- Старение и инсулиновый сигналинг у человека

Содержание
- 2. Старение – биологический процесс, сопровождающийся закономерно возникающими в организме возрастными изменениями, характер которых генетически запрограммирован. Возникновение
- 3. К основным сигнальным механизмам, задействованным в регуляции процессов старения и долголетия, относятся: инсулин/инсулиноподобный фактор роста-1 (ИФР-1),
- 4. Умеренные физические упражнения, ограничение калорийности питания и снижение веса – то есть, воздействия, снижающие уровень циркулирующего
- 5. Наиболее частые аллели гена ИФР-1 (19 и 20 СА-повторов) ассоциированы с максимальным уровнем ИФР-1 в сыворотке
- 6. СХЕМА ИНСУЛИНОВОГО СИГНАЛИНГА
- 7. Циркулирующая глюкоза индуцирует высвобождение инсулина. Секреция ИФР-1 запускается многими факторами, в первую очередь соматотропным гормоном (СТГ).
- 8. Инсулин на внешней поверхности клетки стимулирует тирозинкиназную активность инсулиновых рецепторов (InR или IGF1R), что ведет к
- 9. Одним из важнейших сигнальных каскадов, регулируемых инсулином и ИФР-1 является фосфатидилинозитол-3-киназный путь (метаболический сигнал)
- 10. Модель инсулинового сигналинга
- 11. Инсулин вызывает фосфорилирование белков – инсулин-рецепторных субстанций 1-го и 2-го типов (IRS1 и IRS2). Это приводит
- 12. Активируется фосфатидилинозитол-3-киназа (PI3K), катализирующая образование вторичных посредников фосфатидилинозитола-3,4,5-трифосфата (PI-3,4,5-P(3)) и фосфатидилинозитола-4,5-бифосфата (PI-4,5-P(2)), которые активируют фосфоинозитолзависимую протеинкиназу
- 13. Белок PTEN (Phosphatase and tensin homolog) является антагонистом фосфатидилинозитол-3-киназной активности, стимулирует ядерную локализацию эндогенного FOXO (Forkhead
- 14. Активированная АКТ-киназа: – приводит к транслокации инсулинзависимого глюкозного транспортера 4 типа (GLUT-4) под воздействием протеинкиназы С
- 15. Активированная АКТ-киназа: – контролирует активность киназы-3-гликогенситетазы (GSK3), которая регулирует синтез свободных жирных кислот и гликогена, ингибирует
- 16. Активированная АКТ-киназа: – активирует протеинкиназу mTOR (mammalian Target Of Rapamycin), что приводит к фосфорилированию р70-рибосомальной S6-киназы
- 17. Активированная АКТ-киназа: – тормозит PEPCK (Phosphoenolpyruvate carboxykinase) и IGFBP-1 (Insulin-like growth factor-binding protein 1) и выключаются
- 18. Показано, что мишенью АКТ-киназы также является белок FBP-1 (forkhead-box protein-1), негативный регулятор инсулинового сигналинга, активация которого
- 19. Второй путь инсулинового сигналинга – митоген-активированный протеинкиназный путь (RAS/MAP-киназный) – ростовой сигнал также необходим для нормального
- 20. Инсулин на внешней поверхности мембраны стимулирует тирозинкиназную активность инсулиновых рецепторов. Фосфорилированные изоформы p46Shc и p52Shc локализованные
- 21. Изоформа p66Shc локализована в митохондриальном матриксе и не участвует в активации Ras, а также подавляет Erk1/2
- 22. Активируется белок Ras, который активирует белок гена Raf. В свою очередь активация Raf приводит к фосфорилированию
- 23. В результате активации инсулин/ИФР-1 сигнального пути обеспечивается нормальный рост и развитие организма, поддержание метаболического гомеостаза
- 24. С возрастом происходит увеличение активности PI3K, что приводит к снижению стрессоустойчивости организма. К тому же, с
- 25. Установлено, что фармакологическое ингибирование PI3K приводит к увеличению продолжительности жизни у нематод и коловраток. У дрозофилы,
- 26. Третьим путем является р38/МАРК-опосредованный сигнальный путь При фосфорилировании IRS1/2 активируются белок р38 и JNK (c-Jun N-terminal
- 27. Активация p38 также инициируется внешнесредовыми сигналами. p38 – эволюционно-консервативная изоформа митоген-активируемой протеинкиназы (MAPK). У млекопитающих активируется
- 28. р38 участвует в реакциях синтеза провоспалительных цитокинов, развитии ревматоидного артрита, болезни Альцгеймера, развитии рака. Также р38
- 29. Факторы роста стимулируют деление и выживание клетки путем активации рецептора инсулина, который действует через два главных
- 30. НАРУШЕНИЯ ИНСУЛИНОВОГО СИГНАЛИНГА Метаболический сигналинг инсулина (фосфатидилинозитол-3-киназный путь) резко снижен у тучных лиц и практически отсутствует
- 31. НАРУШЕНИЯ ИНСУЛИНОВОГО СИГНАЛИНГА Гиперинсулинемия приводит к гиперстимуляции митоген-активированного протеин-(МАР)-киназного пути с сохраненной чувствительностью к инсулину, что
- 32. НАРУШЕНИЯ ИНСУЛИНОВОГО СИГНАЛИНГА Гиперинсулинемия удваивает способность ангиотензина II трансактивировать нуклеарный фактор каппа В (NF-kВ), который, в
- 33. Нарушение метаболического сигналинга инсулина приводит к снижению продукции оксида азота, что, в свою очередь, делает эндотелиальные
- 34. ВЛИЯНИЕ ИНСУЛИН/ИФР-1-СИГНАЛИНГА НА ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
- 35. У людей-долгожителей повышена чувствительность к инсулину при сохранении его низкого уровня в плазме крови. Активность инсулинового
- 36. Мутации в генах киназ PI3K, AKT/PKB, PDK сопровождаются увеличением продолжительности жизни животных. Активность фосфатаз PTEN, SHIP1
- 37. Инсулиновый сигналинг подавляет активность механизмов стресс-ответа, связанных с транскрипционным фактором FOXO. Активность FOXO и FOXO-зависимых генов
- 38. Инсулин/ИФР-1-путь регулирует продолжительность жизни не сам по себе, а посредством большого количества генов, в том числе
- 39. В норме у человека чувствительность к инсулину с возрастом снижается. Снижение количества инсулина в плазме крови
- 40. СИСТЕМА СОМАТОТРОПИН/ ИНСУЛИНОПОДОБНЫЙ ФАКТОР РОСТА-1/ИНСУЛИН
- 41. C возрастом ночной пик секреции СТГ снижается как у человека, так и у лабораторных грызунов, сопровождаясь
- 42. У человека, и у лабораторных животных с возрастом развиваются нарушения в системе передачи сигнала рецептора СТГ,
- 43. У итальянских столетних людей концентрация ИФР-1 очень низкая, что и способствует их долголетию
- 44. Введение СТГ и ИФР-1 старым животным оказывало ряд благоприятных эффектов, в частности стимулировало внутриклеточный синтез белка,
- 45. Снижая уровень СТГ и ИФР-1 до нижней границы нормы (5-10 перцентиль) в пожилом возрасте, можно увеличить
- 46. Люди, теряющие функцию рецептора СТГ (синдром Ларона), контролирующего инсулиновый сигналинг, характеризуются: маленьким ростом; уменьшением продолжительности жизни
- 47. «Инсулиновый парадокс» (по теории Благосклонного): инсулин и ИФР-1 активируют TOR. Поэтому уменьшение ИФР-1/инсулин сигналинга снижает активность
- 49. Скачать презентацию

Старение – биологический процесс, сопровождающийся закономерно возникающими в организме возрастными изменениями,
Старение – биологический процесс, сопровождающийся закономерно возникающими в организме возрастными изменениями,

К основным сигнальным механизмам, задействованным в регуляции процессов
старения и долголетия,
К основным сигнальным механизмам, задействованным в регуляции процессов старения и долголетия,

Умеренные физические упражнения, ограничение калорийности питания и снижение веса – то
Умеренные физические упражнения, ограничение калорийности питания и снижение веса – то

Наиболее частые аллели гена ИФР-1 (19 и 20 СА-повторов) ассоциированы с
Наиболее частые аллели гена ИФР-1 (19 и 20 СА-повторов) ассоциированы с

СХЕМА
ИНСУЛИНОВОГО СИГНАЛИНГА
СХЕМА
ИНСУЛИНОВОГО СИГНАЛИНГА

Циркулирующая глюкоза индуцирует высвобождение инсулина. Секреция ИФР-1 запускается многими факторами, в
Циркулирующая глюкоза индуцирует высвобождение инсулина. Секреция ИФР-1 запускается многими факторами, в

Инсулин на внешней поверхности клетки стимулирует тирозинкиназную активность инсулиновых рецепторов (InR
Инсулин на внешней поверхности клетки стимулирует тирозинкиназную активность инсулиновых рецепторов (InR

Одним из важнейших сигнальных каскадов, регулируемых инсулином и ИФР-1 является
фосфатидилинозитол-3-киназный
Одним из важнейших сигнальных каскадов, регулируемых инсулином и ИФР-1 является фосфатидилинозитол-3-киназный
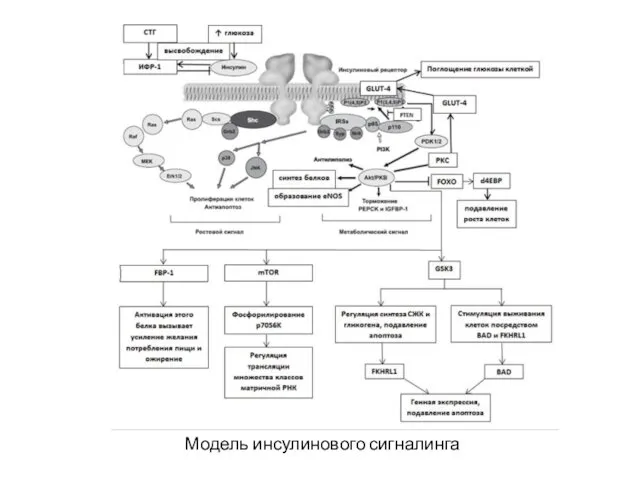
Модель инсулинового сигналинга
Модель инсулинового сигналинга

Инсулин вызывает фосфорилирование белков – инсулин-рецепторных субстанций 1-го и 2-го типов
Инсулин вызывает фосфорилирование белков – инсулин-рецепторных субстанций 1-го и 2-го типов

Активируется фосфатидилинозитол-3-киназа (PI3K), катализирующая образование вторичных посредников фосфатидилинозитола-3,4,5-трифосфата (PI-3,4,5-P(3)) и фосфатидилинозитола-4,5-бифосфата
Активируется фосфатидилинозитол-3-киназа (PI3K), катализирующая образование вторичных посредников фосфатидилинозитола-3,4,5-трифосфата (PI-3,4,5-P(3)) и фосфатидилинозитола-4,5-бифосфата

Белок PTEN (Phosphatase and tensin homolog) является антагонистом фосфатидилинозитол-3-киназной активности, стимулирует
Белок PTEN (Phosphatase and tensin homolog) является антагонистом фосфатидилинозитол-3-киназной активности, стимулирует

Активированная АКТ-киназа:
– приводит к транслокации инсулинзависимого глюкозного транспортера 4 типа (GLUT-4)
Активированная АКТ-киназа: – приводит к транслокации инсулинзависимого глюкозного транспортера 4 типа (GLUT-4)

Активированная АКТ-киназа:
– контролирует активность
киназы-3-гликогенситетазы (GSK3), которая регулирует синтез свободных жирных
Активированная АКТ-киназа: – контролирует активность киназы-3-гликогенситетазы (GSK3), которая регулирует синтез свободных жирных

Активированная АКТ-киназа:
– активирует протеинкиназу mTOR
(mammalian Target Of Rapamycin), что приводит
Активированная АКТ-киназа: – активирует протеинкиназу mTOR (mammalian Target Of Rapamycin), что приводит

Активированная АКТ-киназа:
– тормозит PEPCK (Phosphoenolpyruvate carboxykinase) и IGFBP-1 (Insulin-like growth factor-binding
Активированная АКТ-киназа: – тормозит PEPCK (Phosphoenolpyruvate carboxykinase) и IGFBP-1 (Insulin-like growth factor-binding

Показано, что мишенью АКТ-киназы также является белок FBP-1 (forkhead-box protein-1),
Показано, что мишенью АКТ-киназы также является белок FBP-1 (forkhead-box protein-1),

Второй путь инсулинового сигналинга –
митоген-активированный протеинкиназный путь (RAS/MAP-киназный) –
ростовой
Второй путь инсулинового сигналинга – митоген-активированный протеинкиназный путь (RAS/MAP-киназный) – ростовой

Инсулин на внешней поверхности мембраны стимулирует тирозинкиназную активность инсулиновых рецепторов.
Фосфорилированные
Инсулин на внешней поверхности мембраны стимулирует тирозинкиназную активность инсулиновых рецепторов. Фосфорилированные

Изоформа p66Shc локализована в митохондриальном матриксе и не участвует в активации
Изоформа p66Shc локализована в митохондриальном матриксе и не участвует в активации

Активируется белок Ras, который активирует белок гена Raf.
В свою очередь
Активируется белок Ras, который активирует белок гена Raf. В свою очередь

В результате активации инсулин/ИФР-1 сигнального пути
обеспечивается
нормальный рост и развитие
В результате активации инсулин/ИФР-1 сигнального пути обеспечивается нормальный рост и развитие

С возрастом происходит увеличение активности PI3K, что приводит к снижению стрессоустойчивости
С возрастом происходит увеличение активности PI3K, что приводит к снижению стрессоустойчивости

Установлено, что фармакологическое ингибирование PI3K приводит к
увеличению продолжительности жизни
у
Установлено, что фармакологическое ингибирование PI3K приводит к увеличению продолжительности жизни у

Третьим путем является
р38/МАРК-опосредованный сигнальный путь
При фосфорилировании IRS1/2 активируются белок
Третьим путем является р38/МАРК-опосредованный сигнальный путь При фосфорилировании IRS1/2 активируются белок

Активация p38 также инициируется внешнесредовыми сигналами.
p38 – эволюционно-консервативная изоформа
митоген-активируемой
Активация p38 также инициируется внешнесредовыми сигналами. p38 – эволюционно-консервативная изоформа митоген-активируемой

р38 участвует в реакциях синтеза провоспалительных цитокинов, развитии ревматоидного артрита, болезни
р38 участвует в реакциях синтеза провоспалительных цитокинов, развитии ревматоидного артрита, болезни

Факторы роста
стимулируют деление и выживание клетки
путем активации рецептора инсулина,
Факторы роста стимулируют деление и выживание клетки путем активации рецептора инсулина,

НАРУШЕНИЯ ИНСУЛИНОВОГО СИГНАЛИНГА
Метаболический сигналинг инсулина (фосфатидилинозитол-3-киназный путь)
резко снижен у тучных
НАРУШЕНИЯ ИНСУЛИНОВОГО СИГНАЛИНГА Метаболический сигналинг инсулина (фосфатидилинозитол-3-киназный путь) резко снижен у тучных

НАРУШЕНИЯ ИНСУЛИНОВОГО СИГНАЛИНГА
Гиперинсулинемия приводит к
гиперстимуляции
митоген-активированного протеин-(МАР)-киназного пути
с сохраненной
НАРУШЕНИЯ ИНСУЛИНОВОГО СИГНАЛИНГА Гиперинсулинемия приводит к гиперстимуляции митоген-активированного протеин-(МАР)-киназного пути с сохраненной

НАРУШЕНИЯ ИНСУЛИНОВОГО СИГНАЛИНГА
Гиперинсулинемия
удваивает способность
ангиотензина II трансактивировать
нуклеарный фактор каппа
НАРУШЕНИЯ ИНСУЛИНОВОГО СИГНАЛИНГА Гиперинсулинемия удваивает способность ангиотензина II трансактивировать нуклеарный фактор каппа

Нарушение метаболического сигналинга инсулина приводит к
снижению продукции оксида азота, что,
Нарушение метаболического сигналинга инсулина приводит к снижению продукции оксида азота, что,

ВЛИЯНИЕ
ИНСУЛИН/ИФР-1-СИГНАЛИНГА
НА ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
ВЛИЯНИЕ
ИНСУЛИН/ИФР-1-СИГНАЛИНГА
НА ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ

У людей-долгожителей
повышена чувствительность к инсулину
при сохранении его низкого уровня
У людей-долгожителей повышена чувствительность к инсулину при сохранении его низкого уровня

Мутации в генах киназ
PI3K, AKT/PKB, PDK
сопровождаются
увеличением продолжительности жизни
Мутации в генах киназ PI3K, AKT/PKB, PDK сопровождаются увеличением продолжительности жизни

Инсулиновый сигналинг
подавляет активность механизмов стресс-ответа, связанных с транскрипционным фактором FOXO.
Инсулиновый сигналинг подавляет активность механизмов стресс-ответа, связанных с транскрипционным фактором FOXO.

Инсулин/ИФР-1-путь
регулирует
продолжительность жизни не сам по себе,
а посредством большого
Инсулин/ИФР-1-путь регулирует продолжительность жизни не сам по себе, а посредством большого

В норме у человека
чувствительность к инсулину
с возрастом снижается.
Снижение
В норме у человека чувствительность к инсулину с возрастом снижается. Снижение

СИСТЕМА
СОМАТОТРОПИН/
ИНСУЛИНОПОДОБНЫЙ ФАКТОР РОСТА-1/ИНСУЛИН
СИСТЕМА
СОМАТОТРОПИН/
ИНСУЛИНОПОДОБНЫЙ ФАКТОР РОСТА-1/ИНСУЛИН

C возрастом
ночной пик секреции СТГ
снижается
как у человека, так
C возрастом ночной пик секреции СТГ снижается как у человека, так

У человека, и у лабораторных животных
с возрастом
развиваются нарушения
в
У человека, и у лабораторных животных с возрастом развиваются нарушения в

У итальянских столетних людей
концентрация ИФР-1
очень низкая,
что и способствует
У итальянских столетних людей концентрация ИФР-1 очень низкая, что и способствует

Введение СТГ и ИФР-1
старым животным
оказывало ряд благоприятных эффектов, в
Введение СТГ и ИФР-1 старым животным оказывало ряд благоприятных эффектов, в

Снижая уровень СТГ и ИФР-1
до нижней границы нормы (5-10 перцентиль)
Снижая уровень СТГ и ИФР-1 до нижней границы нормы (5-10 перцентиль)

Люди, теряющие функцию рецептора СТГ
(синдром Ларона),
контролирующего инсулиновый сигналинг,
характеризуются:
Люди, теряющие функцию рецептора СТГ (синдром Ларона), контролирующего инсулиновый сигналинг, характеризуются:

«Инсулиновый парадокс»
(по теории Благосклонного):
инсулин и ИФР-1 активируют TOR.
Поэтому уменьшение
«Инсулиновый парадокс» (по теории Благосклонного): инсулин и ИФР-1 активируют TOR. Поэтому уменьшение
 Общее знакомство с цветковыми растениями
Общее знакомство с цветковыми растениями Листья деревьев. Лабораторные работы
Листья деревьев. Лабораторные работы Центральная нервная система головного мозга
Центральная нервная система головного мозга Элодея Канадская
Элодея Канадская Методы исследования в биологии. Лабораторное оборудование
Методы исследования в биологии. Лабораторное оборудование Узловые моменты прогрессивной эволюции хордовых. Общая характеристика и систематика типа Хордовые
Узловые моменты прогрессивной эволюции хордовых. Общая характеристика и систематика типа Хордовые Решение задач по молекулярной биологии
Решение задач по молекулярной биологии Виды мышц
Виды мышц  Типы галлов. Галлы крушиновой галловой листоблошки. (Лекция 11)
Типы галлов. Галлы крушиновой галловой листоблошки. (Лекция 11) Презентация на тему "Историческое прошлое людей. Расы человека" - скачать презентации по Биологии
Презентация на тему "Историческое прошлое людей. Расы человека" - скачать презентации по Биологии Многообразие пресмыкающихся
Многообразие пресмыкающихся Насекомые - переносчики заболевания человека
Насекомые - переносчики заболевания человека Экосистема озера
Экосистема озера  ДОМАШНИЕ ЖИВОТНЫЕ. Презентация Шумковой Кристины и Алексеевой Светланы. Города Перми.
ДОМАШНИЕ ЖИВОТНЫЕ. Презентация Шумковой Кристины и Алексеевой Светланы. Города Перми. Секреты истинного здоровья
Секреты истинного здоровья Предмет и задачи микробиологии
Предмет и задачи микробиологии Проект на тему Собаки
Проект на тему Собаки Черепно-мозговые нервы № 2
Черепно-мозговые нервы № 2 Синий кит
Синий кит Микробы и окружающая среда (тема 5)
Микробы и окружающая среда (тема 5) Единство химической организации живых организмов. Углеводы
Единство химической организации живых организмов. Углеводы Жизнь леса
Жизнь леса РЕШЕНИЕ БИОЛОГИЧЕСКИХ ЗАДАЧ Электив по разделу «Организм»
РЕШЕНИЕ БИОЛОГИЧЕСКИХ ЗАДАЧ Электив по разделу «Организм» Хищная птица орел
Хищная птица орел Третий закон Менделя
Третий закон Менделя Эволюция человека
Эволюция человека Органы выделения насекомых
Органы выделения насекомых «Природные зоны. Лес. Растения леса.» 4 класс Залесская Ирина Валерьевна учитель начальных классов СОШ № 5 г. Караганды, Казахс
«Природные зоны. Лес. Растения леса.» 4 класс Залесская Ирина Валерьевна учитель начальных классов СОШ № 5 г. Караганды, Казахс