- Ретровирусы и ретровирусные векторы МГУ В.С. Прасолов

Содержание
- 2. Открытие вирусов 1892 год Д.И.Ивановский – инфекционный фильтрующийся агент, вызывающий табачную мозаику. 1898 год M.Beijerinck -
- 3. Ретровирусы
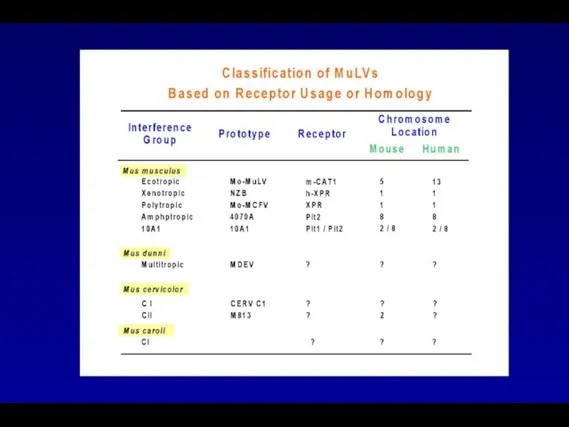
- 4. Классификация ретровирусов
- 5. morphology of a retrovirus (HIV1, a lentivirus)
- 6. Схема строения ретровирусного вириона (MuLV)
- 7. Гены простых и сложных ретровирусов
- 8. Величина геномов различных ретровирусов
- 9. Структура геномной РНК простого ретровируса
- 10. Геномная организация различных ретровирусов
- 11. Смена рамки считывания, реализуемая при трансляции РНК
- 12. Образование двух форм ретровирусной РНК
- 13. Геномная организация РНК трансформирующих вирусов
- 14. SINGLE-CELL REPRODUCTIVE CYCLE OF MuLV
- 15. Организация белков оболочки различных ретровирусов
- 16. RNA-genome of an infectious retrovirus tRNA gag pol env Splicedonor (SD) Splice- acceptor (SA) Ψ R
- 17. gag pol env R U5 PBS PP U3 R AAAA 3´ The process of reverse transcription
- 18. gag pol env PP U3 R AAAA 3´ The process of reverse transcription PBS r u5
- 19. The process of reverse transcription
- 20. The process of reverse transcription u3 r u5 pbs r u5 PBS u3 pbs pp pp
- 21. The process of reverse transcription who was copied to where ?
- 22. Схема обратной транскрипции
- 23. Genomic organization of murine retroviruses
- 24. Схема интеграции ДНК провируса в геном клетки хозяина
- 25. Схема интеграции провируса в геномную ДНК клетки-хозяина
- 26. Образование молекул РНК простого ретровируса
- 27. Transcriptional control of MoMLV integrated provirus (Moloney Murine Leukemia Virus, MoMLV)
- 28. Gene expression of simple retroviruses gag pol env AAAA
- 29. gag-pol region of simple retroviruses
- 30. env-region of retroviruses
- 31. Packaging signal of retroviruses 5´Cap AUG UAG gag pol env +1 AAAA SA
- 32. Смена рамки считывания, реализуемая при трансляции РНК
- 33. Геномная организация РНК трансформирующих вирусов
- 34. Лентивирусы – возбудители медленных инфекций
- 35. morphology of a retrovirus (HIV1, a lentivirus)
- 36. Схематическое изображение вирусной частицы HIV-1
- 37. Этапы развития СПИДа
- 38. Лентивирусы млекопитающих
- 39. Структура генома и схема сплайсинга РНК HIV-1
- 40. Структурные белки HIV-1
- 41. Неструктурные белки HIV-1
- 42. Проникновение HIV-1 в клетку
- 43. Что делает белок Tat
- 44. Зачем нужен белок Rev
- 45. Модуляция экспрессии поверхностного антигена CD4
- 46. Функции вирусных белков 1.Белок Tat увеличивает транскрипцию провирусного генома HIV-1, стимулируя элонгаторную активность РНК-полимеразы II. 2.Белок
- 47. Вирус Т-клеточного лейкоза 1 типа человека
- 48. Синтез регуляторных белков HTLV-1 направляется дважды сплайсированными РНК
- 49. Полиаденилирование и экспорт РНК HTLV-1
- 50. Белок Tax активирует транскрипцию РНК HTLV-1
- 51. Клеточные гены, активируемые вирусным белком Tax
- 60. The concept of Gene Therapy
- 61. Необходимые свойства эффективной системы переноса и экспрессии гена Высокая эффективность переноса выбранного генетического материала в клетки-мишени(in
- 62. Выбор метода переноса и экспрессии целевых генов
- 63. Выбор метода переноса и экспрессии целевых генов (продолжение)
- 64. retroviral vectors components needed in cis MFG * My Favorite Gene U 5
- 65. retroviral vectors components needed in trans
- 66. retroviral replication / packaging of defective particles
- 67. Coexpression strategies using retroviral vectors
- 68. Utilized coexpression strategies HoxB4 eGFP SD internal ribosomal entry IRES fusion protein eGFP HoxB4 SD eGFP
- 69. Calculation of retroviral titers („GFP-Transfer Units“) -> % GFP positive cells x original cell number x
- 70. Titers of retroviral expression vectors
- 71. vector production transduction splicing / nuclear export reverse transcription packaging
- 73. Excision of genes via direct application of Cre-recombinase reporter cell line
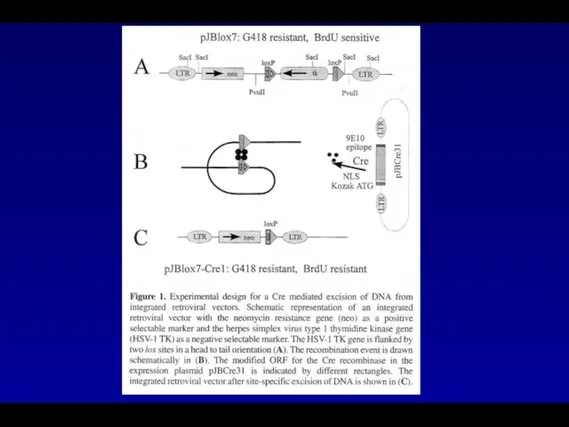
- 76. В процессе работы были получены ретровирусные векторы содержащие в своем составе в качестве маркерного гена ген
- 77. (А) и трансфицированные (В) клетки 293 при микроскопии в видимом свете. Исходные (Б) и трансфицированные (Г)
- 78. Кинетика продукции ретровирусного вектора RV-GFP/07-07 упаковывающими клетками HEK293gp. Сбор вируса осуществлялся через 24 часа после трансфекции
- 79. Кинетика продукции ретровирусного вектора RV-GFP/07-07 упаковывающими клетками HEK293gp. Для переноса использовали 100 мкл культуральной среды трансфицированных
- 80. Создание клеток продуцентов (перевиваемых фибробластов мыши) секретируемого гормона роста человека с помощью ретровирусного вектора RV-GFP/ 07-07.
- 81. Продукция гормона роста человека перевиваемыми клетками мыши линии SC1, трансдуцированных ретровирусным вектором R-hGH-IRES-eGFP Электрофоретической разделение в
- 82. Физическая карта модульного лентивирусного вектора pLPL-mCMV-H4puro Индукция синтеза бета-галактозидазы в перевиваемых клетках рака лёгкого человека H1299,
- 83. Схема конструкции для экспрессии зеленого флуоресцентного белка с локализацией в эндоплазматическом ретикулуме, на основе нового лентивирусного
- 84. Испытание лентивирусного вектора pLCMV для стабильной экспрессии зеленого флуоресцентного белка, направляемого в различные структуры клетки
- 85. Retroviral vectors most commonly used gene transfer system in gene therapy genome integration ensures stable long-term
- 86. Possible mechanisms of (retroviral) insertional mutagenesis gene X 5‘LTR 3‘LTR 1 2 3 4 5 7
- 87. ... theoretical risk assessment Starting cell number: 108 Target cell number: 1x109 (1x107 per kg) Transduction
- 88. Схематическое изображение структурно-функциональных изменений ретровирусного вектора, приводящих к делециям в промоторно-энхансерных областях обоих LTR а) -
- 89. Env protein Transmembrane Domain TM Surface Protein SU Cell Surface Protein ( Receptor) Binding Trimer-Formation MuLV
- 90. Neuropathology of DDD mice infected with Mo-AmphoV alone (A–C), or coinfected with Mo-AmphoV and F-MuLV (D–F).
- 94. M813 expression slightly interferences with ecotropic MuLV infection but not with other MuLVs
- 95. А) PA 317 cells incubated with M813 for 4h B) Uninfected PA317 Syncytia formation induced by
- 96. Selection with Geneticin Target Cells TE671 Infection Packaging the Vector into VSV-G Pseudotypes TE671i mCAT1 Establishing
- 97. Marker-rescue assay (A) Cell with integrated provirus of replication defective virus with marker gene Infection by
- 98. Marker-rescue assay (B)
- 99. M813 Does Not Use mCAT1 for Infection SC-1 TE671-neo TE671-mCAT 10A1 Mo-MuLV M813 79% 55% 56%
- 101. M813 SU Sequences Dictate Receptor Usage MoMuLV MoM813 M813
- 102. Hamster Cells Murine Cells Irradiation Genome Fragmententation Fusion + Selection 96 Hybrid Clones Per clone: 31%
- 103. promising candidate gene on Chr. 16: ► Scl5A3 : multiple membrane spanning surface protein encoding SMIT1:
- 104. Expression of mSMIT1 in Human Cells Imparts Susceptibility to M813 Infection TE671-mSMIT1 52% M1 SC1 M1
- 105. M813 Induces Syncytia Formation in Cells Expressing Its Receptor Te671 + neo Te671 + mSmit1
- 106. M813 belongs to a unique receptor interference group M813 is highly fusogenic in vitro and in
- 107. Acknowledgments Dmitry Ivanov, Pavel Spirin, Tamara Semenova Engelhardt Institute of Molecular Biology Moscow, Russia Sibyll Hein
- 108. MuLV EXPRESSION IN SC-1 CELLS DETERMINES THE FUSOGENIC PROPERTIES OF SC-1 CELLS
- 109. М813 INDUCES SYNCYTIA FORMATION MORE EFFECTIVE THAN OTHER MURINE LEUKEMIA RETROVIRUSES
- 110. Fusion index (FI) = (N - S)/T N – number of nuclei in syncytium S –
- 111. Gene Therapy trials - overview http://www.wiley.co.uk/wileychi/genmed/clinical/
- 112. Gene Therapy trials - overview http://www.wiley.co.uk/wileychi/genmed/clinical/
- 113. Gene Therapy trials - overview http://www.wiley.co.uk/wileychi/genmed/clinical/
- 114. Инсерционный мутагенез Встраивание провируса может приводить к трансформации клетки
- 115. Открытие вирусов 1892 год Д.И.Ивановский – инфекционный фильтрующийся агент, вызывающий табачную мозаику. 1898 год M.Beijerinck -
- 117. Скачать презентацию

Открытие вирусов
1892 год
Д.И.Ивановский – инфекционный фильтрующийся агент, вызывающий табачную мозаику.
1898
Открытие вирусов
1892 год
Д.И.Ивановский – инфекционный фильтрующийся агент, вызывающий табачную мозаику.
1898

Ретровирусы
Ретровирусы

Классификация ретровирусов
Классификация ретровирусов
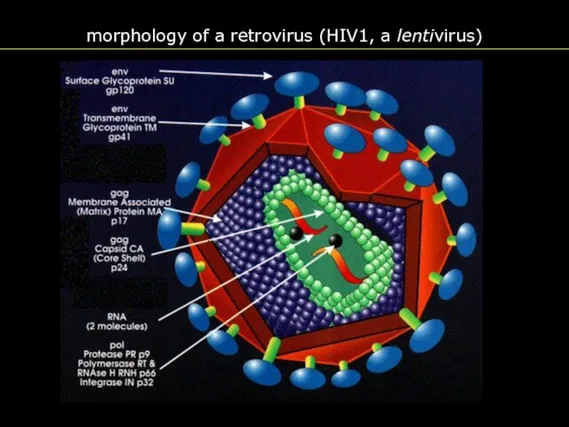
morphology of a retrovirus (HIV1, a lentivirus)
morphology of a retrovirus (HIV1, a lentivirus)
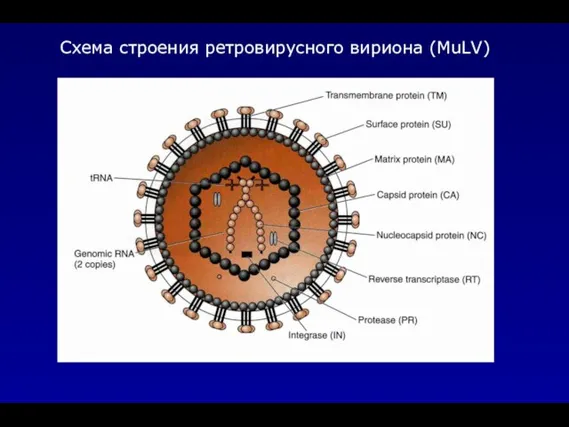
Схема строения ретровирусного вириона (MuLV)
Схема строения ретровирусного вириона (MuLV)
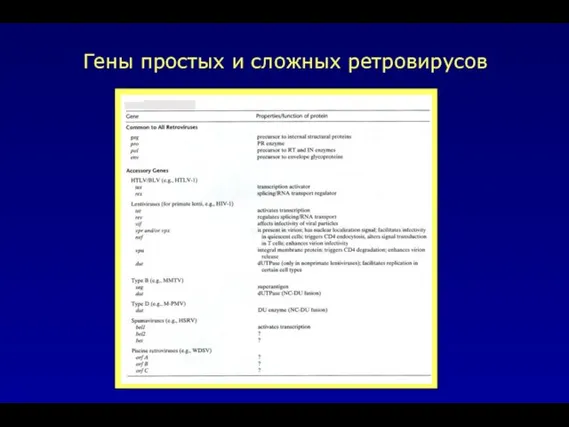
Гены простых и сложных ретровирусов
Гены простых и сложных ретровирусов
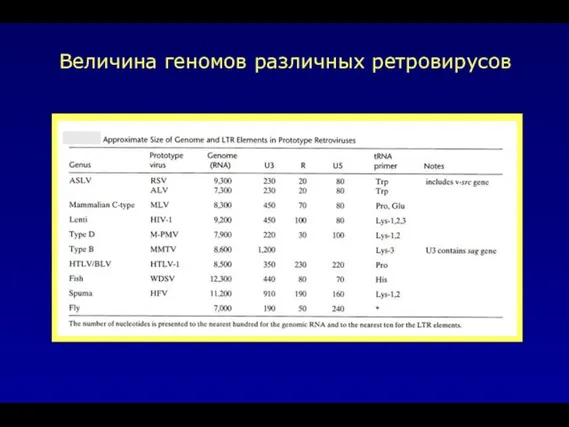
Величина геномов различных ретровирусов
Величина геномов различных ретровирусов

Структура геномной РНК простого ретровируса
Структура геномной РНК простого ретровируса

Геномная организация различных ретровирусов
Геномная организация различных ретровирусов
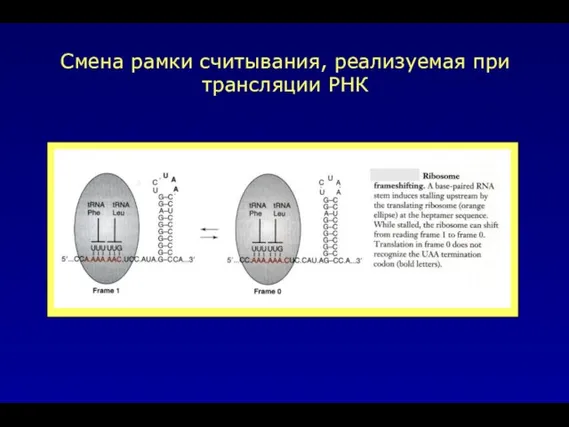
Смена рамки считывания, реализуемая при трансляции РНК
Смена рамки считывания, реализуемая при трансляции РНК
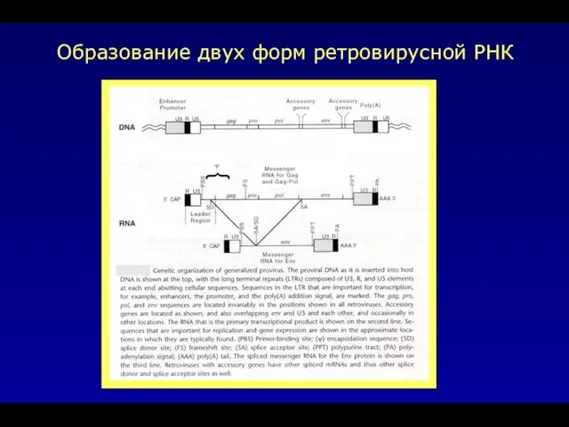
Образование двух форм ретровирусной РНК
Образование двух форм ретровирусной РНК
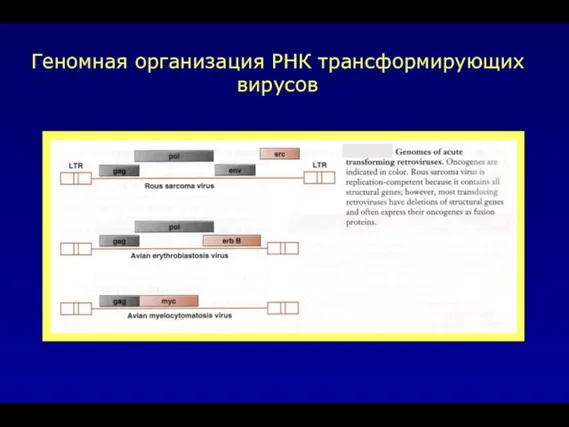
Геномная организация РНК трансформирующих вирусов
Геномная организация РНК трансформирующих вирусов
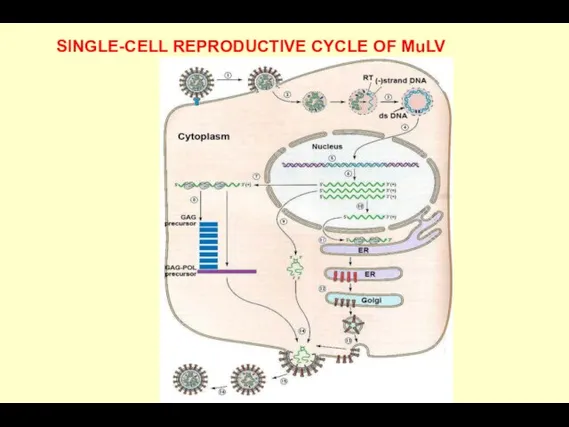
SINGLE-CELL REPRODUCTIVE CYCLE OF MuLV
SINGLE-CELL REPRODUCTIVE CYCLE OF MuLV

Организация белков оболочки различных ретровирусов
Организация белков оболочки различных ретровирусов
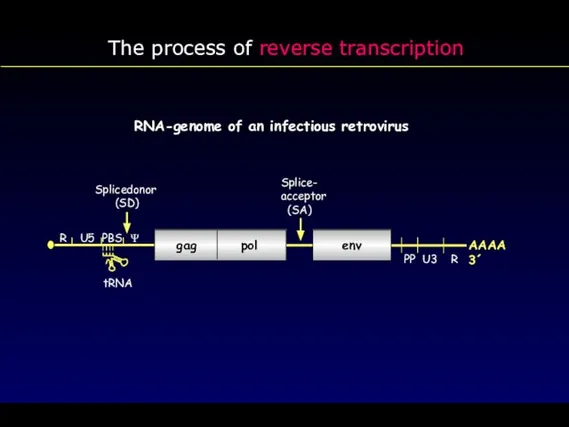
RNA-genome of an infectious retrovirus
tRNA
gag pol
env
Splicedonor
(SD)
Splice-
acceptor
(SA)
Ψ
R
RNA-genome of an infectious retrovirus
tRNA
gag pol
env
Splicedonor
(SD)
Splice-
acceptor
(SA)
Ψ
R
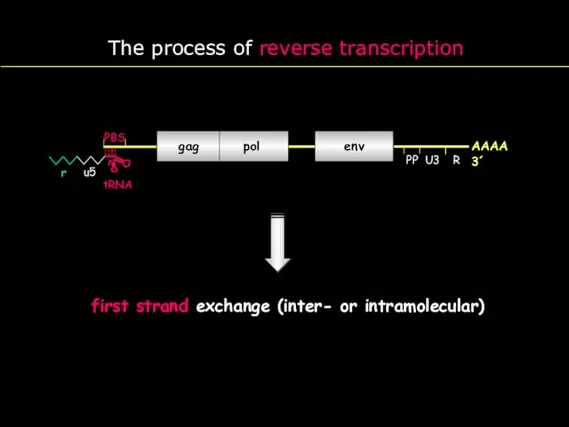
gag pol
env
R U5 PBS
PP U3 R
AAAA 3´
The process
gag pol
env
R U5 PBS
PP U3 R
AAAA 3´
The process
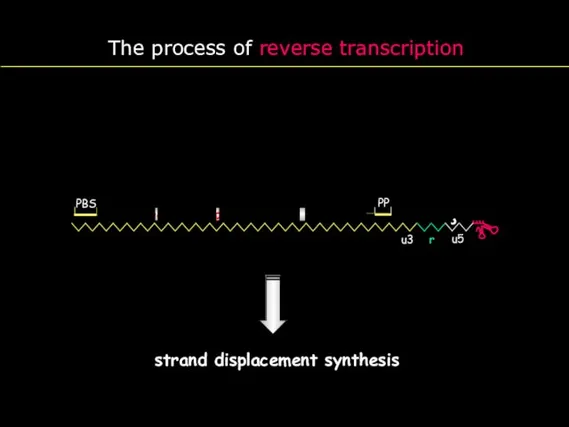
gag pol
env
PP U3 R
AAAA 3´
The process of reverse
gag pol
env
PP U3 R
AAAA 3´
The process of reverse
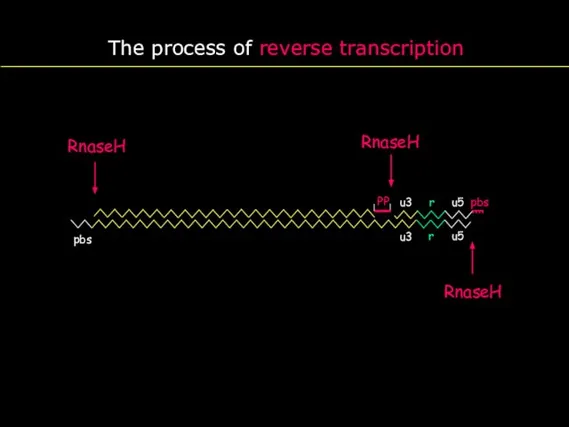
The process of reverse transcription
The process of reverse transcription
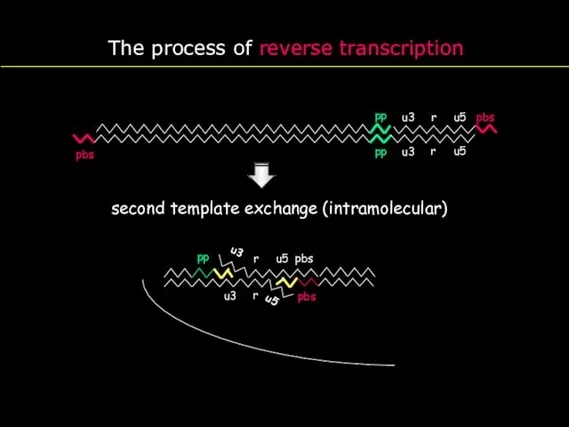
The process of reverse transcription
u3 r u5 pbs
r
u5
PBS
u3
pbs
pp
pp
The process of reverse transcription
u3 r u5 pbs
r
u5
PBS
u3
pbs
pp
pp
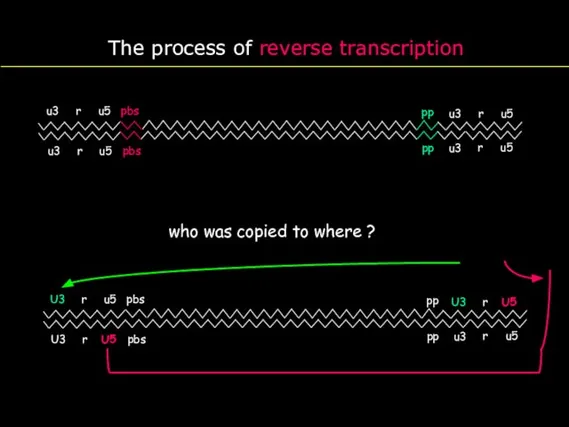
The process of reverse transcription
who was copied to where ?
The process of reverse transcription
who was copied to where ?

Схема обратной транскрипции
Схема обратной транскрипции

Genomic organization of murine retroviruses
Genomic organization of murine retroviruses
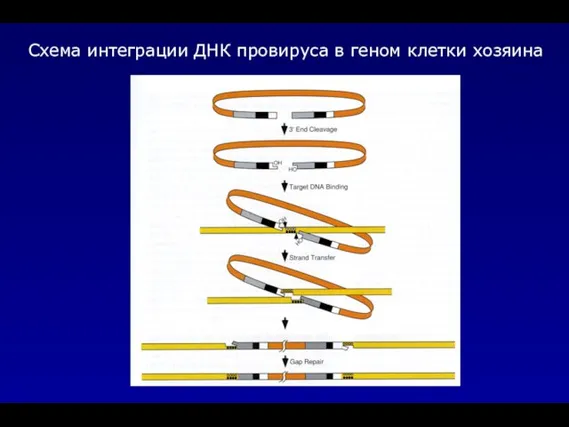
Схема интеграции ДНК провируса в геном клетки хозяина
Схема интеграции ДНК провируса в геном клетки хозяина

Схема интеграции провируса в геномную ДНК клетки-хозяина
Схема интеграции провируса в геномную ДНК клетки-хозяина

Образование молекул РНК простого ретровируса
Образование молекул РНК простого ретровируса
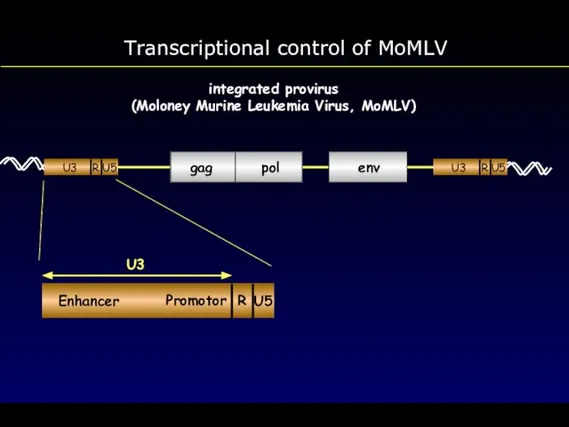
Transcriptional control of MoMLV
integrated provirus
(Moloney Murine Leukemia Virus, MoMLV)
Transcriptional control of MoMLV
integrated provirus
(Moloney Murine Leukemia Virus, MoMLV)
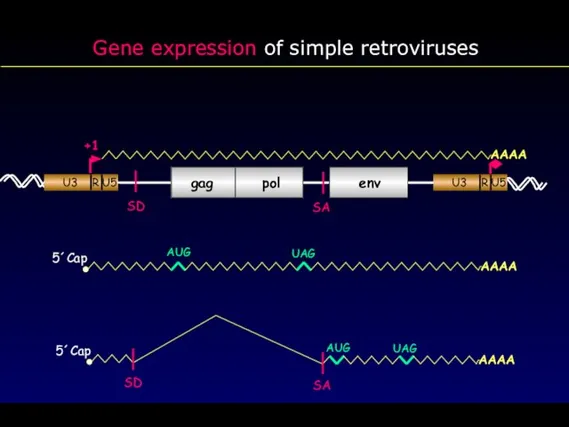
Gene expression of simple retroviruses
gag pol
env
AAAA
Gene expression of simple retroviruses
gag pol
env
AAAA
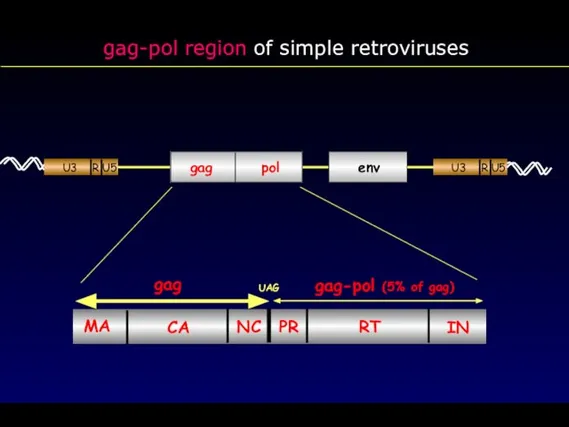
gag-pol region of simple retroviruses
gag-pol region of simple retroviruses
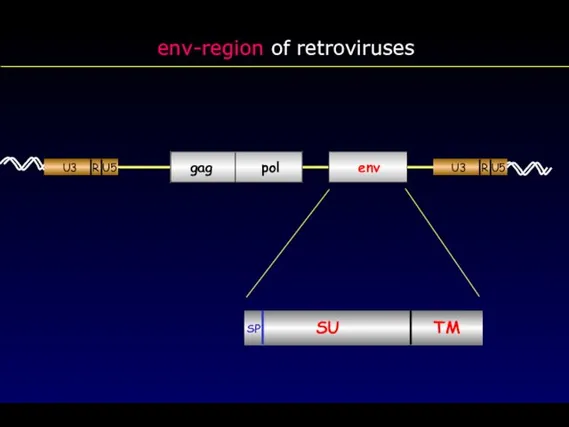
env-region of retroviruses
env-region of retroviruses
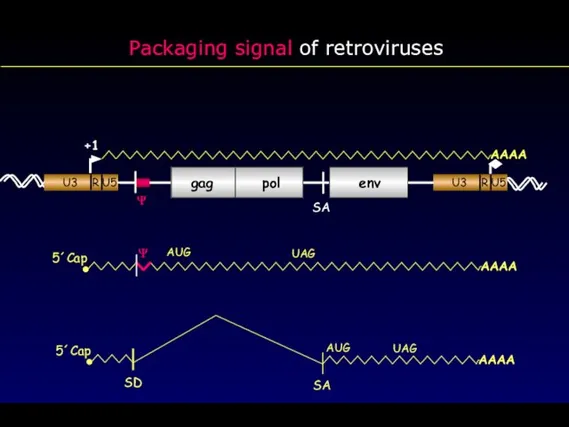
Packaging signal of retroviruses
5´Cap
AUG
UAG
gag pol
env
+1
AAAA
SA
Packaging signal of retroviruses
5´Cap
AUG
UAG
gag pol
env
+1
AAAA
SA
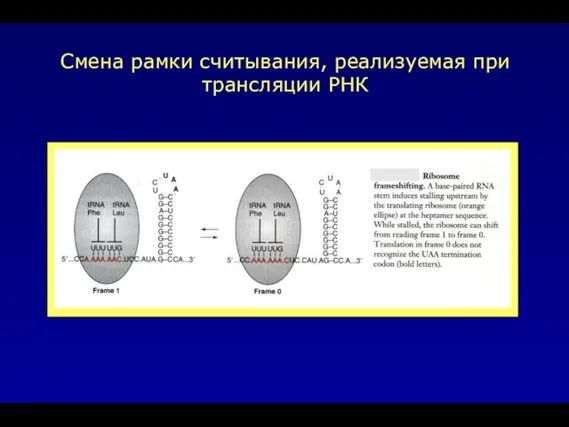
Смена рамки считывания, реализуемая при трансляции РНК
Смена рамки считывания, реализуемая при трансляции РНК
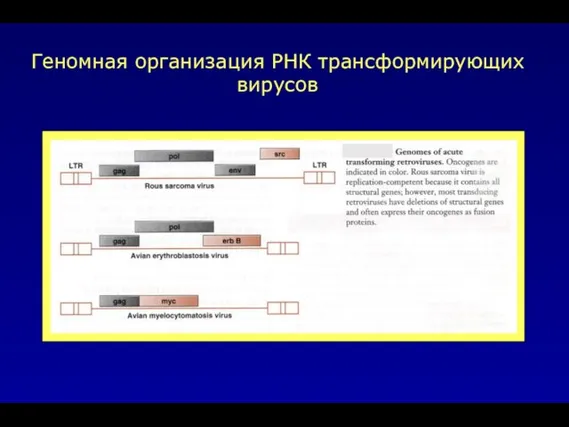
Геномная организация РНК трансформирующих вирусов
Геномная организация РНК трансформирующих вирусов

Лентивирусы – возбудители медленных инфекций
Лентивирусы – возбудители медленных инфекций
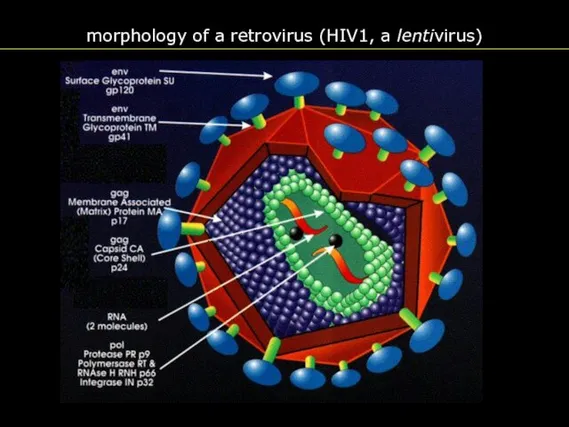
morphology of a retrovirus (HIV1, a lentivirus)
morphology of a retrovirus (HIV1, a lentivirus)
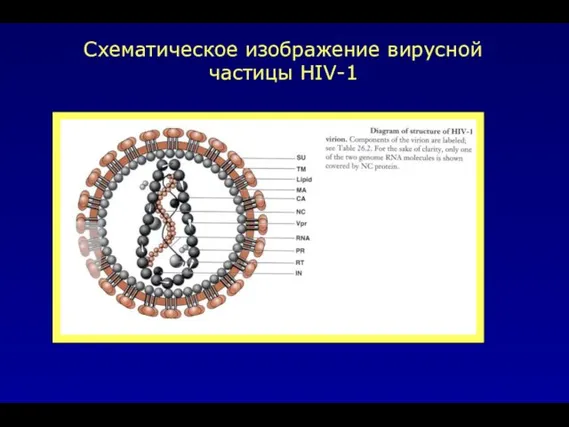
Схематическое изображение вирусной частицы HIV-1
Схематическое изображение вирусной частицы HIV-1
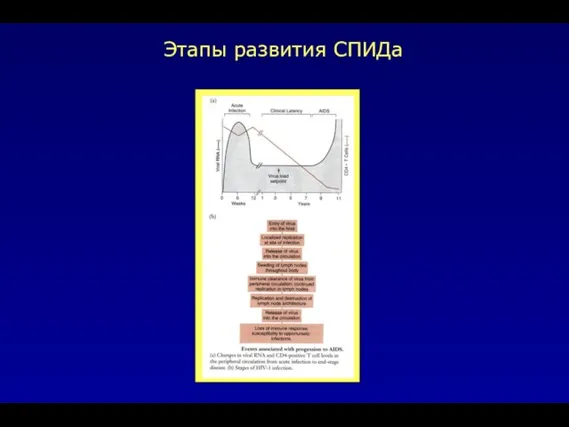
Этапы развития СПИДа
Этапы развития СПИДа

Лентивирусы млекопитающих
Лентивирусы млекопитающих
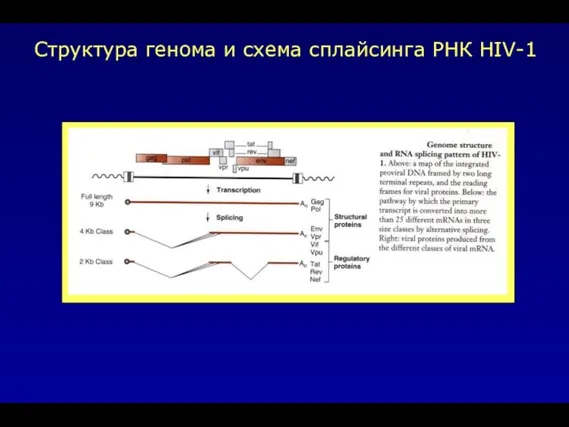
Структура генома и схема сплайсинга РНК HIV-1
Структура генома и схема сплайсинга РНК HIV-1

Структурные белки HIV-1
Структурные белки HIV-1

Неструктурные белки HIV-1
Неструктурные белки HIV-1
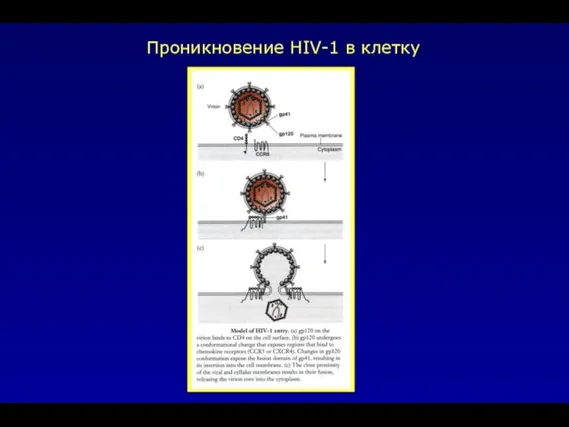
Проникновение HIV-1 в клетку
Проникновение HIV-1 в клетку
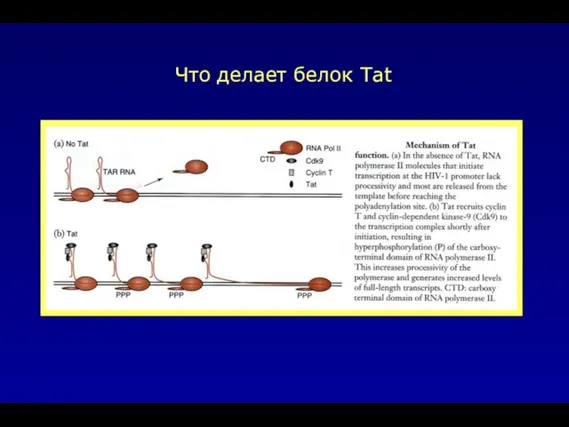
Что делает белок Tat
Что делает белок Tat

Зачем нужен белок Rev
Зачем нужен белок Rev
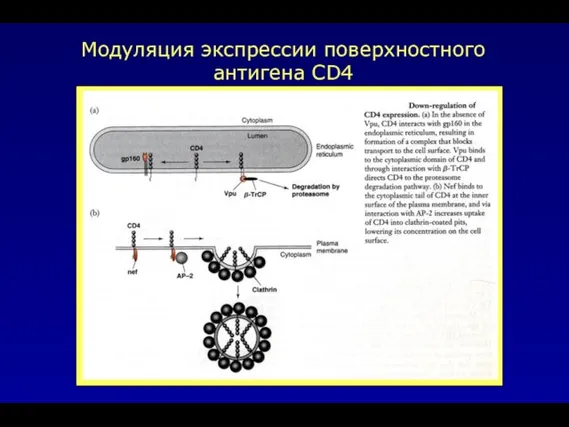
Модуляция экспрессии поверхностного антигена CD4
Модуляция экспрессии поверхностного антигена CD4

Функции вирусных белков
1.Белок Tat увеличивает транскрипцию провирусного генома HIV-1,
стимулируя элонгаторную активность
Функции вирусных белков
1.Белок Tat увеличивает транскрипцию провирусного генома HIV-1,
стимулируя элонгаторную активность

Вирус Т-клеточного лейкоза 1 типа человека
Вирус Т-клеточного лейкоза 1 типа человека
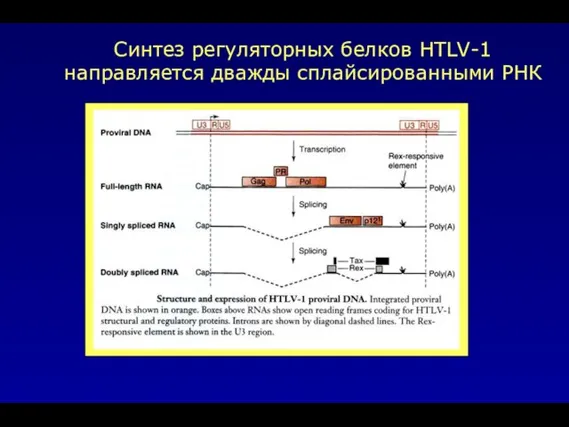
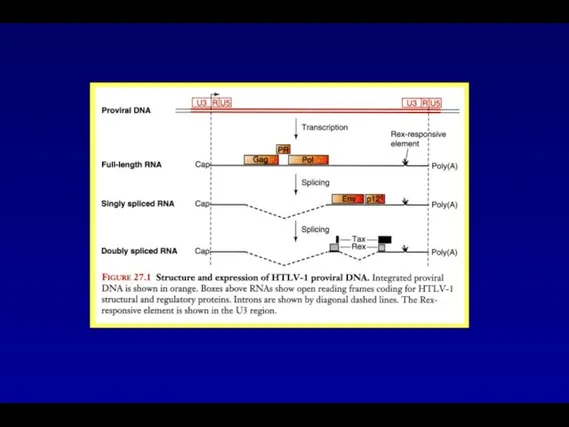
Синтез регуляторных белков HTLV-1 направляется дважды сплайсированными РНК
Синтез регуляторных белков HTLV-1 направляется дважды сплайсированными РНК
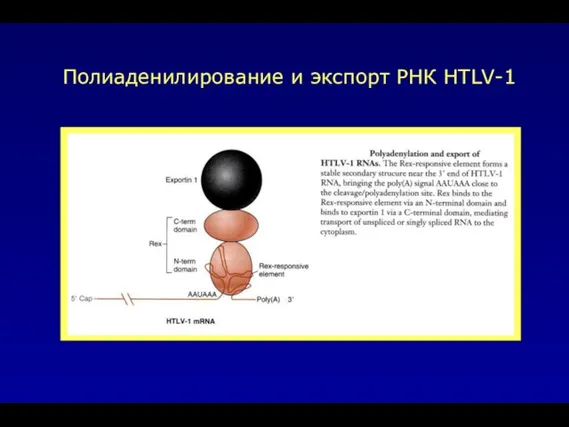
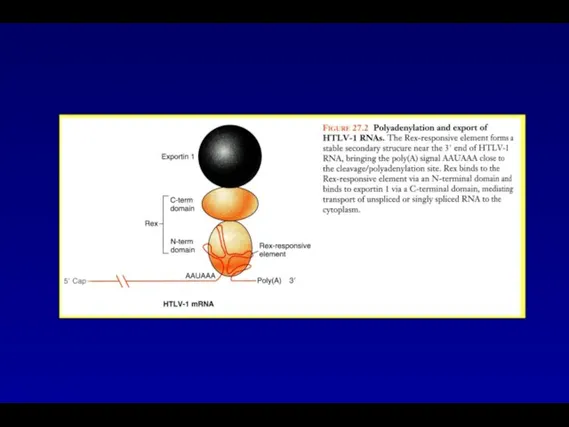
Полиаденилирование и экспорт РНК HTLV-1
Полиаденилирование и экспорт РНК HTLV-1

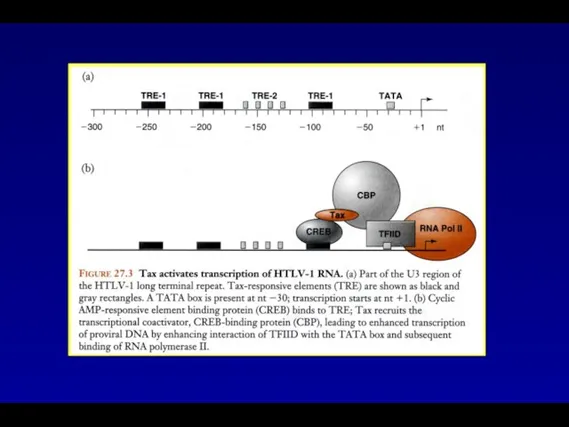
Белок Tax активирует транскрипцию РНК HTLV-1
Белок Tax активирует транскрипцию РНК HTLV-1

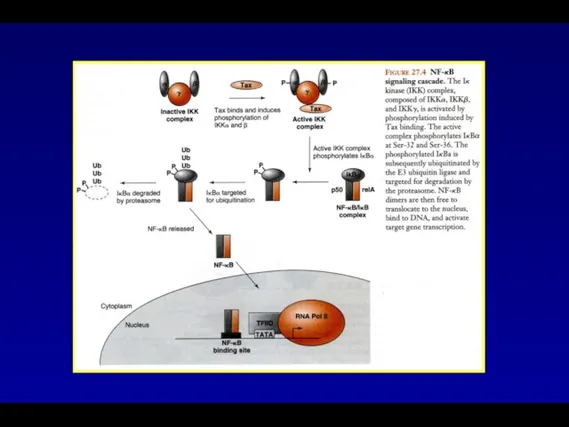
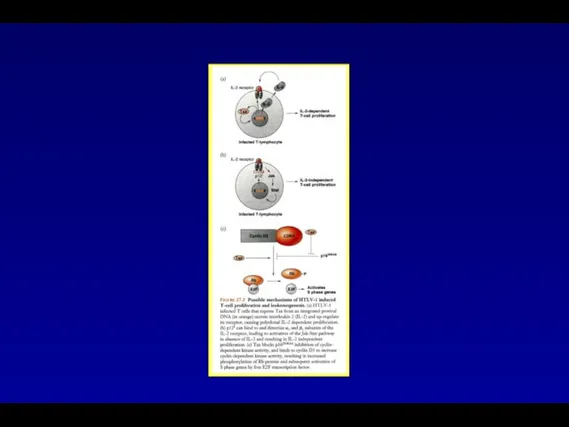
Клеточные гены, активируемые вирусным белком Tax
Клеточные гены, активируемые вирусным белком Tax



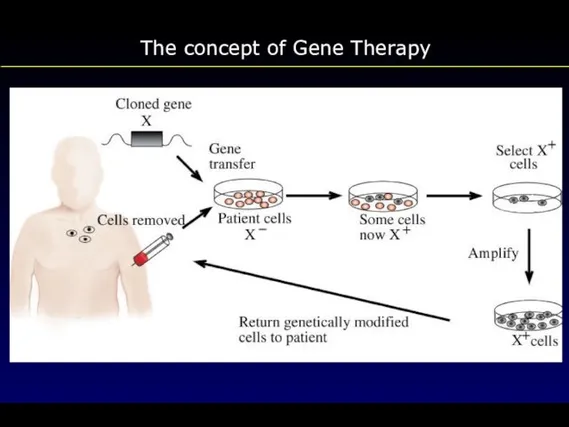
The concept of Gene Therapy
The concept of Gene Therapy

Необходимые свойства эффективной системы переноса и экспрессии гена
Высокая эффективность переноса выбранного
Необходимые свойства эффективной системы переноса и экспрессии гена
Высокая эффективность переноса выбранного

Выбор метода переноса и экспрессии целевых генов
Выбор метода переноса и экспрессии целевых генов

Выбор метода переноса и экспрессии целевых генов (продолжение)
Выбор метода переноса и экспрессии целевых генов (продолжение)
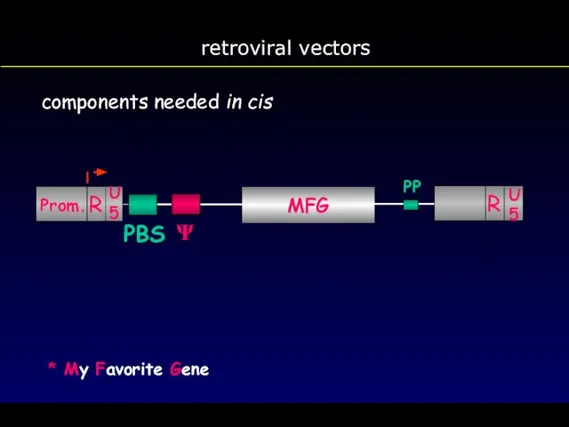
retroviral vectors
components needed in cis
MFG
* My Favorite Gene
U
5
retroviral vectors
components needed in cis
MFG
* My Favorite Gene
U
5
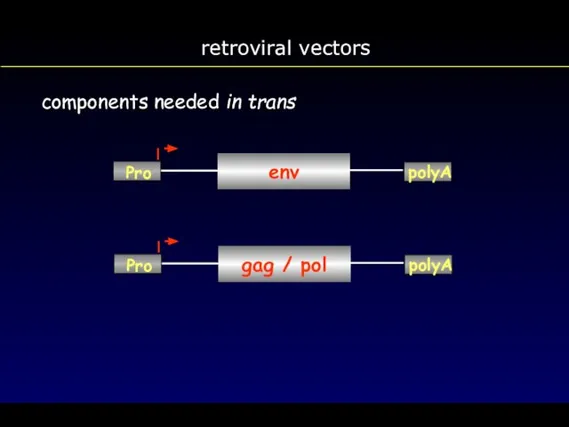
retroviral vectors
components needed in trans
retroviral vectors
components needed in trans
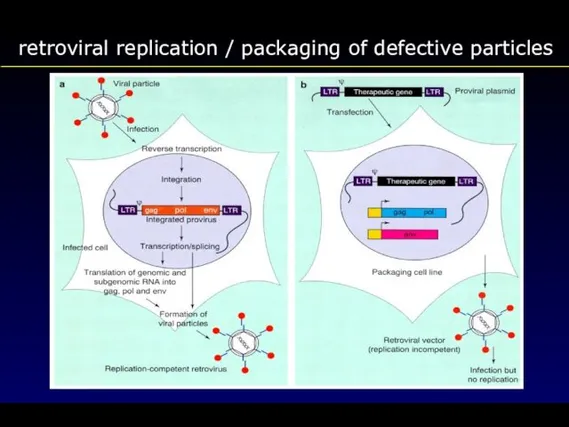
retroviral replication / packaging of defective particles
retroviral replication / packaging of defective particles
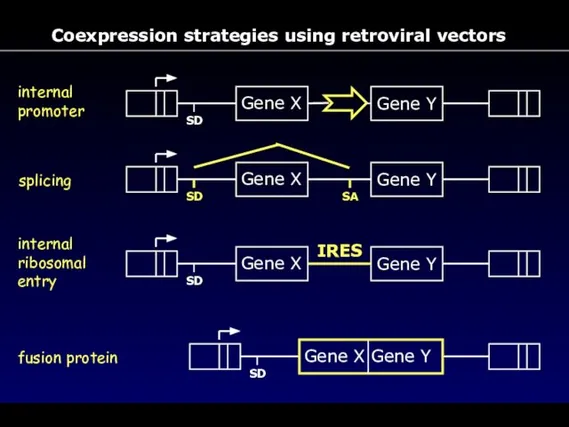
Coexpression strategies using retroviral vectors
Coexpression strategies using retroviral vectors
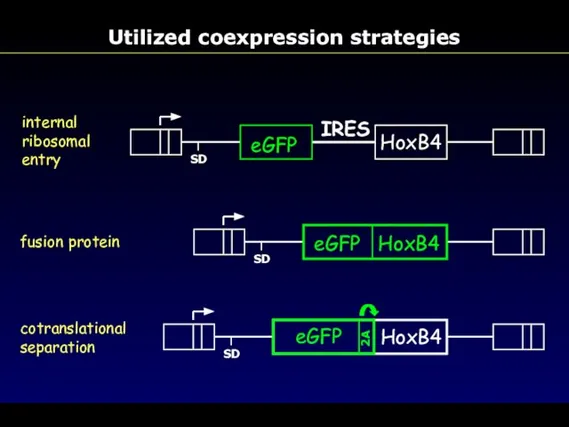
Utilized coexpression strategies
HoxB4
eGFP
SD
internal
ribosomal
entry
IRES
fusion protein
eGFP HoxB4
SD
eGFP
SD
HoxB4
2A
cotranslational
separation
Utilized coexpression strategies
HoxB4
eGFP
SD
internal
ribosomal
entry
IRES
fusion protein
eGFP HoxB4
SD
eGFP
SD
HoxB4
2A
cotranslational
separation
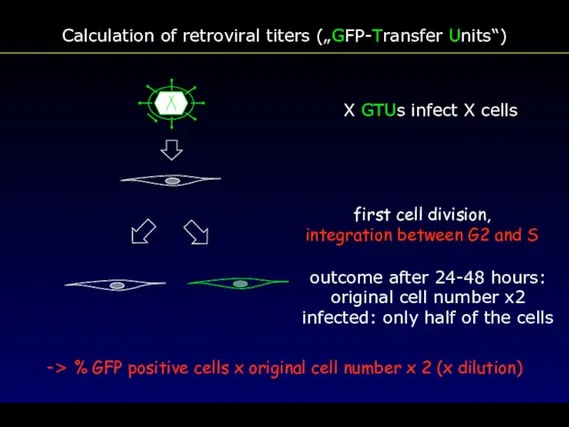
Calculation of retroviral titers („GFP-Transfer Units“)
-> % GFP positive cells x
Calculation of retroviral titers („GFP-Transfer Units“)
-> % GFP positive cells x
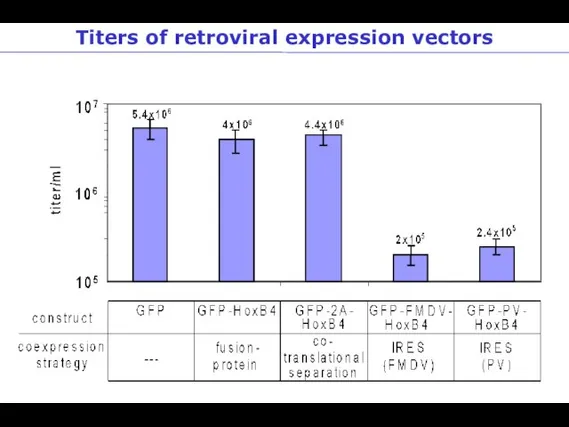
Titers of retroviral expression vectors
Titers of retroviral expression vectors
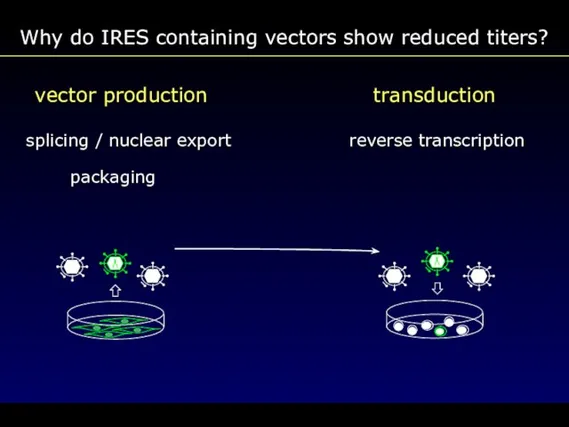
vector production
transduction
splicing / nuclear export
reverse transcription
packaging
vector production
transduction
splicing / nuclear export
reverse transcription
packaging
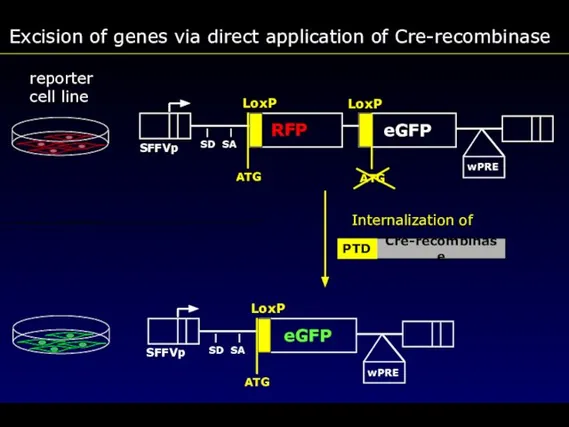
Excision of genes via direct application of Cre-recombinase
reporter
cell line
Excision of genes via direct application of Cre-recombinase
reporter
cell line
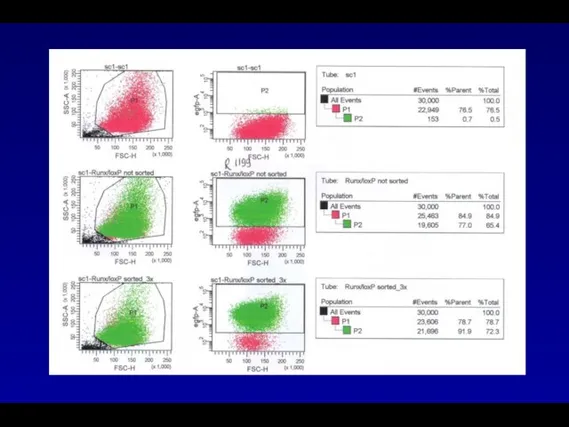
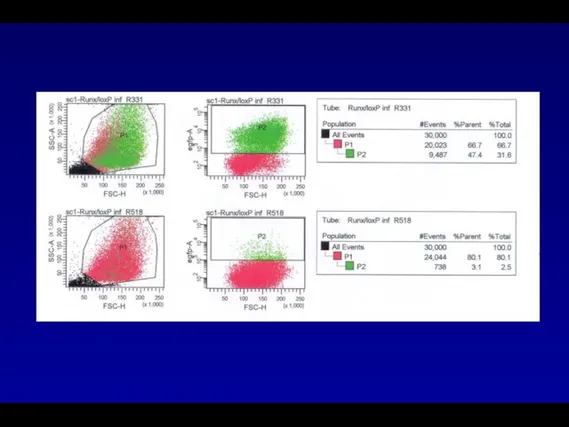

В процессе работы были получены ретровирусные векторы содержащие в своем составе
В процессе работы были получены ретровирусные векторы содержащие в своем составе
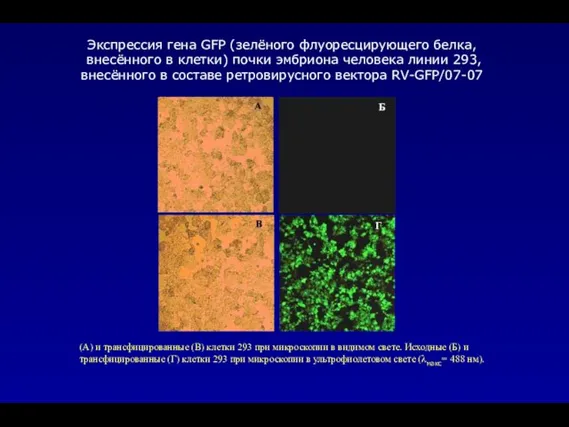
(А) и трансфицированные (В) клетки 293 при микроскопии в видимом свете.
(А) и трансфицированные (В) клетки 293 при микроскопии в видимом свете.
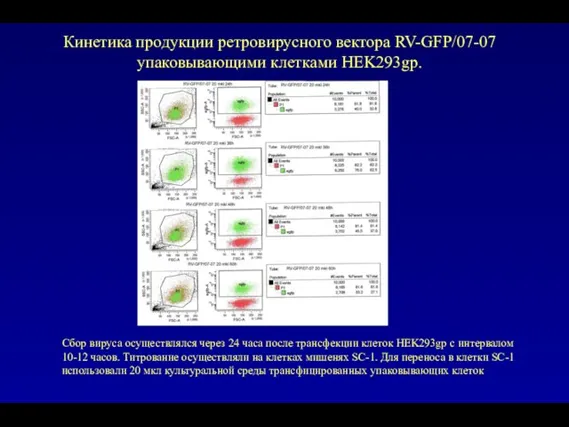
Кинетика продукции ретровирусного вектора RV-GFP/07-07 упаковывающими клетками HEK293gp.
Сбор вируса осуществлялся
Кинетика продукции ретровирусного вектора RV-GFP/07-07 упаковывающими клетками HEK293gp.
Сбор вируса осуществлялся

Кинетика продукции ретровирусного вектора RV-GFP/07-07 упаковывающими клетками HEK293gp.
Для переноса использовали
Кинетика продукции ретровирусного вектора RV-GFP/07-07 упаковывающими клетками HEK293gp.
Для переноса использовали

Создание клеток продуцентов (перевиваемых фибробластов мыши) секретируемого гормона роста человека с
Создание клеток продуцентов (перевиваемых фибробластов мыши) секретируемого гормона роста человека с

Продукция гормона роста человека перевиваемыми клетками мыши линии SC1, трансдуцированных ретровирусным
Продукция гормона роста человека перевиваемыми клетками мыши линии SC1, трансдуцированных ретровирусным
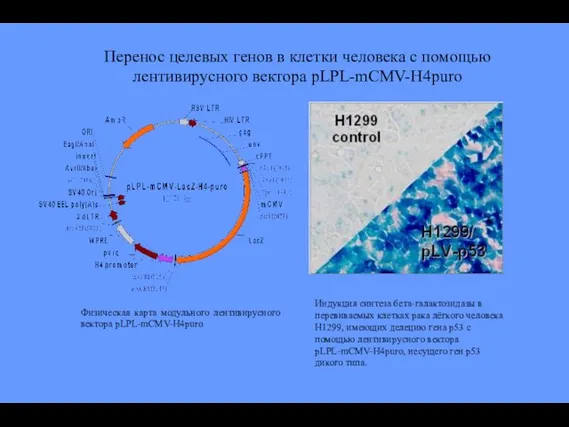
Физическая карта модульного лентивирусного вектора pLPL-mCMV-H4puro
Индукция синтеза бета-галактозидазы в перевиваемых клетках
Физическая карта модульного лентивирусного вектора pLPL-mCMV-H4puro
Индукция синтеза бета-галактозидазы в перевиваемых клетках

Схема конструкции для экспрессии зеленого флуоресцентного белка с локализацией в эндоплазматическом
Схема конструкции для экспрессии зеленого флуоресцентного белка с локализацией в эндоплазматическом
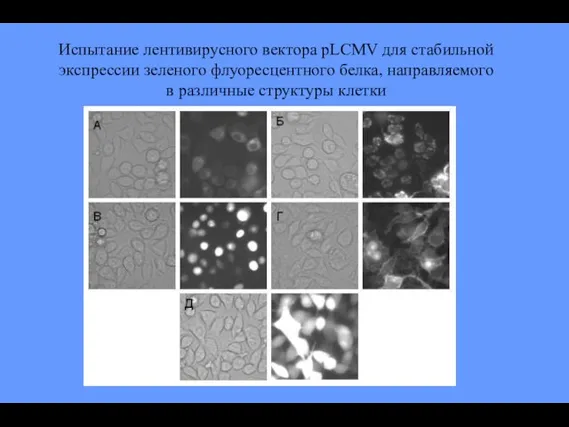
Испытание лентивирусного вектора pLCMV для стабильной экспрессии зеленого флуоресцентного белка, направляемого
Испытание лентивирусного вектора pLCMV для стабильной экспрессии зеленого флуоресцентного белка, направляемого

Retroviral vectors
most commonly used gene transfer system in gene therapy
genome integration
Retroviral vectors
most commonly used gene transfer system in gene therapy
genome integration
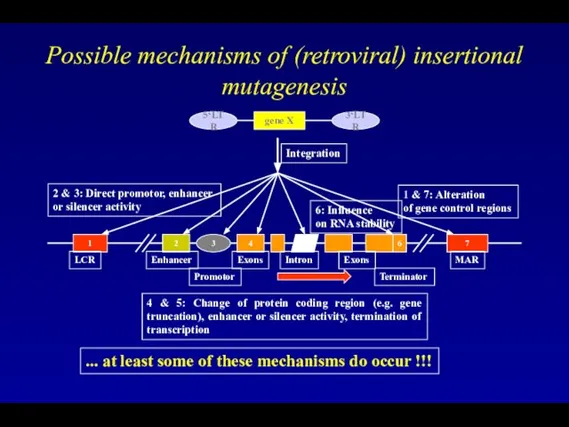
Possible mechanisms of (retroviral) insertional mutagenesis
gene X
5‘LTR
3‘LTR
1
2
3
4
5
7
LCR
Enhancer
Promotor
Terminator
Exons
Intron
Exons
MAR
Integration
6
4 & 5: Change of
Possible mechanisms of (retroviral) insertional mutagenesis
gene X
5‘LTR
3‘LTR
1
2
3
4
5
7
LCR
Enhancer
Promotor
Terminator
Exons
Intron
Exons
MAR
Integration
6
4 & 5: Change of

... theoretical risk assessment
Starting cell number: 108
Target cell number: 1x109
... theoretical risk assessment
Starting cell number: 108
Target cell number: 1x109

Схематическое изображение структурно-функциональных изменений ретровирусного вектора, приводящих к делециям в промоторно-энхансерных
Схематическое изображение структурно-функциональных изменений ретровирусного вектора, приводящих к делециям в промоторно-энхансерных
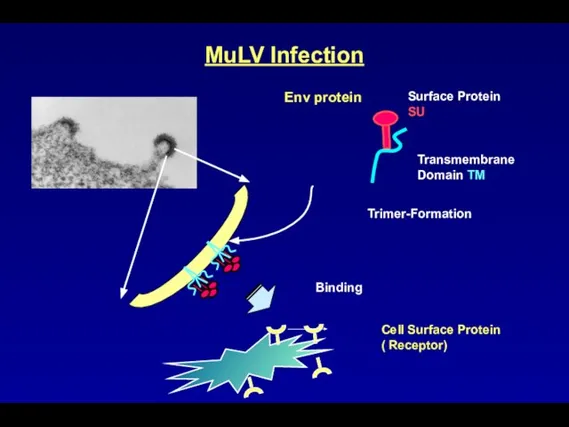
Env protein
Transmembrane Domain TM
Surface Protein
SU
Cell Surface Protein
( Receptor)
Binding
Trimer-Formation
MuLV Infection
Env protein
Transmembrane Domain TM
Surface Protein
SU
Cell Surface Protein
( Receptor)
Binding
Trimer-Formation
MuLV Infection
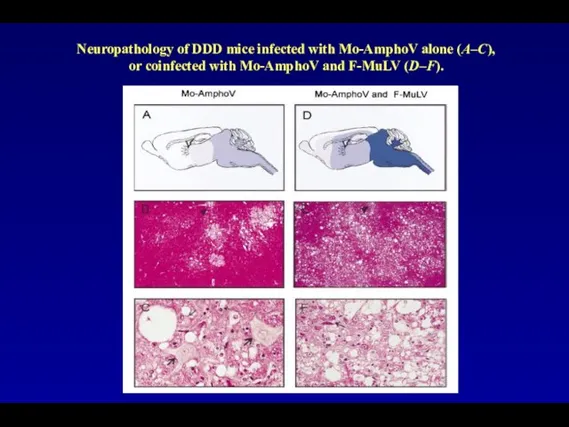
Neuropathology of DDD mice infected with Mo-AmphoV alone (A–C),
or coinfected
Neuropathology of DDD mice infected with Mo-AmphoV alone (A–C),
or coinfected
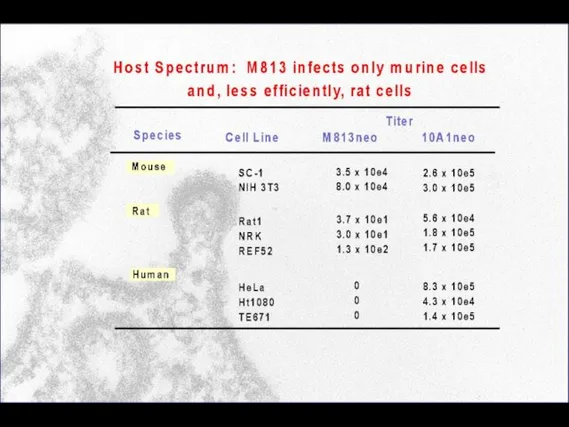


M813 expression slightly interferences with
ecotropic MuLV infection but not with
M813 expression slightly interferences with
ecotropic MuLV infection but not with
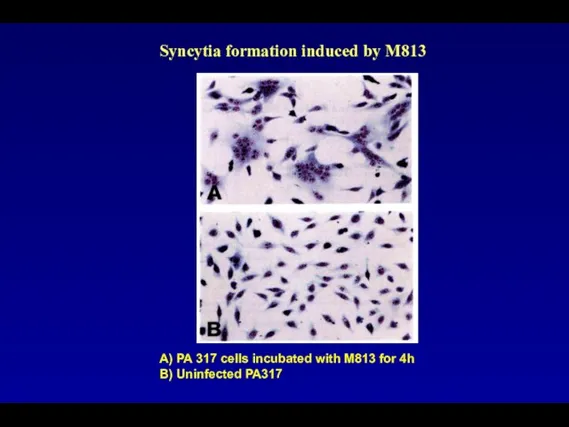
А) PA 317 cells incubated with M813 for 4h
B) Uninfected
А) PA 317 cells incubated with M813 for 4h
B) Uninfected
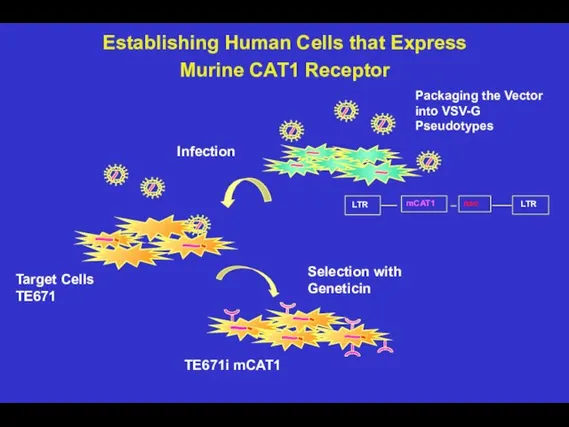
Selection with Geneticin
Target Cells
TE671
Infection
Packaging the Vector into VSV-G Pseudotypes
TE671i mCAT1
Establishing Human
Selection with Geneticin
Target Cells
TE671
Infection
Packaging the Vector into VSV-G Pseudotypes
TE671i mCAT1
Establishing Human
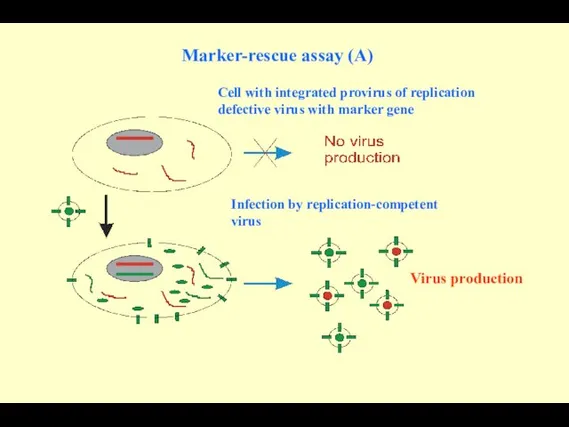
Marker-rescue assay (A)
Cell with integrated provirus of replication
defective virus with marker
Marker-rescue assay (A)
Cell with integrated provirus of replication
defective virus with marker
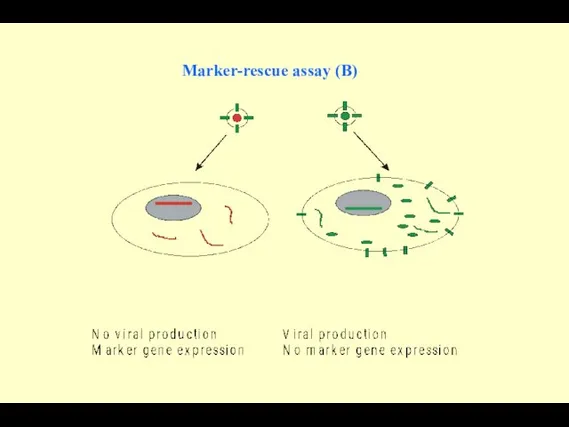
Marker-rescue assay (B)
Marker-rescue assay (B)
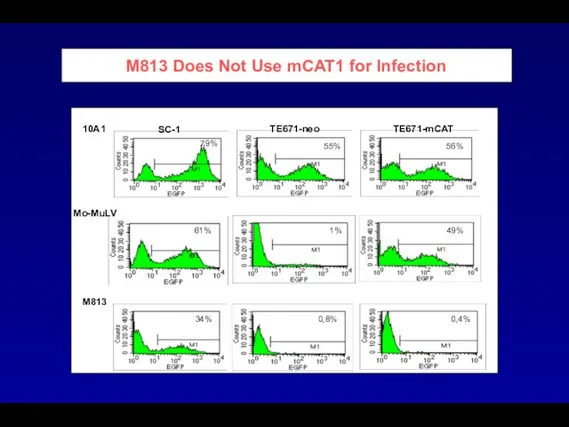
M813 Does Not Use mCAT1 for Infection
SC-1
TE671-neo
TE671-mCAT
10A1
Mo-MuLV
M813
79%
55%
56%
49%
1%
61%
0,4%
0,8%
34%
M813 Does Not Use mCAT1 for Infection
SC-1
TE671-neo
TE671-mCAT
10A1
Mo-MuLV
M813
79%
55%
56%
49%
1%
61%
0,4%
0,8%
34%
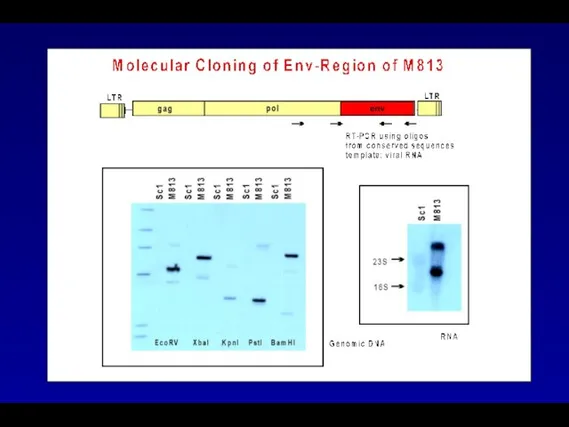

M813 SU Sequences Dictate Receptor Usage
MoMuLV
MoM813
M813
M813 SU Sequences Dictate Receptor Usage
MoMuLV
MoM813
M813
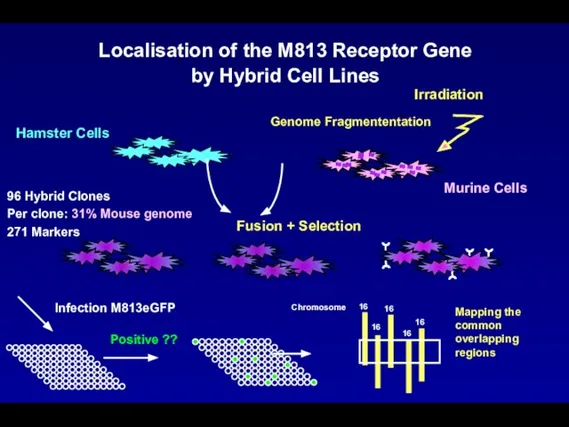
Hamster Cells
Murine Cells
Irradiation
Genome Fragmententation
Fusion + Selection
96 Hybrid Clones
Per clone: 31%
Hamster Cells
Murine Cells
Irradiation
Genome Fragmententation
Fusion + Selection
96 Hybrid Clones
Per clone: 31%
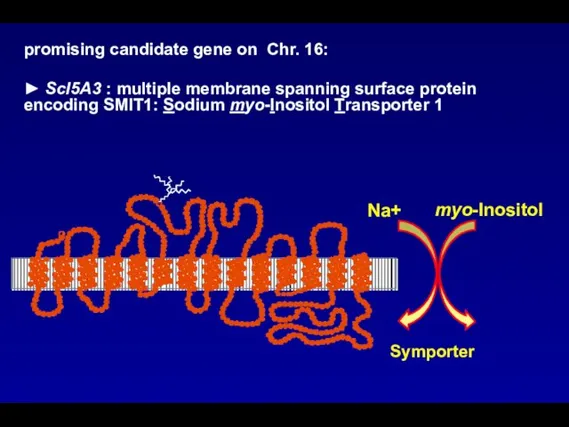
promising candidate gene on Chr. 16:
► Scl5A3 : multiple membrane spanning
promising candidate gene on Chr. 16:
► Scl5A3 : multiple membrane spanning
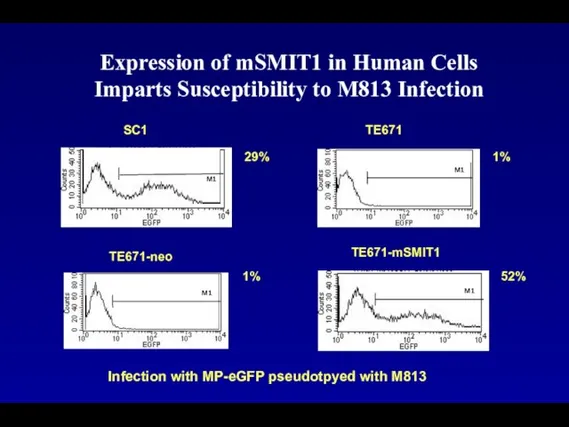
Expression of mSMIT1 in Human Cells
Imparts Susceptibility to M813 Infection
TE671-mSMIT1
52%
M1
SC1
M1
TE671-neo
1%
M1
TE671
1%
29%
M1
M1
Infection
Expression of mSMIT1 in Human Cells
Imparts Susceptibility to M813 Infection
TE671-mSMIT1
52%
M1
SC1
M1
TE671-neo
1%
M1
TE671
1%
29%
M1
M1
Infection
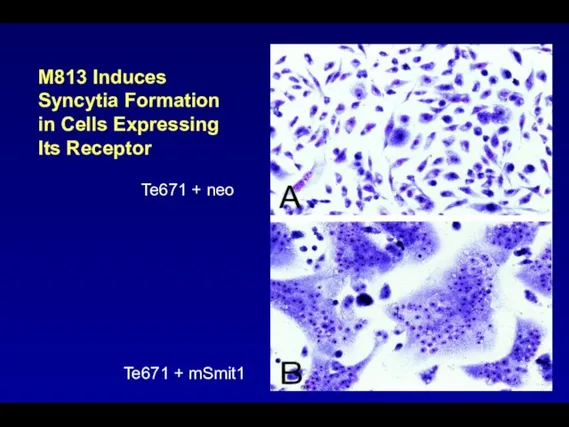
M813 Induces
Syncytia Formation
in Cells Expressing
Its Receptor
Te671 + neo
Te671 + mSmit1
M813 Induces
Syncytia Formation
in Cells Expressing
Its Receptor
Te671 + neo
Te671 + mSmit1
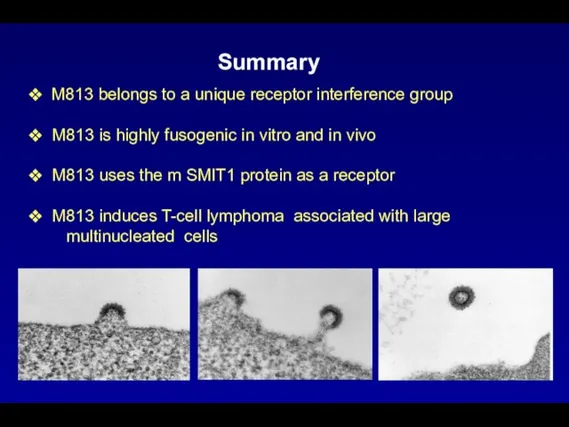
M813 belongs to a unique receptor interference group
M813 is highly
M813 belongs to a unique receptor interference group
M813 is highly

Acknowledgments
Dmitry Ivanov,
Pavel Spirin,
Tamara Semenova
Engelhardt Institute of Molecular Biology
Moscow, Russia
Sibyll Hein
Jürgen
Acknowledgments
Dmitry Ivanov,
Pavel Spirin,
Tamara Semenova
Engelhardt Institute of Molecular Biology
Moscow, Russia
Sibyll Hein
Jürgen
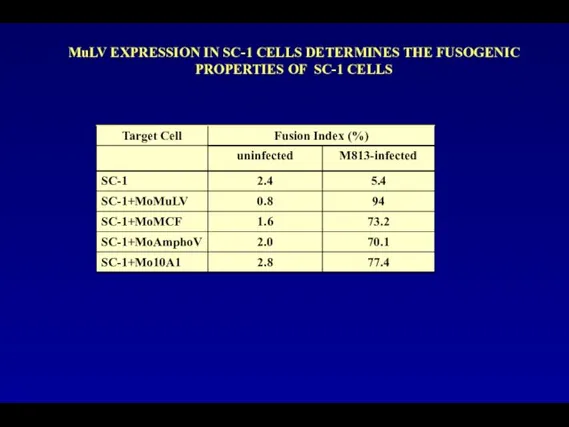
MuLV EXPRESSION IN SC-1 CELLS DETERMINES THE FUSOGENIC
PROPERTIES OF SC-1
MuLV EXPRESSION IN SC-1 CELLS DETERMINES THE FUSOGENIC
PROPERTIES OF SC-1
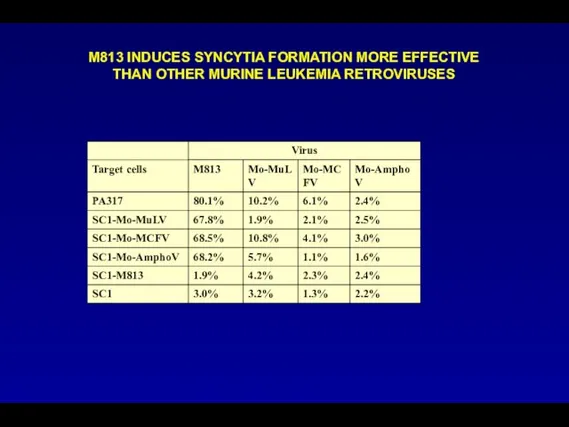
М813 INDUCES SYNCYTIA FORMATION MORE EFFECTIVE
THAN OTHER MURINE LEUKEMIA RETROVIRUSES
М813 INDUCES SYNCYTIA FORMATION MORE EFFECTIVE
THAN OTHER MURINE LEUKEMIA RETROVIRUSES

Fusion index (FI) = (N - S)/T
N – number of nuclei
Fusion index (FI) = (N - S)/T
N – number of nuclei
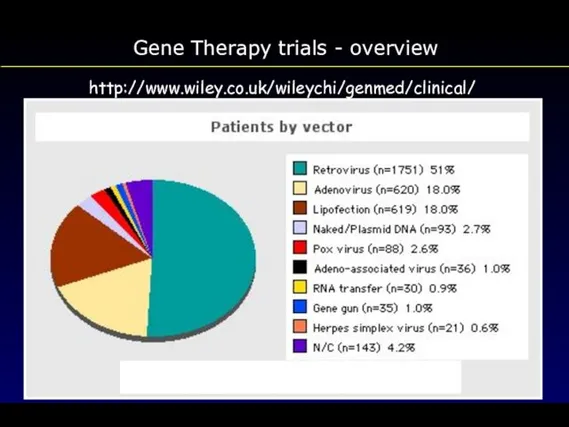
Gene Therapy trials - overview
http://www.wiley.co.uk/wileychi/genmed/clinical/
Gene Therapy trials - overview
http://www.wiley.co.uk/wileychi/genmed/clinical/
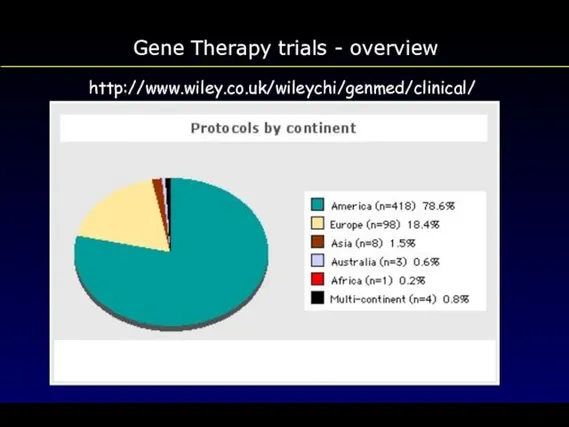
Gene Therapy trials - overview
http://www.wiley.co.uk/wileychi/genmed/clinical/
Gene Therapy trials - overview
http://www.wiley.co.uk/wileychi/genmed/clinical/
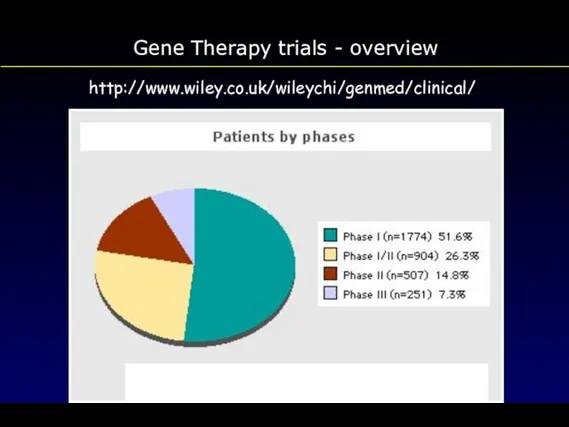
Gene Therapy trials - overview
http://www.wiley.co.uk/wileychi/genmed/clinical/
Gene Therapy trials - overview
http://www.wiley.co.uk/wileychi/genmed/clinical/

Инсерционный мутагенез
Встраивание провируса может приводить к трансформации клетки
Инсерционный мутагенез
Встраивание провируса может приводить к трансформации клетки

Открытие вирусов
1892 год
Д.И.Ивановский – инфекционный фильтрующийся агент, вызывающий табачную мозаику.
1898
Открытие вирусов
1892 год
Д.И.Ивановский – инфекционный фильтрующийся агент, вызывающий табачную мозаику.
1898
 Презентация на тему ДНК и РНК нуклеиновые кислоты
Презентация на тему ДНК и РНК нуклеиновые кислоты Жасушаның құрылысы
Жасушаның құрылысы Современные аспекты генетического анализа
Современные аспекты генетического анализа Этапы эволюции человека
Этапы эволюции человека Презентация на тему Птицы, кормящиеся в воздухе
Презентация на тему Птицы, кормящиеся в воздухе  ОХРАНА ЖИВОЙ ПРИРОДЫ
ОХРАНА ЖИВОЙ ПРИРОДЫ  Тип кишечнополостные
Тип кишечнополостные Презентация по биологии Простейшие Жгутиковые
Презентация по биологии Простейшие Жгутиковые  Насекомые - переносчики заболевания человека
Насекомые - переносчики заболевания человека Презентация на тему "Окружающий мир (3 класс)" - скачать презентации по Биологии
Презентация на тему "Окружающий мир (3 класс)" - скачать презентации по Биологии Нервная ткань
Нервная ткань Каталог гибридов подсолнечника и кукурузы 2021-2022
Каталог гибридов подсолнечника и кукурузы 2021-2022 Головной мозг
Головной мозг Генеалогическое древо моей семьи Два чувства в жизни близки нам. В них обретает сердце пищу: Любовь к отеческим гробам, Любовь к
Генеалогическое древо моей семьи Два чувства в жизни близки нам. В них обретает сердце пищу: Любовь к отеческим гробам, Любовь к  Дикие животные
Дикие животные Вегетативное размножение
Вегетативное размножение Строение растительных клеток
Строение растительных клеток Презентация на тему "Значение млекопитающих в природе и жизни человека" - скачать презентации по Биологии
Презентация на тему "Значение млекопитающих в природе и жизни человека" - скачать презентации по Биологии Презентация на тему "Василёчки - Васильки" - скачать презентации по Биологии
Презентация на тему "Василёчки - Васильки" - скачать презентации по Биологии Царство грибы. Общая характеристика грибов. Отдел Миксомицеты. Отдел Хитридиомицеты. Отдел Оомицеты. Отдел Зигомицеты
Царство грибы. Общая характеристика грибов. Отдел Миксомицеты. Отдел Хитридиомицеты. Отдел Оомицеты. Отдел Зигомицеты Строение и функции гемоглобина и миоглобина человека. Кривые диссоциации кислорода для гемоглобина и миоглобина человека
Строение и функции гемоглобина и миоглобина человека. Кривые диссоциации кислорода для гемоглобина и миоглобина человека Миграция радионуклидов по пищевым цепочкам
Миграция радионуклидов по пищевым цепочкам Оценка сортов яровой пшеницы в условиях Лысковского сортоучастка
Оценка сортов яровой пшеницы в условиях Лысковского сортоучастка Презентация Отряды насекомых
Презентация Отряды насекомых Introduction to the Nervous System
Introduction to the Nervous System Животный мир Северной Америки
Животный мир Северной Америки Класс Млекопитающие. 7 класс
Класс Млекопитающие. 7 класс ТЕМА УРОКА ВЛИЯНИЕ НАРКОГЕННЫХ ВЕЩЕСТВ НА ЗДОРОВЬЕ И СУДЬБУ ЧЕЛОВЕКА
ТЕМА УРОКА ВЛИЯНИЕ НАРКОГЕННЫХ ВЕЩЕСТВ НА ЗДОРОВЬЕ И СУДЬБУ ЧЕЛОВЕКА