- Электрон - транспортная цепь митохондрий

Содержание
- 2. Дыхание растений
- 3. Общая схема дыхательной ЭТЦ: четыре белковых комплекса, объединяемые подвижными переносчиками е- Переносчики е- в ЭТЦ выстаиваются
- 4. Хиноны (убихиноны и пластохиноны) – липофильные молекулы с Red-Ox свойствами: перенос 2ē + 2Н+ E0’: от
- 5. Флавинадениндинуклеотид (ФАД, FAD) и флавинмононуклеотид (ФМН, FMN) – компоненты многих Red-Ox ферментов: перенос 2ē + 2Н+
- 6. Железо-серные белки - 2Fe-2S и 4Fe-4S: перенос только одного ē Eо’: от – 0,42V (Fd) до
- 7. Гемы – коферменты цитохромов: перенос только одного ē Eо’: от -0,18 V (cyt b6) до +0,55
- 8. Подвижные переносчики: Убихиноны – их много («пул убихинонов») 2. Цитохром с – 104 ак. Локализован с
- 9. Комплекс I: NADH-дегидрогеназа – похож на старый башмак Матрикс Межмембранное пространство Мембрана
- 10. Комплекс I Матрикс Межмембранное пространство Мембрана ФМН НАДН- связывающий сайт FeS-кластеры FeS-кластер Убихинон- связывающий сайт Карбоангидраза
- 11. Комплекс I: NADH-дегидрогеназа
- 12. Особенность ЭТЦ дыхания растений – дополнительные NAD(P)H-дегидрогеназы
- 13. КомплексII: сукцинатдегидрогеназа Единственный фермент цикла Кребса, встроенный в мембрану митохондрий
- 14. Структура: димер, мономер - до 11 белков, min – 3 белка: Cyt b (2 гема bL
- 15. Комплекс III: почти как в фотосинтезе Межмембранное пространство Межмембранное пространство матрикс матрикс
- 16. Цитохром с: белок, ассоциированный с мембраной D-L DH D-Лактат Пируват GL DH L-Галактоно- лактон L-Аскорбат cyt
- 17. Комплекс IV: цитохром a/a3, цитохромоксидаза Структура: димер 125 kDa. Мономеры: 3 больших полипептида, кодируются в m:
- 18. Комплекс IV: цитохром a/a3, цитохромоксидаза CuA гем а гем а3 -СuB H+ H+ H+ D K
- 19. Предполагаемая схема работы цитохрома а-а3 O2 О● ē 2H+ O H H 2H2O ē 2H+ ē
- 20. АТФ-синтаза: вальсирующий комплекс Структура: Две субъединицы: F0 и F1 F1: α3β3γδε α - 59, β -
- 21. Альтернативная оксидаза
- 22. 2QH2 +O2 → 2Q + 2H2O Альтернативная оксидаза Термогенез ароидных растений Arum maculatum, Symplocarpus foetidus, Sauromatum
- 23. Sauromatum guttatum
- 24. Symplocarpus foetidus
- 25. Helicodeceros muscivorus
- 26. Особенность ЭТЦ дыхания растений – альтернативная оксидаза Гомодимер, 37 kDa, ~ 350 а-к. Передает ē от
- 27. Альтернативная оксидаза
- 28. Зачем нужна альтернативная оксидаза и как она включается? Гипотезы: 1. Для шунтирования ЭТЦ – модели «перелива»
- 29. Регуляция экспрессии и активности альтернативной оксидазы
- 30. Вторичный метаболизм: цианогенные гликозиды Лирическое отступление
- 31. Откуда берется цианид? Цианогенные гликозиды C≡N ⎟ глюкоза – О – C – СН3 ⎟ R
- 32. Цианогенные гликозиды
- 33. Цианогенные гликозиды продуцируют 2500-3000 видов растений сем. бобовых, розоцветных, мимозовых Клевер, кассава, лен, сорго, лотос, гевея,
- 34. Цианогенные гликозиды Trifolium repens Клевер ползучий Lupinus polyphyllus Люпин многолистный Syringa pannonica Сирень венгерская Padus racemosa
- 35. Маниок съедобный ( Manihot esculenta ) Сем. Молочайные ( Euphorbiaceae )
- 36. Маниок съедобный ( Manihot esculenta ) Сем. Молочайные ( Euphorbiaceae ) Родина: Бразилия (культивируется 6–7 тыс.
- 37. Маниок съедобный ( Manihot esculenta ) Сем. Молочайные ( Euphorbiaceae )
- 38. Маниок съедобный ( Manihot esculenta ) Сем. Молочайные ( Euphorbiaceae ) HCN цианид, синильная кислота Линамарин:
- 39. C≡N ⎟ глюкоза – О – C – СН3 ⎟ R О ⎟⎟ C – СН3
- 40. Маниок съедобный ( Manihot esculenta ) Сем. Молочайные ( Euphorbiaceae )
- 41. Маниок съедобный ( Manihot esculenta ) Сем. Молочайные ( Euphorbiaceae )
- 42. Дыхание растений: итоги
- 43. Итак, ЭТЦ дыхания растений: У растений обнаружили UCP (разобщающий белок), как и у животных
- 44. NADPH Порины Порины Цитозоль (симпласт) Пластида Митохондрия ЭТЦ Н2О Цит b/f ФC II ФC I 3-ФГК
- 45. NAD+- зависимая МДГ декарб. Пируват- карбоксилаза СО2 Пируват (С3) NADPH Порины Порины Цитозоль (симпласт) Пластида Митохондрия
- 46. ЭТЦ дыхания растений: последние новости… DHO DH Дигидро- оротат Оротат Пиримидины G-3-P DH 3-Ф-глицерин ДГАФ Гликолиз
- 47. ЭТЦ дыхания растений: образование суперкомплексов, респирасома Состав суперкомплексов: I+III2 I2+III4 III2+IV III2+IV2 I+III2+IV I+III2+IV2 I+III2+IV4 I2+III2+IV2
- 48. ЭТЦ дыхания растений: образование суперкомплексов КС-I КС-III КС-IV I2+III2+IV2
- 49. ЭТЦ дыхания растений: образование суперкомплексов АТФ-синтазы Места сборки супрекомплексов АТФ-синтазы соответствуют местам инициации крист
- 50. ЭТЦ дыхания растений: образование суперкомплексов АТФ-синтазы Кристы работают как протонные «ловушки»
- 51. Генетическая функция митохондрий
- 52. Множество кольцевых молекул митохондриальной ДНК растений – результат гомологичных рекомбинаций по повторам.
- 53. Предполагаемая структура «мастер-хромосомы» митохондрий кукурузы
- 54. Линейная ДНК митохондрий хламидомонады
- 55. Сопоставление хлоропластного и митохондриального геномов риса
- 56. Варианты редактирования хлоропластных и митохондриальных РНК растений
- 57. Гены митохондрий Синтез белка. - 3 гена рРНК (оперон rrn) - 10 генов белков пластидных рибосом
- 58. Ядерный генов контролирует : некоторые т РНК ДНК – полимеразу РНК – полимеразу Некоторые рибосомальные белки
- 60. Fzo – это гигантские ГТФ-азы, расположенные во внешней мембране митохондрий Fzo (fuzzy onion)
- 61. Цитоплазматическая мужская стерильность (ЦМС) «Техасская» - Т-ЦМС «USDA»-тип - S-ЦМС «Молдавская» - M-ЦМС «Парагвайская» (Charrua-тип) -
- 62. АТФ-синтаза: ключевой фермент «Техасской» ЦМС 13 kDa-белок появляется в митохондриях как результат незаконной рекомбинации c геном
- 63. АТФ-синтаза: ключевой фермент «Техасской» ЦМС 13 kDa-белок появляется в митохондриях как результат незаконной рекомбинации c геном
- 64. АТФ-синтаза: ключевой фермент ЦМС у петунии У петунии незаконная рекомбинация происходит с участием гена atp9. Изменения
- 65. Цитоплазматическая мужская стерильность (ЦМС) «USDA»-тип - S-ЦМС «Молдавская» - Т-ЦМС Ядерный ген Rf3 При S-ЦМС(=М-ЦМС) появляются
- 66. Цитоплазматическая мужская стерильность (ЦМС): спорофитный и гаметофитный контроль Стерильность Полное (!) восстановление фертильности «USDA»-тип - S-ЦМС
- 67. Цитоплазматическая мужская стерильность (ЦМС): спорофитный и гаметофитный контроль «Техасская» - Т-ЦМС Ядерные гены Rf1 и Rf2
- 70. Локализация ферментов глиоксилатного цикла
- 71. РАСТИТЕЛЬНЫЕ ПЕРОКСИСОМЫ – ИСТОЧНИК СИГНАЛЬНЫХ МОЛЕКУЛ
- 72. Метаболический переход от гетеротрофного к фототрофному питанию
- 74. Скачать презентацию

Дыхание растений
Дыхание растений

Общая схема дыхательной ЭТЦ: четыре белковых комплекса, объединяемые подвижными переносчиками е-
Переносчики
Общая схема дыхательной ЭТЦ: четыре белковых комплекса, объединяемые подвижными переносчиками е-
Переносчики

Хиноны (убихиноны и пластохиноны) – липофильные молекулы с Red-Ox свойствами: перенос
Хиноны (убихиноны и пластохиноны) – липофильные молекулы с Red-Ox свойствами: перенос
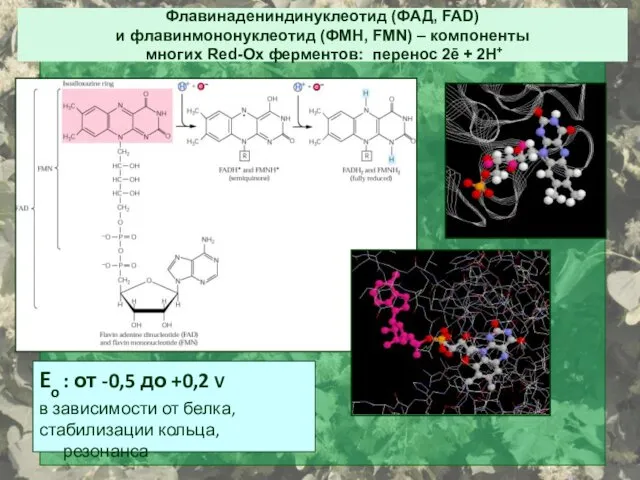
Флавинадениндинуклеотид (ФАД, FAD)
и флавинмононуклеотид (ФМН, FMN) – компоненты
многих Red-Ox
Флавинадениндинуклеотид (ФАД, FAD) и флавинмононуклеотид (ФМН, FMN) – компоненты многих Red-Ox
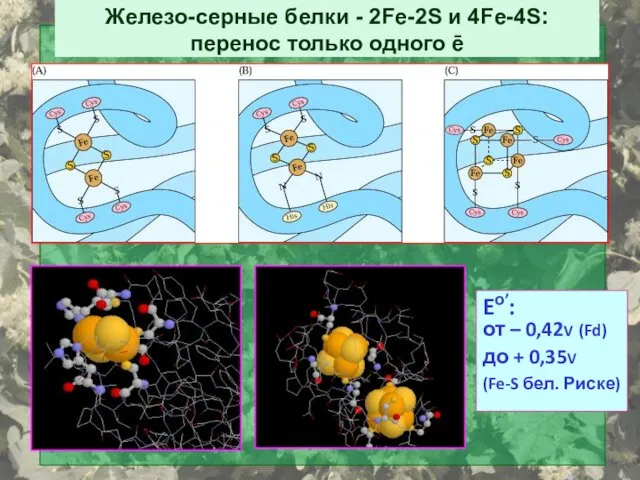
Железо-серные белки - 2Fe-2S и 4Fe-4S: перенос только одного ē
Eо’:
от
Железо-серные белки - 2Fe-2S и 4Fe-4S: перенос только одного ē
Eо’:
от
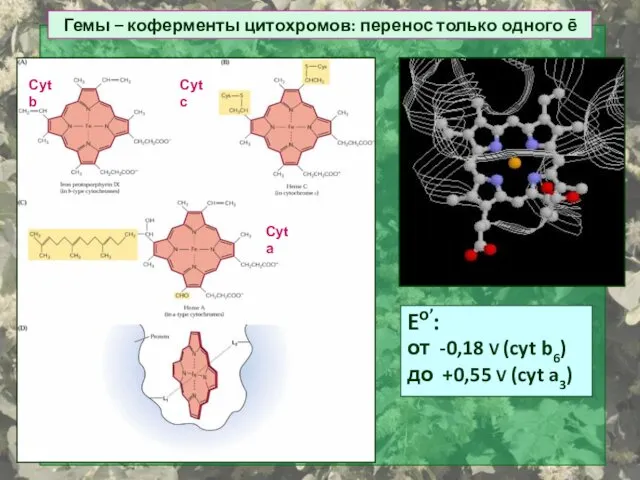
Гемы – коферменты цитохромов: перенос только одного ē
Eо’:
от -0,18
Гемы – коферменты цитохромов: перенос только одного ē
Eо’:
от -0,18
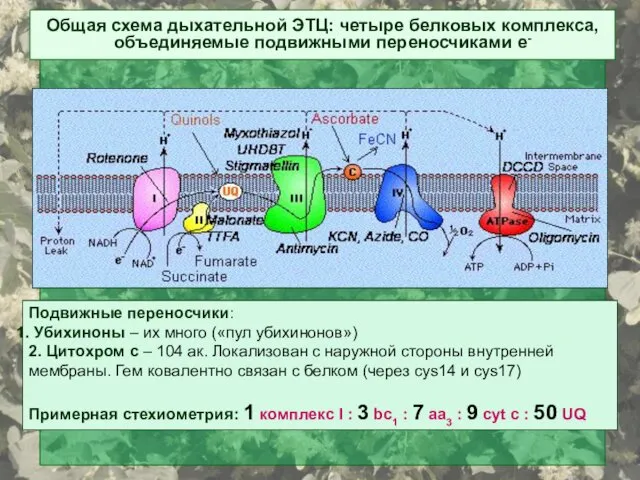
Подвижные переносчики:
Убихиноны – их много («пул убихинонов»)
2. Цитохром с –
Подвижные переносчики:
Убихиноны – их много («пул убихинонов»)
2. Цитохром с –
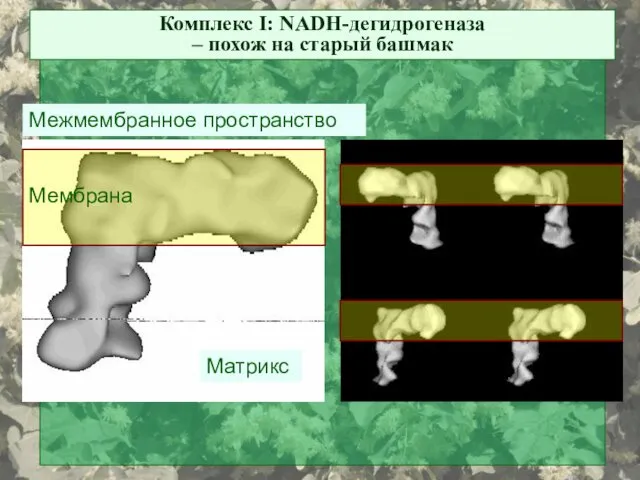
Комплекс I: NADH-дегидрогеназа
– похож на старый башмак
Матрикс
Межмембранное пространство
Мембрана
Комплекс I: NADH-дегидрогеназа
– похож на старый башмак
Матрикс
Межмембранное пространство
Мембрана
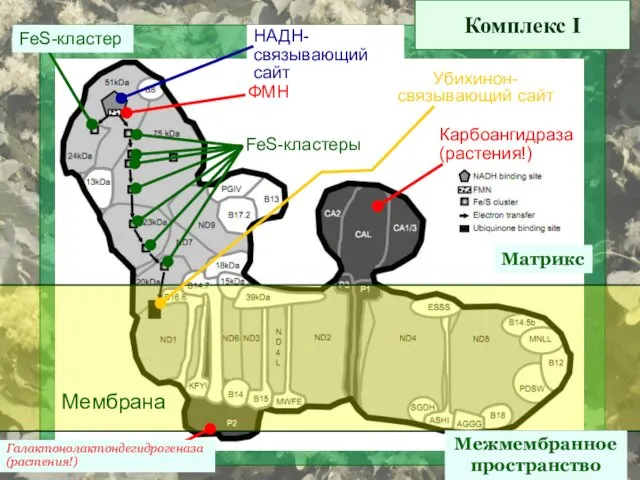
Комплекс I
Матрикс
Межмембранное
пространство
Мембрана
ФМН
НАДН-
связывающий сайт
FeS-кластеры
FeS-кластер
Убихинон-
связывающий сайт
Карбоангидраза
(растения!)
Галактонолактондегидрогеназа
(растения!)
Комплекс I
Матрикс
Межмембранное
пространство
Мембрана
ФМН
НАДН-
связывающий сайт
FeS-кластеры
FeS-кластер
Убихинон-
связывающий сайт
Карбоангидраза
(растения!)
Галактонолактондегидрогеназа
(растения!)
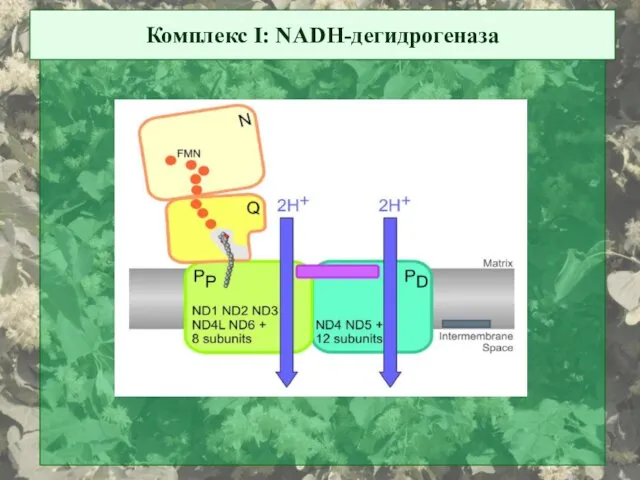
Комплекс I: NADH-дегидрогеназа
Комплекс I: NADH-дегидрогеназа
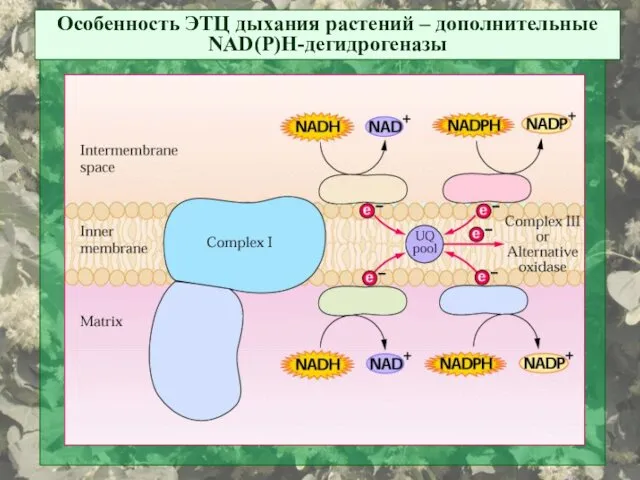
Особенность ЭТЦ дыхания растений – дополнительные NAD(P)H-дегидрогеназы
Особенность ЭТЦ дыхания растений – дополнительные NAD(P)H-дегидрогеназы
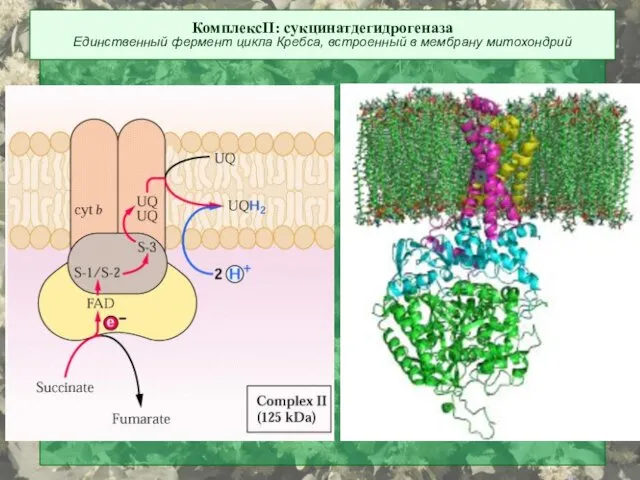
КомплексII: сукцинатдегидрогеназа
Единственный фермент цикла Кребса, встроенный в мембрану митохондрий
КомплексII: сукцинатдегидрогеназа
Единственный фермент цикла Кребса, встроенный в мембрану митохондрий
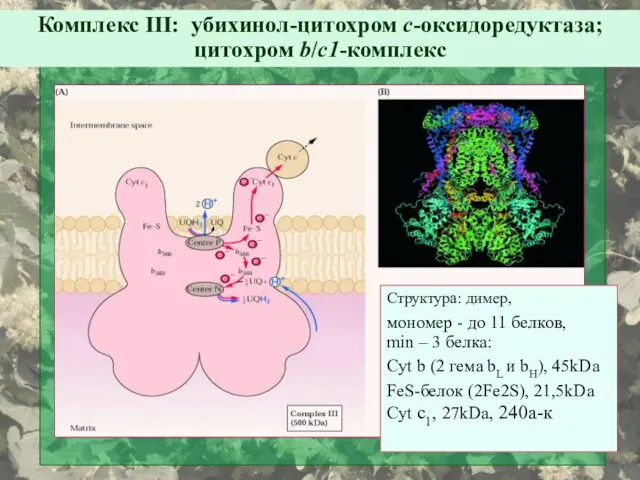
Структура: димер,
мономер - до 11 белков,
min – 3 белка:
Cyt b
Структура: димер,
мономер - до 11 белков,
min – 3 белка:
Cyt b
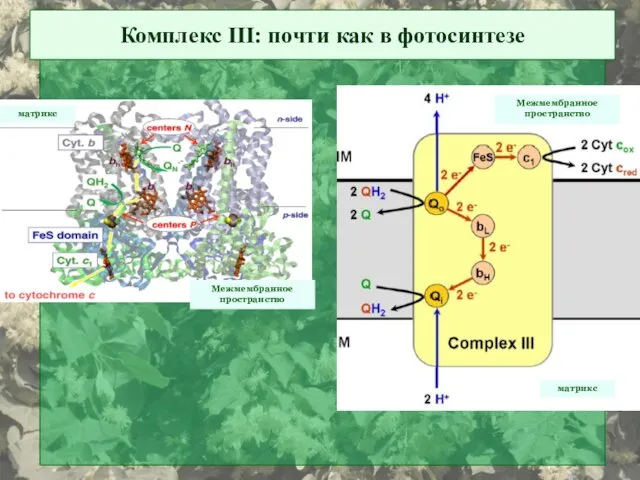
Комплекс III: почти как в фотосинтезе
Межмембранное
пространство
Межмембранное
пространство
матрикс
матрикс
Комплекс III: почти как в фотосинтезе
Межмембранное
пространство
Межмембранное
пространство
матрикс
матрикс
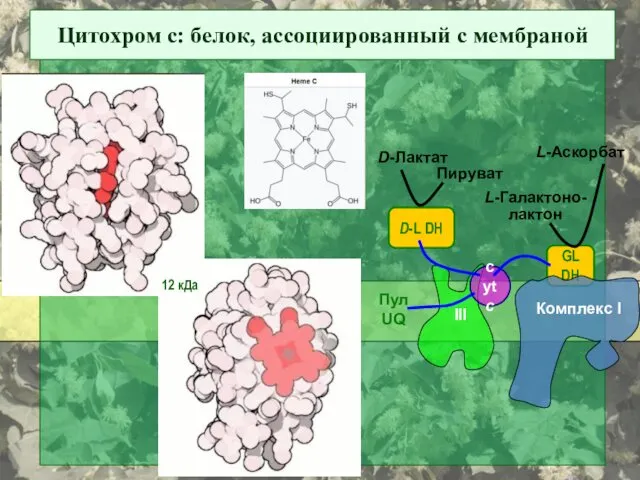
Цитохром с: белок, ассоциированный с мембраной
D-L DH
D-Лактат
Пируват
GL DH
L-Галактоно-
лактон
L-Аскорбат
cyt c
Пул
UQ
Цитохром с: белок, ассоциированный с мембраной
D-L DH
D-Лактат
Пируват
GL DH
L-Галактоно-
лактон
L-Аскорбат
cyt c
Пул
UQ
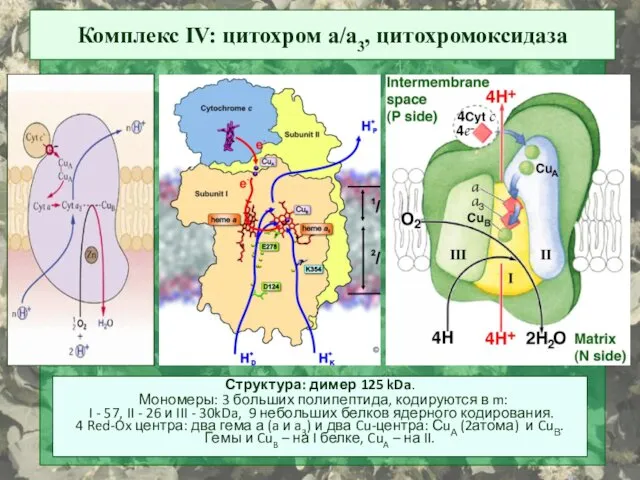
Комплекс IV: цитохром a/a3, цитохромоксидаза
Структура: димер 125 kDa.
Мономеры: 3 больших
Комплекс IV: цитохром a/a3, цитохромоксидаза
Структура: димер 125 kDa.
Мономеры: 3 больших

Комплекс IV: цитохром a/a3, цитохромоксидаза
CuA
гем а
гем а3 -СuB
H+ H+ H+
Комплекс IV: цитохром a/a3, цитохромоксидаза
CuA
гем а
гем а3 -СuB
H+ H+ H+
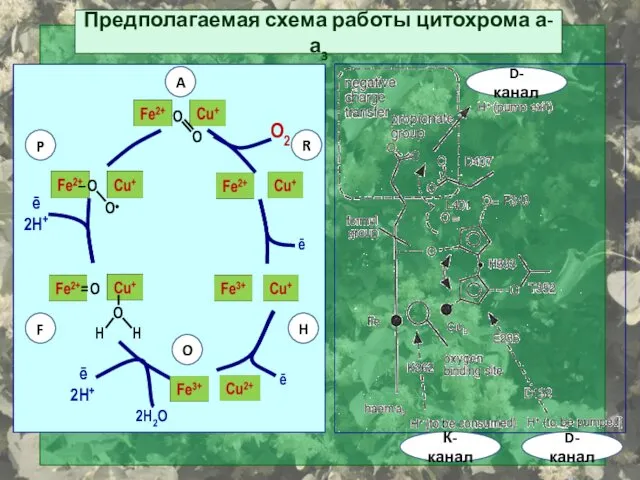
Предполагаемая схема работы цитохрома а-а3
O2
О●
ē
2H+
O
H
H
2H2O
ē
2H+
ē
ē
= O
– O
A
P
F
O
H
R
К-канал
D-канал
D-канал
Предполагаемая схема работы цитохрома а-а3
O2
О●
ē
2H+
O
H
H
2H2O
ē
2H+
ē
ē
= O
– O
A
P
F
O
H
R
К-канал
D-канал
D-канал
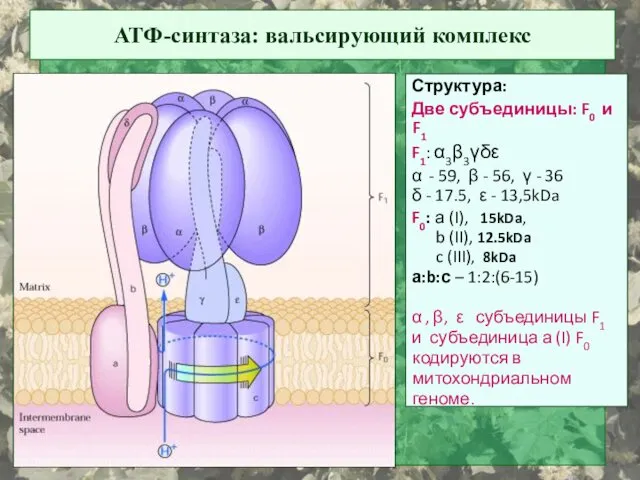
АТФ-синтаза: вальсирующий комплекс
Структура:
Две субъединицы: F0 и F1
F1: α3β3γδε
α - 59, β
АТФ-синтаза: вальсирующий комплекс
Структура:
Две субъединицы: F0 и F1
F1: α3β3γδε
α - 59, β

Альтернативная оксидаза
Альтернативная оксидаза

2QH2 +O2 → 2Q + 2H2O
Альтернативная оксидаза
Термогенез ароидных растений
Arum maculatum,
2QH2 +O2 → 2Q + 2H2O
Альтернативная оксидаза
Термогенез ароидных растений
Arum maculatum,

Sauromatum guttatum
Sauromatum guttatum

Symplocarpus foetidus
Symplocarpus foetidus
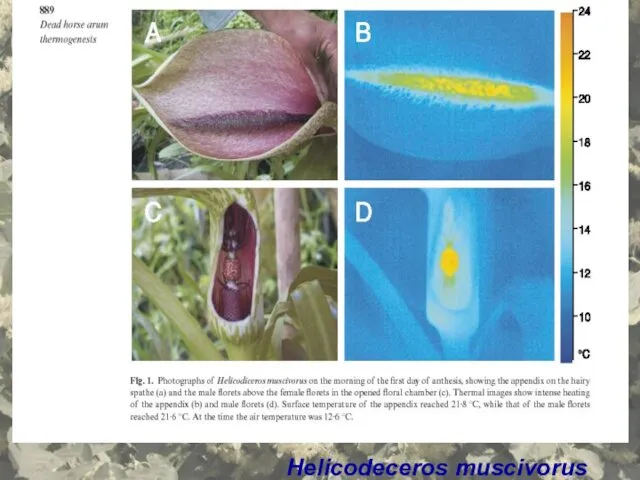
Helicodeceros muscivorus
Helicodeceros muscivorus
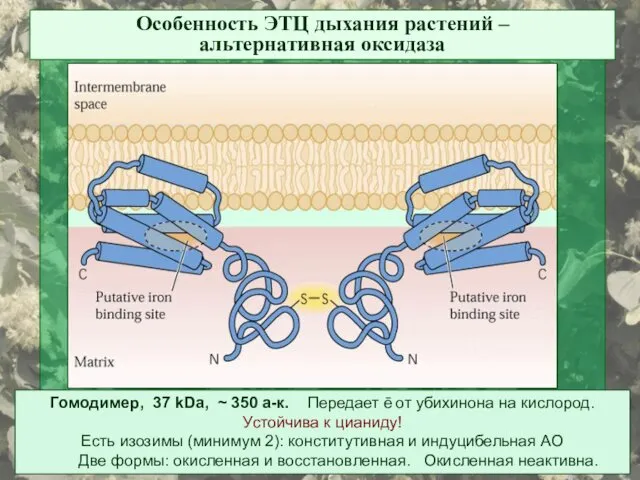
Особенность ЭТЦ дыхания растений –
альтернативная оксидаза
Гомодимер, 37 kDa, ~ 350
Особенность ЭТЦ дыхания растений –
альтернативная оксидаза
Гомодимер, 37 kDa, ~ 350
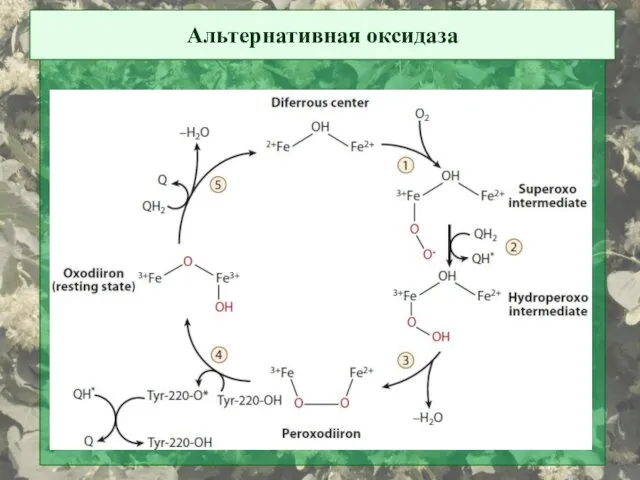
Альтернативная оксидаза
Альтернативная оксидаза

Зачем нужна альтернативная оксидаза
и как она включается?
Гипотезы:
1. Для шунтирования ЭТЦ
Зачем нужна альтернативная оксидаза
и как она включается?
Гипотезы:
1. Для шунтирования ЭТЦ

Регуляция экспрессии и активности альтернативной оксидазы
Регуляция экспрессии и активности альтернативной оксидазы

Вторичный метаболизм: цианогенные гликозиды
Лирическое отступление
Вторичный метаболизм: цианогенные гликозиды
Лирическое отступление

Откуда берется цианид?
Цианогенные гликозиды
C≡N
⎟
глюкоза – О – C –
Откуда берется цианид?
Цианогенные гликозиды
C≡N
⎟
глюкоза – О – C –

Цианогенные гликозиды
Цианогенные гликозиды

Цианогенные гликозиды продуцируют 2500-3000 видов растений
сем. бобовых, розоцветных, мимозовых
Клевер, кассава,
Цианогенные гликозиды продуцируют 2500-3000 видов растений
сем. бобовых, розоцветных, мимозовых
Клевер, кассава,

Цианогенные гликозиды
Trifolium repens
Клевер ползучий
Lupinus polyphyllus
Люпин многолистный
Syringa pannonica
Сирень венгерская
Padus racemosa
Черемуха обыкновенная
Padus serotina
Черемуха
Цианогенные гликозиды
Trifolium repens
Клевер ползучий
Lupinus polyphyllus
Люпин многолистный
Syringa pannonica
Сирень венгерская
Padus racemosa
Черемуха обыкновенная
Padus serotina
Черемуха
Маниок съедобный ( Manihot esculenta ) Сем. Молочайные ( Euphorbiaceae )
Маниок съедобный ( Manihot esculenta ) Сем. Молочайные ( Euphorbiaceae )

Маниок съедобный ( Manihot esculenta ) Сем. Молочайные ( Euphorbiaceae )
Маниок съедобный ( Manihot esculenta ) Сем. Молочайные ( Euphorbiaceae )

Маниок съедобный ( Manihot esculenta ) Сем. Молочайные ( Euphorbiaceae )
Маниок съедобный ( Manihot esculenta ) Сем. Молочайные ( Euphorbiaceae )
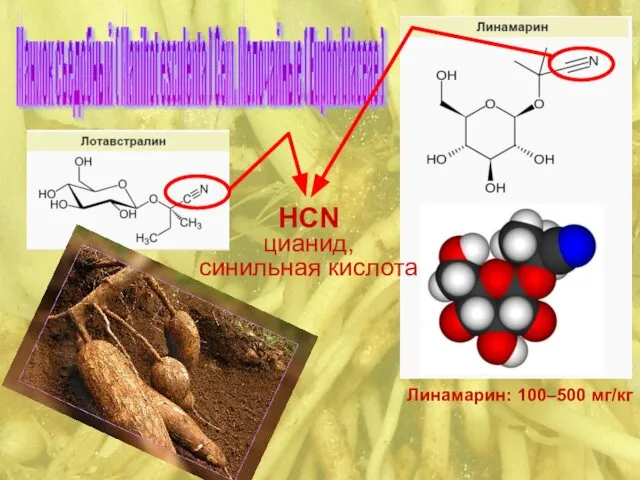
Маниок съедобный ( Manihot esculenta ) Сем. Молочайные ( Euphorbiaceae )
Маниок съедобный ( Manihot esculenta ) Сем. Молочайные ( Euphorbiaceae )
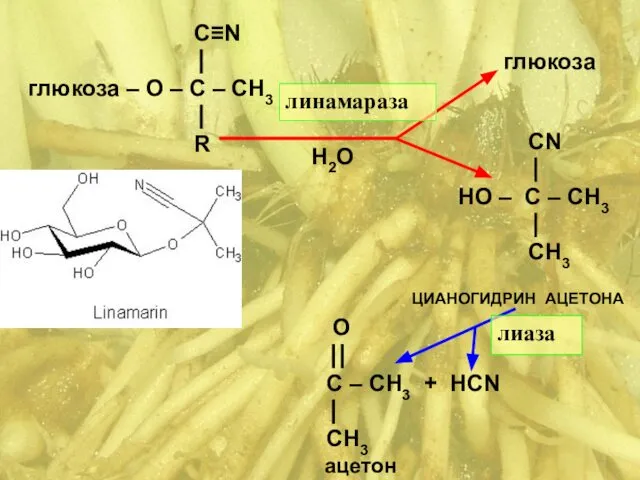
C≡N
⎟
глюкоза – О – C – СН3
⎟
C≡N
⎟
глюкоза – О – C – СН3
⎟

Маниок съедобный ( Manihot esculenta ) Сем. Молочайные ( Euphorbiaceae )
Маниок съедобный ( Manihot esculenta ) Сем. Молочайные ( Euphorbiaceae )

Маниок съедобный ( Manihot esculenta ) Сем. Молочайные ( Euphorbiaceae )
Маниок съедобный ( Manihot esculenta ) Сем. Молочайные ( Euphorbiaceae )

Дыхание растений: итоги
Дыхание растений: итоги
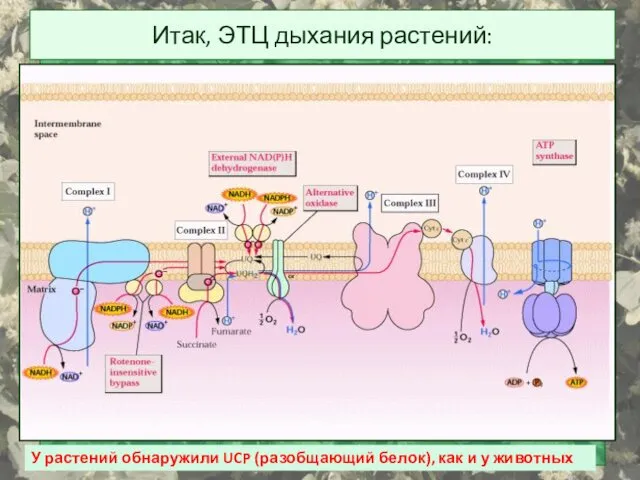
Итак, ЭТЦ дыхания растений:
У растений обнаружили UCP (разобщающий белок), как и
Итак, ЭТЦ дыхания растений:
У растений обнаружили UCP (разобщающий белок), как и

NADPH
Порины
Порины
Цитозоль
(симпласт)
Пластида
Митохондрия
ЭТЦ
Н2О
Цит b/f
ФC II
ФC I
3-ФГК (С3)
3-ФГА(С3)
NADP+
NADH
NAD+
О2
Н2О
ē
Гликолиз
Гликолиз,
Цикл Кальвина
1,3-ДФГК (С3)
АТФ
АДФ
3-ФГК (С3)
3-ФГА (С3)
1,3-ДФГК (С3)
АТФ
АДФ
Ф
Ф
Ф
Ф
NADPH
Порины
Порины
Цитозоль
(симпласт)
Пластида
Митохондрия
ЭТЦ
Н2О
Цит b/f
ФC II
ФC I
3-ФГК (С3)
3-ФГА(С3)
NADP+
NADH
NAD+
О2
Н2О
ē
Гликолиз
Гликолиз,
Цикл Кальвина
1,3-ДФГК (С3)
АТФ
АДФ
3-ФГК (С3)
3-ФГА (С3)
1,3-ДФГК (С3)
АТФ
АДФ
Ф
Ф
Ф
Ф

NAD+-
зависимая
МДГ декарб.
Пируват-
карбоксилаза
СО2
Пируват (С3)
NADPH
Порины
Порины
Цитозоль
(симпласт)
Пластида
Митохондрия
ЭТЦ
Н2О
Цит b/f
ФC II
ФC I
Оксалоацетат (С4)
Малат (С4)
NADP+-
зависимая
МДГ декарб.
Оксалоацетат (С4)
Малат
Оксалоацетат
NADP+
СО2
NADH
NAD+
Пируват (С3)
О2
Н2О
NADH
NAD+
Малат
NAD+-
зависимая
МДГ декарб.
Пируват-
карбоксилаза
СО2
Пируват (С3)
NADPH
Порины
Порины
Цитозоль
(симпласт)
Пластида
Митохондрия
ЭТЦ
Н2О
Цит b/f
ФC II
ФC I
Оксалоацетат (С4)
Малат (С4)
NADP+-
зависимая
МДГ декарб.
Оксалоацетат (С4)
Малат
Оксалоацетат
NADP+
СО2
NADH
NAD+
Пируват (С3)
О2
Н2О
NADH
NAD+
Малат
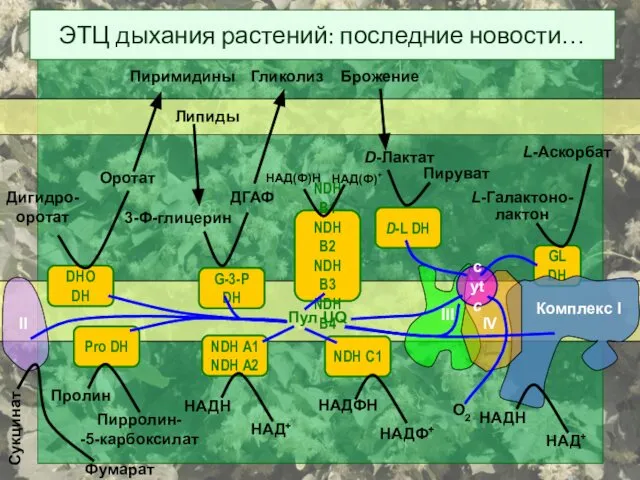
ЭТЦ дыхания растений: последние новости…
DHO DH
Дигидро-
оротат
Оротат
Пиримидины
G-3-P DH
3-Ф-глицерин
ДГАФ
Гликолиз
Pro DH
Пролин
Пирролин-
-5-карбоксилат
Пул UQ
D-L DH
D-Лактат
Пируват
GL DH
L-Галактоно-
лактон
L-Аскорбат
cyt
ЭТЦ дыхания растений: последние новости…
DHO DH
Дигидро-
оротат
Оротат
Пиримидины
G-3-P DH
3-Ф-глицерин
ДГАФ
Гликолиз
Pro DH
Пролин
Пирролин-
-5-карбоксилат
Пул UQ
D-L DH
D-Лактат
Пируват
GL DH
L-Галактоно-
лактон
L-Аскорбат
cyt
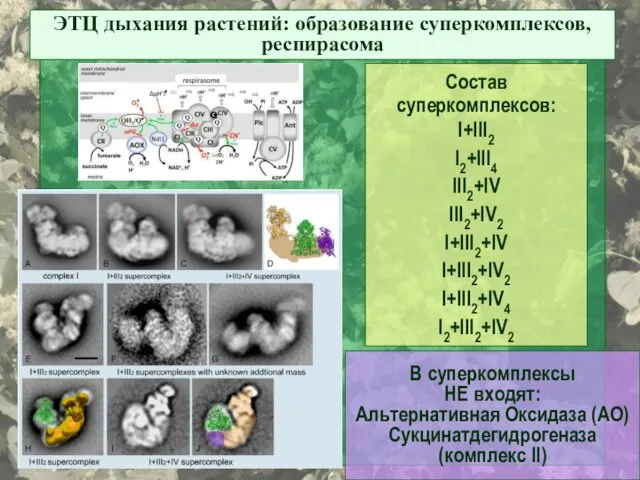
ЭТЦ дыхания растений: образование суперкомплексов, респирасома
Состав суперкомплексов:
I+III2
I2+III4
III2+IV
III2+IV2
I+III2+IV
I+III2+IV2
I+III2+IV4
I2+III2+IV2
В суперкомплексы
НЕ входят:
Альтернативная Оксидаза
ЭТЦ дыхания растений: образование суперкомплексов, респирасома
Состав суперкомплексов:
I+III2
I2+III4
III2+IV
III2+IV2
I+III2+IV
I+III2+IV2
I+III2+IV4
I2+III2+IV2
В суперкомплексы
НЕ входят:
Альтернативная Оксидаза
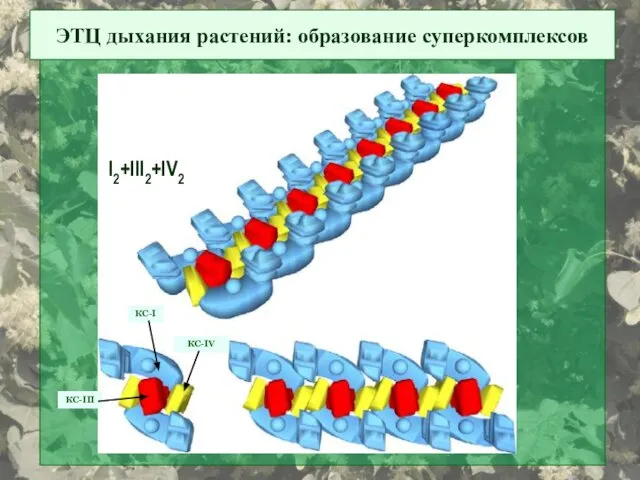
ЭТЦ дыхания растений: образование суперкомплексов
КС-I
КС-III
КС-IV
I2+III2+IV2
ЭТЦ дыхания растений: образование суперкомплексов
КС-I
КС-III
КС-IV
I2+III2+IV2
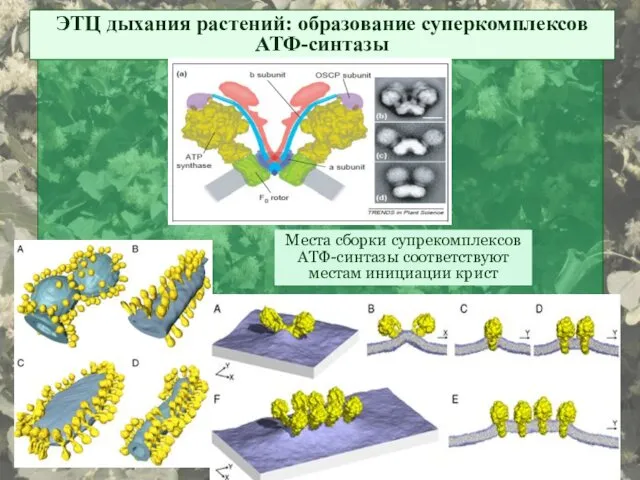
ЭТЦ дыхания растений: образование суперкомплексов АТФ-синтазы
Места сборки супрекомплексов АТФ-синтазы соответствуют местам
ЭТЦ дыхания растений: образование суперкомплексов АТФ-синтазы
Места сборки супрекомплексов АТФ-синтазы соответствуют местам

ЭТЦ дыхания растений: образование суперкомплексов АТФ-синтазы
Кристы работают как протонные «ловушки»
ЭТЦ дыхания растений: образование суперкомплексов АТФ-синтазы
Кристы работают как протонные «ловушки»

Генетическая функция митохондрий
Генетическая функция митохондрий
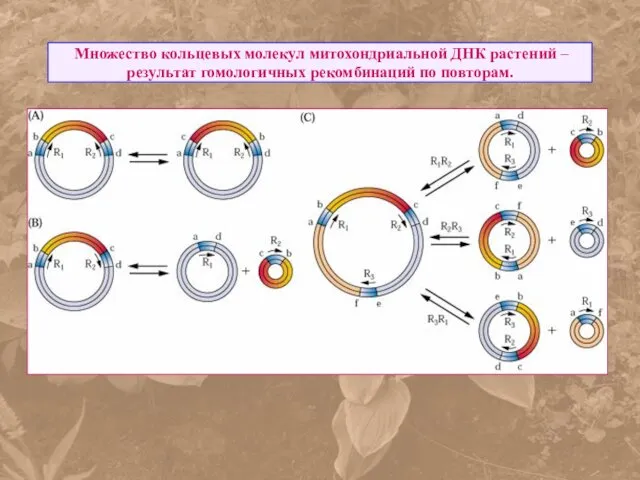
Множество кольцевых молекул митохондриальной ДНК растений – результат гомологичных рекомбинаций
Множество кольцевых молекул митохондриальной ДНК растений – результат гомологичных рекомбинаций
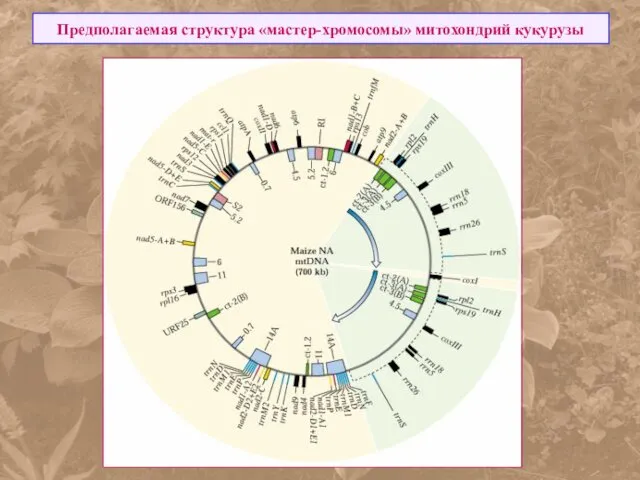
Предполагаемая структура «мастер-хромосомы» митохондрий кукурузы
Предполагаемая структура «мастер-хромосомы» митохондрий кукурузы
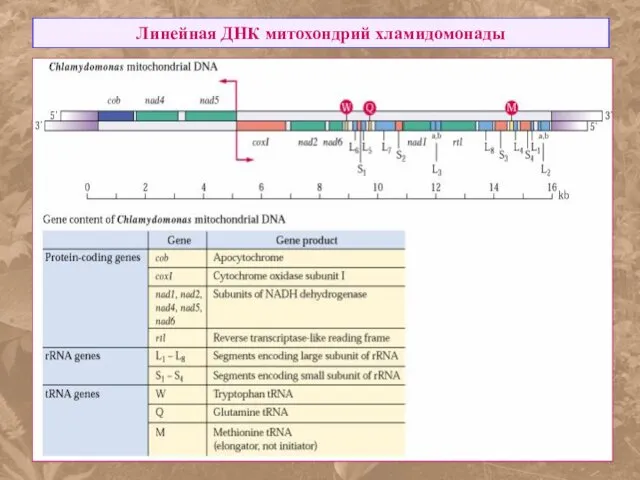
Линейная ДНК митохондрий хламидомонады
Линейная ДНК митохондрий хламидомонады
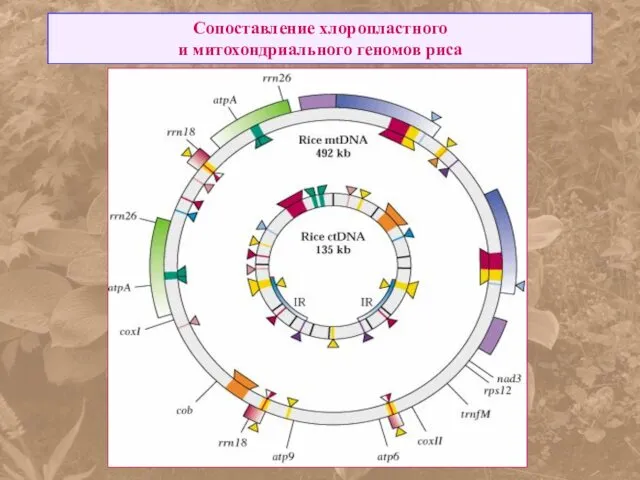
Сопоставление хлоропластного
и митохондриального геномов риса
Сопоставление хлоропластного
и митохондриального геномов риса
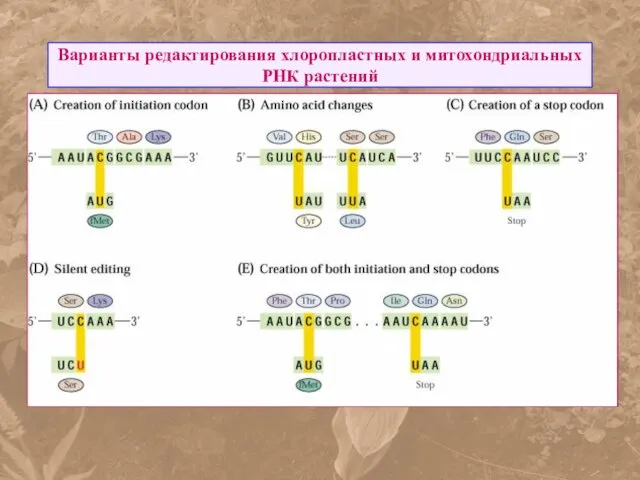
Варианты редактирования хлоропластных и митохондриальных РНК растений
Варианты редактирования хлоропластных и митохондриальных РНК растений

Гены митохондрий
Синтез белка. - 3 гена рРНК (оперон rrn)
- 10
Гены митохондрий
Синтез белка. - 3 гена рРНК (оперон rrn)
- 10

Ядерный генов контролирует :
некоторые т РНК
ДНК – полимеразу
РНК – полимеразу
Некоторые рибосомальные
Ядерный генов контролирует :
некоторые т РНК
ДНК – полимеразу
РНК – полимеразу
Некоторые рибосомальные

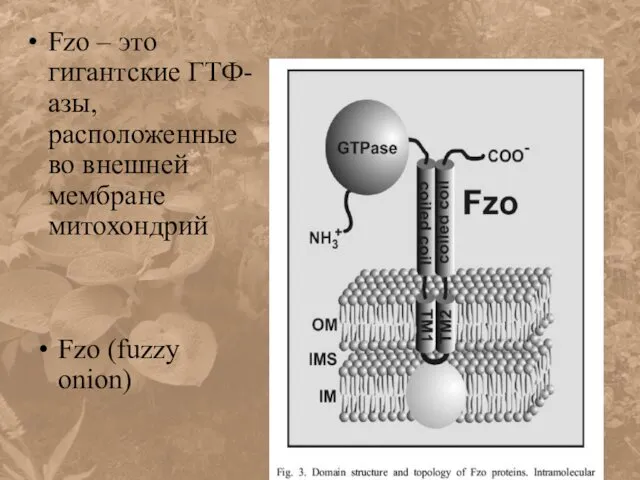
Fzo – это гигантские ГТФ-азы, расположенные во внешней мембране митохондрий
Fzo (fuzzy
Fzo – это гигантские ГТФ-азы, расположенные во внешней мембране митохондрий
Fzo (fuzzy
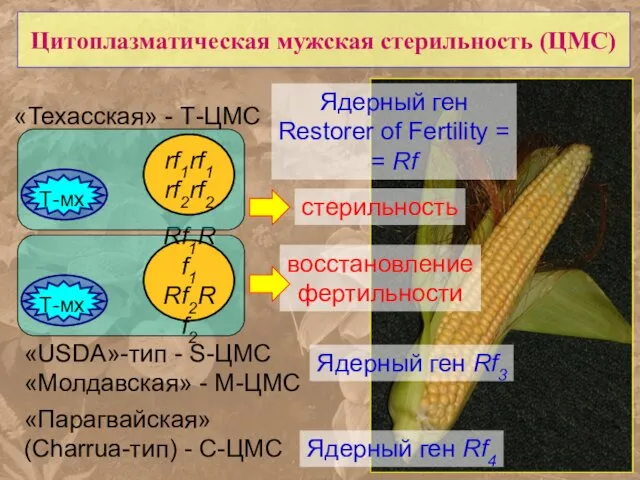
Цитоплазматическая мужская стерильность (ЦМС)
«Техасская» - Т-ЦМС
«USDA»-тип - S-ЦМС
«Молдавская» - M-ЦМС
«Парагвайская»
Цитоплазматическая мужская стерильность (ЦМС)
«Техасская» - Т-ЦМС
«USDA»-тип - S-ЦМС
«Молдавская» - M-ЦМС
«Парагвайская»
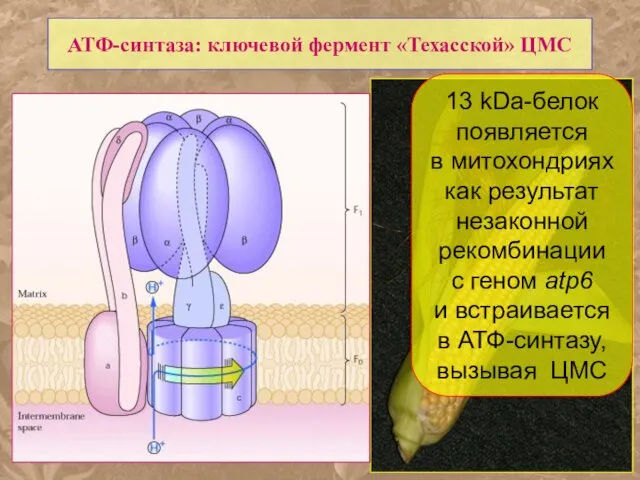
АТФ-синтаза: ключевой фермент «Техасской» ЦМС
13 kDa-белок
появляется
в митохондриях
как результат
АТФ-синтаза: ключевой фермент «Техасской» ЦМС
13 kDa-белок
появляется
в митохондриях
как результат

АТФ-синтаза: ключевой фермент «Техасской» ЦМС
13 kDa-белок
появляется
в митохондриях
как результат
АТФ-синтаза: ключевой фермент «Техасской» ЦМС
13 kDa-белок
появляется
в митохондриях
как результат

АТФ-синтаза: ключевой фермент ЦМС у петунии
У петунии
незаконная
рекомбинация
происходит
с
АТФ-синтаза: ключевой фермент ЦМС у петунии
У петунии
незаконная
рекомбинация
происходит
с

Цитоплазматическая мужская стерильность (ЦМС)
«USDA»-тип - S-ЦМС
«Молдавская» - Т-ЦМС
Ядерный ген Rf3
При
Цитоплазматическая мужская стерильность (ЦМС)
«USDA»-тип - S-ЦМС
«Молдавская» - Т-ЦМС
Ядерный ген Rf3
При

Цитоплазматическая мужская стерильность (ЦМС):
спорофитный и гаметофитный контроль
Стерильность
Полное (!)
восстановление
фертильности
«USDA»-тип - S-ЦМС
«Молдавская»
Цитоплазматическая мужская стерильность (ЦМС):
спорофитный и гаметофитный контроль
Стерильность
Полное (!)
восстановление
фертильности
«USDA»-тип - S-ЦМС
«Молдавская»
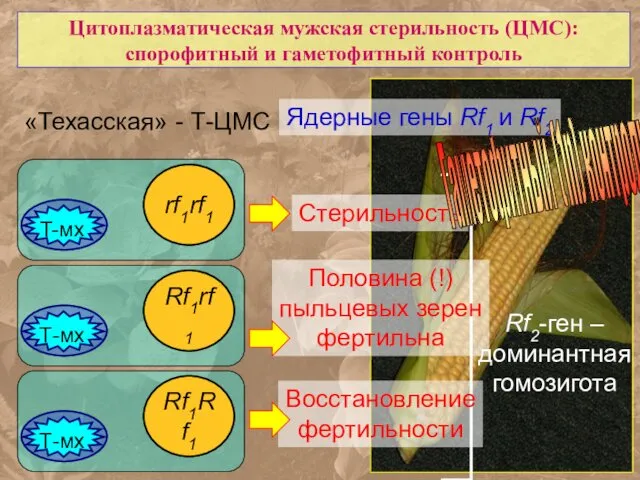
Цитоплазматическая мужская стерильность (ЦМС):
спорофитный и гаметофитный контроль
«Техасская» - Т-ЦМС
Ядерные гены
Цитоплазматическая мужская стерильность (ЦМС):
спорофитный и гаметофитный контроль
«Техасская» - Т-ЦМС
Ядерные гены


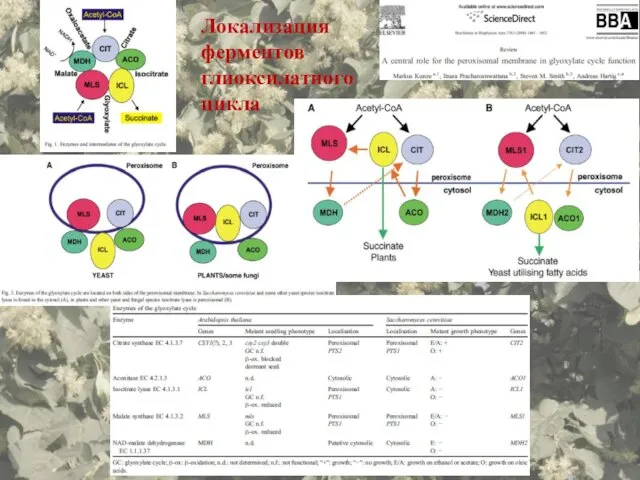
Локализация ферментов глиоксилатного цикла
Локализация ферментов глиоксилатного цикла
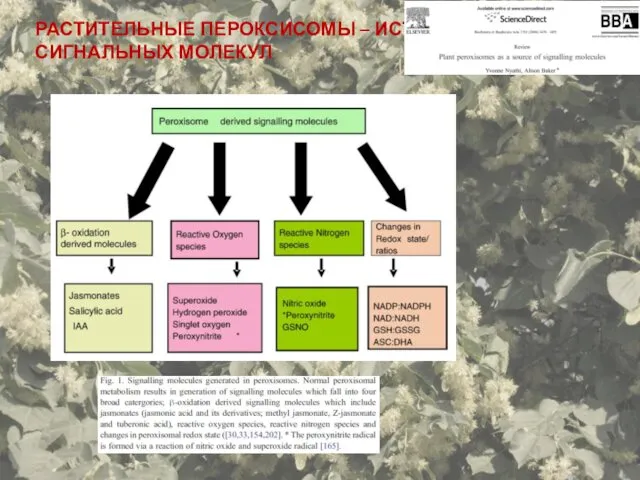
РАСТИТЕЛЬНЫЕ ПЕРОКСИСОМЫ – ИСТОЧНИК СИГНАЛЬНЫХ МОЛЕКУЛ
РАСТИТЕЛЬНЫЕ ПЕРОКСИСОМЫ – ИСТОЧНИК СИГНАЛЬНЫХ МОЛЕКУЛ
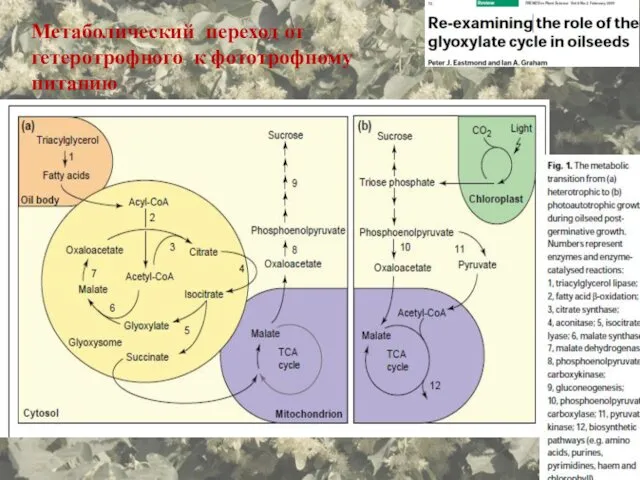
Метаболический переход от гетеротрофного к фототрофному питанию
Метаболический переход от гетеротрофного к фототрофному питанию
 Любопытные факты о страусах
Любопытные факты о страусах Чай. Сорта чая. Приготовление чая
Чай. Сорта чая. Приготовление чая Идея развития органического мира
Идея развития органического мира Сенсорные системы
Сенсорные системы Тигр (лат. Panthera tigris)
Тигр (лат. Panthera tigris) Спортивная биохимия. (Часть 3)
Спортивная биохимия. (Часть 3) Негізгі тағамдық азықтардың тамақтық және биологиялық құндылығы
Негізгі тағамдық азықтардың тамақтық және биологиялық құндылығы Матричные процессы. Этапы транскрипции
Матричные процессы. Этапы транскрипции Шоюн Александр Саноолович — поэт, прозаик
Шоюн Александр Саноолович — поэт, прозаик Отряд грызунов
Отряд грызунов Современные исследования в кролиководстве
Современные исследования в кролиководстве Круглые черви. Класс: нематоды представители: аскарида человеческая острица, ришта
Круглые черви. Класс: нематоды представители: аскарида человеческая острица, ришта Презентация на тему "Покрытосеменные" - скачать презентации по Биологии
Презентация на тему "Покрытосеменные" - скачать презентации по Биологии Биосоциальная природа человека. Лекция 2
Биосоциальная природа человека. Лекция 2 Презентация на тему "Семейство Астровые, или Сложноцветные" - скачать бесплатно презентации по Биологии
Презентация на тему "Семейство Астровые, или Сложноцветные" - скачать бесплатно презентации по Биологии Презентация на тему "Животные со дна океана" - скачать бесплатно презентации по Биологии
Презентация на тему "Животные со дна океана" - скачать бесплатно презентации по Биологии Презентация на тему "Сообщества живых организмов" - скачать бесплатно презентации по Биологии
Презентация на тему "Сообщества живых организмов" - скачать бесплатно презентации по Биологии Наследование признаков, сцепленных с полом. (Задачи № 322, 335, 348)
Наследование признаков, сцепленных с полом. (Задачи № 322, 335, 348) Характеристики и виды обмена
Характеристики и виды обмена Трансгенные животные и растения
Трансгенные животные и растения Интересные факты о насекомых
Интересные факты о насекомых ФИЗИОЛОГИЯ И БИОХИМИЯ МИКРООРГАНИЗМОВ
ФИЗИОЛОГИЯ И БИОХИМИЯ МИКРООРГАНИЗМОВ Разнообразие круглых червей
Разнообразие круглых червей Конечный мозг. Нервная система
Конечный мозг. Нервная система Интеграционная образовательно-исследовательская программа в области молекулярной биологии
Интеграционная образовательно-исследовательская программа в области молекулярной биологии Строение эукариотической клетки
Строение эукариотической клетки ДНК и РНК нуклеиновые кислоты
ДНК и РНК нуклеиновые кислоты Царство (подцарство) Простейшие
Царство (подцарство) Простейшие