- Формирование околоводной растительности

Содержание
- 2. Выход на сушу В 1859 году канадский геолог Джеймс Досон в девонских отложениях на полуострове Гаспе
- 3. Риния — одно из древнейших сосудистых растений (ранний девон, около 410 млн лет назад).
- 5. Псилот голый (Psilotum nudum)
- 6. породы раннедевонского возраста - 415 млн. лет назад. различные виды бактерий, водорослей и грибов - полностью
- 7. Выход на сушу : приспособления Тело растения -две части: надземная и подземная. Одни ответвления - теломы,
- 8. Специализация подземных органов Увеличивалась поглощающая способность - обильное ветвление и возникновению волосконосного слоя. Защиту апикальной меристемы
- 9. надземная часть телом риниофитов - преобразовался в побег Побег - стебли и листья. В качестве листьев
- 10. В процессе эволюции совершенствовалась и система ветвления телома. У самых примитивных высших растений телом ветвился вильчато
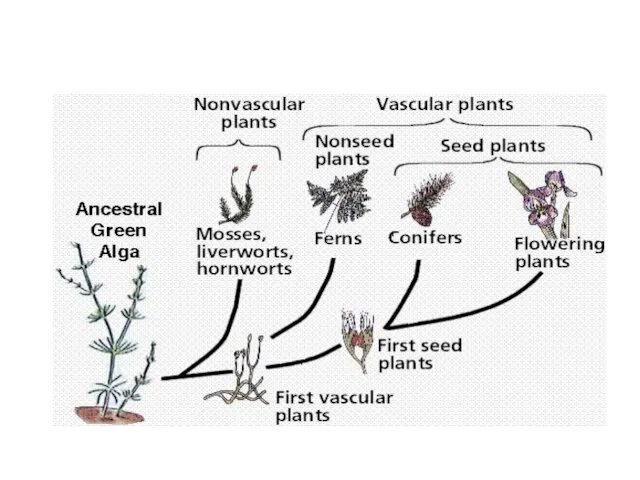
- 11. Наземные растения Бессосудистые споровые: Мохообразные – Bryophyta Сосудисто-споровые: Псилотовидные – Psilotophyta Плауновидные – Lycopodiophyta Хвощевидные –
- 13. Соотношение спорофита и гаметофита
- 14. ОТДЕЛ Моховидные (BRYOPHYTA) Моховидные - древняя группа слепая ветвь в эволюции растений. Класс Бриофиты ( Bryopsida)
- 15. Экология мхов В условиях повышенного увлажнения в умеренных широтах северного и южного полушарий идет болотообразовательный процесс,
- 16. Жизненный цикл из 2-х фаз развития, или ядерных фаз, растение с диплоидным набором хромосом – спорофит
- 17. Моховидные в жизненном цикле преобладает гаметофит. обеспечивает половое размножение выполняет основные вегетативные функции - фотосинтез, водоснабжение
- 18. групповые формы роста – плотные дерновины. возможность переносить неблагоприятные условия поглощать и сохранять влагу обеспечивает половое
- 19. Жизненный цикл У однодомных - половые органы - антеридии и архегонии на одном растении, у двудомных
- 20. Жизненный цикл Антеридии - мешочки на ножке с однослойной оболочкой. Внутри -сперматогенная ткань. Из каждой клетки
- 23. Оплодотворение при наличии капельножидкой воды Сперматозоиды - к архегонию - по слизистому каналу При слиянии гамет
- 24. Споры образуются в результате редукционного деления спорогенной ткани высеивание спор пленочка - эпифрагма крепится к краям
- 25. Водные мхи Мхи интенсивно развиваются в стоячих водах, образуя моховые болота. К листостебельным мхам и близкой
- 26. У мха Drepanocladus побеги стелющиеся, буроватые. Он растет на мелководьях под тростником и осокой. Его мелкие
- 27. Fontinalis antipyretica
- 28. Отдел Псилотовидные – Psilotophyta включает класс Psilotopsida порядок – Psilotales семейство – Psilotaceae. 2 рода –
- 29. Отдел Плауновидные типичные листостебельные растения всегда есть корни микрофиллия - мелкие листья на стебле. Известны начиная
- 30. Листья плауновидных простые, реже слегка вильчато разветвленные на верхушке, с одной жилкой. Листовые прорывы стели отсутствуют.
- 32. На верхней стороне спорофиллов располагаются спорангии. Зрелый спорангий имеет почковидную форму и прикрепляется к спорофиллу короткой
- 33. Попав на землю, спора потоками воды уносится в углубления (трещины) почвы, где на глубине в несколько
- 34. На верхней поверхности заростка образуются антеридии и архегонии. Антеридии целиком погружены в ткань заростка. В них
- 37. Хвощевидные Современные хвощи — небольшие травянистые растения около 80 — 100 см высотой и 2 —
- 38. Надземные побеги хвоща полевого двух видов. Одни из них вегетативные, зеленые, мутовчато-разветвленные, летние. Надземные стебли ветвятся.
- 39. Однако для многих видов (например, Е. arvense) экспериментально установлено, что, поливая питательным раствором мужские заростки, на
- 40. У всех видов хвоща стебли обладают выраженной метамерией, то есть правильным чередованием узлов и междоузлий. Листья
- 41. Вторичноводные растения. Эволюция наземных растений. Лекция № 6
- 42. Отдел Папоротникообразные Папоротники относятся к числу наиболее древних групп высших растений. По своей древности они уступают
- 43. Вегетативные органы папоротникообразных Среди современных папоротников преобладают травянистые формы с крупными листьями и неразвитым стеблем. Деревянистые
- 44. Вегетативные органы папоротникообразных По сравнению со мхами, папоротники значительно лучше приспособлены к сухопутному образу жизни: крупные
- 45. Вегетативные органы папоротникообразных Крупные листья – вайи. В отличие от обычных листьев, вайи нарастают не основанием,
- 46. Генеративные органы папоротников На нижней стороне листовых пластинок появляются скопления спорангиев, именуемые сорусами. Сорус содержит булавовидные
- 47. Генеративные органы папоротников В спорангиях из диплоидной спорогенной ткани в результате мейоза образуются гаплоидные споры. Стенка
- 49. Королевский папоротник
- 50. Вайи
- 51. Генеративные органы папоротников
- 53. Жизненный цикл Попав в благоприятные условия (влажная почва) спора прорастает в небольшую (диаметром около 1 см)
- 54. Жизненный цикл Под пластинкой заростка скапливается испаряющаяся из почвы влага, поэтому вышедшие из антеридиев сперматозоиды обычно
- 56. Водные папоротники
- 57. Водные папоротники К порядку Сальвиниевые — водных папоротников — относится ряд приспособившихся к обитанию в воде
- 58. Водные папоротники Наиболее распространенные роды сальвиниевых — Марсилия и Сальвиния. Первая представляет собой небольшие прибрежные и
- 60. Скачать презентацию

Выход на сушу
В 1859 году канадский геолог Джеймс Досон в девонских
Выход на сушу
В 1859 году канадский геолог Джеймс Досон в девонских

Риния — одно из древнейших сосудистых растений (ранний девон, около 410 млн лет
Риния — одно из древнейших сосудистых растений (ранний девон, около 410 млн лет


Псилот голый (Psilotum nudum)
Псилот голый (Psilotum nudum)

породы раннедевонского возраста - 415 млн. лет назад.
различные виды бактерий,
породы раннедевонского возраста - 415 млн. лет назад.
различные виды бактерий,

Выход на сушу : приспособления
Тело растения -две части: надземная и подземная.
Выход на сушу : приспособления
Тело растения -две части: надземная и подземная.

Специализация подземных органов
Увеличивалась поглощающая способность - обильное ветвление и возникновению волосконосного
Специализация подземных органов
Увеличивалась поглощающая способность - обильное ветвление и возникновению волосконосного

надземная часть
телом риниофитов - преобразовался в побег
Побег - стебли и
надземная часть
телом риниофитов - преобразовался в побег
Побег - стебли и

В процессе эволюции совершенствовалась и система ветвления телома. У самых примитивных
В процессе эволюции совершенствовалась и система ветвления телома. У самых примитивных

Наземные растения
Бессосудистые споровые:
Мохообразные – Bryophyta
Сосудисто-споровые:
Псилотовидные – Psilotophyta
Плауновидные – Lycopodiophyta
Хвощевидные –
Наземные растения
Бессосудистые споровые:
Мохообразные – Bryophyta
Сосудисто-споровые:
Псилотовидные – Psilotophyta
Плауновидные – Lycopodiophyta
Хвощевидные –
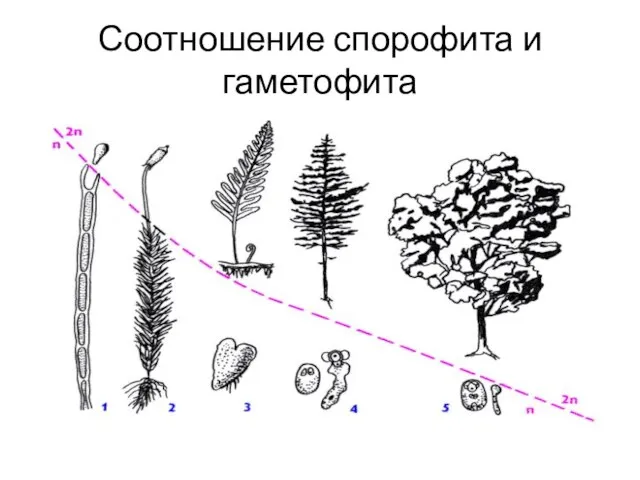
Соотношение спорофита и гаметофита
Соотношение спорофита и гаметофита

ОТДЕЛ Моховидные
(BRYOPHYTA)
Моховидные - древняя группа
слепая ветвь в эволюции растений.
Класс Бриофиты ( Bryopsida)
Класс
ОТДЕЛ Моховидные
(BRYOPHYTA)
Моховидные - древняя группа
слепая ветвь в эволюции растений.
Класс Бриофиты ( Bryopsida)
Класс

Экология мхов
В условиях повышенного увлажнения в умеренных широтах северного и южного
Экология мхов
В условиях повышенного увлажнения в умеренных широтах северного и южного

Жизненный цикл
из 2-х фаз развития, или ядерных фаз,
растение с
Жизненный цикл
из 2-х фаз развития, или ядерных фаз,
растение с

Моховидные
в жизненном цикле преобладает гаметофит.
обеспечивает половое размножение
выполняет основные вегетативные
Моховидные
в жизненном цикле преобладает гаметофит.
обеспечивает половое размножение
выполняет основные вегетативные

групповые формы роста – плотные дерновины.
возможность переносить неблагоприятные условия
поглощать и
групповые формы роста – плотные дерновины.
возможность переносить неблагоприятные условия
поглощать и

Жизненный цикл
У однодомных - половые органы - антеридии и архегонии на
Жизненный цикл
У однодомных - половые органы - антеридии и архегонии на

Жизненный цикл
Антеридии - мешочки на ножке с однослойной оболочкой. Внутри -сперматогенная
Жизненный цикл
Антеридии - мешочки на ножке с однослойной оболочкой. Внутри -сперматогенная
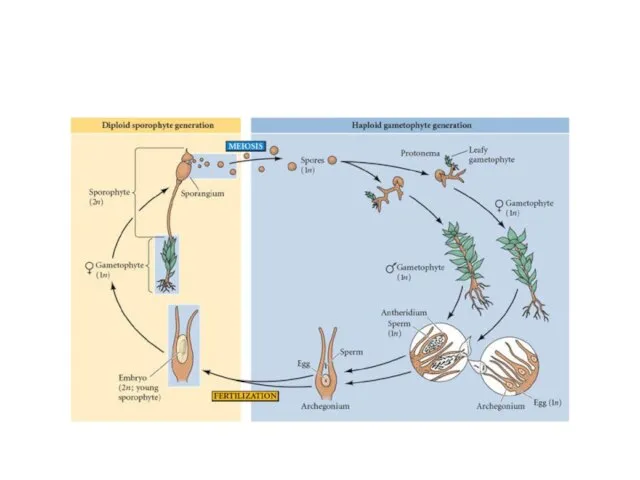

Оплодотворение
при наличии капельножидкой воды
Сперматозоиды - к архегонию - по
Оплодотворение
при наличии капельножидкой воды
Сперматозоиды - к архегонию - по

Споры
образуются в результате редукционного деления спорогенной ткани
высеивание спор
пленочка -
Споры
образуются в результате редукционного деления спорогенной ткани
высеивание спор
пленочка -

Водные мхи
Мхи интенсивно развиваются в стоячих водах, образуя моховые болота.
К
Водные мхи
Мхи интенсивно развиваются в стоячих водах, образуя моховые болота.
К

У мха Drepanocladus побеги стелющиеся, буроватые. Он растет на мелководьях под
У мха Drepanocladus побеги стелющиеся, буроватые. Он растет на мелководьях под

Fontinalis antipyretica
Fontinalis antipyretica

Отдел Псилотовидные – Psilotophyta
включает класс Psilotopsida
порядок – Psilotales
семейство – Psilotaceae.
2 рода – Tmesipteris (10 видов), Psilotum (2 вида).
Растения
Отдел Псилотовидные – Psilotophyta
включает класс Psilotopsida
порядок – Psilotales
семейство – Psilotaceae.
2 рода – Tmesipteris (10 видов), Psilotum (2 вида).
Растения

Отдел Плауновидные
типичные листостебельные растения
всегда есть корни
микрофиллия - мелкие листья
Отдел Плауновидные
типичные листостебельные растения
всегда есть корни
микрофиллия - мелкие листья

Листья плауновидных простые, реже слегка вильчато разветвленные на верхушке, с одной
Листья плауновидных простые, реже слегка вильчато разветвленные на верхушке, с одной
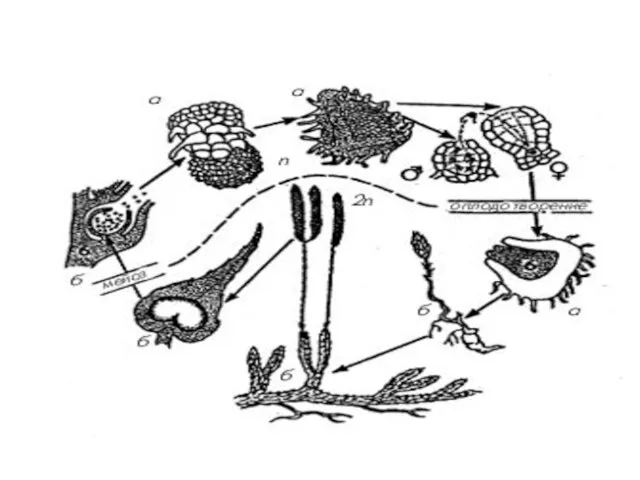

На верхней стороне спорофиллов располагаются спорангии. Зрелый спорангий имеет почковидную форму
На верхней стороне спорофиллов располагаются спорангии. Зрелый спорангий имеет почковидную форму

Попав на землю, спора потоками воды уносится в углубления (трещины) почвы,
Попав на землю, спора потоками воды уносится в углубления (трещины) почвы,

На верхней поверхности заростка образуются антеридии и архегонии. Антеридии целиком погружены
На верхней поверхности заростка образуются антеридии и архегонии. Антеридии целиком погружены



Хвощевидные
Современные хвощи — небольшие травянистые растения около 80 — 100 см
Хвощевидные
Современные хвощи — небольшие травянистые растения около 80 — 100 см

Надземные побеги хвоща полевого двух видов. Одни из них вегетативные, зеленые,
Надземные побеги хвоща полевого двух видов. Одни из них вегетативные, зеленые,

Однако для многих видов (например, Е. arvense) экспериментально установлено, что, поливая
Однако для многих видов (например, Е. arvense) экспериментально установлено, что, поливая

У всех видов хвоща стебли обладают выраженной метамерией, то есть правильным
У всех видов хвоща стебли обладают выраженной метамерией, то есть правильным

Вторичноводные растения.
Эволюция наземных растений.
Лекция № 6
Вторичноводные растения.
Эволюция наземных растений.
Лекция № 6

Отдел Папоротникообразные
Папоротники относятся к числу наиболее древних групп высших растений.
Отдел Папоротникообразные
Папоротники относятся к числу наиболее древних групп высших растений.

Вегетативные органы папоротникообразных
Среди современных папоротников преобладают травянистые формы с крупными листьями
Вегетативные органы папоротникообразных
Среди современных папоротников преобладают травянистые формы с крупными листьями

Вегетативные органы папоротникообразных
По сравнению со мхами, папоротники значительно лучше приспособлены к
Вегетативные органы папоротникообразных
По сравнению со мхами, папоротники значительно лучше приспособлены к

Вегетативные органы папоротникообразных
Крупные листья – вайи.
В отличие от обычных листьев,
Вегетативные органы папоротникообразных
Крупные листья – вайи.
В отличие от обычных листьев,

Генеративные органы папоротников
На нижней стороне листовых пластинок появляются скопления спорангиев, именуемые
Генеративные органы папоротников
На нижней стороне листовых пластинок появляются скопления спорангиев, именуемые

Генеративные органы папоротников
В спорангиях из диплоидной спорогенной ткани в результате мейоза
Генеративные органы папоротников
В спорангиях из диплоидной спорогенной ткани в результате мейоза


Королевский папоротник
Королевский папоротник

Вайи
Вайи

Генеративные органы папоротников
Генеративные органы папоротников


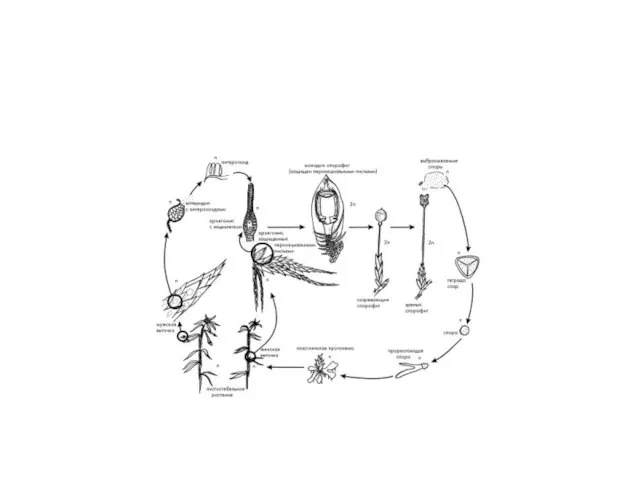
Жизненный цикл
Попав в благоприятные условия (влажная почва) спора прорастает в небольшую
Жизненный цикл
Попав в благоприятные условия (влажная почва) спора прорастает в небольшую

Жизненный цикл
Под пластинкой заростка скапливается испаряющаяся из почвы влага, поэтому вышедшие
Жизненный цикл
Под пластинкой заростка скапливается испаряющаяся из почвы влага, поэтому вышедшие


Водные папоротники
Водные папоротники

Водные папоротники
К порядку Сальвиниевые — водных папоротников — относится ряд приспособившихся к обитанию
Водные папоротники
К порядку Сальвиниевые — водных папоротников — относится ряд приспособившихся к обитанию

Водные папоротники
Наиболее распространенные роды сальвиниевых — Марсилия и Сальвиния. Первая представляет собой
Водные папоротники
Наиболее распространенные роды сальвиниевых — Марсилия и Сальвиния. Первая представляет собой
 Особенности Розоцветных Розоцветные (лат. Rosáles) — порядок двудольных растений, состоящий из девяти семейств, самым типичным из ко
Особенности Розоцветных Розоцветные (лат. Rosáles) — порядок двудольных растений, состоящий из девяти семейств, самым типичным из ко Кровообращение
Кровообращение Rosengewächse (Pfirsich)
Rosengewächse (Pfirsich) Зоология. Тесты
Зоология. Тесты Транспорт газов кровью. Регуляция дыхания
Транспорт газов кровью. Регуляция дыхания Взаимоотношение микро- и макро- организма. Понятие инфекция
Взаимоотношение микро- и макро- организма. Понятие инфекция Селекция животных
Селекция животных  Применение_эфирных_масел_в_составе_лекарственных_средств
Применение_эфирных_масел_в_составе_лекарственных_средств Зертханалық жұмыс. Сабақтың тақырыбы: Өсімдік түрлерін анықтау Өткізілу күні: 16.01.2013 Өткізілу уақыты: Өткізілу орны:Ақсу қаласын
Зертханалық жұмыс. Сабақтың тақырыбы: Өсімдік түрлерін анықтау Өткізілу күні: 16.01.2013 Өткізілу уақыты: Өткізілу орны:Ақсу қаласын Особенностей строения паразитов на всех стадиях развития
Особенностей строения паразитов на всех стадиях развития Презентация на тему "Влияние минеральных удобрений на рост пеларгонии зональной" - скачать презентации по Биологии
Презентация на тему "Влияние минеральных удобрений на рост пеларгонии зональной" - скачать презентации по Биологии Презентация по биологии Мозг человека
Презентация по биологии Мозг человека Сенсорная Система
Сенсорная Система Хлороводород. Соляная кислота.
Хлороводород. Соляная кислота. Иммунная система организма Урок-презентация для учащихся 9 класса. Составитель: учитель естествознания ГСКОУ № 46 «Центр Рим»
Иммунная система организма Урок-презентация для учащихся 9 класса. Составитель: учитель естествознания ГСКОУ № 46 «Центр Рим»  ПЛАСТИКОВАЯ БОМБА ? Куда мы идём? К прогрессу? Тогда почему привычным стал пейзаж, украшенный разорванными пластиковыми пакетами и
ПЛАСТИКОВАЯ БОМБА ? Куда мы идём? К прогрессу? Тогда почему привычным стал пейзаж, украшенный разорванными пластиковыми пакетами и Деление клетки. Митоз
Деление клетки. Митоз Растительный мир Восточной Сибири и Дальнего Востока
Растительный мир Восточной Сибири и Дальнего Востока Презентация на тему "Барсук" - скачать презентации по Биологии
Презентация на тему "Барсук" - скачать презентации по Биологии Презентация на тему "Путешествие в мир Простейших" - скачать презентации по Биологии
Презентация на тему "Путешествие в мир Простейших" - скачать презентации по Биологии ЦАРСТВО ПРОКАРИОТЫ ПОДЦАРСТВО БАКТЕРИИ
ЦАРСТВО ПРОКАРИОТЫ ПОДЦАРСТВО БАКТЕРИИ О пользе питьевой воды
О пользе питьевой воды Селекция
Селекция Види кровотеч
Види кровотеч  Метаболизм углеводов
Метаболизм углеводов Сельскохозяйственные животные
Сельскохозяйственные животные Особенности внутреннего строения и размножения птиц
Особенности внутреннего строения и размножения птиц Презентация на тему МЫШЦЫ МЫШЕЧНАЯ СИСТЕМА
Презентация на тему МЫШЦЫ МЫШЕЧНАЯ СИСТЕМА