- Транспорт газов кровью. Регуляция дыхания

Содержание
- 2. Транспорт газов кровью гемоглобин, его формы кривая диссоциации гемоглобина транспорт углекислого газа газообмен между кровью и
- 3. 1. ТРАНСПОРТ ГАЗОВ КРОВЬЮ
- 4. Физическое растворение О2 и СО2 подчиняется закону Генри, количество растворенного в жидкости газа пропорционально его парциальному
- 5. Гемоглобин – сложный белок класса хромопротеинов: простетическая группа – гем – порфириновое ядро содержит железо (Fe++)
- 6. Гемоглобин А (взрослого): 4 полипептидных субъединицы (2 α и 2 β), 4 гема Структура Нв –
- 7. Формы гемоглобина Гемоглобин плода (HbF): 2 γ-цепи вместо β имеет большее сродство к O2, чем дефинитивные
- 8. Формы гемоглобина крови Оксигемоглобин (HbO2) - форма переноса O2 к тканям оксигенированная кровь светло-красная, Дезоксигенированный Hb
- 9. Формы гемоглобина крови Метгемоглобин (MetHb) - содержит Fe3+; прочно связывает O2, диссоциация затруднена, метгемоглобиненмия м. б.
- 10. Аномальные формы гемоглобина Известно более 1000 мутаций разных глобинов, значительно изменяющих свойства Hb, в первую очередь
- 11. Серповидно-клеточная анемия (Африка) Гемоглобин S тенденция к полимеризации в деоксигенированном состоянии, причина серповидной формы Э. Во
- 12. Формы транспорта кислорода в крови ≈ 1.5% O2 в плазме в растворенном виде, ≈ 98.5% связано
- 13. Взаимосвязь между парциальным давлением О2 (РО2) и % HbO2 (насыщение, сатурация) описывается кривой диссоциации оксигемоглобина Насыщение
- 14. Пульсоксиметрия (оксигемометрия, гемоксиметрия) — неинвазивный метод определения степени насыщения крови кислородом. В основе –спектрофотометрический способ оценки
- 15. Факторы, влияющие на сродство гемоглобина к О2 pH, температура, 2,3-дифосфоглицерат (2,3-ДФГ). Индекс описывающий смещения кривой диссоциации
- 16. 2,3-Дифосфоглицерат (ДФГ) промежуточный продукт гликолиза в эритроцитах, взаимодействует с β‑субъединицей Hb, сдвигает кривую диссоциации Нв вправо,
- 17. Эффект Кристиана Бора (сдвиг кривой вправо) влияние CO2 и H+ на аффинитет О2 к Hb физиологическое
- 18. Факторы, влияющие на образование и диссоциацию НвО2
- 19. Влияние СО на связывание гемоглобином О2 крови Если построить кривую, отражающую половину кислородной емкости крови (что
- 20. гипербола на обратимое связывание миоглобина мышечной ткани и О2 влияет только PO2 миоглобин имеет очень высокое
- 21. Кислородная ёмкость крови максимальное возможное количество связанного с Hb О2 теоретически составляет 0,062 ммоль О2 (1,39
- 22. На 1 мм рт. ст. PO2 - 0.003 мл O2 растворяется в 100 мл крови при
- 23. Транспорт углекислого газа кровью
- 24. Транспорт углекислого газа кровью НСОз- с помощью белка-переносчика выводятся из Э. в плазму, а на их
- 25. Почти прямолинейная зависимость небольшая разница PCO2 артериальной и венозной крови (40 мм рт. ст. против 45
- 26. Диффузия О2 от тканевых капилляров к клеткам Артериальный конец капилляра Венозный конец капилляра
- 27. Захват СО2 кровью из тканей в тканевые капилляры Артериальный конец капилляра Венозный конец капилляра
- 28. 2. РЕГУЛЯЦИЯ ДЫХАНИЯ
- 29. Регуляция дыхания Система регуляции дыхания: рецепторы, воспринимающие информацию и передающие ее в дыхательный центр дыхательный центр/центральный
- 30. Центральный регулятор (дыхательные центры продолговатого мозга, варолиева моста и др.отделов ЦНС) Афферентные импульсы Эфферентные импульсы рецепторы
- 31. Центральная регуляция дыхания Группа дыхательных центров в стволе головного мозга → формируют автоматическую дыхательную активность. Основной
- 32. Ствол мозга продолговатый мозг чередование вдоха и выдоха варолиев мост – пневмотаксический центр – тонкая настройка
- 34. Рецепторы, участвующие в механизме регуляции дыхания центральные хеморецепторы, периферические хеморецепторы, рецепторы легких, прочие рецепторы носовой полости,
- 35. Центральные хеморецепторы мониторинг Рсо2, клетки-рецепторы - в продолговатом мозге. хеморецепторы ответ на отклонения в [Н+] и
- 36. Периферические хеморецепторы каротидные тельца – бифуркация сонной артерии, аортальные тельца – дуга аорты Два типа рецепторов:
- 37. Повышение PCO2 и снижение pH в артериальной крови и их влияние на уровень вентиляции
- 38. Стимуляция периферических хеморецепторов сниженным РО2
- 39. PO2 в норме мало влияет на вентиляцию, но при длительной гипоксемии (PO2 вентиляция в большей степени
- 40. При хронической гиперкапнии центральные хеморецепторы теряют чувствительность к CO2 при ХОБЛ - «гипоксический респираторный драйв» опосредован
- 41. Механорецепторы легких Ирритантные рецепторы – быстро адаптирующиеся рецепторы растяжения между эпителиальными клетками воздухоносных путей отвечают на
- 42. Рецепторы растяжения легких: медленно адаптирующиеся рецепторы растяжения (легочные механорецепторы вагуса) в ГМК воздухоносных путей, стимулируются растяжением
- 43. Юкстакапиллярные рецепторы: свободные нервные окончания в альвеолярной стенке вблизи легочных капилляров возбуждение – при ↑ кровенаполнения
- 44. Дополнительные дыхательные чувствительные структуры Рецепторы верхних воздухоносных путей: ответ на механические и химические стимулы, рефлекторный кашель,
- 45. Произвольный контроль Дыхания (истерия, возбуждение) Химические стимулы Нейрогенные стимулы Дыхательный центр стимулируется Каротидные и Аортальные тельца
- 46. Паттерны дыхания в норме и патологии В норме ЧД и глубина могут значительно изменяться. При сильном
- 48. Схематические представления активности дыхательного центра Импульсы, идущие через афферентные нейроны (темные красные линии), поступают к центральным
- 50. Скачать презентацию

Транспорт газов кровью
гемоглобин, его формы
кривая диссоциации гемоглобина
транспорт углекислого газа
газообмен между кровью
Транспорт газов кровью
гемоглобин, его формы
кривая диссоциации гемоглобина
транспорт углекислого газа
газообмен между кровью

1. ТРАНСПОРТ ГАЗОВ КРОВЬЮ
1. ТРАНСПОРТ ГАЗОВ КРОВЬЮ

Физическое растворение О2 и СО2 подчиняется закону Генри,
количество растворенного в жидкости газа
Физическое растворение О2 и СО2 подчиняется закону Генри,
количество растворенного в жидкости газа
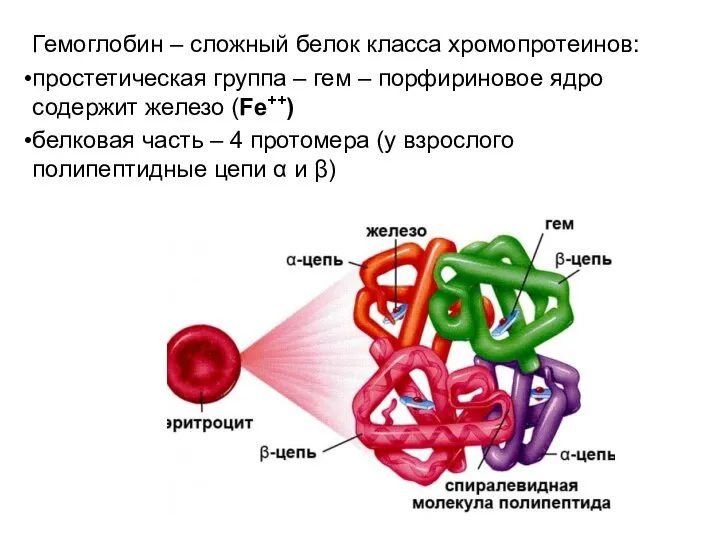
Гемоглобин – сложный белок класса хромопротеинов:
простетическая группа – гем – порфириновое
Гемоглобин – сложный белок класса хромопротеинов:
простетическая группа – гем – порфириновое
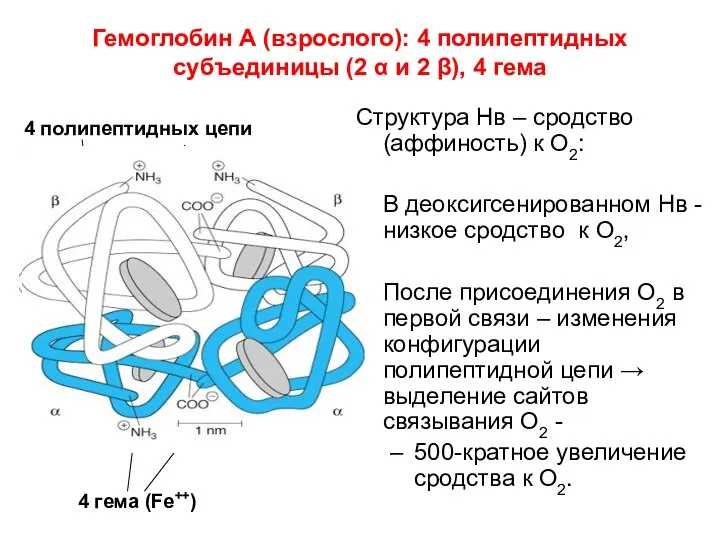
Гемоглобин А (взрослого): 4 полипептидных субъединицы (2 α и 2 β),
Гемоглобин А (взрослого): 4 полипептидных субъединицы (2 α и 2 β),
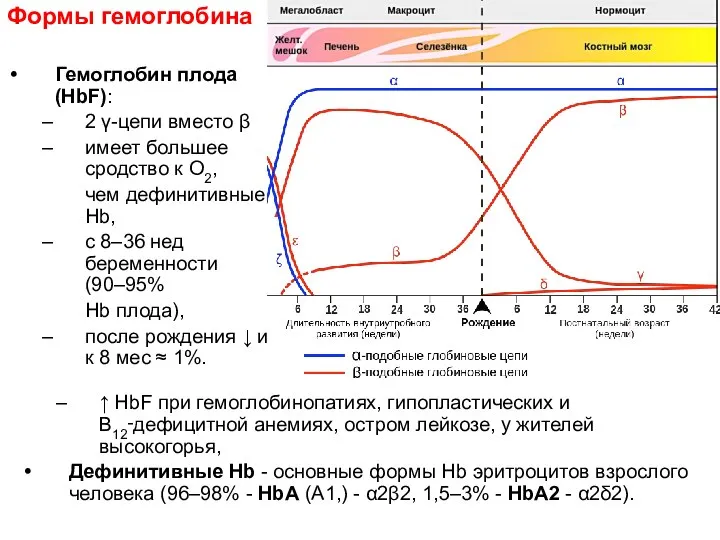
Формы гемоглобина
Гемоглобин плода (HbF):
2 γ-цепи вместо β
имеет большее сродство к
Формы гемоглобина
Гемоглобин плода (HbF):
2 γ-цепи вместо β
имеет большее сродство к

Формы гемоглобина крови
Оксигемоглобин (HbO2) - форма переноса O2 к тканям
оксигенированная кровь
Формы гемоглобина крови
Оксигемоглобин (HbO2) - форма переноса O2 к тканям
оксигенированная кровь

Формы гемоглобина крови
Метгемоглобин (MetHb) - содержит Fe3+; прочно связывает O2, диссоциация затруднена,
метгемоглобиненмия
Формы гемоглобина крови
Метгемоглобин (MetHb) - содержит Fe3+; прочно связывает O2, диссоциация затруднена,
метгемоглобиненмия

Аномальные формы гемоглобина
Известно более 1000 мутаций разных глобинов, значительно изменяющих свойства
Аномальные формы гемоглобина
Известно более 1000 мутаций разных глобинов, значительно изменяющих свойства
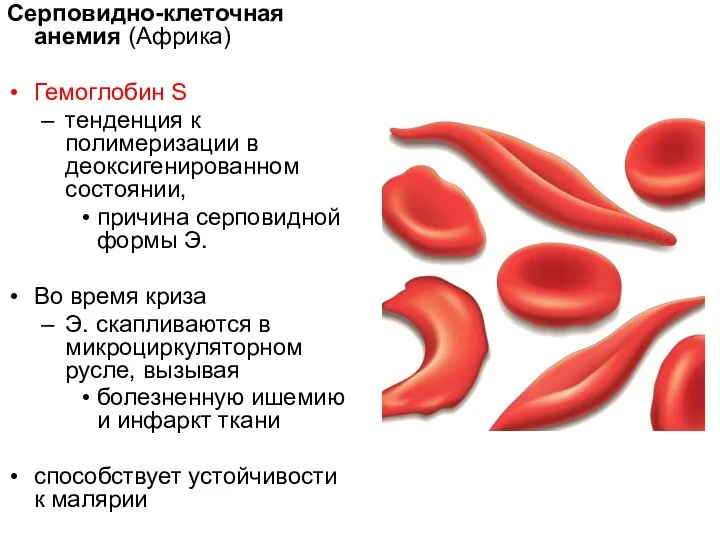
Серповидно-клеточная анемия (Африка)
Гемоглобин S
тенденция к полимеризации в деоксигенированном состоянии,
причина серповидной
Серповидно-клеточная анемия (Африка)
Гемоглобин S
тенденция к полимеризации в деоксигенированном состоянии,
причина серповидной
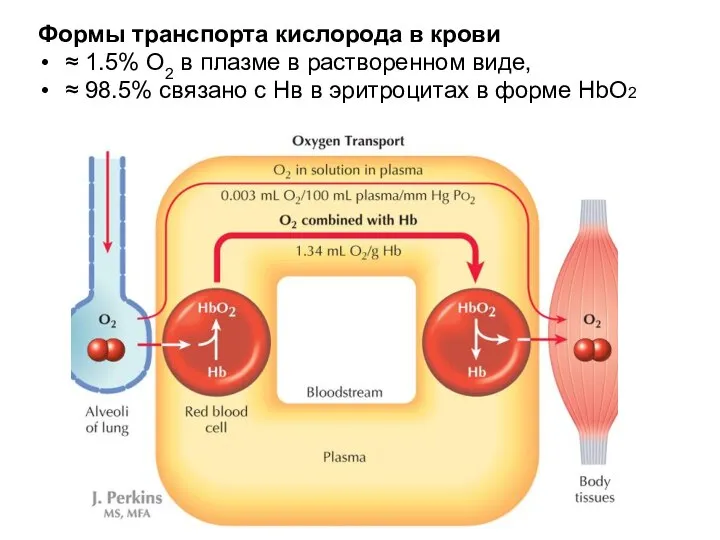
Формы транспорта кислорода в крови
≈ 1.5% O2 в плазме в растворенном
Формы транспорта кислорода в крови
≈ 1.5% O2 в плазме в растворенном

Взаимосвязь между парциальным давлением О2 (РО2) и % HbO2 (насыщение, сатурация)
Взаимосвязь между парциальным давлением О2 (РО2) и % HbO2 (насыщение, сатурация)

Пульсоксиметрия (оксигемометрия, гемоксиметрия) — неинвазивный метод определения степени насыщения крови кислородом.
В
Пульсоксиметрия (оксигемометрия, гемоксиметрия) — неинвазивный метод определения степени насыщения крови кислородом.
В

Факторы, влияющие на сродство гемоглобина к О2
pH,
температура,
2,3-дифосфоглицерат (2,3-ДФГ).
Индекс описывающий
Факторы, влияющие на сродство гемоглобина к О2
pH,
температура,
2,3-дифосфоглицерат (2,3-ДФГ).
Индекс описывающий

2,3-Дифосфоглицерат (ДФГ)
промежуточный продукт гликолиза в эритроцитах,
взаимодействует с β‑субъединицей Hb,
сдвигает кривую диссоциации
2,3-Дифосфоглицерат (ДФГ)
промежуточный продукт гликолиза в эритроцитах,
взаимодействует с β‑субъединицей Hb,
сдвигает кривую диссоциации
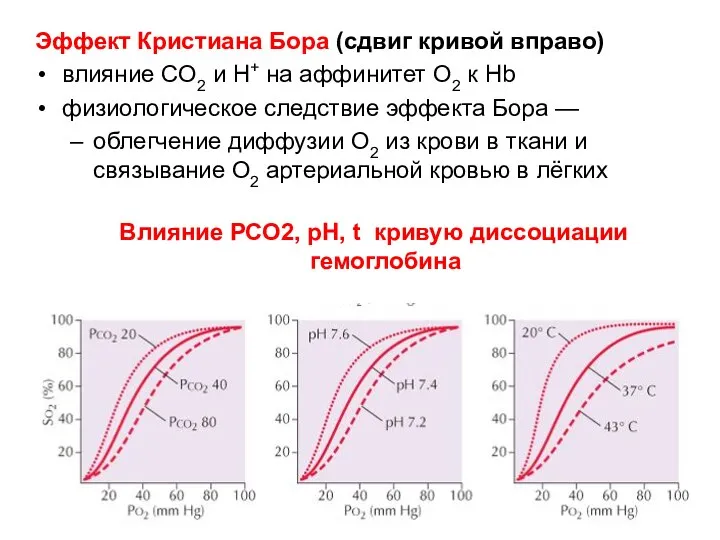
Эффект Кристиана Бора (сдвиг кривой вправо)
влияние CO2 и H+ на аффинитет
Эффект Кристиана Бора (сдвиг кривой вправо)
влияние CO2 и H+ на аффинитет
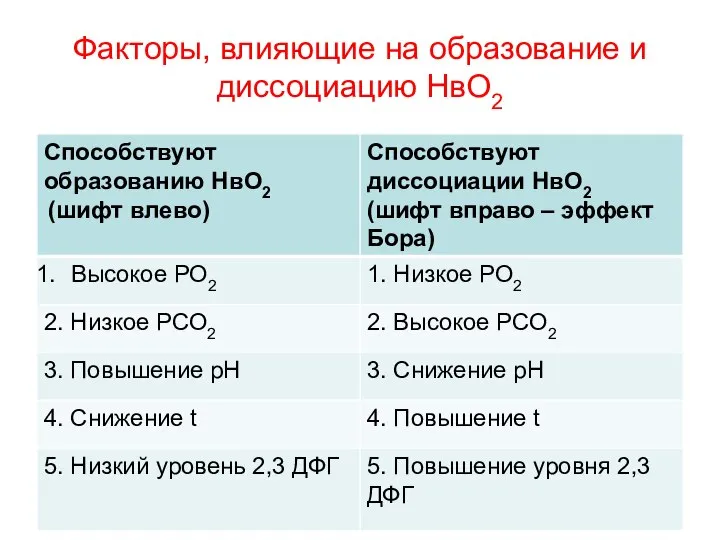
Факторы, влияющие на образование и диссоциацию НвО2
Факторы, влияющие на образование и диссоциацию НвО2

Влияние СО на связывание гемоглобином О2 крови
Если построить кривую, отражающую половину кислородной
Влияние СО на связывание гемоглобином О2 крови
Если построить кривую, отражающую половину кислородной

гипербола
на обратимое связывание миоглобина мышечной ткани и О2 влияет только PO2
миоглобин
гипербола
на обратимое связывание миоглобина мышечной ткани и О2 влияет только PO2
миоглобин

Кислородная ёмкость крови
максимальное возможное количество связанного с Hb О2
теоретически составляет 0,062
Кислородная ёмкость крови
максимальное возможное количество связанного с Hb О2
теоретически составляет 0,062

На 1 мм рт. ст. PO2 - 0.003 мл O2 растворяется
На 1 мм рт. ст. PO2 - 0.003 мл O2 растворяется
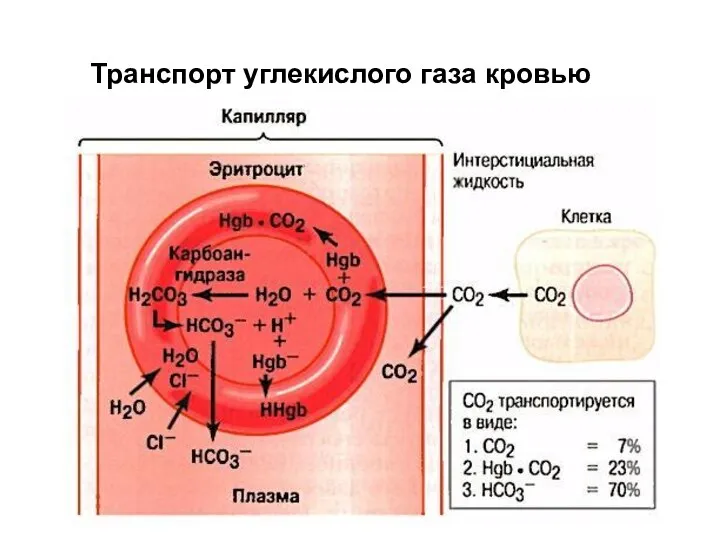
Транспорт углекислого газа кровью
Транспорт углекислого газа кровью

Транспорт углекислого газа кровью
НСОз- с помощью белка-переносчика выводятся из Э. в
Транспорт углекислого газа кровью
НСОз- с помощью белка-переносчика выводятся из Э. в
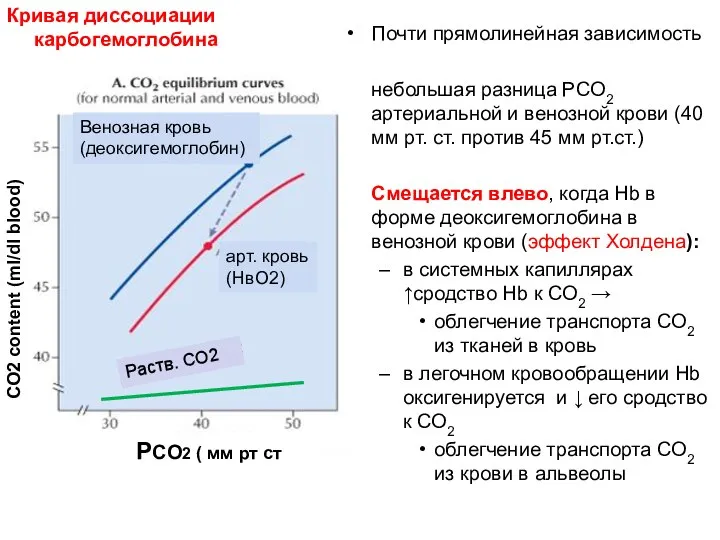
Почти прямолинейная зависимость
небольшая разница PCO2 артериальной и венозной крови (40 мм
Почти прямолинейная зависимость
небольшая разница PCO2 артериальной и венозной крови (40 мм
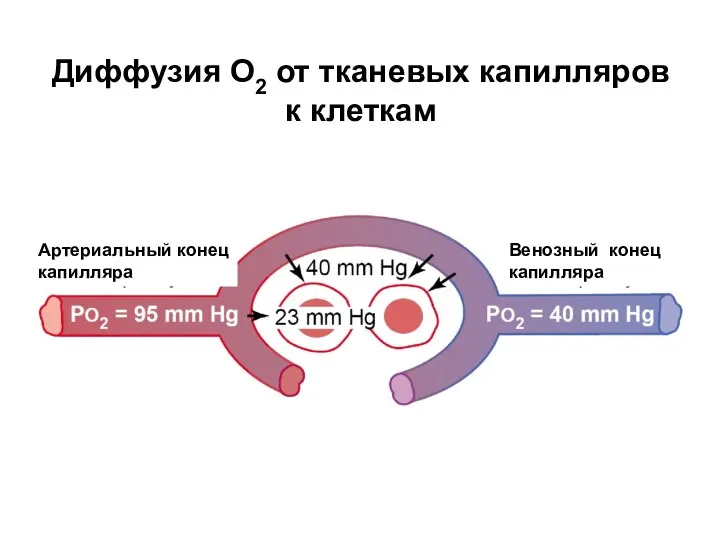
Диффузия О2 от тканевых капилляров к клеткам
Артериальный конец
капилляра
Венозный конец
капилляра
Диффузия О2 от тканевых капилляров к клеткам
Артериальный конец
капилляра
Венозный конец
капилляра

Захват СО2 кровью из тканей в тканевые капилляры
Артериальный конец
капилляра
Венозный конец
капилляра
Захват СО2 кровью из тканей в тканевые капилляры
Артериальный конец
капилляра
Венозный конец
капилляра

2. РЕГУЛЯЦИЯ ДЫХАНИЯ
2. РЕГУЛЯЦИЯ ДЫХАНИЯ

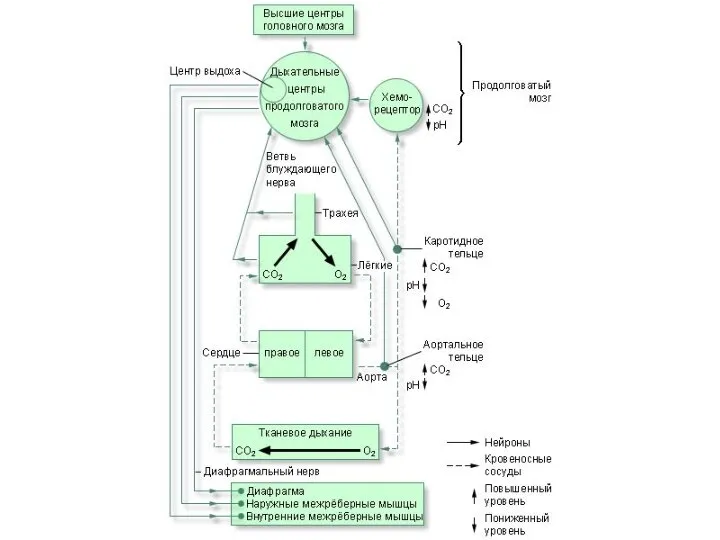
Регуляция дыхания
Система регуляции дыхания:
рецепторы, воспринимающие информацию и передающие ее в дыхательный
Регуляция дыхания
Система регуляции дыхания:
рецепторы, воспринимающие информацию и передающие ее в дыхательный

Центральный регулятор
(дыхательные центры продолговатого мозга, варолиева моста и др.отделов ЦНС)
Афферентные импульсы
Эфферентные
Центральный регулятор
(дыхательные центры продолговатого мозга, варолиева моста и др.отделов ЦНС)
Афферентные импульсы
Эфферентные
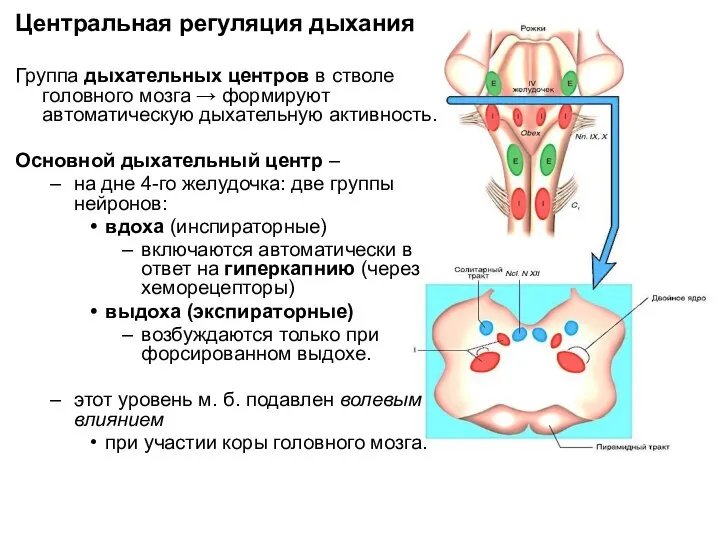
Центральная регуляция дыхания
Группа дыхательных центров в стволе головного мозга → формируют
Центральная регуляция дыхания
Группа дыхательных центров в стволе головного мозга → формируют
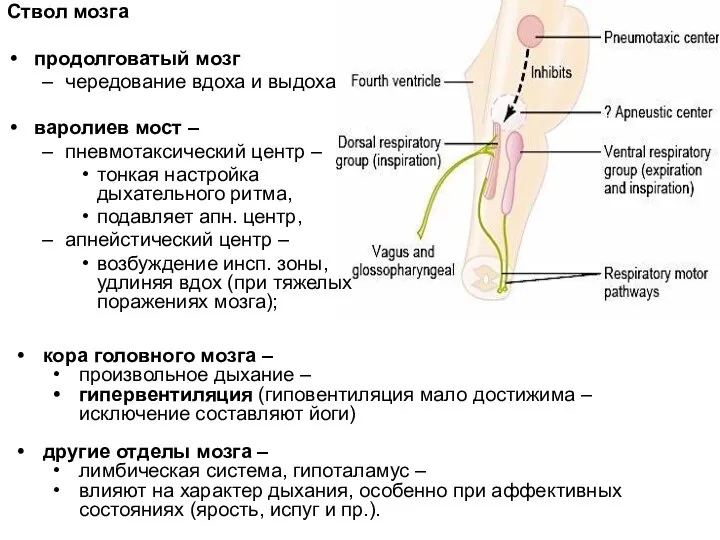
Ствол мозга
продолговатый мозг
чередование вдоха и выдоха
варолиев мост –
пневмотаксический
Ствол мозга
продолговатый мозг
чередование вдоха и выдоха
варолиев мост –
пневмотаксический

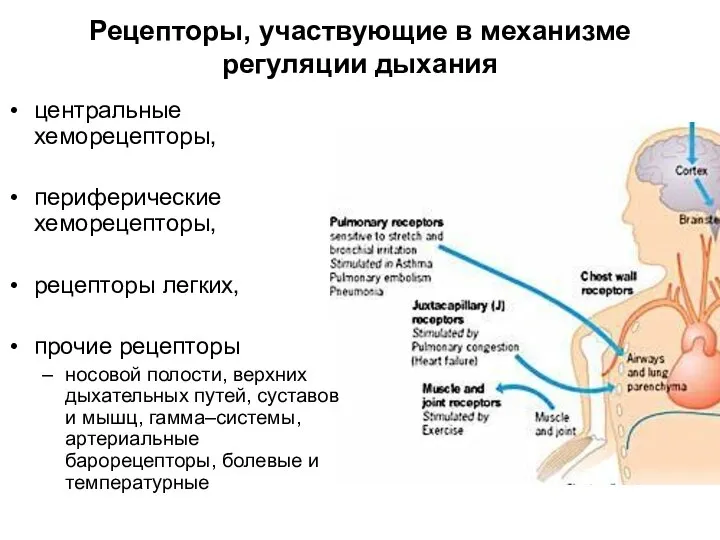
Рецепторы, участвующие в механизме регуляции дыхания
центральные хеморецепторы,
периферические хеморецепторы,
рецепторы легких,
прочие рецепторы
носовой
Рецепторы, участвующие в механизме регуляции дыхания
центральные хеморецепторы,
периферические хеморецепторы,
рецепторы легких,
прочие рецепторы
носовой
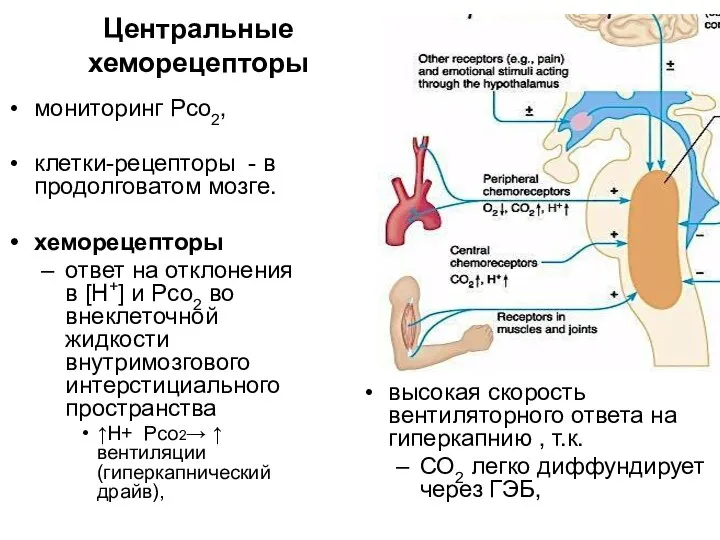
Центральные хеморецепторы
мониторинг Рсо2,
клетки-рецепторы - в продолговатом мозге.
хеморецепторы
ответ на отклонения в
Центральные хеморецепторы
мониторинг Рсо2,
клетки-рецепторы - в продолговатом мозге.
хеморецепторы
ответ на отклонения в

Периферические хеморецепторы
каротидные тельца – бифуркация сонной артерии,
аортальные тельца – дуга аорты
Два
Периферические хеморецепторы
каротидные тельца – бифуркация сонной артерии,
аортальные тельца – дуга аорты
Два
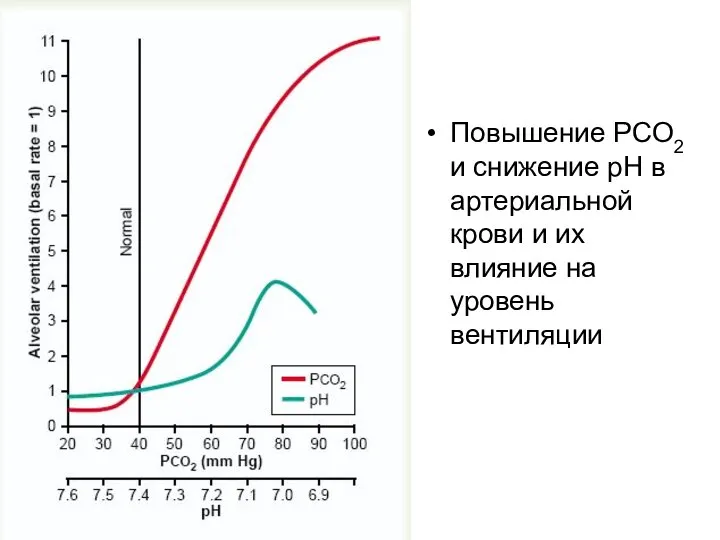
Повышение PCO2 и снижение pH в артериальной крови и их влияние
Повышение PCO2 и снижение pH в артериальной крови и их влияние

Стимуляция периферических хеморецепторов сниженным РО2
Стимуляция периферических хеморецепторов сниженным РО2
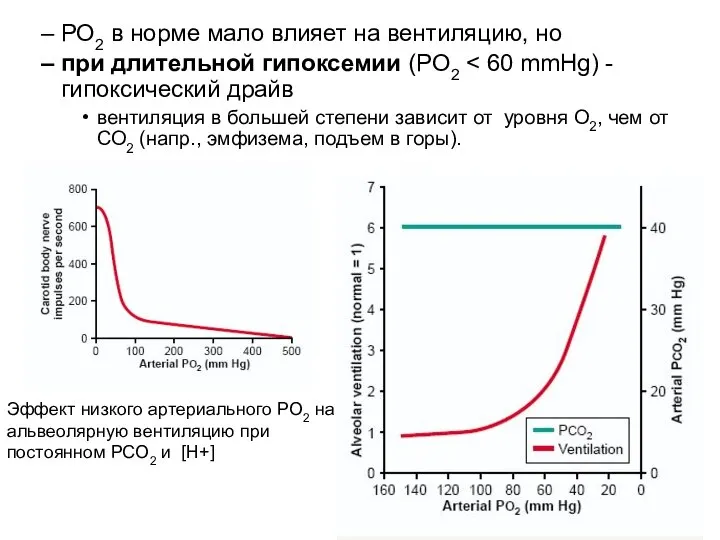
PO2 в норме мало влияет на вентиляцию, но
при длительной гипоксемии (PO2
PO2 в норме мало влияет на вентиляцию, но
при длительной гипоксемии (PO2
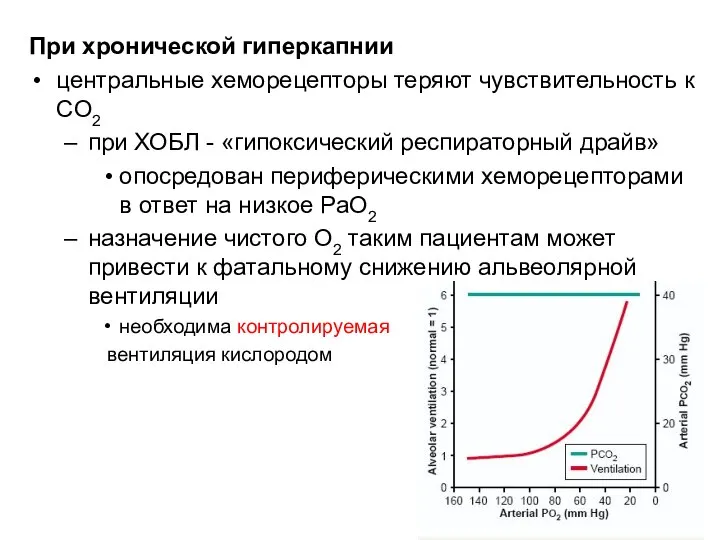
При хронической гиперкапнии
центральные хеморецепторы теряют чувствительность к CO2
при ХОБЛ -
При хронической гиперкапнии
центральные хеморецепторы теряют чувствительность к CO2
при ХОБЛ -
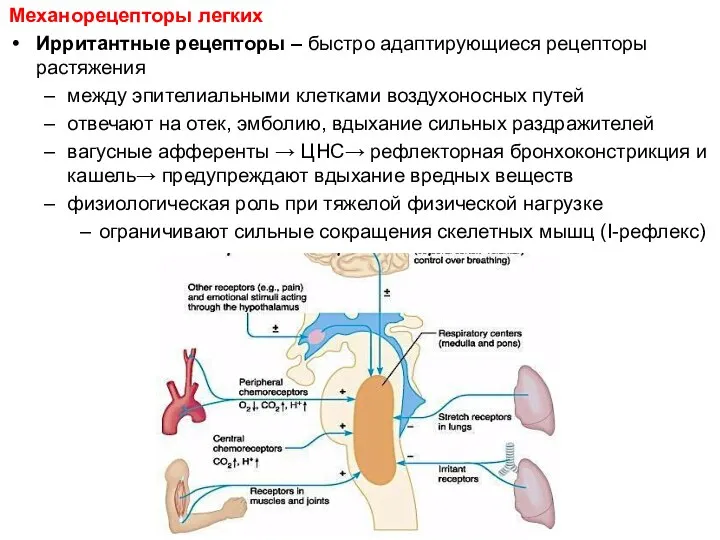
Механорецепторы легких
Ирритантные рецепторы – быстро адаптирующиеся рецепторы растяжения
между эпителиальными клетками воздухоносных
Механорецепторы легких
Ирритантные рецепторы – быстро адаптирующиеся рецепторы растяжения
между эпителиальными клетками воздухоносных
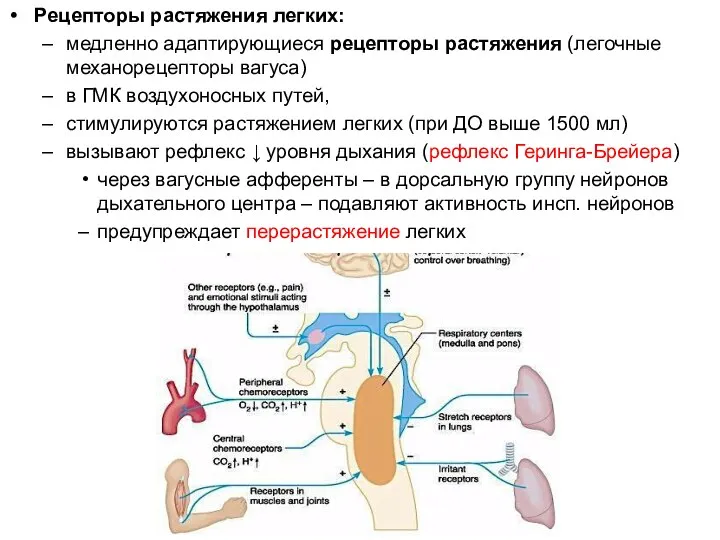
Рецепторы растяжения легких:
медленно адаптирующиеся рецепторы растяжения (легочные механорецепторы вагуса)
в ГМК воздухоносных
Рецепторы растяжения легких:
медленно адаптирующиеся рецепторы растяжения (легочные механорецепторы вагуса)
в ГМК воздухоносных
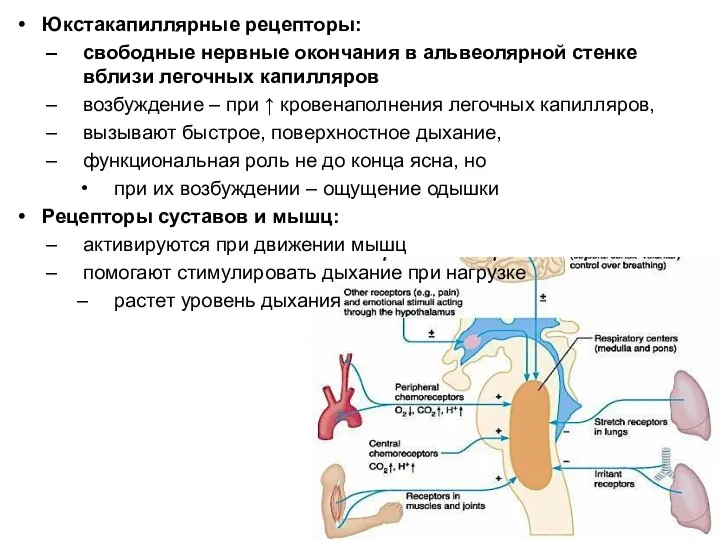
Юкстакапиллярные рецепторы:
свободные нервные окончания в альвеолярной стенке вблизи легочных капилляров
возбуждение
Юкстакапиллярные рецепторы:
свободные нервные окончания в альвеолярной стенке вблизи легочных капилляров
возбуждение

Дополнительные дыхательные чувствительные структуры
Рецепторы верхних воздухоносных путей:
ответ на механические и
Дополнительные дыхательные чувствительные структуры
Рецепторы верхних воздухоносных путей:
ответ на механические и
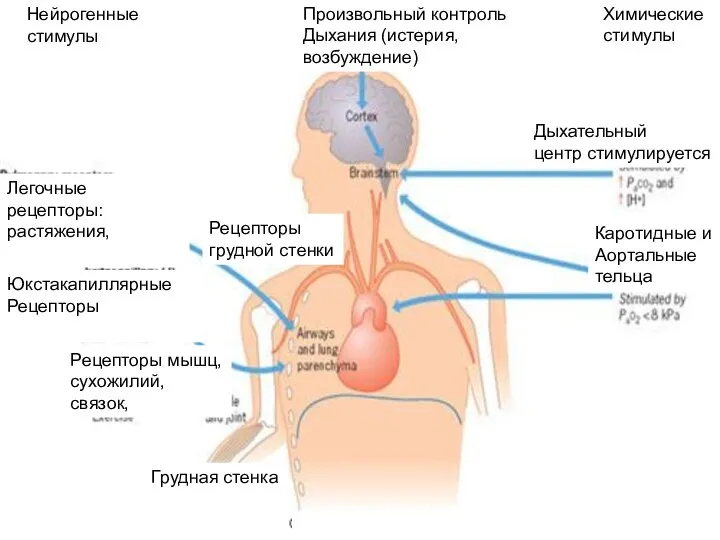
Произвольный контроль
Дыхания (истерия,
возбуждение)
Химические
стимулы
Нейрогенные
стимулы
Дыхательный
центр стимулируется
Каротидные и
Аортальные
тельца
Рецепторы
грудной стенки
Легочные рецепторы:
растяжения,
Юкстакапиллярные
Рецепторы
Рецепторы мышц,
сухожилий,
связок,
Грудная стенка
Произвольный контроль
Дыхания (истерия,
возбуждение)
Химические
стимулы
Нейрогенные
стимулы
Дыхательный
центр стимулируется
Каротидные и
Аортальные
тельца
Рецепторы
грудной стенки
Легочные рецепторы:
растяжения,
Юкстакапиллярные
Рецепторы
Рецепторы мышц,
сухожилий,
связок,
Грудная стенка
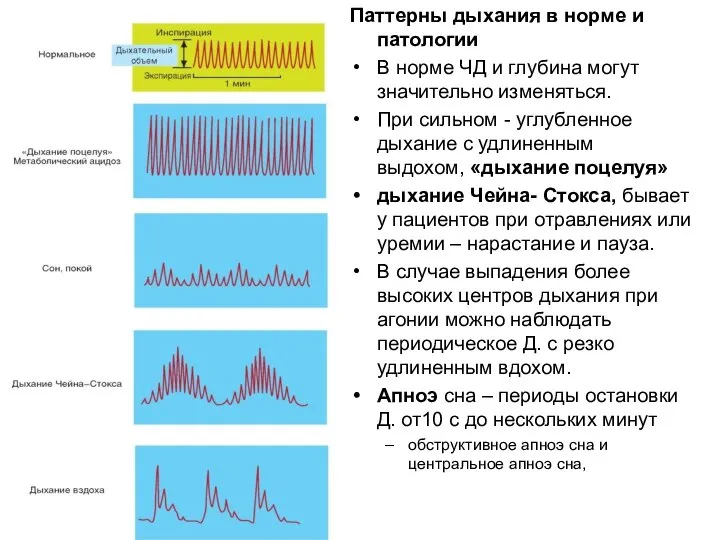
Паттерны дыхания в норме и патологии
В норме ЧД и глубина
Паттерны дыхания в норме и патологии
В норме ЧД и глубина
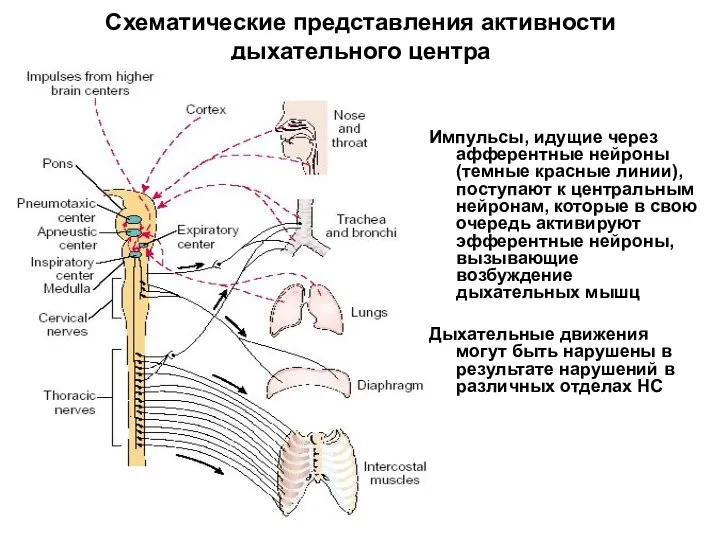
Схематические представления активности дыхательного центра
Импульсы, идущие через афферентные нейроны (темные красные
Схематические представления активности дыхательного центра
Импульсы, идущие через афферентные нейроны (темные красные
 Презентация на тему "Строение пищеварительной системы" - скачать бесплатно презентации по Биологии_
Презентация на тему "Строение пищеварительной системы" - скачать бесплатно презентации по Биологии_ Практическая работа №1. Анализ почвы пришкольного участка
Практическая работа №1. Анализ почвы пришкольного участка Удосконалення способів створення багатонасінних заплювачів буряка цукрового в умовах лісостепу України
Удосконалення способів створення багатонасінних заплювачів буряка цукрового в умовах лісостепу України Животные Крыма
Животные Крыма Нокардии
Нокардии Птица удод
Птица удод Общая анатомия развитие скелета. Кость как орган
Общая анатомия развитие скелета. Кость как орган Презентация к уроку «Значение растений в природе и жизни человека» 5 класс Лемешкина Ирина Евгеньевна, учитель биологии МБОУ Ле
Презентация к уроку «Значение растений в природе и жизни человека» 5 класс Лемешкина Ирина Евгеньевна, учитель биологии МБОУ Ле Характеристика Косуль
Характеристика Косуль Презентация на тему "Скелет Строение и Состав костей" - скачать бесплатно презентации по Биологии
Презентация на тему "Скелет Строение и Состав костей" - скачать бесплатно презентации по Биологии Задачи по экологической физиологии
Задачи по экологической физиологии ПРОЕКТНАЯ РАБОТА Тема: «Паукообразные: интересные и разные»
ПРОЕКТНАЯ РАБОТА Тема: «Паукообразные: интересные и разные» Производство чугуна и стали
Производство чугуна и стали Игры по лексической теме весна для детей 4-5 лет
Игры по лексической теме весна для детей 4-5 лет Фенотипическая или модификационная изменчивость
Фенотипическая или модификационная изменчивость Ядовитые грибы
Ядовитые грибы Социально-биологические основы физической культуры
Социально-биологические основы физической культуры Оомицеты Лутина Мария
Оомицеты Лутина Мария Кожно-мышечная чувствительность. Обоняние. Вкус. Презентация по биологии
Кожно-мышечная чувствительность. Обоняние. Вкус. Презентация по биологии Рептилии. Происхождение рептилий
Рептилии. Происхождение рептилий Изменчивость. Генные и хромосомные мутации
Изменчивость. Генные и хромосомные мутации Технические и санитарные средства борьбы с выделениями органической пыли и конидий грибов
Технические и санитарные средства борьбы с выделениями органической пыли и конидий грибов Морской скат
Морской скат Виды и разновидности фиалок
Виды и разновидности фиалок Социально-биологические основы физической культуры и здоровья
Социально-биологические основы физической культуры и здоровья Презентация на тему "Химический состав растений" - скачать презентации по Биологии
Презентация на тему "Химический состав растений" - скачать презентации по Биологии Кровоснабжение организма человека.
Кровоснабжение организма человека. Биологиялыќ белсенді заттар
Биологиялыќ белсенді заттар