- Эндокринная система

Содержание
- 2. Эндокринология Начало этой науки положили опыты немецкого физиолога Адольфа Бер-тольда, которому в 1849 году удалось установить,
- 3. Опыт О. Леви. Нобелевская премия по физиологии и медицине, Нобелевская премия по физиологии и медицине, 1936
- 4. Что такое гормоны? Гормоны - (от греч. hormáo - привожу в движение, побуждаю), биологически активные вещества,
- 5. Физиологическое действие гормонов направлено на: 1) обеспечение гуморальной, т.е. осуществляемой через кровь, регуляции биологических процессов. 2)
- 6. Эффекты действия гормонов Метаболическое действие – изменяют проницаемость мембран клетки и активность транспортных систем; активность ферментов,
- 7. 4. Корригирующее действие – изменяет интенсивность функции органа (например, продукцию инсулина, в ответ на образование гликогена
- 8. Классическим гормонам присущ ряд признаков: Дистантность действия – синтез в железах внутренней секреции, а регуляция отдаленных
- 9. Гармональный баланс В нормальном состоянии существует строгий баланс между активностью эндокринных желез, состоянием нервной системы и
- 10. Регуляция уровня гормонов в организме Изменение концентрации метаболитов в клетках-мишенях по механизму отрицательной обратной связи подавляет
- 11. Организация нервно-гормональной регуляции Существует строгая иерархия или соподчиненность гормонов. Поддержание уровня гормонов в организме в большинстве
- 13. I. Центральные регуляторные образования эндокринной системы: гипоталамус (нейросекреторные ядра); гипофиз (аденогипофиз и нейрогипофиз); эпифиз. II. Периферические
- 14. Различные формы гуморальных связей между клетками
- 19. Классификация гормонов по химическому строению; по биологическим функциям; по механизму действия;
- 20. 1. По химической структуре гормоны, подразделяют на: пептиды, производные аминокислот, стероиды, производные арахидоновой кислоты.
- 21. Пептидные гормоны - секретируются путем экзоцитоза, - относятся к полярным веществам, поэтому не могут проникать через
- 22. Некоторые представители пептидных гормонов: тиролиберина (пироглу-гис-про-NН2), инсулина и соматостатина.
- 23. Производные аминокислот: Производные тирозина – йодсодержащие гормоны щитовидной железы, норадреналин, адреналин и дофамин. Молекулы тироксина (T4)
- 24. Стероидные гормоны - производные холестерола (минералокортикоиды, глюкокортикоиды, андрогены, эстрогены, прогестины, кальцитриол). - относятся к неполярным, поэтому
- 25. Производные арахидоновой кислоты (эйкозаноиды) Предшественником всех эйкозаноидов является арахидоновая кислота. Они делятся на 3 группы: простагландины,
- 26. Сигнальные пути арахидоновой кислоты ПГ - простагландин, ЛГ - лейкотриен, ГПЭТЕ - гидропероксиэйкозатетраеноат, ГЭТЕ - гидроксиэйкозатетраеноат,
- 27. Характеристика разных групп эйказоноидов Простагландины (Pg) — синтезируются практически во всех клетках, кроме эритроцитов и лимфоцитов.
- 28. Тромбоксаны и лейкотриены Тромбоксаны (Tx) образуются в тромбоцитах, стимулируют их агрегацию и вызывают сужение мелких сосудов.
- 29. 2. биологические функции гормонов
- 30. 3. Основные механизмы нейроэндокринной регуляции клеток
- 32. По механизму действия различают гормоны: 1)взаимодействующие с мембранными рецепторами (пептидные гормоны, адреналин, эйкозаноиды); 2)взаимодействующие с внутриклеточными
- 33. Передача гормонального сигнала через внутриклеточные рецепторы для стероидных гормонов (гормоны коры надпочечников и половые гормоны), тиреодных
- 34. Передача гормонального сигнала через ядерный рецептор.
- 35. Передача гормонального сигнала через мембранные рецепторы Передача информации от первичного посредника гормона осуществляется через рецептор. Этот
- 36. Сигнал от гормона трансформируется в виде изменения концентрации вторичных посредников – цАМФ, цГТФ, ИФ3, ДАГ, Са2+
- 37. Вторичные посредники гормонов Вторичные посредники (вторичные мессенджеры, англ. second messengers) — это малые сигнальные молекулы, которые
- 38. КЛАССИЧЕСКИЕ ВТОРИЧНЫЕ МЕССЕНДЖЕРЫ цАМФ (циклический аденозинмонофосфат); цГМФ (циклический гуанозинмонофосфат); ИФ3 (инозитолтрифосфат ); ДИАЦИЛГЛИЦЕРОЛ (ДАГ) ИОНЫ КАЛЬЦИЯ
- 40. G-белки (англ. G proteins) — это семейство белков, относящихся к ГТФазам и функционирующих в качестве посредников
- 41. G-белки делятся на две основных группы — гетеротримерные («большие») и «малые». Гетеротримерные G-белки — это белки
- 42. Цикл работы гетеротримерного G-белка, запускающего дальнейшую цепь событий с помощью своей α-субъединицы.
- 43. Принцип работы мономерных ГТФ-связывающих белков (мономерных G-белков).
- 44. Система аденилатциклаза - цАМФ Так действуют активирующие аденилатциклазу гормоны (глюкагон, тиротропин, паратирин и др.). 1. Образование
- 45. Система аденилатциклаза - цАМФ
- 49. Холера – это инфекция, вызывающая диарею, которая может быстро привести к обезвоживанию организма и смерти. Диарея
- 50. АДЦиклазы – 9 изоформ (от I до IX) Характеризуется в основном по их чувствительности к Са2+
- 51. Фосфодиастеразы (PDES) PDE1,-2,-3,-4,-5,-6,-7. 7 генных семейств определяют синтез изоформ 1 – 7, где >50 различных PDES.
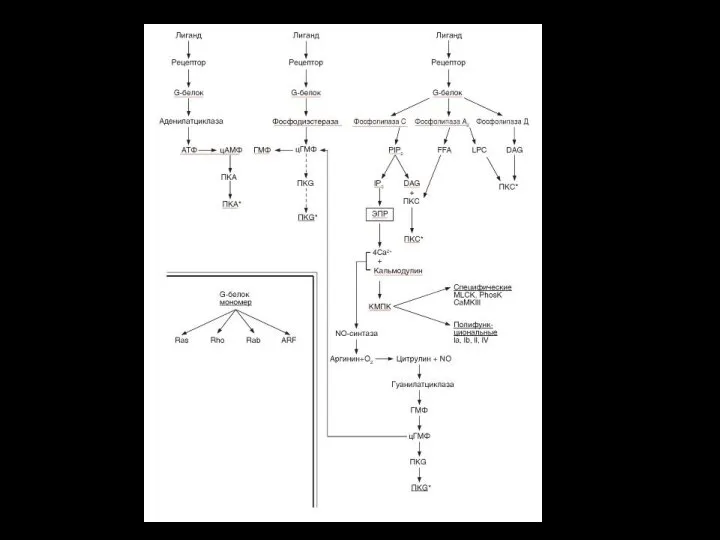
- 52. Система фосфолипаза С - ИФ3 -ДГ Этапы: 1. Образование гормон-рецепторного комплекса с G- белком активирует мембранную
- 54. Медленная и продолжительная активация РКC
- 55. ИФ3 связывается с рецептором – каналоформером, расположенным на эндоплазматической сети и вызывает выход Са2+ в цитоплазму.
- 56. Система кальций-кальмодулин Вызываемое гормональным стимулом кратковременное увеличение в клетке кальция и его связывание с кальмодулином является
- 57. Система кальций-кальмодулин Этапы: Са2+ поступает в клетку после образования гормон-рецепторного комплекса : а) либо из внеклеточной
- 58. Кальмодулин EF - Са2+-связывающие домены кальмодулина
- 59. Система гуанилатциклаза - цГМФ Этапы: 1. Активация гуанилатциклазы происходит опосредованно через ионизированный кальций (ацетилхолин, натрийуретический гормон)
- 61. NO как вторичный мессенджер Образуется при окислении L-аргинина ферментом NO-синтазой (NOS), присутствующего в нервной ткани, эндотелии
- 62. Механизм действия В клетках-мишенях, например, эндотелиальных клетках NO взаимодействует с входящим в активный центр гуанилатциклазы ионом
- 65. Взаимосвязи вторичных посредников Между вторичными посредниками устанавливаются различные взаимоотношения: равнозначное участие, когда разные посредники необходимы для
- 71. Скачать презентацию

Эндокринология
Начало этой науки положили опыты немецкого физиолога Адольфа Бер-тольда, которому
Эндокринология
Начало этой науки положили опыты немецкого физиолога Адольфа Бер-тольда, которому
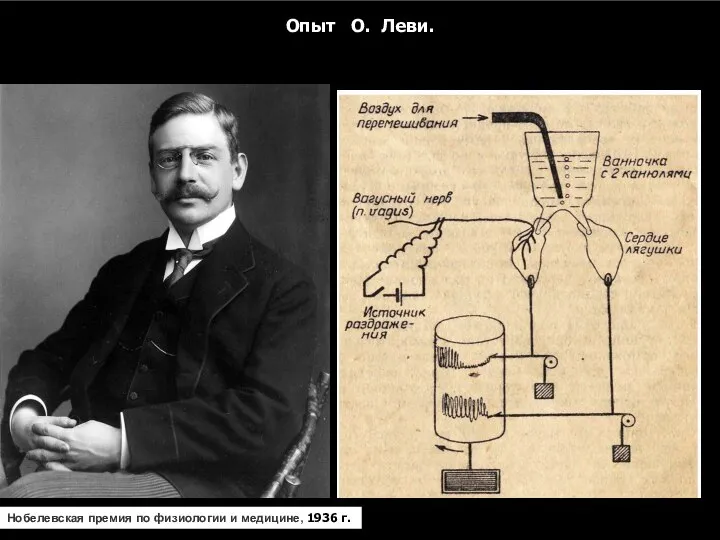
Опыт О. Леви.
Нобелевская премия по физиологии и медицине,
Нобелевская премия по физиологии и медицине, 1936 г.
Опыт О. Леви.
Нобелевская премия по физиологии и медицине,
Нобелевская премия по физиологии и медицине, 1936 г.

Что такое гормоны?
Гормоны - (от греч. hormáo - привожу в
Что такое гормоны?
Гормоны - (от греч. hormáo - привожу в

Физиологическое действие гормонов направлено на:
1) обеспечение гуморальной, т.е. осуществляемой через
Физиологическое действие гормонов направлено на:
1) обеспечение гуморальной, т.е. осуществляемой через

Эффекты действия гормонов
Метаболическое действие – изменяют проницаемость мембран клетки и активность
Эффекты действия гормонов
Метаболическое действие – изменяют проницаемость мембран клетки и активность

4. Корригирующее действие – изменяет интенсивность функции органа (например, продукцию инсулина,
4. Корригирующее действие – изменяет интенсивность функции органа (например, продукцию инсулина,

Классическим гормонам присущ ряд признаков:
Дистантность действия – синтез в железах внутренней
Классическим гормонам присущ ряд признаков:
Дистантность действия – синтез в железах внутренней

Гармональный баланс
В нормальном состоянии существует строгий баланс между активностью эндокринных
Гармональный баланс
В нормальном состоянии существует строгий баланс между активностью эндокринных

Регуляция уровня гормонов в организме
Изменение концентрации метаболитов в клетках-мишенях по механизму
Регуляция уровня гормонов в организме
Изменение концентрации метаболитов в клетках-мишенях по механизму

Организация нервно-гормональной регуляции
Существует строгая иерархия или соподчиненность гормонов.
Поддержание уровня гормонов в
Организация нервно-гормональной регуляции
Существует строгая иерархия или соподчиненность гормонов.
Поддержание уровня гормонов в
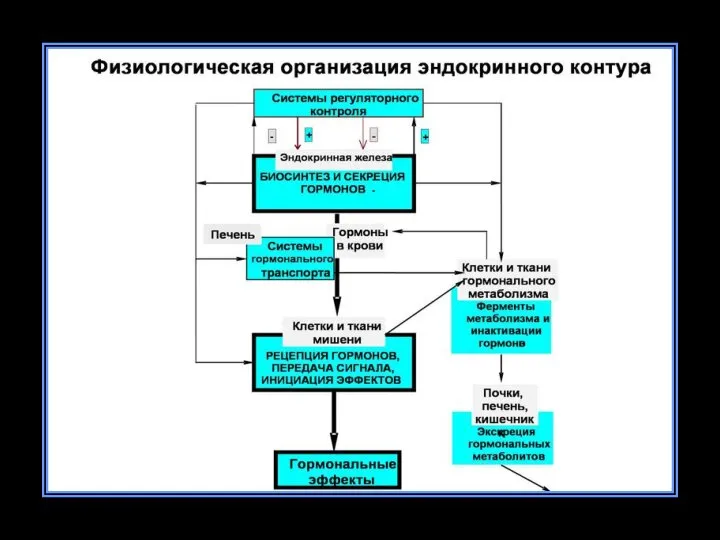

I. Центральные регуляторные образования эндокринной системы:
гипоталамус (нейросекреторные ядра);
гипофиз (аденогипофиз
I. Центральные регуляторные образования эндокринной системы:
гипоталамус (нейросекреторные ядра);
гипофиз (аденогипофиз
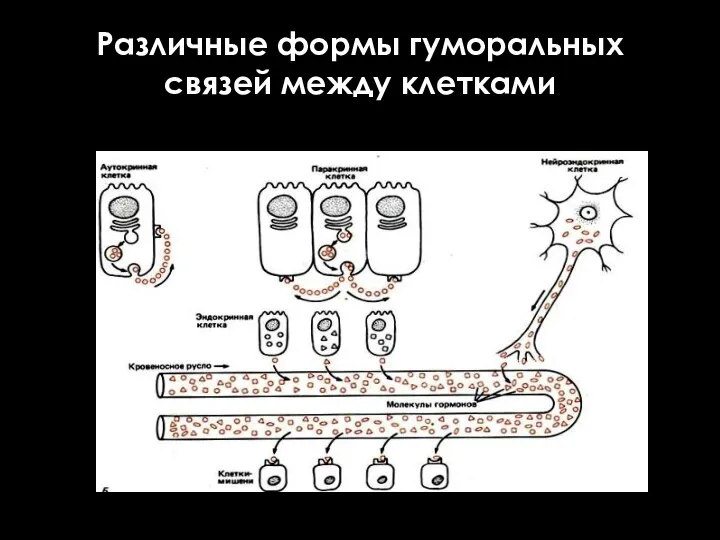
Различные формы гуморальных связей между клетками
Различные формы гуморальных связей между клетками




Классификация гормонов
по химическому строению;
по биологическим функциям;
по механизму действия;
Классификация гормонов
по химическому строению;
по биологическим функциям;
по механизму действия;

1. По химической структуре гормоны, подразделяют на:
пептиды,
производные аминокислот,
стероиды,
производные арахидоновой
1. По химической структуре гормоны, подразделяют на:
пептиды,
производные аминокислот,
стероиды,
производные арахидоновой

Пептидные гормоны
- секретируются путем экзоцитоза,
- относятся к полярным веществам, поэтому не
Пептидные гормоны
- секретируются путем экзоцитоза,
- относятся к полярным веществам, поэтому не
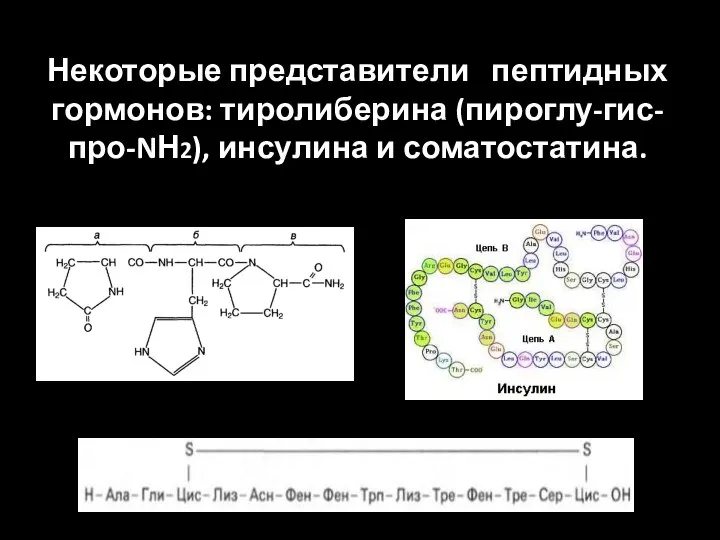
Некоторые представители пептидных
гормонов: тиролиберина (пироглу-гис-про-NН2), инсулина и соматостатина.
Некоторые представители пептидных
гормонов: тиролиберина (пироглу-гис-про-NН2), инсулина и соматостатина.
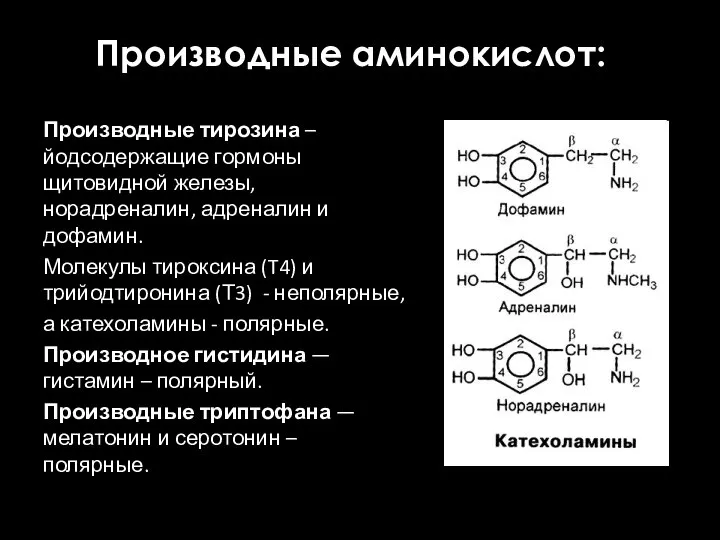
Производные аминокислот:
Производные тирозина – йодсодержащие гормоны щитовидной железы, норадреналин, адреналин и
Производные аминокислот:
Производные тирозина – йодсодержащие гормоны щитовидной железы, норадреналин, адреналин и
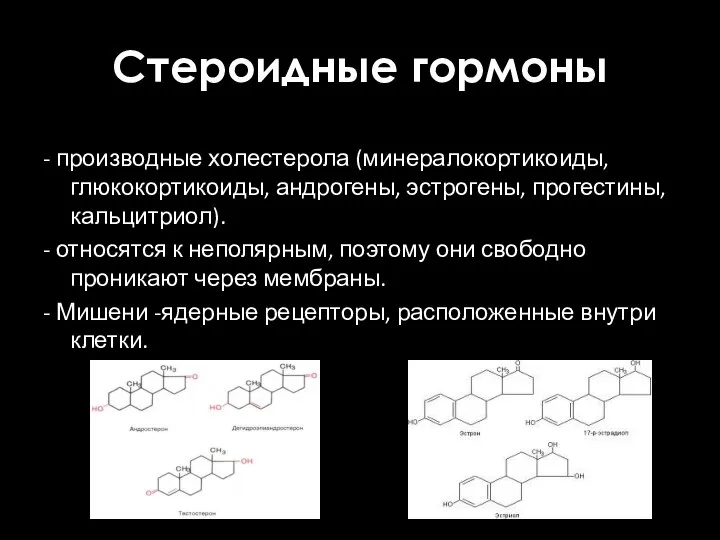
Стероидные гормоны
- производные холестерола (минералокортикоиды, глюкокортикоиды, андрогены, эстрогены, прогестины, кальцитриол).
-
Стероидные гормоны
- производные холестерола (минералокортикоиды, глюкокортикоиды, андрогены, эстрогены, прогестины, кальцитриол).
-

Производные арахидоновой кислоты (эйкозаноиды)
Предшественником всех эйкозаноидов является арахидоновая кислота.
Они
Производные арахидоновой кислоты (эйкозаноиды)
Предшественником всех эйкозаноидов является арахидоновая кислота.
Они
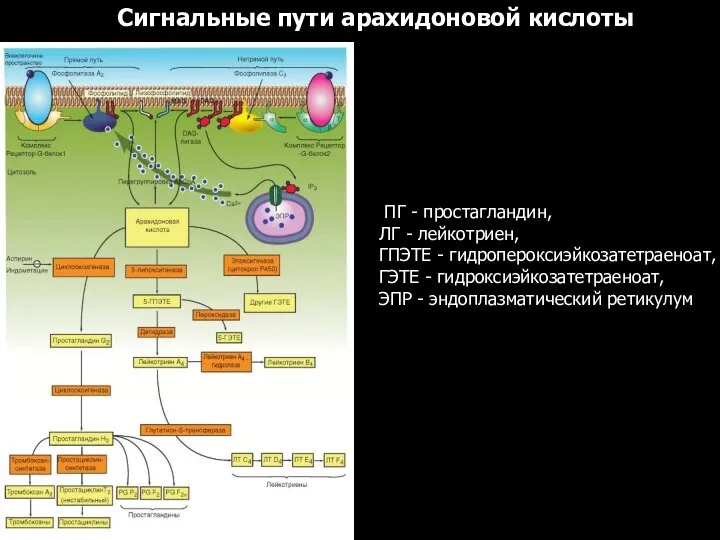
Сигнальные пути арахидоновой кислоты
ПГ - простагландин,
ЛГ - лейкотриен,
ГПЭТЕ
Сигнальные пути арахидоновой кислоты
ПГ - простагландин,
ЛГ - лейкотриен,
ГПЭТЕ

Характеристика разных групп эйказоноидов
Простагландины (Pg) — синтезируются практически во всех клетках, кроме
Характеристика разных групп эйказоноидов
Простагландины (Pg) — синтезируются практически во всех клетках, кроме

Тромбоксаны и лейкотриены
Тромбоксаны (Tx) образуются в тромбоцитах, стимулируют их агрегацию и
Тромбоксаны и лейкотриены
Тромбоксаны (Tx) образуются в тромбоцитах, стимулируют их агрегацию и

2. биологические функции гормонов
2. биологические функции гормонов
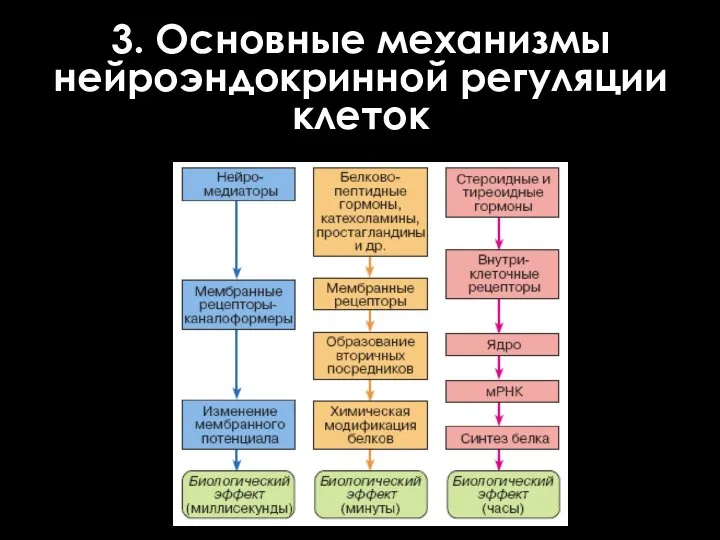
3. Основные механизмы нейроэндокринной регуляции клеток
3. Основные механизмы нейроэндокринной регуляции клеток
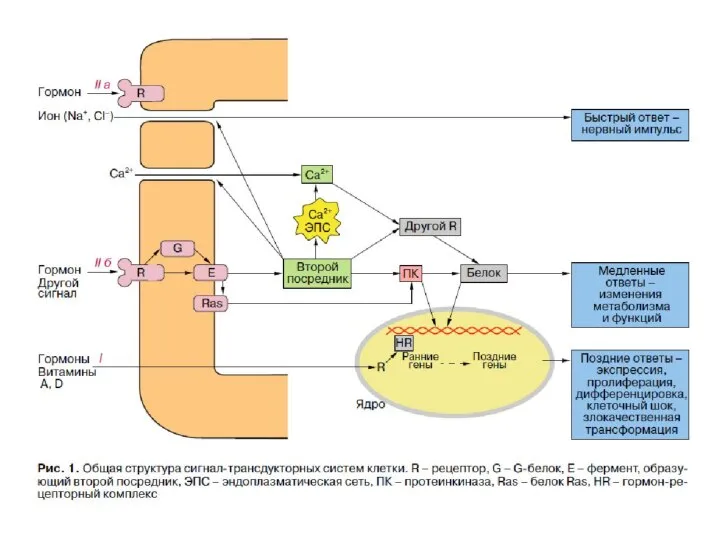

По механизму действия различают гормоны:
1)взаимодействующие с мембранными рецепторами (пептидные гормоны,
По механизму действия различают гормоны: 1)взаимодействующие с мембранными рецепторами (пептидные гормоны,
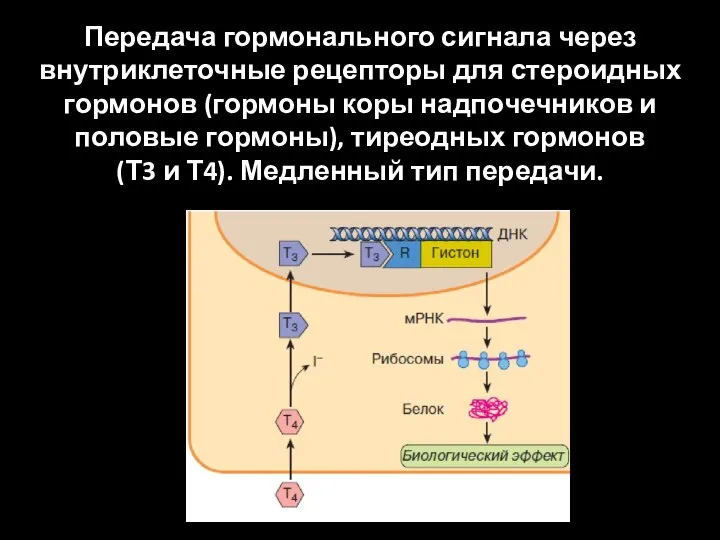
Передача гормонального сигнала через
внутриклеточные рецепторы для стероидных гормонов (гормоны коры
Передача гормонального сигнала через внутриклеточные рецепторы для стероидных гормонов (гормоны коры
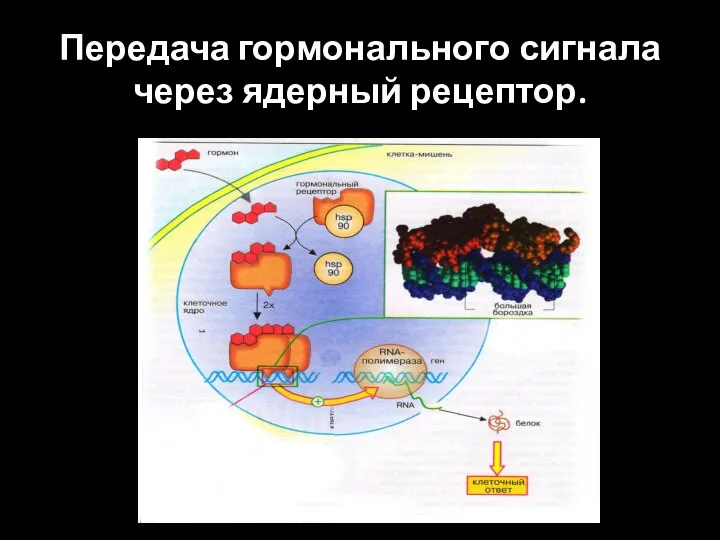
Передача гормонального сигнала через ядерный рецептор.
Передача гормонального сигнала через ядерный рецептор.

Передача гормонального сигнала через мембранные рецепторы
Передача информации от первичного посредника гормона
Передача гормонального сигнала через мембранные рецепторы
Передача информации от первичного посредника гормона
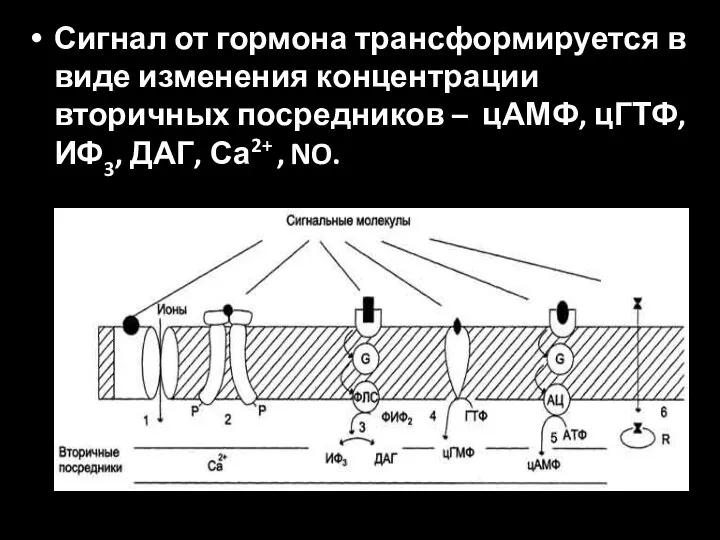
Сигнал от гормона трансформируется в виде изменения концентрации вторичных посредников –
Сигнал от гормона трансформируется в виде изменения концентрации вторичных посредников –

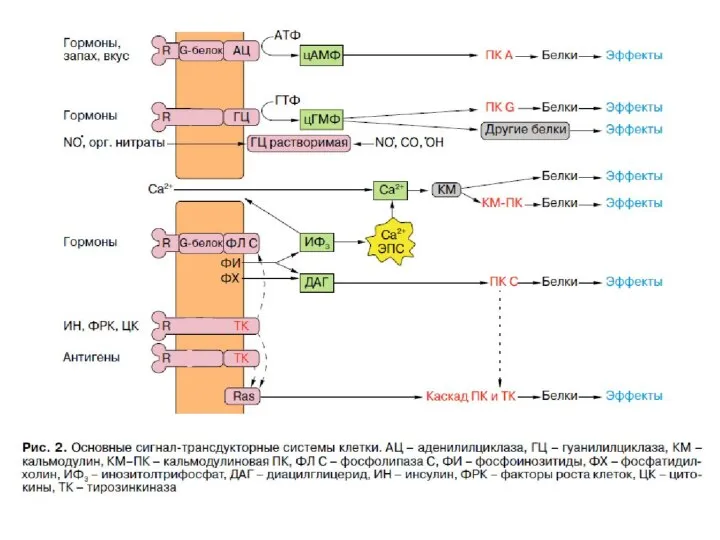
Вторичные посредники гормонов
Вторичные посредники (вторичные мессенджеры, англ. second messengers) — это малые сигнальные
Вторичные посредники гормонов
Вторичные посредники (вторичные мессенджеры, англ. second messengers) — это малые сигнальные

КЛАССИЧЕСКИЕ ВТОРИЧНЫЕ МЕССЕНДЖЕРЫ
цАМФ (циклический аденозинмонофосфат);
цГМФ (циклический гуанозинмонофосфат);
ИФ3 (инозитолтрифосфат );
ДИАЦИЛГЛИЦЕРОЛ (ДАГ)
ИОНЫ КАЛЬЦИЯ
КЛАССИЧЕСКИЕ ВТОРИЧНЫЕ МЕССЕНДЖЕРЫ
цАМФ (циклический аденозинмонофосфат);
цГМФ (циклический гуанозинмонофосфат);
ИФ3 (инозитолтрифосфат );
ДИАЦИЛГЛИЦЕРОЛ (ДАГ)
ИОНЫ КАЛЬЦИЯ
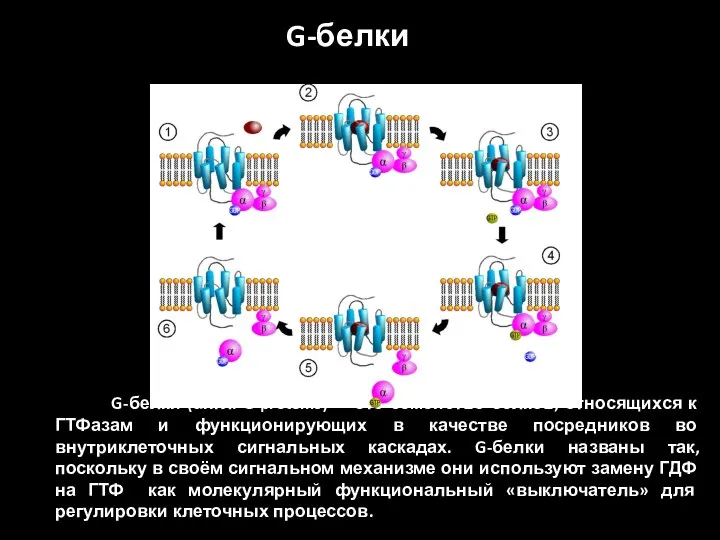
G-белки (англ. G proteins) — это семейство белков, относящихся к
G-белки (англ. G proteins) — это семейство белков, относящихся к

G-белки делятся на две основных группы — гетеротримерные («большие») и «малые».
G-белки делятся на две основных группы — гетеротримерные («большие») и «малые».
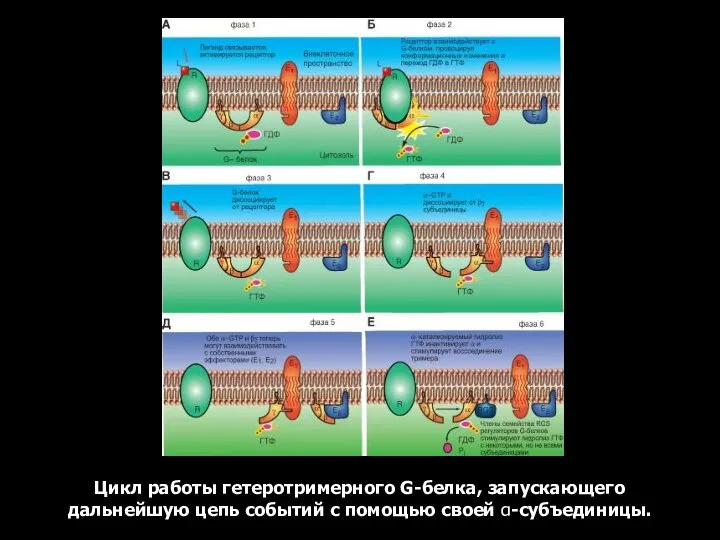
Цикл работы гетеротримерного G-белка, запускающего дальнейшую цепь событий с помощью своей α-субъединицы.
Цикл работы гетеротримерного G-белка, запускающего дальнейшую цепь событий с помощью своей α-субъединицы.
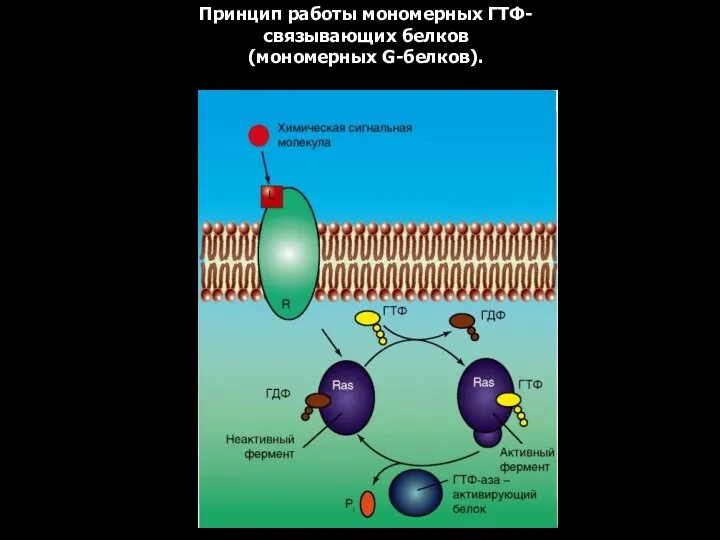
Принцип работы мономерных ГТФ-связывающих белков (мономерных G-белков).
Принцип работы мономерных ГТФ-связывающих белков (мономерных G-белков).
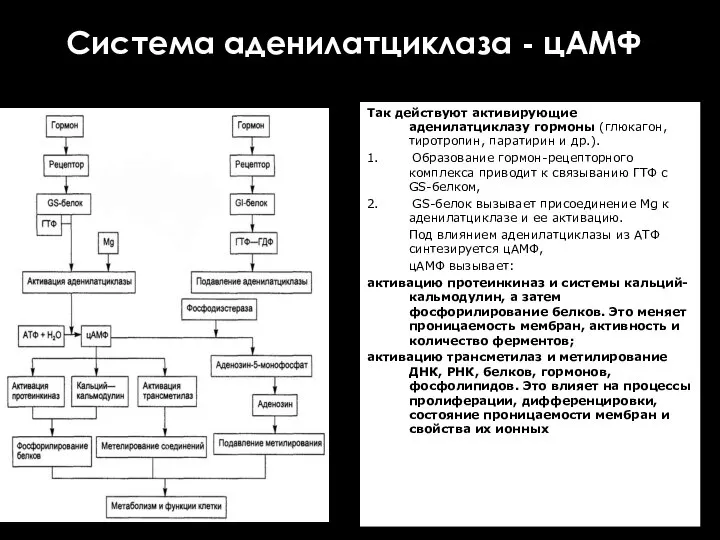
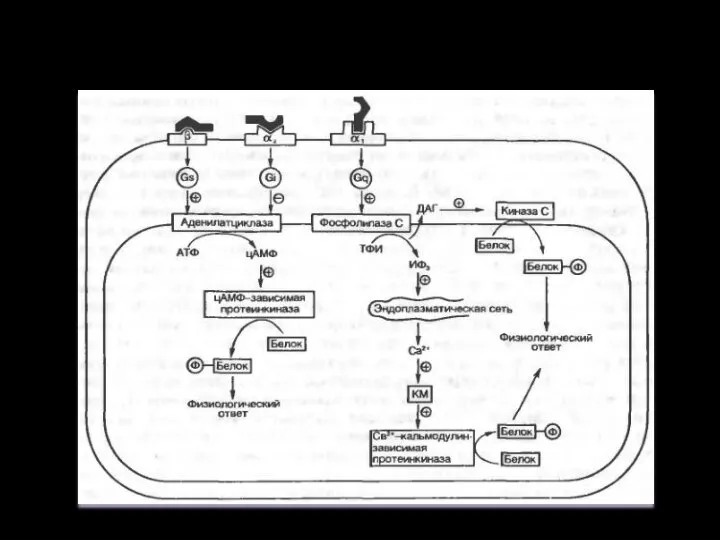
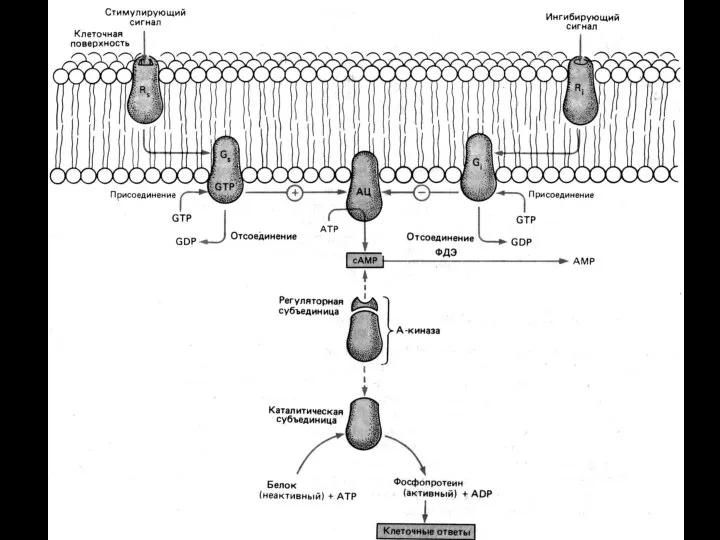
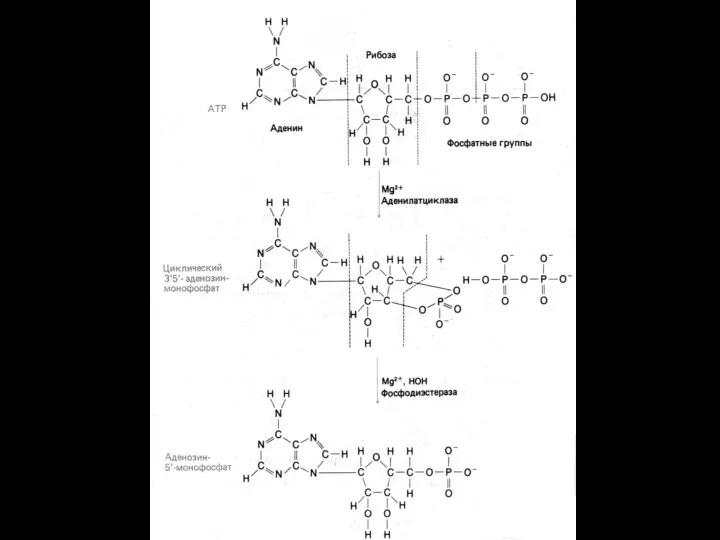
Система аденилатциклаза - цАМФ
Так действуют активирующие аденилатциклазу гормоны (глюкагон, тиротропин,
Система аденилатциклаза - цАМФ
Так действуют активирующие аденилатциклазу гормоны (глюкагон, тиротропин,
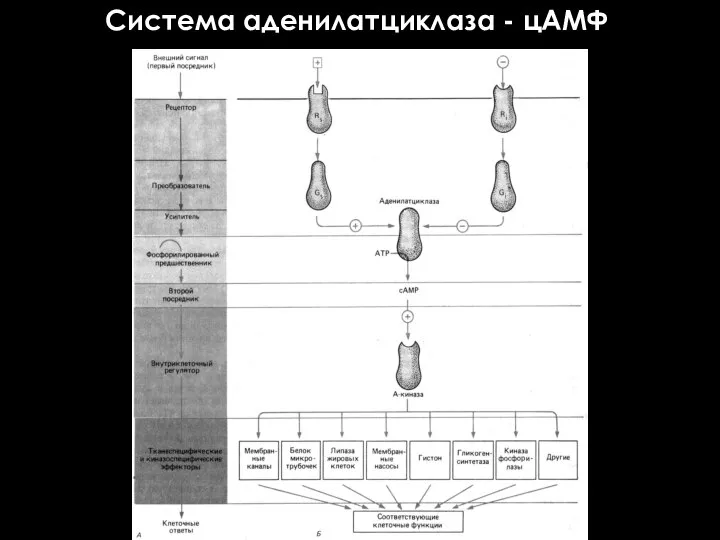
Система аденилатциклаза - цАМФ
Система аденилатциклаза - цАМФ
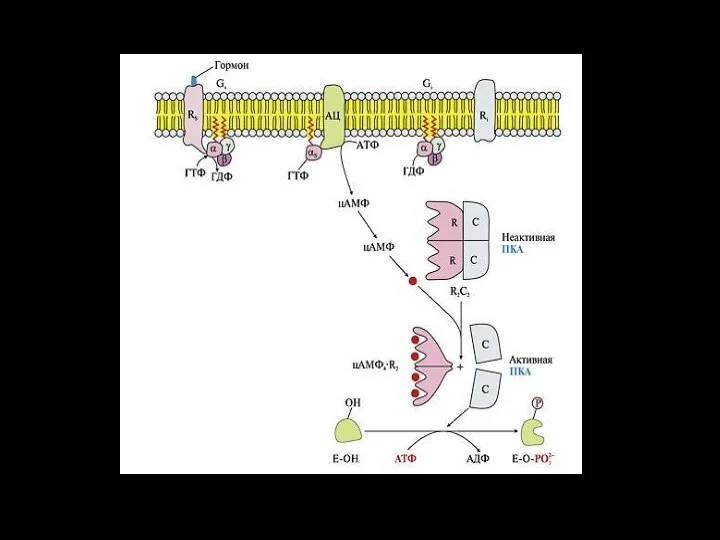

Холера – это инфекция, вызывающая диарею, которая может быстро привести к
Холера – это инфекция, вызывающая диарею, которая может быстро привести к

АДЦиклазы – 9 изоформ (от I до IX)
Характеризуется в основном по
АДЦиклазы – 9 изоформ (от I до IX)
Характеризуется в основном по

Фосфодиастеразы (PDES) PDE1,-2,-3,-4,-5,-6,-7.
7 генных семейств определяют синтез изоформ 1 – 7,
Фосфодиастеразы (PDES) PDE1,-2,-3,-4,-5,-6,-7.
7 генных семейств определяют синтез изоформ 1 – 7,
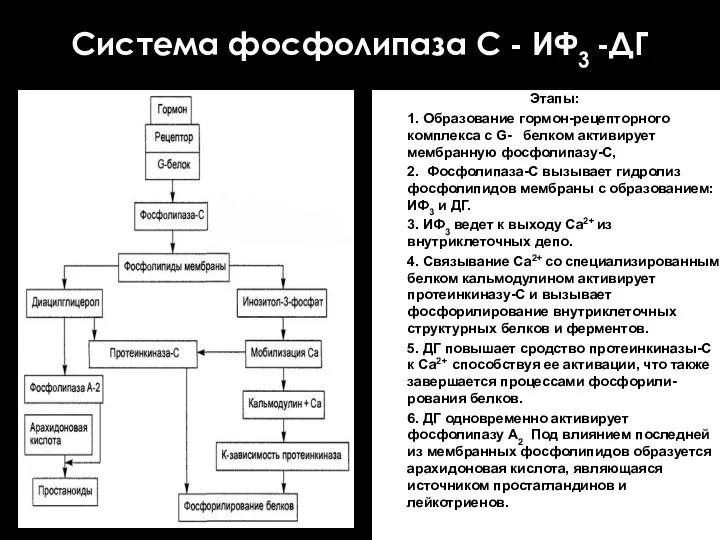
Система фосфолипаза С - ИФ3 -ДГ
Этапы:
1. Образование гормон-рецепторного комплекса с
Система фосфолипаза С - ИФ3 -ДГ
Этапы:
1. Образование гормон-рецепторного комплекса с
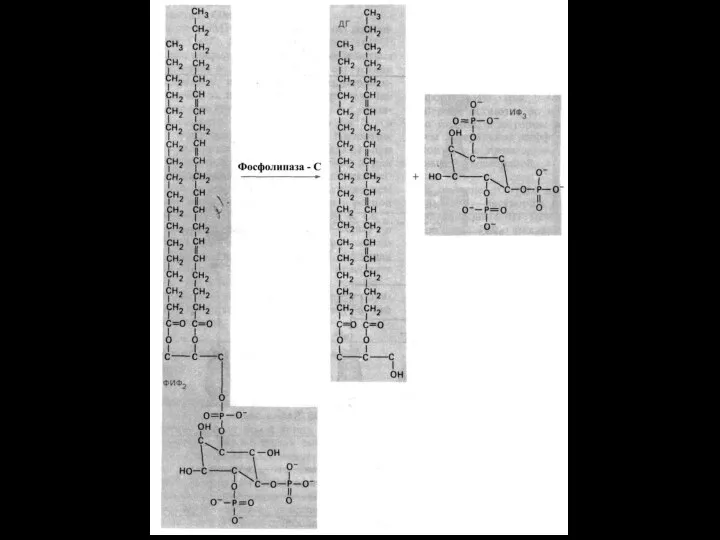
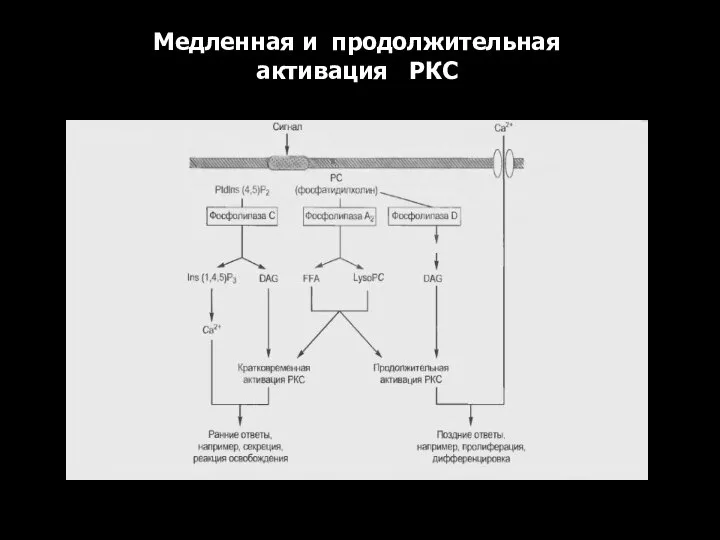
Медленная и продолжительная активация РКC
Медленная и продолжительная активация РКC
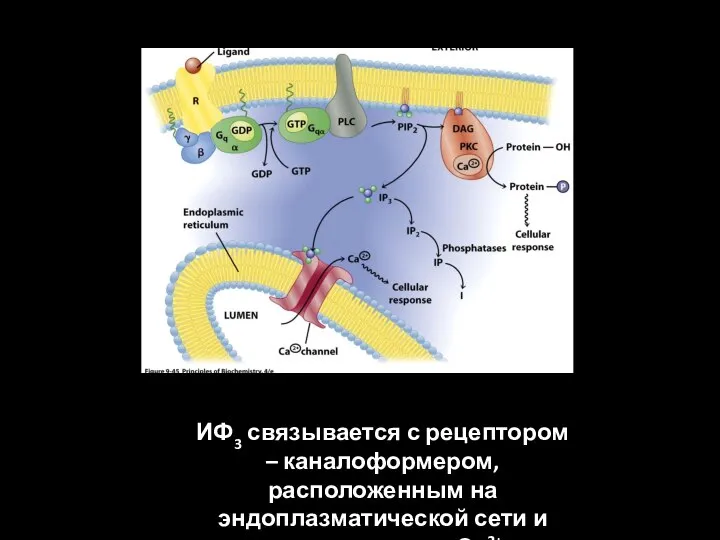
ИФ3 связывается с рецептором – каналоформером, расположенным на эндоплазматической сети и
ИФ3 связывается с рецептором – каналоформером, расположенным на эндоплазматической сети и
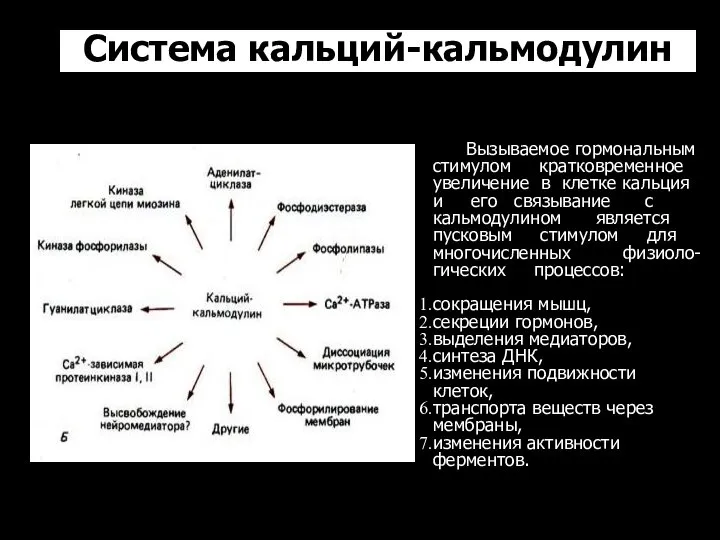
Система кальций-кальмодулин
Вызываемое гормональным
стимулом кратковременное
увеличение в клетке кальция
и
Система кальций-кальмодулин
Вызываемое гормональным
стимулом кратковременное
увеличение в клетке кальция
и
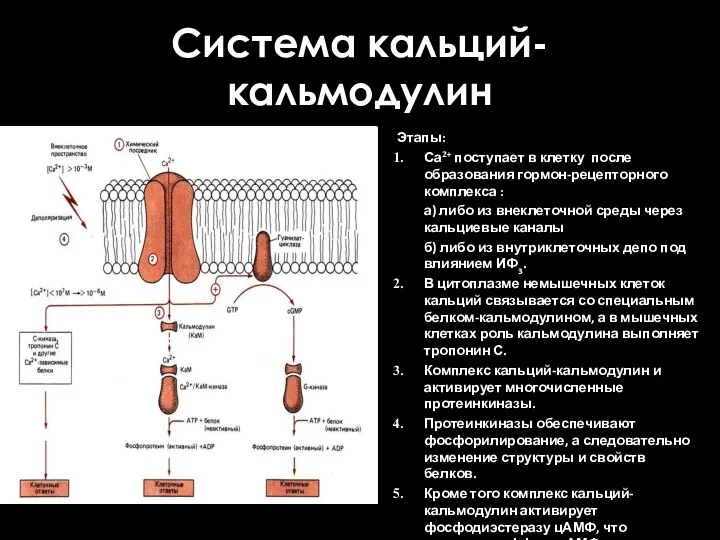
Система кальций-кальмодулин
Этапы:
Са2+ поступает в клетку после образования гормон-рецепторного комплекса :
а) либо
Система кальций-кальмодулин
Этапы:
Са2+ поступает в клетку после образования гормон-рецепторного комплекса :
а) либо
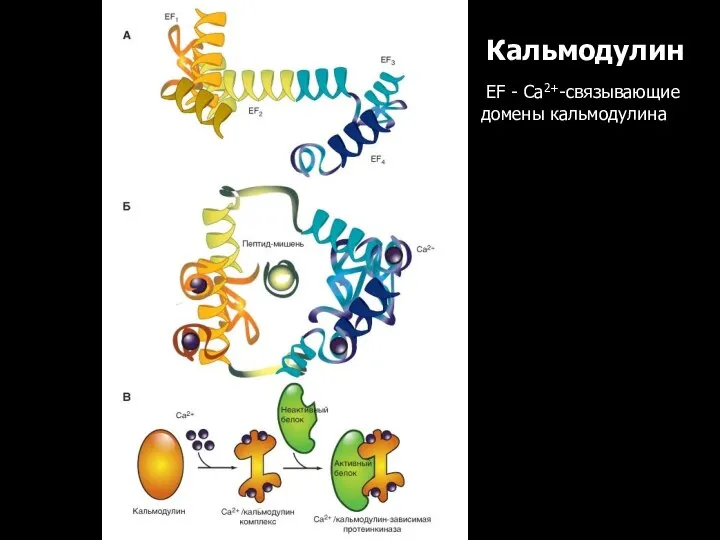
Кальмодулин
EF - Са2+-связывающие домены кальмодулина
Кальмодулин
EF - Са2+-связывающие домены кальмодулина
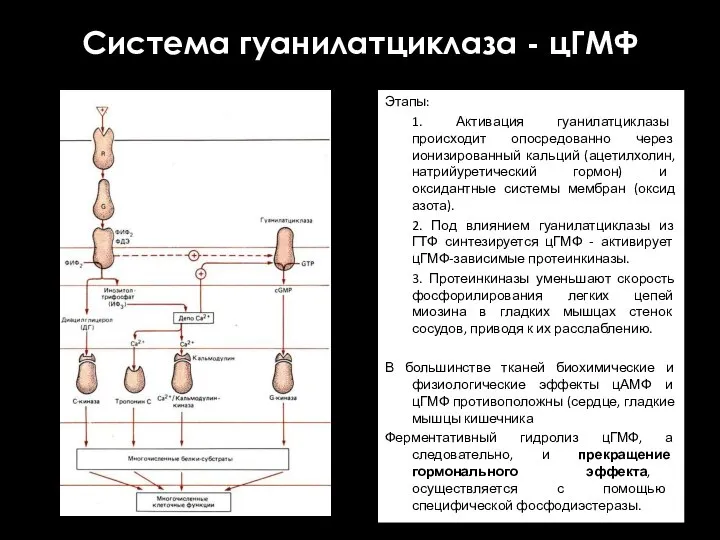
Система гуанилатциклаза - цГМФ
Этапы:
1. Активация гуанилатциклазы происходит опосредованно через ионизированный
Система гуанилатциклаза - цГМФ
Этапы:
1. Активация гуанилатциклазы происходит опосредованно через ионизированный

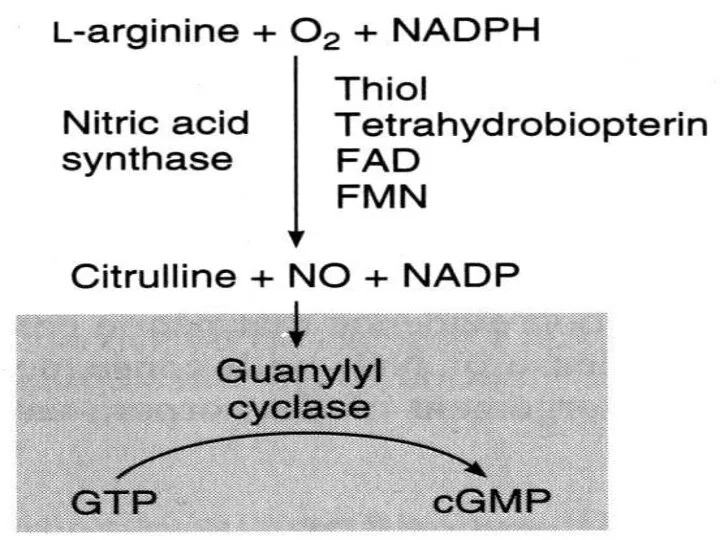
NO
как вторичный мессенджер
Образуется при окислении L-аргинина ферментом NO-синтазой (NOS), присутствующего
NO
как вторичный мессенджер
Образуется при окислении L-аргинина ферментом NO-синтазой (NOS), присутствующего

Механизм действия
В клетках-мишенях, например, эндотелиальных клетках NO взаимодействует с входящим в
Механизм действия
В клетках-мишенях, например, эндотелиальных клетках NO взаимодействует с входящим в
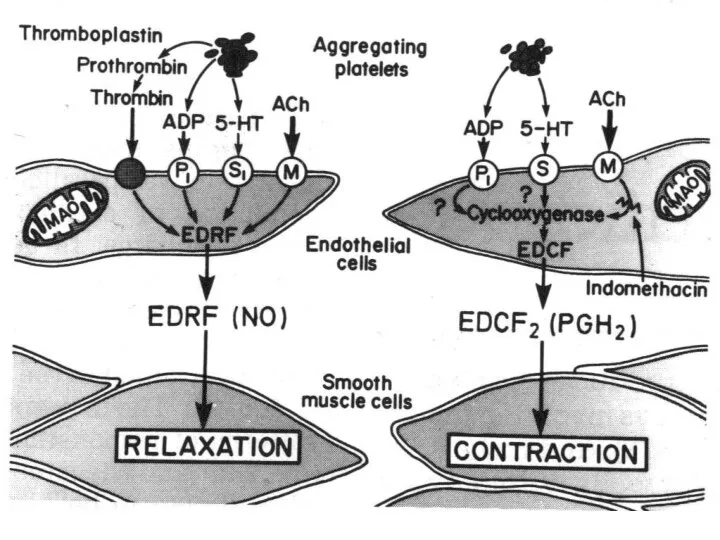

Взаимосвязи вторичных посредников
Между вторичными посредниками устанавливаются различные взаимоотношения:
равнозначное участие, когда
Взаимосвязи вторичных посредников
Между вторичными посредниками устанавливаются различные взаимоотношения:
равнозначное участие, когда
 Викторина про флору и фауну
Викторина про флору и фауну Видоизмененные корни
Видоизмененные корни Презентация на тему "Основные этапы развития растительного мира" - скачать презентации по Биологии
Презентация на тему "Основные этапы развития растительного мира" - скачать презентации по Биологии «Наука не имеет отечества , но учёный не бывает без отечества» (Луи Пастер) «Наука не имеет отечества , но учёный не бывает бе
«Наука не имеет отечества , но учёный не бывает без отечества» (Луи Пастер) «Наука не имеет отечества , но учёный не бывает бе МОУ «Владимировская средняя общеобразовательная школа» «Вода – источник жизни» Воспитатель ГПД:
МОУ «Владимировская средняя общеобразовательная школа» «Вода – источник жизни» Воспитатель ГПД:  Інфекційні захворювання Корєшкова А.В. 11-М
Інфекційні захворювання Корєшкова А.В. 11-М  Презентация на тему "Биологический аукцион «Простейшие»" - скачать презентации по Биологии
Презентация на тему "Биологический аукцион «Простейшие»" - скачать презентации по Биологии Ярмарка знаний. Цветок, плод, семя. 6 класс
Ярмарка знаний. Цветок, плод, семя. 6 класс Тип Круглые черви
Тип Круглые черви Антропогенез. Долгопяты: величие и закат. (часть 5)
Антропогенез. Долгопяты: величие и закат. (часть 5) Ретровирусы и ретровирусные векторы МГУ В.С. Прасолов
Ретровирусы и ретровирусные векторы МГУ В.С. Прасолов Профессия - флорист
Профессия - флорист Биологические ресурсы
Биологические ресурсы История генетики
История генетики Кости черепа
Кости черепа Презентация на тему Строение органов дыхания
Презентация на тему Строение органов дыхания  Презентация на тему "Папоротники, хвощи, плауны" - скачать презентации по Биологии
Презентация на тему "Папоротники, хвощи, плауны" - скачать презентации по Биологии Микропрепараты
Микропрепараты Побег и почки
Побег и почки Спланхнология бойынша түсінік. Ас қорыту, тыныс алу жүйе ағзаларының жалпы анатомиясы
Спланхнология бойынша түсінік. Ас қорыту, тыныс алу жүйе ағзаларының жалпы анатомиясы Значение воды
Значение воды Репродуктивные органы растений
Репродуктивные органы растений Биологические ритмы и их влияние на работоспособность человека
Биологические ритмы и их влияние на работоспособность человека Жалпақ құрттар типі
Жалпақ құрттар типі Микроэволюция. Видообразование
Микроэволюция. Видообразование Образцы опорных конспектов. 7 класс
Образцы опорных конспектов. 7 класс Строение клетки
Строение клетки Выделительная система ребенка
Выделительная система ребенка