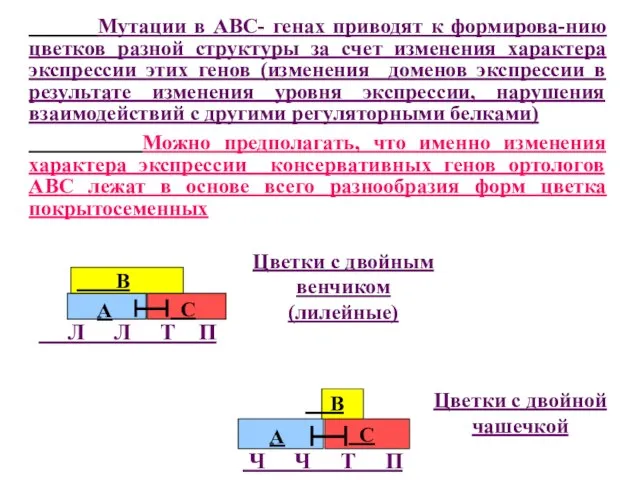
Цветки с двойным венчиком (лилейные)
Цветки с двойной чашечкой
Мутации в АВС-
Цветки с двойным венчиком (лилейные)
Цветки с двойной чашечкой
Мутации в АВС-
Использование методов «генетической хирургии» для изучения взаимодействия клеток и тканей в
Использование методов «генетической хирургии» для изучения взаимодействия клеток и тканей в

Day, Galgoci, Irish, 1995.
Генетическое удаление лепестков и тычинок для выяснения
Day, Galgoci, Irish, 1995. Генетическое удаление лепестков и тычинок для выяснения
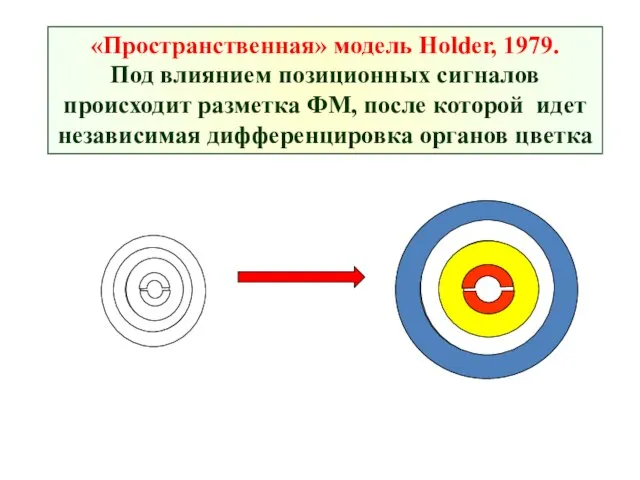
«Пространственная» модель Holder, 1979.
Под влиянием позиционных сигналов происходит разметка ФМ,
«Пространственная» модель Holder, 1979. Под влиянием позиционных сигналов происходит разметка ФМ,

В основе «генетической хирургии» лежит метод трансформации растений химерными генами следующей
В основе «генетической хирургии» лежит метод трансформации растений химерными генами следующей
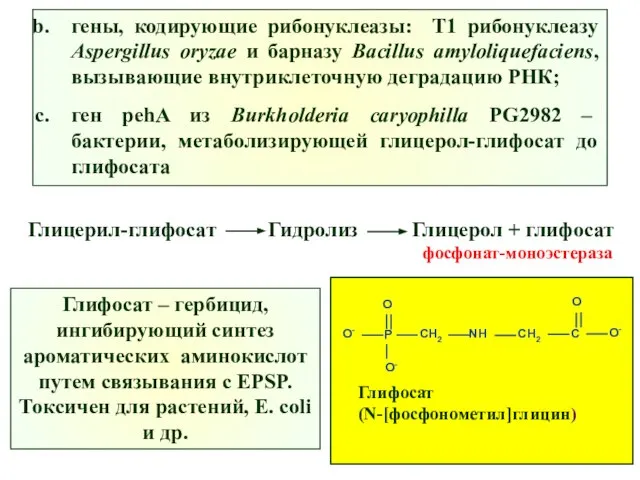
гены, кодирующие рибонуклеазы: Т1 рибонуклеазу Aspergillus oryzae и барназу Bacillus amyloliquefaciens,
гены, кодирующие рибонуклеазы: Т1 рибонуклеазу Aspergillus oryzae и барназу Bacillus amyloliquefaciens,
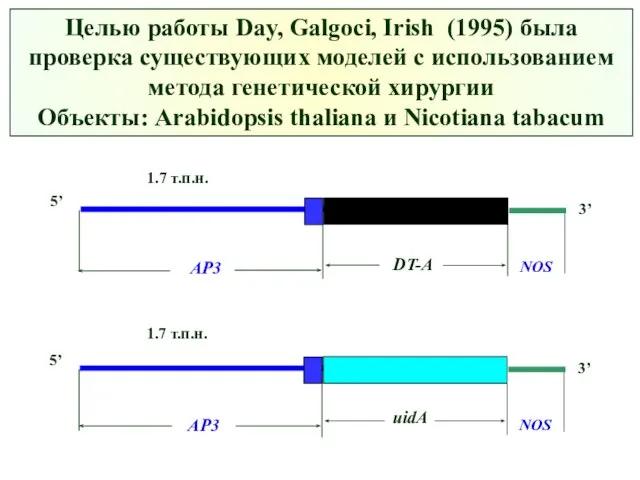
Целью работы Day, Galgoci, Irish (1995) была проверка существующих моделей с
Целью работы Day, Galgoci, Irish (1995) была проверка существующих моделей с
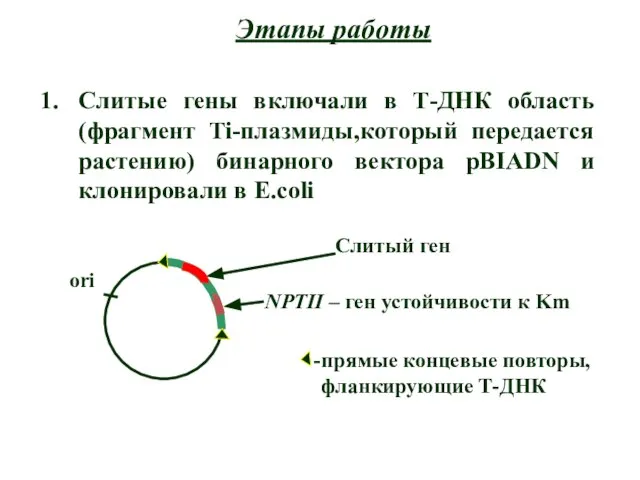
Этапы работы
Слитые гены включали в Т-ДНК область (фрагмент Ti-плазмиды,который передается растению)
Этапы работы
Слитые гены включали в Т-ДНК область (фрагмент Ti-плазмиды,который передается растению)

У резушки получено 5 трансгенных растений, в геноме которых присутствовали от
У резушки получено 5 трансгенных растений, в геноме которых присутствовали от
Использование трансформации для изучения роли гормонов в развитиии растений
Использование трансформации для изучения роли гормонов в развитиии растений

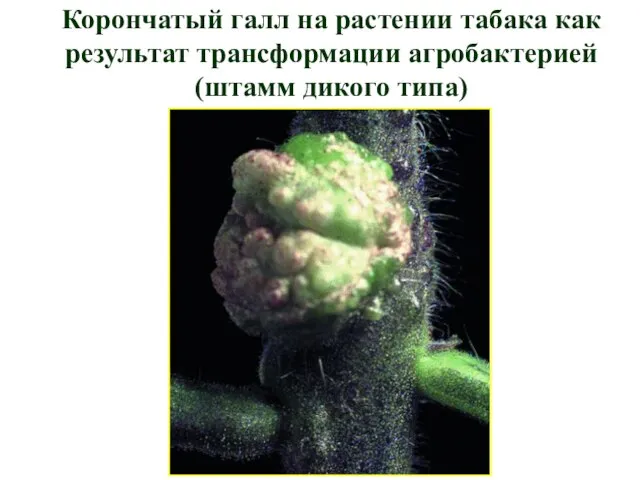
Корончатый галл на растении табака как результат трансформации агробактерией (штамм дикого
Корончатый галл на растении табака как результат трансформации агробактерией (штамм дикого

Трансгенные рстения с геном tmr
Трансгенные рстения с геном tmr

Ген tmr в трансгенных растениях, экспрессирующийся в плодах (промотор 2А11)
Ген tmr в трансгенных растениях, экспрессирующийся в плодах (промотор 2А11)

Опухолеобразование у редиса – генетический признак
Опухолеобразование у редиса – генетический признак

Фенокопии опухолеобразования у трансгенных растений с геном ipt
Фенокопии опухолеобразования у трансгенных растений с геном ipt

Прямая генетика:
фенотип ген
Обратная генетика:
ген фенотип
Прямая и обратная генетика в
Прямая генетика:
фенотип ген
Обратная генетика:
ген фенотип
Прямая и обратная генетика в

- гомологичная рекомбинация/замещение гена
-замещение гена: альтернативные подходы (Zn-fingers nucleases, TALEN)
- РНК-интерференция/
- гомологичная рекомбинация/замещение гена -замещение гена: альтернативные подходы (Zn-fingers nucleases, TALEN) - РНК-интерференция/
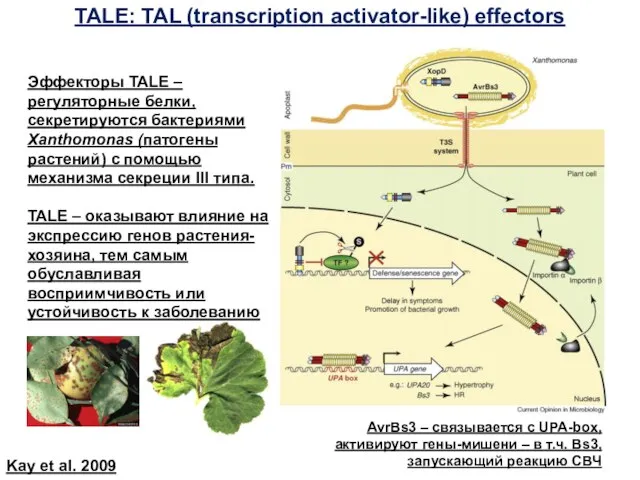
TALE: TAL (transcription activator-like) effectors
Эффекторы TALE – регуляторные белки, секретируются бактериями
TALE: TAL (transcription activator-like) effectors
Эффекторы TALE – регуляторные белки, секретируются бактериями
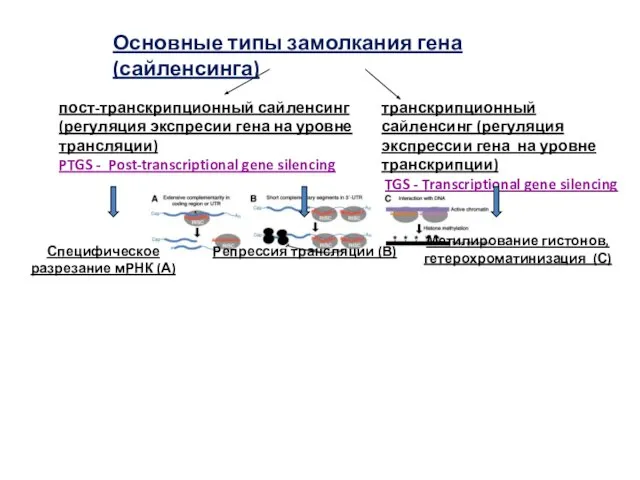
Основные типы замолкания гена (сайленсинга)
пост-транскрипционный сайленсинг
(регуляция экспресии гена на уровне трансляции)
PTGS
Основные типы замолкания гена (сайленсинга)
пост-транскрипционный сайленсинг
(регуляция экспресии гена на уровне трансляции)
PTGS
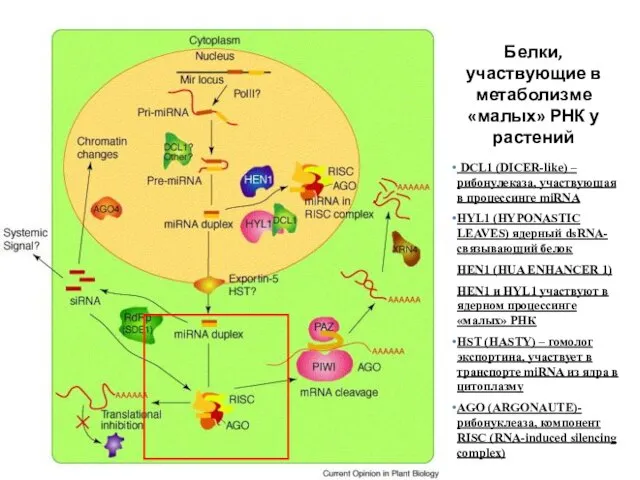
Белки, участвующие в метаболизме «малых» РНК у растений
DCL1 (DICER-like) –
Белки, участвующие в метаболизме «малых» РНК у растений
DCL1 (DICER-like) –
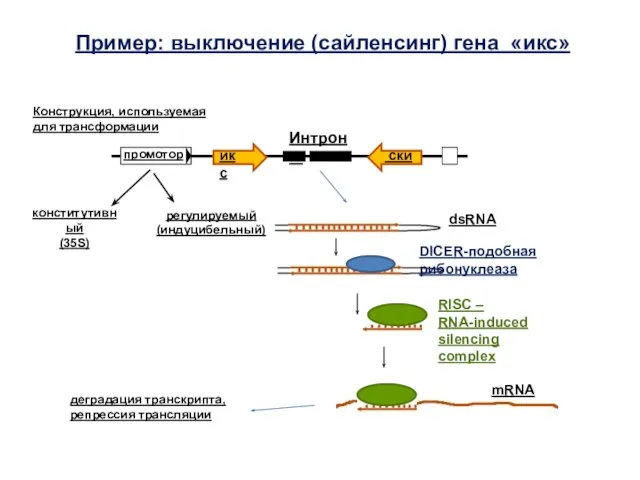
Пример: выключение (сайленсинг) гена «икс»
конститутивный
(35S)
регулируемый
(индуцибельный)
Интроны
промотор
икс
ски
dsRNA
RISC –
RNA-induced
silencing
Пример: выключение (сайленсинг) гена «икс»
конститутивный
(35S)
регулируемый
(индуцибельный)
Интроны
промотор
икс
ски
dsRNA
RISC –
RNA-induced
silencing
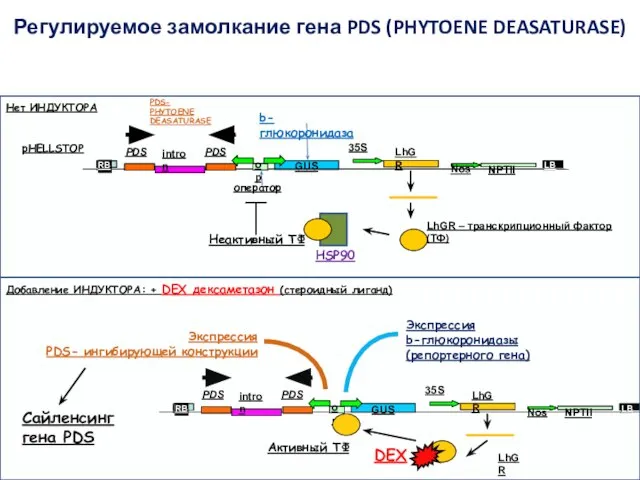
Регулируемое замолкание гена PDS (PHYTOENE DEASATURASE)
LB
RB
RB
LB
35S
LhGR
GUS
op
PDS
PDS
intron
pHELLSTOP
PDS-
PHYTOENE
DEASATURASE
оператор
b-глюкоронидаза
Нет ИНДУКТОРА
Добавление ИНДУКТОРА: + DEX дексаметазон
Регулируемое замолкание гена PDS (PHYTOENE DEASATURASE)
LB
RB
RB
LB
35S
LhGR
GUS
op
PDS
PDS
intron
pHELLSTOP
PDS-
PHYTOENE
DEASATURASE
оператор
b-глюкоронидаза
Нет ИНДУКТОРА
Добавление ИНДУКТОРА: + DEX дексаметазон

Регулируемое замолкание (индуцибельный сайленсинг)
гена PDS (PHYTOENE DEASATURASE)
No Dex
Dex
Регулируемое замолкание (индуцибельный сайленсинг)
гена PDS (PHYTOENE DEASATURASE)
No Dex
Dex
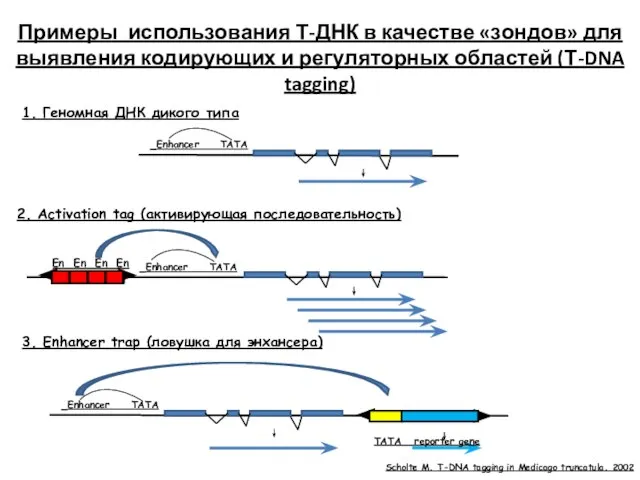
Enhancer TATA
En En En En
Enhancer TATA
Enhancer TATA
3. Enhancer
Enhancer TATA
En En En En
Enhancer TATA
Enhancer TATA
3. Enhancer
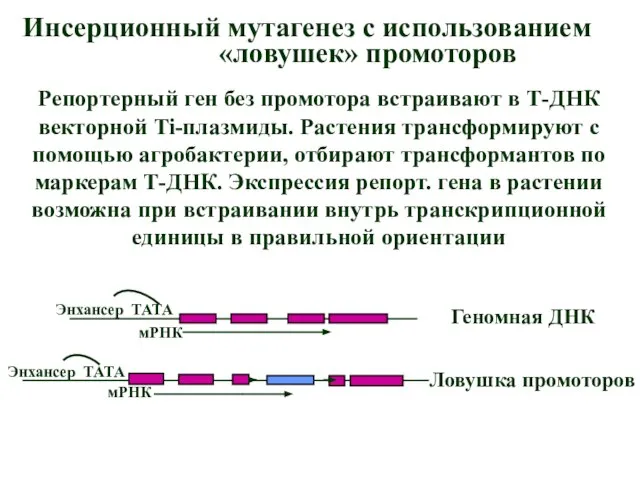
Инсерционный мутагенез с использованием «ловушек» промоторов
Репортерный ген без промотора встраивают
Инсерционный мутагенез с использованием «ловушек» промоторов
Репортерный ген без промотора встраивают
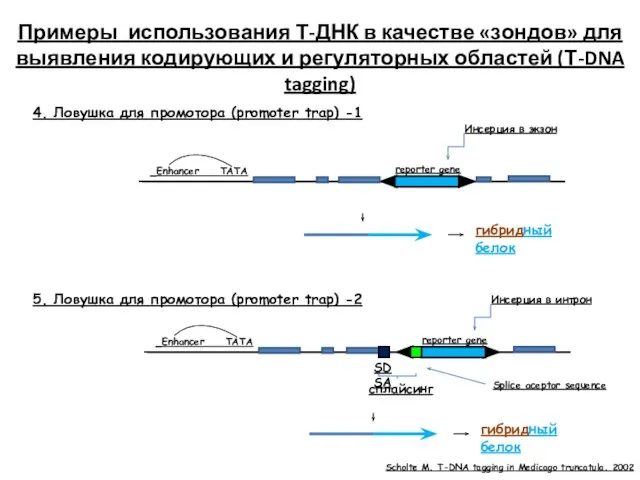
Enhancer TATA
4. Ловушка для промотора (promoter trap) -1
reporter gene
гибридный белок
Enhancer TATA
4. Ловушка для промотора (promoter trap) -1
reporter gene
гибридный белок
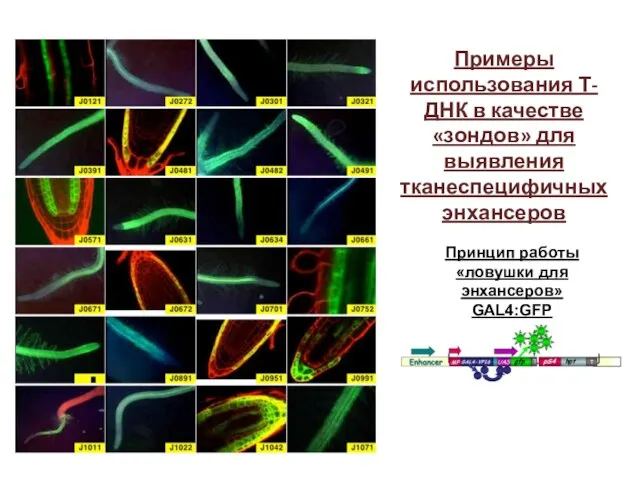
Принцип работы
«ловушки для энхансеров»
GAL4:GFP
Примеры использования Т-ДНК в качестве «зондов»
Принцип работы
«ловушки для энхансеров»
GAL4:GFP
Примеры использования Т-ДНК в качестве «зондов»
Методы редактирования генома растений
Методы редактирования генома растений

Модификации генома: разновидности
Изменение случайного места
в геноме
Редактирование генома
Модификации генома: разновидности
Изменение случайного места
в геноме
Редактирование генома
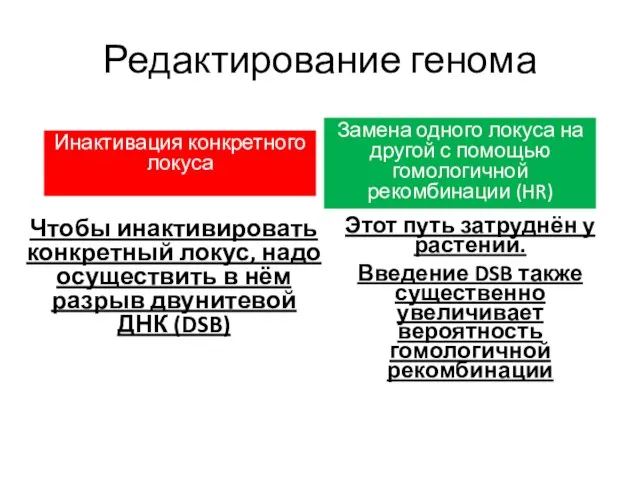
Редактирование генома
Инактивация конкретного локуса
Замена одного локуса на другой с помощью гомологичной
Редактирование генома
Инактивация конкретного локуса
Замена одного локуса на другой с помощью гомологичной

Программируемые нуклеазы
ZFN (Zink-Finger Nucleases)
TALENs (Transcription Activator-Like Effector Nucleases)
CRISPR-Cas система (Clustered regularly
Программируемые нуклеазы
ZFN (Zink-Finger Nucleases)
TALENs (Transcription Activator-Like Effector Nucleases)
CRISPR-Cas система (Clustered regularly

Нуклеазы с цинковыми пальцами
В качестве ДНК-связывающего домена используется универсальный домен типа
Нуклеазы с цинковыми пальцами
В качестве ДНК-связывающего домена используется универсальный домен типа


Как программируют ZFN?
Wolfe et al., 1999
Каждый цинковый палец распознаёт 3 (4)
Как программируют ZFN?
Wolfe et al., 1999
Каждый цинковый палец распознаёт 3 (4)

TALEN (Transcription Activator-Like Effectors Nucleases)
TALEN (Transcription Activator-Like Effectors Nucleases)
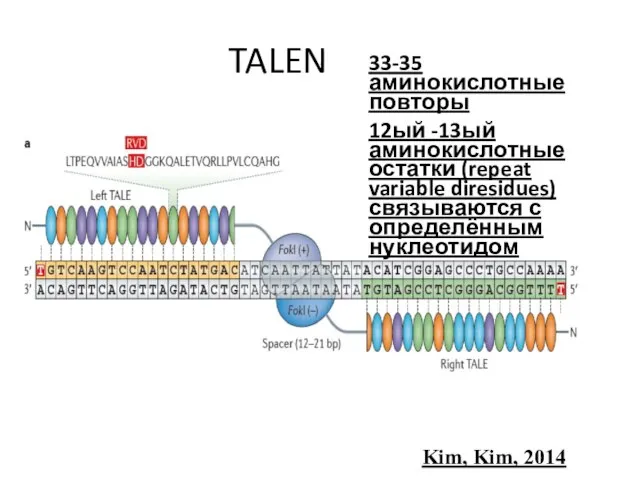
TALEN
Kim, Kim, 2014
33-35 аминокислотные повторы
12ый -13ый аминокислотные остатки (repeat variable
TALEN
Kim, Kim, 2014
33-35 аминокислотные повторы
12ый -13ый аминокислотные остатки (repeat variable

Применение TALEN
Потеря функции гена LOX3 (кодирует липоксигеназу, создающую перекись водорода) увеличивает
Применение TALEN
Потеря функции гена LOX3 (кодирует липоксигеназу, создающую перекись водорода) увеличивает

Система CRISPR/Cas9 (Clustered regularly Interspaced Palindromic Repeats- CRISPR associated protein)
Система CRISPR/Cas9 (Clustered regularly Interspaced Palindromic Repeats- CRISPR associated protein)
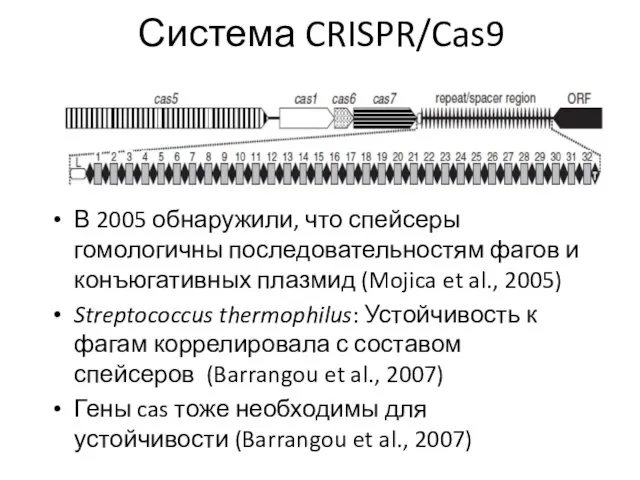
Система CRISPR/Cas9
В 2005 обнаружили, что спейсеры гомологичны последовательностям фагов и конъюгативных
Система CRISPR/Cas9
В 2005 обнаружили, что спейсеры гомологичны последовательностям фагов и конъюгативных
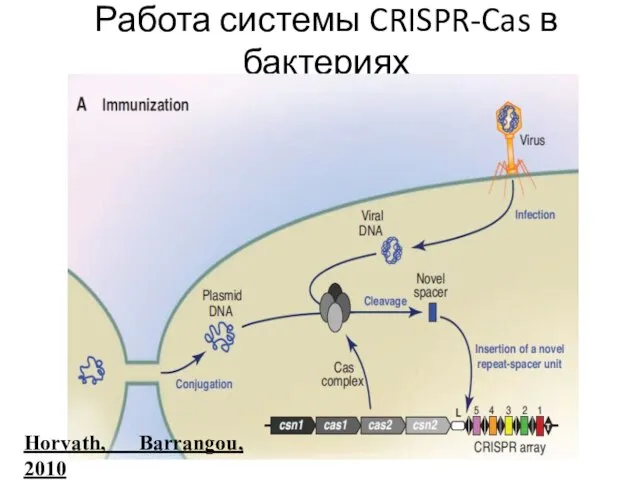
Работа системы CRISPR-Cas в бактериях
Horvath, Barrangou, 2010
Работа системы CRISPR-Cas в бактериях
Horvath, Barrangou, 2010

Čermák et al., 2015
Čermák et al., 2015
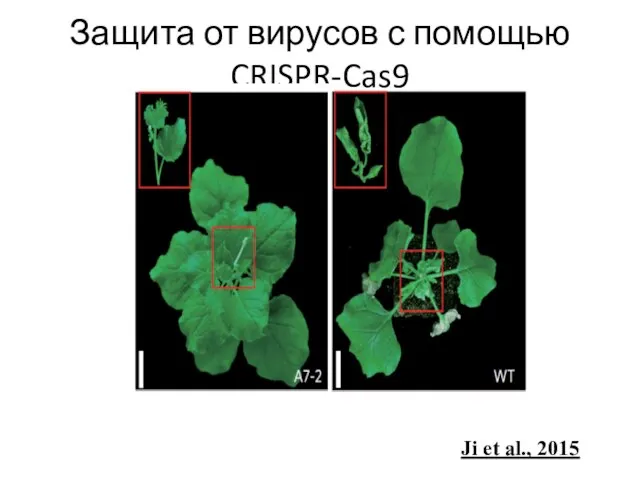
Защита от вирусов с помощью CRISPR-Cas9
Ji et al., 2015
Защита от вирусов с помощью CRISPR-Cas9
Ji et al., 2015
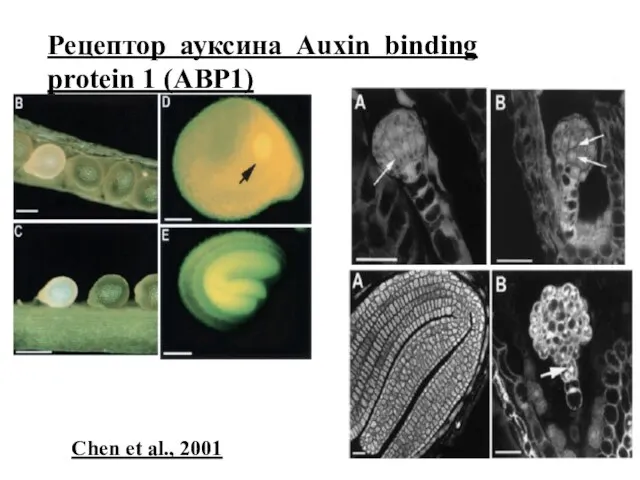
Рецептор ауксина Auxin binding protein 1 (ABP1)
Chen et al., 2001
Рецептор ауксина Auxin binding protein 1 (ABP1)
Chen et al., 2001
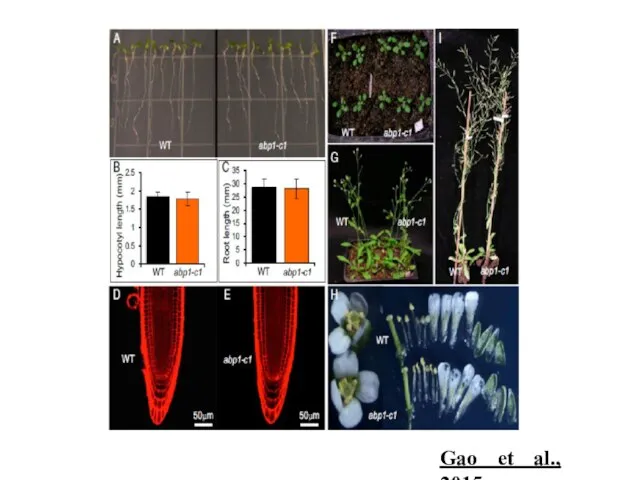
Gao et al., 2015
Gao et al., 2015
 LAMARK (1)
LAMARK (1) Основные виды питательных веществ и их значение в питании человека
Основные виды питательных веществ и их значение в питании человека Презентация на тему Отряд Пингвины
Презентация на тему Отряд Пингвины  Семейство Крестоцветные
Семейство Крестоцветные Презентация на тему Класс Паукообразные
Презентация на тему Класс Паукообразные  Старение и инсулиновый сигналинг у человека
Старение и инсулиновый сигналинг у человека Биофизика мышечного сокращения
Биофизика мышечного сокращения Устойчивость микроорганизмов к воздействию тепла и штатные процессы тепловой консервации
Устойчивость микроорганизмов к воздействию тепла и штатные процессы тепловой консервации Изучение эффекта синергизма между вирусным и бактериальным агентами при борьбе с непарным шелкопрядом
Изучение эффекта синергизма между вирусным и бактериальным агентами при борьбе с непарным шелкопрядом Взгляды и гипотезы о происхождении жизни на Земле Урок биологии в 9классе Учитель биологии МОУСОШ №2
Взгляды и гипотезы о происхождении жизни на Земле Урок биологии в 9классе Учитель биологии МОУСОШ №2  Свойства и функции белков
Свойства и функции белков Минеральное (корневое) питание растений
Минеральное (корневое) питание растений Выполнил: Мустафин Тимур МОУ СОШ №156 2а класс Екатеринбург, 2011 г
Выполнил: Мустафин Тимур МОУ СОШ №156 2а класс Екатеринбург, 2011 г  Сны и сновидения. Выполнили: Ученицы 8 «В» класса Лицея №1 им. Г.С.Титова Барткив София и Сысоева Елизавета
Сны и сновидения. Выполнили: Ученицы 8 «В» класса Лицея №1 им. Г.С.Титова Барткив София и Сысоева Елизавета Многообразие стеблей и видоизменения побегов.
Многообразие стеблей и видоизменения побегов. Витамины - укрепляют организм
Витамины - укрепляют организм Юные исследователи
Юные исследователи Презентация на тему Синдром Шерешевского-Тернера
Презентация на тему Синдром Шерешевского-Тернера  Физиологические основы голода и насщыения. Регуляция аппетита
Физиологические основы голода и насщыения. Регуляция аппетита Биосферный центральный заповедник
Биосферный центральный заповедник Леса Азии
Леса Азии Переломы Работа учащихся Даньковской основной школы
Переломы Работа учащихся Даньковской основной школы Пищеварительная система
Пищеварительная система Этапы урока. Организация начала занятия. Проверка выполнения домашнего задания. Подготовка к усвоению новых знаний Изучение
Этапы урока. Организация начала занятия. Проверка выполнения домашнего задания. Подготовка к усвоению новых знаний Изучение  Земноводные. Амфибии
Земноводные. Амфибии Что такое онтогенез?
Что такое онтогенез? Презентация Загрязнение атмосферы
Презентация Загрязнение атмосферы Список препаратов набор 1
Список препаратов набор 1