- Эйкозаноиды и их функция

Содержание
- 2. Продолжение лекции 2 Рецепторы желчных кислот Существуют внутриклеточные ядерные активируемые фарнезолом* X рецепторы (FXR -члены семейства
- 3. Сопряженный с G-белком рецептор для желчных кислот (TRG) Обработка клеток, экспрессирующих TGR на поверхности клеток, активирует
- 4. Синергизм в действии рецепторов желчных кислот Последние данные показывают, что желчные кислоты через FXR влияют на
- 5. Эйкозаноиды В середине 30-х годов 20 века шведский ученый Эйлер (V. Euler) обнаружил в экстракте из
- 6. Функции эйкозаноидов Эйкозаноиды регулируют сокращение гладкой мускулатуры, влияя на диаметр сосудов и, таким образом, на артериальное
- 7. Что такое простагландины? Простагландины – биологически активные вещества, являющиеся производными ненасыщенных жирных кислот, состоящие из 20
- 8. Классификация простагландинов В зависимости от строения цикла и характера боковых цепей простагландины делят на несколько (9)
- 9. Разделение простагландинов на типы А В С D E F H J 1 2 3 4
- 10. Простациклины PGI Имеют 2 сопряженных кольца: одно пятичленное (как другие простагландины), а другое – тоже пятичленное,
- 11. ТРОМБОКСАНЫ Имеют шестичленное кольцо, включающее атом кислорода, и пероксидный мостик. Cинтезируются только в тромбоцитах, стимулируя их
- 12. Структура арахидоновой кислоты СН3(СН2)4-(СН=СН-СН2)3-СН=СН-(СН2)3СООН 20:4 (5с,8с,11с,14с) Эйкозаноиды являются производными арахидоновой кислоты, которая является омега-6 ненасыщенной аминокислотой.
- 13. Синтез простагландинов семейств 1, 2 и 3 из разных субстратов Арахидоновая кислота (20:4; С5, С8, С11,
- 14. Источники субстратов Полиеновые кислоты либо поступают в организм с пищей, либо образуются из незаменимых жирных кислот
- 15. Биологические эффекты различных простагландинов PGE2 (большинство тканей) – расслабление гладкой мускулатуры: расширение сосудов, но стимуляция сокращения
- 16. Действие фосфолипаз на фосфолипиды Инозитол 4,5-бисфосфат Активируются гормонами Фосфолипаза А1 Фосфолипаза А2 Фосфолипаза С Фосфолипаза D
- 17. Синтез эйкозаноидов из арахидоновой кислоты Катехоламины, брадикинин, ангиотензин II, гистамин, цитокины Циклооксигеназа 1/2 5’-липоксигеназа
- 18. Циклооксигеназный и липоксигеназный пути превращения полиеновых жирных кислот Синтез простагландинов начинается после освобождения полиеновых кислот из
- 19. Синтез простагландинов и тромбоксанов Время полужизни PGH2 0-100 s) Время полужизни PGH2 90 c 1. 2.
- 20. Для синтеза простагландинов необходим набор ферментов Получение первого интермедиата PGH2 обеспечивает мембранный фермент простагландин (РGH2)-синтетаза (циклооксигеназа),
- 21. Циклооксигеназа Циклооксигеназа (ЦОГ), простагландин G/H синтетаза, ЕС 1.14.99.1) обеспечивает первые две стадии синтеза простагландинов. Находится в
- 22. Структура и активные центры циклооксигеназы Cтруктура циклооксигеназы (простагландин-эндопероксидазы). Это гомодимер c аналогом арахидоновой кислоты (фторбипрофен (желтый)
- 23. Циклооксигеназа Активный центр циклооксигеназы находится на вершине канала в виде буквы L, сформированного мембранным доменом. Вход
- 24. Активный центр пероксидазы Модель активного центра пероксидазы со связанным простагландином PGG2. Гем обозначен красным вместе с
- 25. Аспирин (ацетилсалициловая кислота) – ингибитор циклоксигеназы одно из НПВС Салициловый эфир уксусной кислоты известен под торговой
- 26. Ингибирование циклооксигеназы ацетилсалициловой кислотой (аспирином). Необратимое ацетилирование остатка серина в положении 529 (530) в молекуле циклооксигеназы
- 27. Лейкотриены Характерная особенность лейкотриенов – отсутствие циклической структуры и наличие трех сопряженных связей (три-ен). Вырабатываются они
- 28. Образование лейкотриенов 5-липоксигеназа Липоксигеназы проводят окисление 5-го, 12-го и 15-го атома углерода в зависимости от типа
- 29. Лейкотриен В4 Лейкоциты и клетки эпителия сосудов
- 30. Лейкотриены LTD4 и LTE4 Лейкротриен LTС4 в 6-м положении содержит глутатион, трипептид, состоящий из трех аминокислот
- 31. Функции лейкотриенов LTС4, LTD4, LTE4, клетки белой крови, альвеолярные макрофаги: расширение сосудов, увеличение их проницаемости, сокращение
- 32. Фармакологическое устранение эффектов лейкотриенов В настоящее время имеется два способа фармакологического устранения эффектов лейкотриенов: антагонисты рецепторов
- 33. Регуляция скорости синтеза простагландинов Скорость биосинтеза эйкозаноидов зависит от общего количества свободных жирных кислот. Вещества, влияющие
- 34. Липокортин (аннексин) Липокортин 1 (аннексин-1) белок из 346 аминокислот с молекулярной массой 38,716 Да. Липокортин-1 —
- 35. Инактивация эйкозаноидов Все эйкозаноиды инактивируются быстро, величина Т0,5 для свободных эйкозаноидов оставляет обычно от нескольких секунд
- 36. Трансклеточный синтез эйкозаноидов Некоторые клетки имеют полный набор ферментов, необходимых для продукции биологически активных простагландинов и
- 37. Трансклеточный синтез простагландинов Превращение арахидоновой кислоты осуществляется в клетке одного типа (клетка донор), а затем интермедиат
- 38. Роль эйкозаноидов в развитиии воспаления Воспаление – реакция организма на повреждение или инфекцию, направленная на уничтожение
- 39. Роль эйкозаноиов в формировании тромба В норме клетки эндотелия продуцируют простациклин I2, который препятствует агрегации тромбоцитов
- 40. Механизмы действия простагландинов Простагландины взаимодействуют с рецепторами цитоплазматических мембран; Один и тот же простагландин может действовать
- 41. Мембранные рецепторы простагландинов PGE2: четыре подтипа рецепторов (EP-1 четыре подтипа рецепторов (EP-1 , EP-2 четыре подтипа
- 42. Рецепторы, взаимодействующие с G-белками, которые регулируют активность ферментов, производящих вторичные мессенджеры Компоненты системы: 1. Рецептор с
- 43. Изоформы ЦОГ У человека есть два гена, кодирующих ЦОГ: COX-1 и COX-2. ЦОГ-1 считали конститутивной формой,
- 44. Изоформы ЦОГ ЦОГ-2 считали индуцибельной изоформой, которая включается при определённых ситуациях, например, при воспалении. ЦОГ-2 экспрессируется
- 45. Изоформы циклооксигеназы (ЦОГ-2 гипотеза) Циклооксигеназа (ЦОГ) ЦОГ-1 конститутивный фермент ЦОГ-2 конститутивный фермент в мозге и корковом
- 46. Изоформы ЦОГ Ген ЦОГ-1, Ptgs-1, кодирует иРНК, которая является относительно стабильной. Ген ЦОГ-2, Ptgs-2, активируется под
- 47. Неспецифические ингибиторы ЦОГ Аспирин – необратимо ингибирует ЦОГ-1 и ЦОГ-2 за счет ацетилирования серина вблизи активного
- 48. Специфические ингибиторы изоформ ЦОГ Селективные ингибиторы ЦОГ-2 (вальдекоксиб, рофекоксиб, целекоксиб) были синтезированы для уменьшения токсического воздействия
- 49. С чем связаны проблемы применения ингибиторов ЦОГ-2? Воздействия на ЦОГ отражаются на синтезе регуляторов гемостаза -
- 50. С чем связаны проблемы применения ингибиторов ЦОГ-2? Простациклин (PGI2) - вазодилататор и ингибитор активации тромбоцитов. Он
- 51. Физиологический антагонизм между простациклином (PGI 2) из эндотелия кровеносных сосудов и тромбоксаном А2 (TXA2) из тромбоцитов
- 52. С чем связаны проблемы применения ингибиторов ЦОГ-2? У здоровых людей возрастание риска тромбообразования при ингибировании ЦОГ-2
- 53. Последствия ингибирования ЦОГ-2 в нормальных и атеросклеротических артериях норма атеросклероз Ингибитор ЦОГ-2 PGI TxA2 TxA2 PGI
- 54. Новый взгляд на изоформы ЦОГ Исследование мышей генетически дефицитных по ЦОГ-1 или ЦОГ-2 , как и
- 56. Скачать презентацию

Продолжение лекции 2
Рецепторы желчных кислот
Существуют внутриклеточные ядерные активируемые фарнезолом* X
Продолжение лекции 2
Рецепторы желчных кислот
Существуют внутриклеточные ядерные активируемые фарнезолом* X

Сопряженный с G-белком рецептор для желчных кислот (TRG)
Обработка клеток, экспрессирующих TGR
Сопряженный с G-белком рецептор для желчных кислот (TRG)
Обработка клеток, экспрессирующих TGR

Синергизм в действии рецепторов желчных кислот
Последние данные показывают, что желчные кислоты
Синергизм в действии рецепторов желчных кислот
Последние данные показывают, что желчные кислоты

Эйкозаноиды
В середине 30-х годов 20 века шведский ученый Эйлер (V. Euler) обнаружил в
Эйкозаноиды
В середине 30-х годов 20 века шведский ученый Эйлер (V. Euler) обнаружил в

Функции эйкозаноидов
Эйкозаноиды регулируют сокращение гладкой мускулатуры, влияя на диаметр сосудов и,
Функции эйкозаноидов
Эйкозаноиды регулируют сокращение гладкой мускулатуры, влияя на диаметр сосудов и,
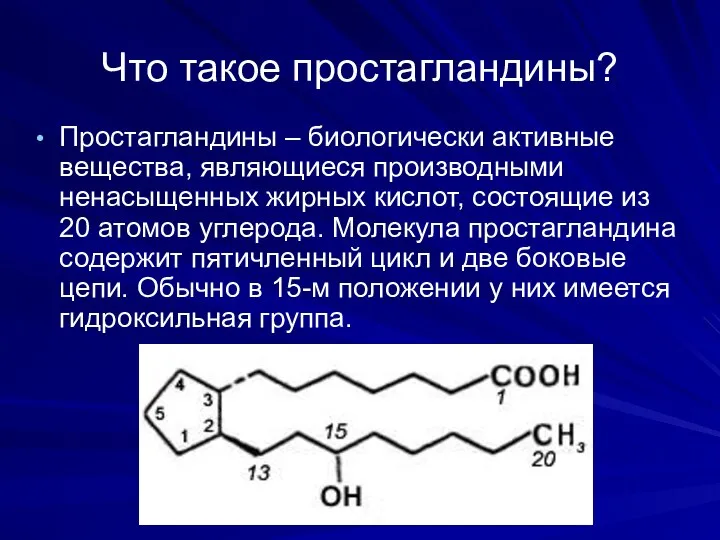
Что такое простагландины?
Простагландины – биологически активные вещества, являющиеся производными ненасыщенных жирных
Что такое простагландины?
Простагландины – биологически активные вещества, являющиеся производными ненасыщенных жирных

Классификация простагландинов
В зависимости от строения цикла и характера боковых цепей простагландины
Классификация простагландинов
В зависимости от строения цикла и характера боковых цепей простагландины
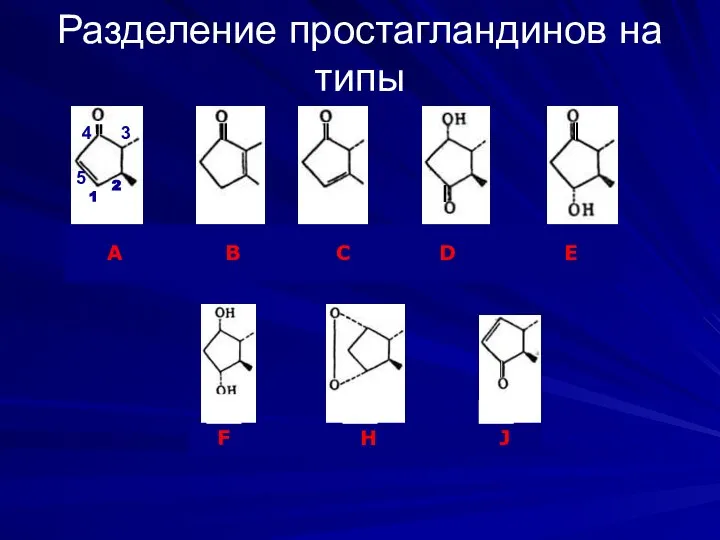
Разделение простагландинов на типы
А В С D E
F H J
1
2
3
4
5
Разделение простагландинов на типы
А В С D E
F H J
1
2
3
4
5
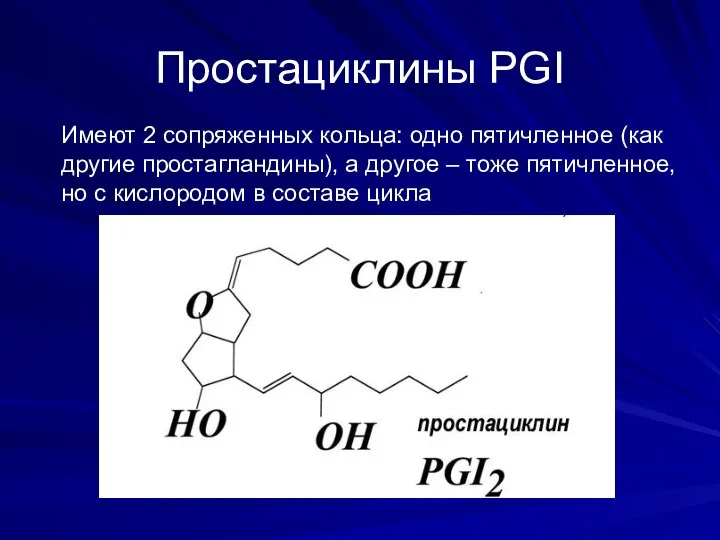
Простациклины PGI
Имеют 2 сопряженных кольца: одно пятичленное (как другие простагландины), а
Простациклины PGI
Имеют 2 сопряженных кольца: одно пятичленное (как другие простагландины), а
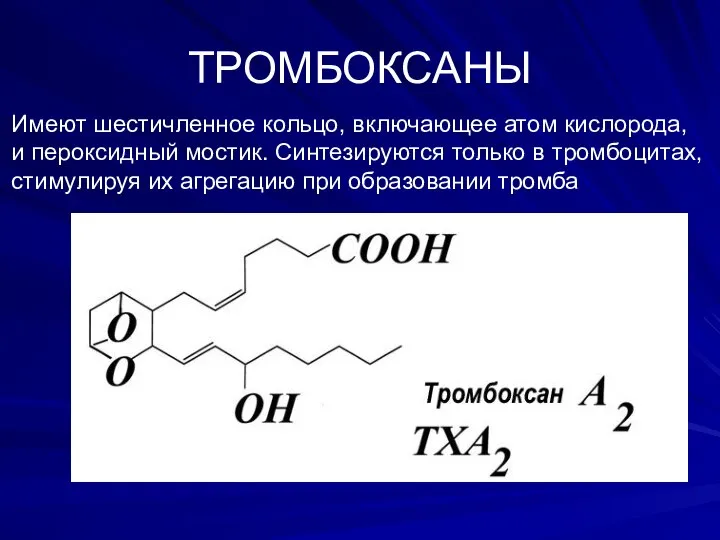
ТРОМБОКСАНЫ
Имеют шестичленное кольцо, включающее атом кислорода,
и пероксидный мостик. Cинтезируются только в
ТРОМБОКСАНЫ
Имеют шестичленное кольцо, включающее атом кислорода,
и пероксидный мостик. Cинтезируются только в

Структура арахидоновой кислоты
СН3(СН2)4-(СН=СН-СН2)3-СН=СН-(СН2)3СООН
20:4 (5с,8с,11с,14с)
Эйкозаноиды являются производными арахидоновой кислоты, которая
Структура арахидоновой кислоты
СН3(СН2)4-(СН=СН-СН2)3-СН=СН-(СН2)3СООН
20:4 (5с,8с,11с,14с)
Эйкозаноиды являются производными арахидоновой кислоты, которая

Синтез простагландинов семейств 1, 2 и 3 из разных субстратов
Арахидоновая
Синтез простагландинов семейств 1, 2 и 3 из разных субстратов
Арахидоновая

Источники субстратов
Полиеновые кислоты либо поступают в организм с пищей, либо образуются
Источники субстратов
Полиеновые кислоты либо поступают в организм с пищей, либо образуются

Биологические эффекты различных простагландинов
PGE2 (большинство тканей) – расслабление гладкой мускулатуры: расширение
Биологические эффекты различных простагландинов
PGE2 (большинство тканей) – расслабление гладкой мускулатуры: расширение
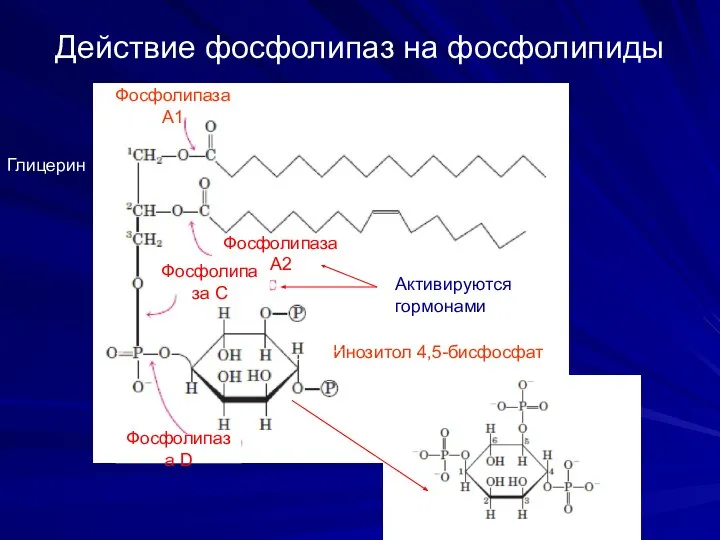
Действие фосфолипаз на фосфолипиды
Инозитол 4,5-бисфосфат
Активируются
гормонами
Фосфолипаза А1
Фосфолипаза А2
Фосфолипаза С
Фосфолипаза D
Глицерин
Действие фосфолипаз на фосфолипиды
Инозитол 4,5-бисфосфат
Активируются
гормонами
Фосфолипаза А1
Фосфолипаза А2
Фосфолипаза С
Фосфолипаза D
Глицерин
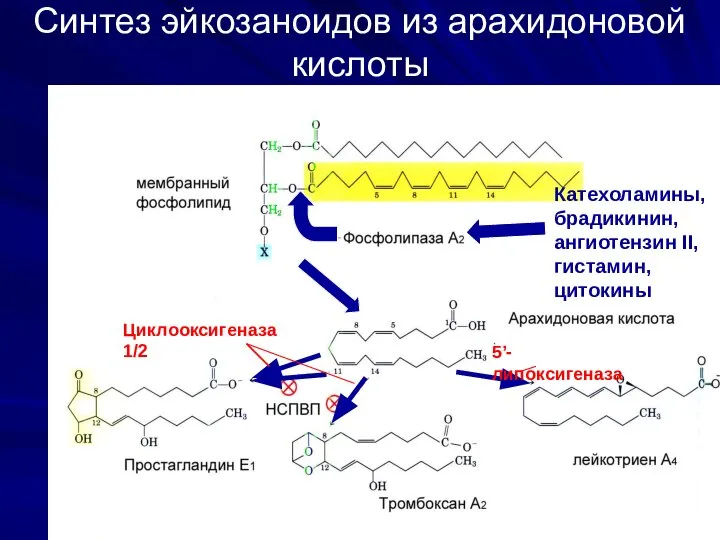
Синтез эйкозаноидов из арахидоновой кислоты
Катехоламины, брадикинин,
ангиотензин II,
гистамин, цитокины
Циклооксигеназа 1/2
5’-липоксигеназа
Синтез эйкозаноидов из арахидоновой кислоты
Катехоламины, брадикинин,
ангиотензин II,
гистамин, цитокины
Циклооксигеназа 1/2
5’-липоксигеназа

Циклооксигеназный и липоксигеназный пути превращения полиеновых жирных кислот
Синтез простагландинов начинается после
Циклооксигеназный и липоксигеназный пути превращения полиеновых жирных кислот
Синтез простагландинов начинается после
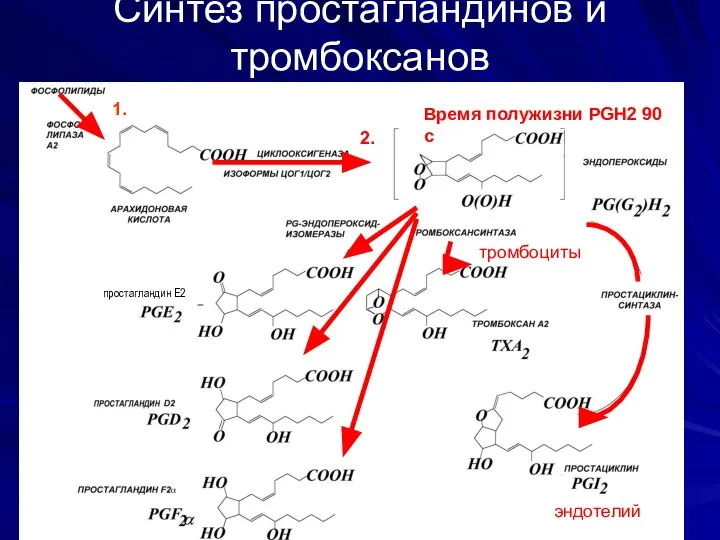
Синтез простагландинов и тромбоксанов
Время полужизни PGH2 0-100 s)
Время полужизни PGH2
Синтез простагландинов и тромбоксанов
Время полужизни PGH2 0-100 s)
Время полужизни PGH2

Для синтеза простагландинов необходим набор ферментов
Получение первого интермедиата PGH2 обеспечивает мембранный
Для синтеза простагландинов необходим набор ферментов
Получение первого интермедиата PGH2 обеспечивает мембранный

Циклооксигеназа
Циклооксигеназа (ЦОГ), простагландин G/H синтетаза, ЕС 1.14.99.1) обеспечивает первые две
Циклооксигеназа
Циклооксигеназа (ЦОГ), простагландин G/H синтетаза, ЕС 1.14.99.1) обеспечивает первые две
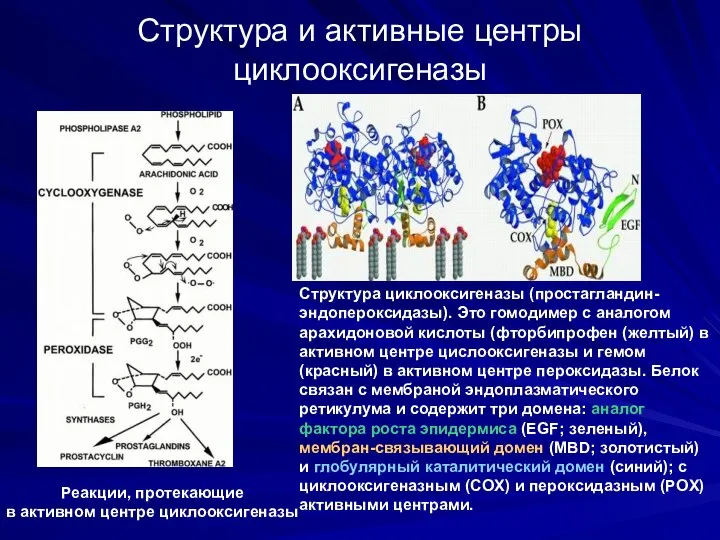
Структура и активные центры циклооксигеназы
Cтруктура циклооксигеназы (простагландин-эндопероксидазы). Это гомодимер c аналогом
Структура и активные центры циклооксигеназы
Cтруктура циклооксигеназы (простагландин-эндопероксидазы). Это гомодимер c аналогом
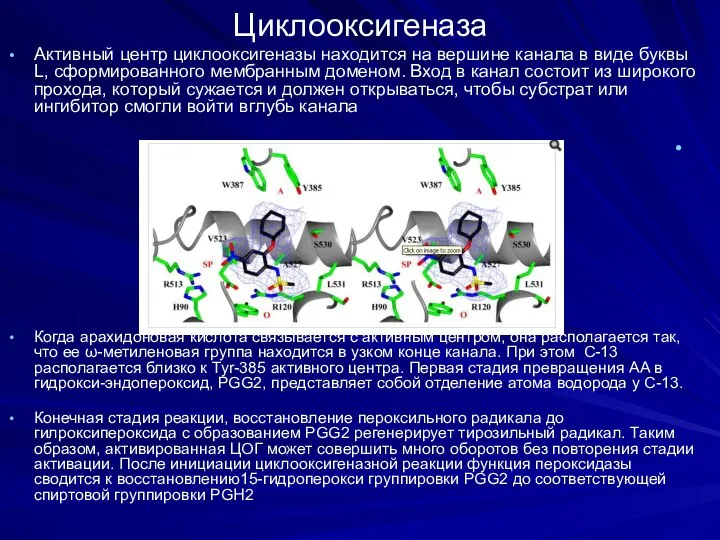
Циклооксигеназа
Активный центр циклооксигеназы находится на вершине канала в виде буквы L,
Циклооксигеназа
Активный центр циклооксигеназы находится на вершине канала в виде буквы L,
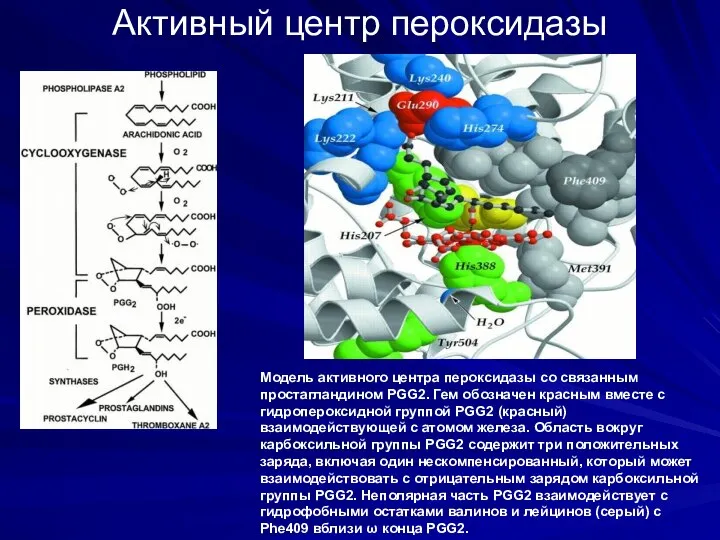
Активный центр пероксидазы
Модель активного центра пероксидазы со связанным простагландином PGG2. Гем
Активный центр пероксидазы
Модель активного центра пероксидазы со связанным простагландином PGG2. Гем
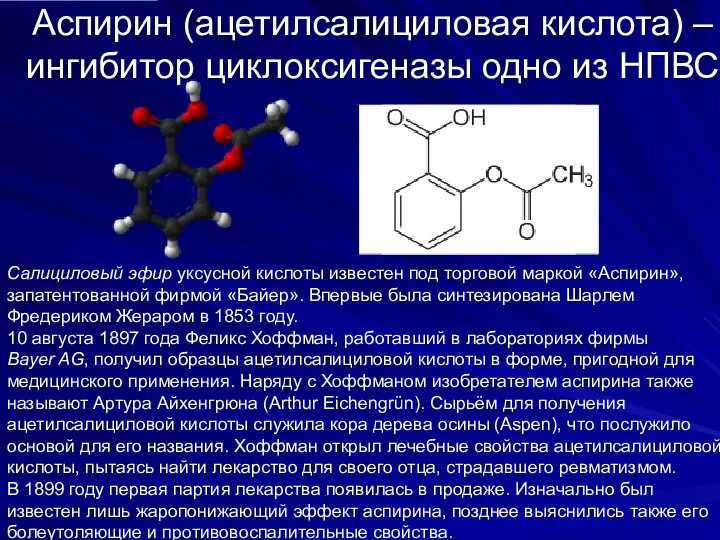
Аспирин (ацетилсалициловая кислота) – ингибитор циклоксигеназы одно из НПВС
Салициловый эфир уксусной
Аспирин (ацетилсалициловая кислота) – ингибитор циклоксигеназы одно из НПВС
Салициловый эфир уксусной
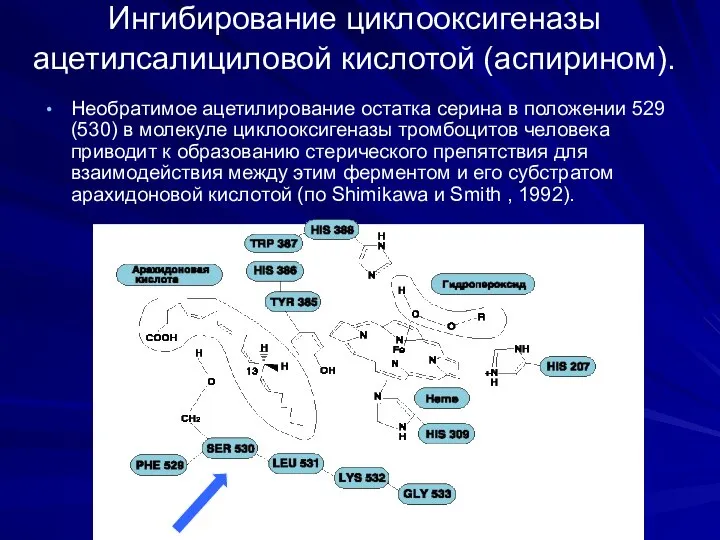
Ингибирование циклооксигеназы ацетилсалициловой кислотой (аспирином).
Необратимое ацетилирование остатка серина в положении
Ингибирование циклооксигеназы ацетилсалициловой кислотой (аспирином).
Необратимое ацетилирование остатка серина в положении

Лейкотриены
Характерная особенность лейкотриенов – отсутствие циклической структуры и наличие трех сопряженных
Лейкотриены
Характерная особенность лейкотриенов – отсутствие циклической структуры и наличие трех сопряженных
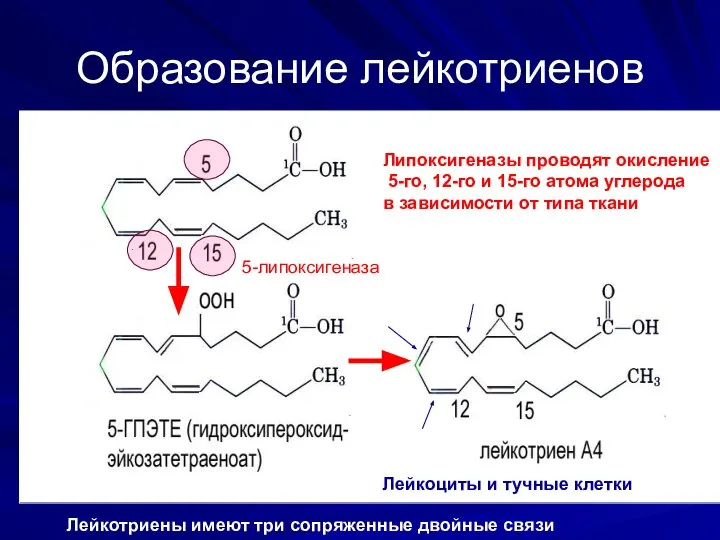
Образование лейкотриенов
5-липоксигеназа
Липоксигеназы проводят окисление
5-го, 12-го и 15-го атома углерода
в
Образование лейкотриенов
5-липоксигеназа
Липоксигеназы проводят окисление
5-го, 12-го и 15-го атома углерода
в
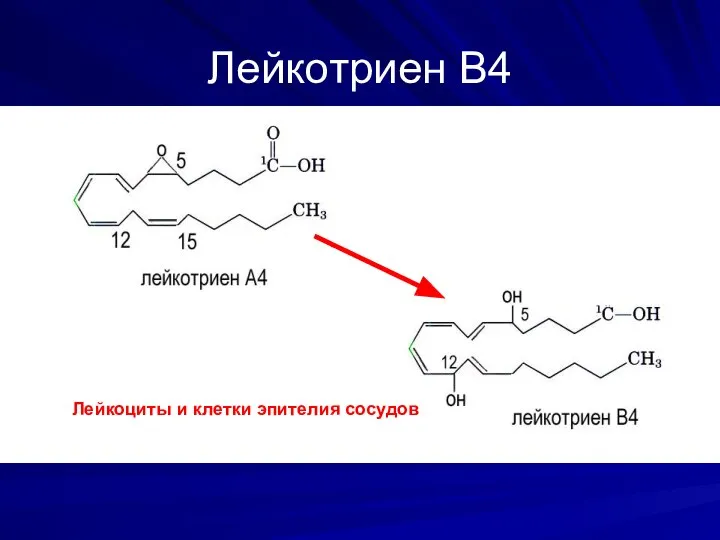
Лейкотриен В4
Лейкоциты и клетки эпителия сосудов
Лейкотриен В4
Лейкоциты и клетки эпителия сосудов
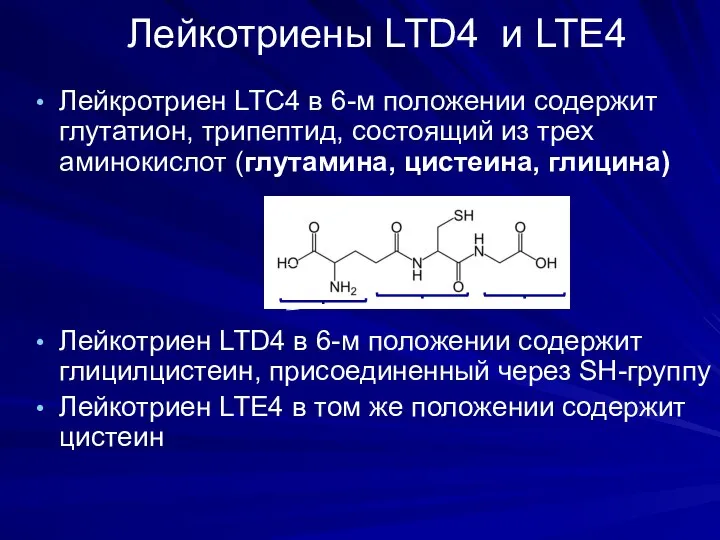
Лейкотриены LTD4 и LTE4
Лейкротриен LTС4 в 6-м положении содержит глутатион, трипептид,
Лейкотриены LTD4 и LTE4
Лейкротриен LTС4 в 6-м положении содержит глутатион, трипептид,

Функции лейкотриенов
LTС4, LTD4, LTE4, клетки белой крови, альвеолярные макрофаги: расширение сосудов,
Функции лейкотриенов
LTС4, LTD4, LTE4, клетки белой крови, альвеолярные макрофаги: расширение сосудов,

Фармакологическое устранение эффектов лейкотриенов
В настоящее время имеется два способа фармакологического устранения
Фармакологическое устранение эффектов лейкотриенов
В настоящее время имеется два способа фармакологического устранения

Регуляция скорости синтеза простагландинов
Скорость биосинтеза эйкозаноидов зависит от общего количества свободных
Регуляция скорости синтеза простагландинов
Скорость биосинтеза эйкозаноидов зависит от общего количества свободных

Липокортин (аннексин)
Липокортин 1 (аннексин-1) белок из 346 аминокислот с молекулярной массой
Липокортин (аннексин)
Липокортин 1 (аннексин-1) белок из 346 аминокислот с молекулярной массой

Инактивация эйкозаноидов
Все эйкозаноиды инактивируются быстро, величина Т0,5 для свободных эйкозаноидов оставляет
Инактивация эйкозаноидов
Все эйкозаноиды инактивируются быстро, величина Т0,5 для свободных эйкозаноидов оставляет

Трансклеточный синтез эйкозаноидов
Некоторые клетки имеют полный набор ферментов, необходимых для продукции
Трансклеточный синтез эйкозаноидов
Некоторые клетки имеют полный набор ферментов, необходимых для продукции
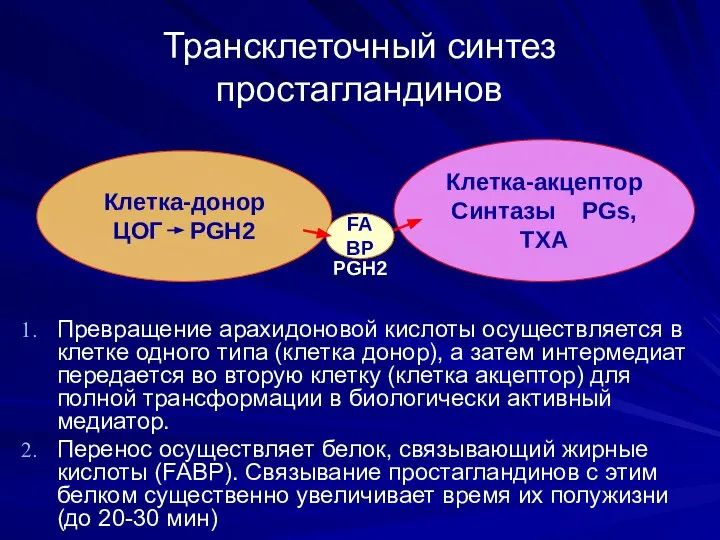
Трансклеточный синтез простагландинов
Превращение арахидоновой кислоты осуществляется в клетке одного типа (клетка
Трансклеточный синтез простагландинов
Превращение арахидоновой кислоты осуществляется в клетке одного типа (клетка

Роль эйкозаноидов в развитиии воспаления
Воспаление – реакция организма на повреждение или
Роль эйкозаноидов в развитиии воспаления
Воспаление – реакция организма на повреждение или

Роль эйкозаноиов в формировании тромба
В норме клетки эндотелия продуцируют простациклин I2,
Роль эйкозаноиов в формировании тромба
В норме клетки эндотелия продуцируют простациклин I2,

Механизмы действия простагландинов
Простагландины взаимодействуют с рецепторами цитоплазматических мембран;
Один и тот же
Механизмы действия простагландинов
Простагландины взаимодействуют с рецепторами цитоплазматических мембран;
Один и тот же

Мембранные рецепторы простагландинов
PGE2: четыре подтипа рецепторов (EP-1 четыре подтипа рецепторов (EP-1
Мембранные рецепторы простагландинов
PGE2: четыре подтипа рецепторов (EP-1 четыре подтипа рецепторов (EP-1
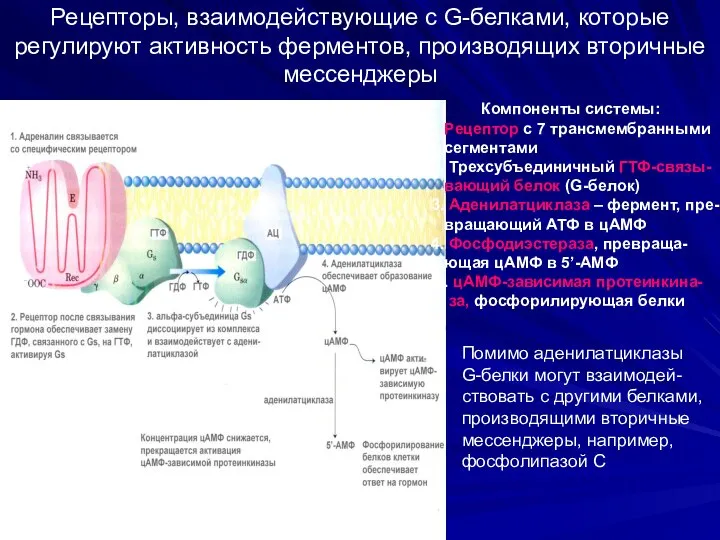
Рецепторы, взаимодействующие с G-белками, которые регулируют активность ферментов, производящих вторичные мессенджеры
Компоненты
Рецепторы, взаимодействующие с G-белками, которые регулируют активность ферментов, производящих вторичные мессенджеры
Компоненты

Изоформы ЦОГ
У человека есть два гена, кодирующих ЦОГ: COX-1 и COX-2.
Изоформы ЦОГ
У человека есть два гена, кодирующих ЦОГ: COX-1 и COX-2.

Изоформы ЦОГ
ЦОГ-2 считали индуцибельной изоформой, которая включается при определённых ситуациях, например,
Изоформы ЦОГ
ЦОГ-2 считали индуцибельной изоформой, которая включается при определённых ситуациях, например,

Изоформы циклооксигеназы (ЦОГ-2 гипотеза)
Циклооксигеназа (ЦОГ)
ЦОГ-1 конститутивный
фермент
ЦОГ-2 конститутивный фермент
в мозге
Изоформы циклооксигеназы (ЦОГ-2 гипотеза)
Циклооксигеназа (ЦОГ)
ЦОГ-1 конститутивный
фермент
ЦОГ-2 конститутивный фермент
в мозге

Изоформы ЦОГ
Ген ЦОГ-1, Ptgs-1, кодирует иРНК, которая является относительно стабильной. Ген
Изоформы ЦОГ
Ген ЦОГ-1, Ptgs-1, кодирует иРНК, которая является относительно стабильной. Ген

Неспецифические ингибиторы ЦОГ
Аспирин – необратимо ингибирует ЦОГ-1 и ЦОГ-2 за счет
Неспецифические ингибиторы ЦОГ
Аспирин – необратимо ингибирует ЦОГ-1 и ЦОГ-2 за счет

Специфические ингибиторы изоформ ЦОГ
Селективные ингибиторы ЦОГ-2 (вальдекоксиб, рофекоксиб, целекоксиб) были
Специфические ингибиторы изоформ ЦОГ
Селективные ингибиторы ЦОГ-2 (вальдекоксиб, рофекоксиб, целекоксиб) были

С чем связаны проблемы применения ингибиторов ЦОГ-2?
Воздействия на ЦОГ отражаются на
С чем связаны проблемы применения ингибиторов ЦОГ-2?
Воздействия на ЦОГ отражаются на

С чем связаны проблемы применения ингибиторов ЦОГ-2?
Простациклин (PGI2) - вазодилататор и
С чем связаны проблемы применения ингибиторов ЦОГ-2?
Простациклин (PGI2) - вазодилататор и
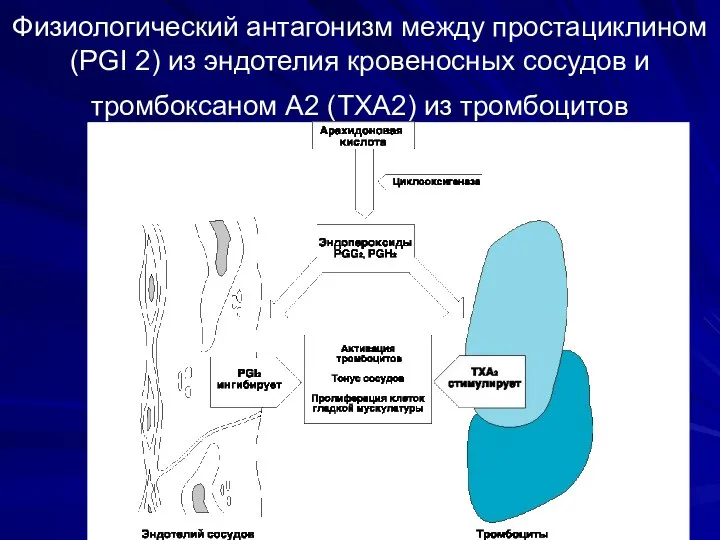
Физиологический антагонизм между простациклином (PGI 2) из эндотелия кровеносных сосудов и
Физиологический антагонизм между простациклином (PGI 2) из эндотелия кровеносных сосудов и

С чем связаны проблемы применения ингибиторов ЦОГ-2?
У здоровых людей возрастание риска
С чем связаны проблемы применения ингибиторов ЦОГ-2?
У здоровых людей возрастание риска
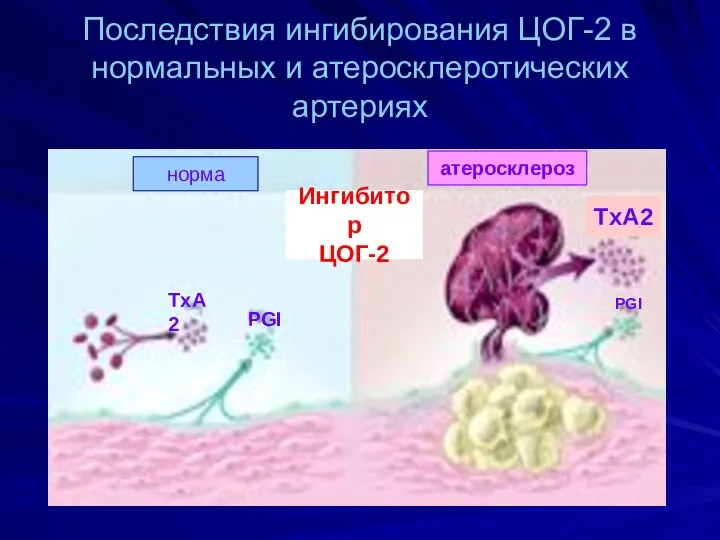
Последствия ингибирования ЦОГ-2 в нормальных и атеросклеротических артериях
норма
атеросклероз
Ингибитор
ЦОГ-2
PGI
TxA2
TxA2
PGI
Последствия ингибирования ЦОГ-2 в нормальных и атеросклеротических артериях
норма
атеросклероз
Ингибитор
ЦОГ-2
PGI
TxA2
TxA2
PGI

Новый взгляд на изоформы ЦОГ
Исследование мышей генетически дефицитных по ЦОГ-1 или
Новый взгляд на изоформы ЦОГ
Исследование мышей генетически дефицитных по ЦОГ-1 или
 Лекция 3. Тип Ресничные (Ciliophora)
Лекция 3. Тип Ресничные (Ciliophora) Презентация на тему Экология атмосферы
Презентация на тему Экология атмосферы  СИСТЕМА: КЛЕТКА- ГОСУДАРСТВО-ЦАРСТВО ОБОБЩАЮЩИЙ УРОК ПО ТЕМЕ «СТРОЕНИЕ КЛЕТКИ» Учитель биологии Пелыньо Ирина Васильевна
СИСТЕМА: КЛЕТКА- ГОСУДАРСТВО-ЦАРСТВО ОБОБЩАЮЩИЙ УРОК ПО ТЕМЕ «СТРОЕНИЕ КЛЕТКИ» Учитель биологии Пелыньо Ирина Васильевна  Изучение растений Красной книги Кубани
Изучение растений Красной книги Кубани Биологические и антропогенные факторы
Биологические и антропогенные факторы Рефлексы.
Рефлексы. Про абиогенез
Про абиогенез Презентация на тему Отряд Поденки
Презентация на тему Отряд Поденки Классификация лесов
Классификация лесов Сувели и прочие аномальные разрастания стебля
Сувели и прочие аномальные разрастания стебля Рыба и рыбные товары
Рыба и рыбные товары Плесневые грибы
Плесневые грибы Образцы опорных конспектов. 8 класс
Образцы опорных конспектов. 8 класс Микропрепараты. Растительные ткани
Микропрепараты. Растительные ткани Биологический вид. Критерии вида
Биологический вид. Критерии вида Capybara
Capybara Наследование при дигибридном скрещивании
Наследование при дигибридном скрещивании  Регуляция физиологических функций организма
Регуляция физиологических функций организма Информация и эволюция живого мира Работа студента группы 212-псо Кузнецова Даниила.
Информация и эволюция живого мира Работа студента группы 212-псо Кузнецова Даниила. Дарвинский заповедник
Дарвинский заповедник Cells and Organelles
Cells and Organelles Сельскохозяйственные животные
Сельскохозяйственные животные Выделительная система ребенка
Выделительная система ребенка Презентация на тему "Дарвин о причинах эволюции" - скачать презентации по Биологии
Презентация на тему "Дарвин о причинах эволюции" - скачать презентации по Биологии “Лелекоподібні” Виконала учениця 8 класу Прачук Анастасія
“Лелекоподібні” Виконала учениця 8 класу Прачук Анастасія Дигибридное и полигибридное скрещивание. Третий закон Менделя – закон независимого комбинирования Урок по основам генетики 10 кл
Дигибридное и полигибридное скрещивание. Третий закон Менделя – закон независимого комбинирования Урок по основам генетики 10 кл Эволюция человека Подготовил учитель биологии высшей квалификационной категории Пету
Эволюция человека Подготовил учитель биологии высшей квалификационной категории Пету Структура и функции растительной клетки
Структура и функции растительной клетки