- Фотосинтез. Вклад ученых в развитие учения о фотосинтезе

Содержание
- 2. Вклад ученых в развитие учения о фотосинтезе Джозеф Пристли ( 1771 г.) сделал открытие о роли
- 3. Сущность фотосинтеза Фотосинтез - это процесс преобразования световой энергии в потенциальную энергию химических связей с последующим
- 4. Фотосинтезирующие структуры мезосомы у бактерий хроматофоры и хлоропласты у водорослей хлоропласты (до 40 у высших растений
- 5. Строение хлоропласта Структурно - функциональной единицей хлоропласта является тилакоид (в одной гране от 10 до 20
- 6. Строение хлоропласта
- 7. Функции хлоропласта 1. Фотосинтез 2. Фосфорилирование (синтез АТФ и АДФ) 3. Синтез и гидролиз липидов 4.
- 8. Разновидности хлорофилла (известно около десяти) У всех высших растений содержатся хлорофиллы а (основной пигмент) и b
- 9. Строение хлорофилла В основе строения лежит Мg- порфирированный скелет. Порфириновая «головка» с атомами Мg-в центре (гидрофильна
- 10. Молекулы хлорофилла а и b Хлорофилл b отличается тем, что содержит на два атома водорода меньше
- 11. Оптический спектр поглощения хлорофиллов a и b Молекулы хлорофилла в основном поглощают красные (680нм) и частично
- 12. Строение квантосомы Схема строения квантосомы кукурузы (по Жиро): Хл — хлорофилл; Лп — липопротеидный комплекс; Пп
- 13. Фотосистема хлоропласта ФОТОСИСТЕМА – это совокупность фотосинтетических пигментов (около 250 – 400 молекул), погруженных в мембраны
- 14. Строение фотосистемы Состоит из: 1. Пигментов - сборщиков или антенн( хлорофиллы в, с, каротиноиды, фикобилины );
- 15. Схема фотосистемы
- 16. Строение фотосистемы
- 17. Фотосистемы - упакованные комплексы
- 18. Фотосистемы в мембране тилакоида
- 20. Спектры поглощения пигментов-сборщиков Энергия фотона света, поглощенная пигментами-сборщиками переносится на молекулу хлорофилла а (пигмент-ловушку)
- 21. Типы фотосинтеза АНАЭРОБНЫЙ ФОТОСИНТЕЗ: характерен для фотосинтезирующих бактерий; фотосинтезирующим пигментом у них является бактериохлорофилл; кислород при
- 22. Фотосинтез состоит из двух периодов: светового (энергетического), который идет на мембранах тилакоидов при участии хлорофилла, белков
- 23. Световой период делится на два этапа Фотофизический Поглощение кванта света пигментами -сборщиками, с последующим возбуждением электронов
- 24. Процессы светового периода А) циклическое фотофосфорилирование Б) нециклическое фотофосфорилирование (Z-схема) Фотофосфорилирование - это процесс присоединения к
- 25. Циклическое и нециклическое фотофосфорилирование
- 26. Циклическое фотофосфорилирование Фотон света попадает на ФС I и приводит к возбуждению пигментов- сборщиков, которое передается
- 27. Циклическое фотофосфорилирование С избытком энергии фотона света электрон двигается по электроннотранспортной цепи переходя от одного переносчика
- 28. Переносчики электронов Переносчики электронов локализованы в мембранах асимметрично. При этом последовательно чередуются переносчики электронов (цитохромы) с
- 31. Нециклическое фотофосфорилирование - однонаправленный поток электронов 1. Фотон света попадает на ФС I I и приводит
- 32. Цепь переноса электронов
- 33. Нециклическое фотофосфорилирование - фотолиз воды 3. Под действием фотона света вода, которая находится во внутреннем пространстве
- 34. Нециклическое фотофосфорилирование 4. Реакционный центр ФС I, получив избыток энергии от антенных пигментов переходит в возбужденное
- 35. Свет, попадая на молекулы хлорофилла, приводит их в возбужденное состояние. В результате этого электроны сходят со
- 36. Цикл Кальвина (С3–путь) Цикл восстановления СО2 до шести углеводородного соединения глюкозы Для образования глюкозы используются: СО2,
- 39. Карбоксилирование СО2 входит в цикл Кальвина и фиксируется на молекуле рибулозо-1,5-бисфосфата. ● Образуемое при этом соединение
- 40. Восстановление Молекулы 3-фосфоглицерата восстанавливаются до глицеральдегид-3-фосфата. ● Две молекулы глицеральдегид-3-фосфата используется для синтеза глюкозы и других
- 41. Регенерация Десять молекул глицеральдегид-3-фосфата включаются в ряд реакций пентозного цикла (5С3 → 3С5), в результате которого
- 42. Факторы, влияющие на фотосинтез 1. Свет - скорость фотосинтеза прямо пропорциональна интенсивности света (при большой интенсивности
- 44. Скачать презентацию

Вклад ученых в развитие учения о фотосинтезе
Джозеф Пристли ( 1771 г.) сделал
Вклад ученых в развитие учения о фотосинтезе
Джозеф Пристли ( 1771 г.) сделал

Сущность фотосинтеза
Фотосинтез - это процесс преобразования световой энергии в потенциальную
Сущность фотосинтеза
Фотосинтез - это процесс преобразования световой энергии в потенциальную

Фотосинтезирующие структуры
мезосомы у бактерий
хроматофоры и хлоропласты у водорослей
хлоропласты (до
Фотосинтезирующие структуры
мезосомы у бактерий
хроматофоры и хлоропласты у водорослей
хлоропласты (до

Строение хлоропласта
Структурно - функциональной единицей хлоропласта является тилакоид
(в одной гране
Строение хлоропласта
Структурно - функциональной единицей хлоропласта является тилакоид
(в одной гране

Строение хлоропласта
Строение хлоропласта

Функции хлоропласта
1. Фотосинтез
2. Фосфорилирование (синтез АТФ и АДФ)
3. Синтез и гидролиз
Функции хлоропласта
1. Фотосинтез
2. Фосфорилирование (синтез АТФ и АДФ)
3. Синтез и гидролиз

Разновидности хлорофилла
(известно около десяти)
У всех высших растений содержатся хлорофиллы а (основной
Разновидности хлорофилла
(известно около десяти)
У всех высших растений содержатся хлорофиллы а (основной

Строение хлорофилла
В основе строения лежит
Мg- порфирированный скелет. Порфириновая «головка» с
Строение хлорофилла
В основе строения лежит
Мg- порфирированный скелет. Порфириновая «головка» с

Молекулы хлорофилла а и b
Хлорофилл b отличается тем, что содержит на
Молекулы хлорофилла а и b
Хлорофилл b отличается тем, что содержит на

Оптический спектр поглощения хлорофиллов a и b
Молекулы хлорофилла в основном
Оптический спектр поглощения хлорофиллов a и b
Молекулы хлорофилла в основном

Строение квантосомы
Схема строения квантосомы кукурузы (по Жиро): Хл — хлорофилл; Лп
Строение квантосомы
Схема строения квантосомы кукурузы (по Жиро): Хл — хлорофилл; Лп

Фотосистема хлоропласта
ФОТОСИСТЕМА – это совокупность фотосинтетических пигментов (около 250 – 400
Фотосистема хлоропласта
ФОТОСИСТЕМА – это совокупность фотосинтетических пигментов (около 250 – 400

Строение фотосистемы
Состоит из:
1. Пигментов - сборщиков или антенн( хлорофиллы в,
Строение фотосистемы
Состоит из:
1. Пигментов - сборщиков или антенн( хлорофиллы в,

Схема фотосистемы
Схема фотосистемы

Строение фотосистемы
Строение фотосистемы

Фотосистемы - упакованные комплексы
Фотосистемы - упакованные комплексы
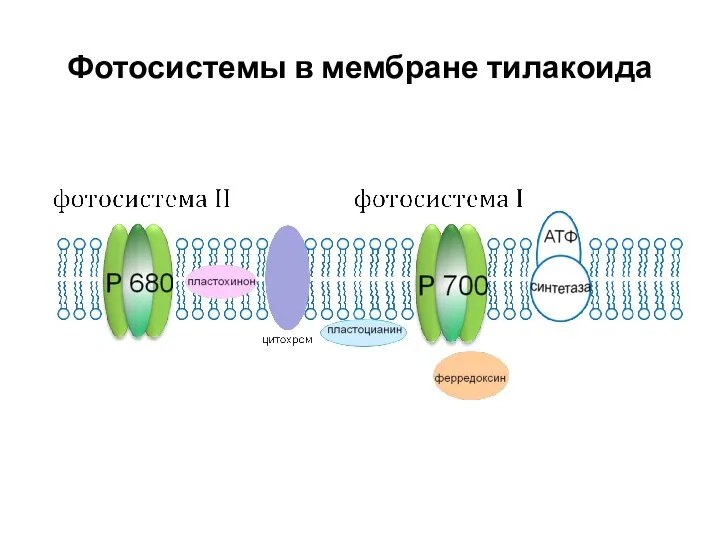
Фотосистемы в мембране тилакоида
Фотосистемы в мембране тилакоида

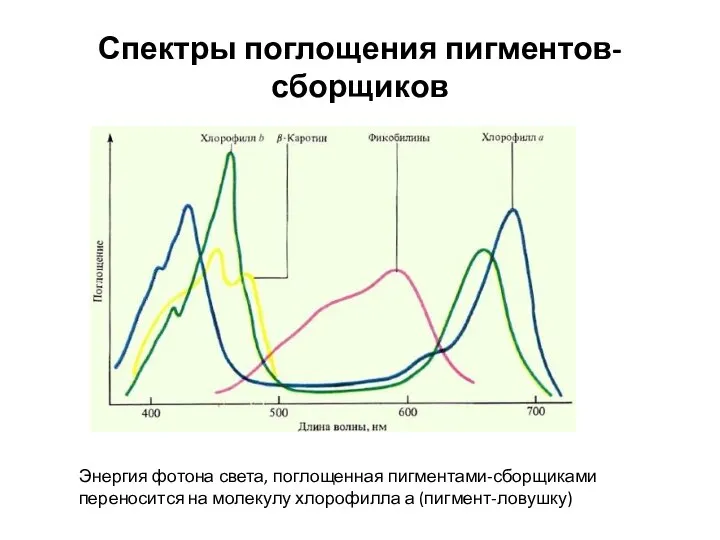
Спектры поглощения пигментов-сборщиков
Энергия фотона света, поглощенная пигментами-сборщиками переносится на молекулу хлорофилла
Спектры поглощения пигментов-сборщиков
Энергия фотона света, поглощенная пигментами-сборщиками переносится на молекулу хлорофилла

Типы фотосинтеза
АНАЭРОБНЫЙ ФОТОСИНТЕЗ:
характерен для фотосинтезирующих бактерий; фотосинтезирующим пигментом у них
Типы фотосинтеза
АНАЭРОБНЫЙ ФОТОСИНТЕЗ:
характерен для фотосинтезирующих бактерий; фотосинтезирующим пигментом у них

Фотосинтез состоит из двух периодов:
светового (энергетического), который идет на мембранах тилакоидов
Фотосинтез состоит из двух периодов:
светового (энергетического), который идет на мембранах тилакоидов

Световой период делится на два этапа
Фотофизический
Поглощение кванта света пигментами -сборщиками, с
Световой период делится на два этапа
Фотофизический
Поглощение кванта света пигментами -сборщиками, с

Процессы светового периода
А) циклическое фотофосфорилирование
Б) нециклическое фотофосфорилирование (Z-схема)
Фотофосфорилирование - это процесс
Процессы светового периода
А) циклическое фотофосфорилирование
Б) нециклическое фотофосфорилирование (Z-схема)
Фотофосфорилирование - это процесс

Циклическое и нециклическое фотофосфорилирование
Циклическое и нециклическое фотофосфорилирование

Циклическое фотофосфорилирование
Фотон света попадает на ФС I и приводит к возбуждению
Циклическое фотофосфорилирование
Фотон света попадает на ФС I и приводит к возбуждению

Циклическое фотофосфорилирование
С избытком энергии фотона света электрон двигается по электроннотранспортной цепи
Циклическое фотофосфорилирование
С избытком энергии фотона света электрон двигается по электроннотранспортной цепи

Переносчики электронов
Переносчики электронов локализованы в мембранах асимметрично. При этом последовательно чередуются
Переносчики электронов
Переносчики электронов локализованы в мембранах асимметрично. При этом последовательно чередуются



Нециклическое фотофосфорилирование - однонаправленный поток электронов
1. Фотон света попадает на ФС
Нециклическое фотофосфорилирование - однонаправленный поток электронов
1. Фотон света попадает на ФС
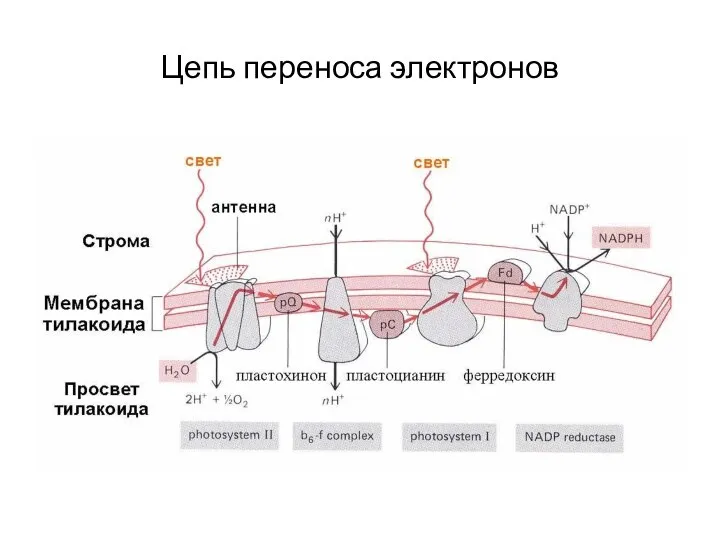
Цепь переноса электронов
Цепь переноса электронов

Нециклическое фотофосфорилирование -
фотолиз воды
3. Под действием фотона света вода, которая
Нециклическое фотофосфорилирование -
фотолиз воды
3. Под действием фотона света вода, которая

Нециклическое фотофосфорилирование
4. Реакционный центр ФС I, получив избыток энергии от антенных
Нециклическое фотофосфорилирование
4. Реакционный центр ФС I, получив избыток энергии от антенных
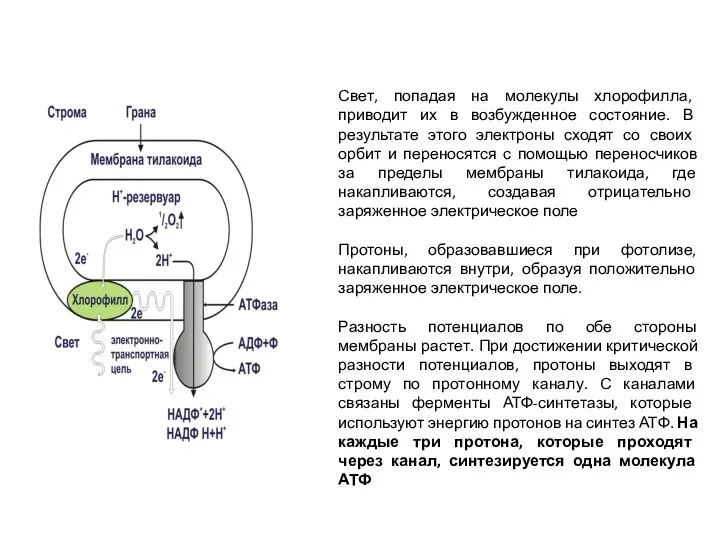
Свет, попадая на молекулы хлорофилла, приводит их в возбужденное состояние.
Свет, попадая на молекулы хлорофилла, приводит их в возбужденное состояние.

Цикл Кальвина (С3–путь)
Цикл восстановления СО2 до шести углеводородного соединения глюкозы
Для
Цикл Кальвина (С3–путь)
Цикл восстановления СО2 до шести углеводородного соединения глюкозы
Для



Карбоксилирование
СО2 входит в цикл Кальвина и фиксируется на молекуле рибулозо-1,5-бисфосфата.
Карбоксилирование
СО2 входит в цикл Кальвина и фиксируется на молекуле рибулозо-1,5-бисфосфата.

Восстановление
Молекулы 3-фосфоглицерата восстанавливаются до глицеральдегид-3-фосфата.
● Две молекулы глицеральдегид-3-фосфата
Восстановление
Молекулы 3-фосфоглицерата восстанавливаются до глицеральдегид-3-фосфата.
● Две молекулы глицеральдегид-3-фосфата

Регенерация
Десять молекул глицеральдегид-3-фосфата включаются в ряд реакций пентозного цикла (5С3
Регенерация
Десять молекул глицеральдегид-3-фосфата включаются в ряд реакций пентозного цикла (5С3

Факторы, влияющие на фотосинтез
1. Свет - скорость фотосинтеза прямо пропорциональна интенсивности
Факторы, влияющие на фотосинтез
1. Свет - скорость фотосинтеза прямо пропорциональна интенсивности
 Восприятие пространства и времени в спортивной деятельности как фактор адаптации организма человека к физическим нагрузкам
Восприятие пространства и времени в спортивной деятельности как фактор адаптации организма человека к физическим нагрузкам Методический комплекс элективного курса
Методический комплекс элективного курса Выращивание овощных культур
Выращивание овощных культур Необычное об обычной воде
Необычное об обычной воде Семейство кошачьих
Семейство кошачьих Обонятельный и вкусовой анализаторы
Обонятельный и вкусовой анализаторы Нервные растения
Нервные растения Соединительные ткани животных
Соединительные ткани животных Презентация на тему "Инфузория-туфелька" - скачать презентации по Биологии
Презентация на тему "Инфузория-туфелька" - скачать презентации по Биологии Презентация____
Презентация____ Генетические задачи. Дигибридное скрещивание. Практикум
Генетические задачи. Дигибридное скрещивание. Практикум Развитие артериальных жаберных дуг
Развитие артериальных жаберных дуг Презентация на тему Анатомия человека
Презентация на тему Анатомия человека  Как живут растения
Как живут растения Ферменты (энзимы)
Ферменты (энзимы) Строение клетки
Строение клетки Развитие жизни на Земле: архей и протерозой
Развитие жизни на Земле: архей и протерозой Презентация на тему Шляпочные
Презентация на тему Шляпочные  Газообмен в легких и тканях. Дыхательные движения
Газообмен в легких и тканях. Дыхательные движения  Значение живых организмов в природе
Значение живых организмов в природе Органы растений: корень, стебель и лист
Органы растений: корень, стебель и лист Практическое занятие №5 Тема: Биотические взаимоотношения организмов в биоценозе
Практическое занятие №5 Тема: Биотические взаимоотношения организмов в биоценозе  Мёд
Мёд Дыхание растений
Дыхание растений Презентация на тему " МЫ ЗА ЗДОРОВЫЙ ОБРАЗ ЖИЗНИ!" - скачать презентации по Биологии
Презентация на тему " МЫ ЗА ЗДОРОВЫЙ ОБРАЗ ЖИЗНИ!" - скачать презентации по Биологии Анатомия и физиология пищеварительной системы
Анатомия и физиология пищеварительной системы Домашние животные
Домашние животные Эффективная система омоложения. Пептидная косметика
Эффективная система омоложения. Пептидная косметика