- Функциональная биохимия тканей. Биохимия мышц и печени. (Тема 6)

Содержание
- 2. Строение мышц. Главные компоненты сократительных систем. Мышцы составляют около половины массы всего тела. Основная динамическая функция
- 3. Строение мышц. Главные компоненты сократительных систем. Саркомер включает пучок миозиновых нитей, серединой прикрепленные к М-пластине (М-линия),пучки
- 4. Механизм сокращения Миозин – белок миозиновых нитей содержит две идентичные цепи, скрученные между собой, N –концы
- 5. Механизм мышечного сокращения А. Са++ связывается с тропонином актиновых нитей и открывается в актине центр связывания
- 6. Мышечное сокращения. Условия. Сила сокращения зависит от количества миозиновых головок, включенных в работу, а значит, и
- 7. Источники энергии (АТФ)для мышечного сокращения. Мышца, работающая с максимальной активностью потребляет энергии в сотни раз больше,
- 8. Источники энергии для мышечного сокращения Креатинфосфат образуется из креатина и АТФ. Креатин - трипептид синтезируется в
- 9. Креатин, креатинин. Диагностическое значение. норма дистрофия мышц Мышцы Печень Мышцы Гли Креатин гли Креатин Арг креатин
- 10. Функциональная биохимия печени Печень занимает центральное место в обмене веществ, что определяется своеобразием топографии и кровоснабжением
- 11. Метаболическая и гомеостатическая функция Выполнение этой функции обусловлено участием печени в обмене углеводов, липидов, белков, пигментном
- 12. Роль печени в липидном обмене Печень участвует во всех этапах обмена липидов, включая переваривание и всасывание
- 13. Роль печени в белковом обмене. Около половины белков организма синтезируется в печени как для собственных нужд,
- 14. Желчеобразовательная и экскреторная функция. В печени образуются желчные кислоты из холестерола под действием фермента 7α-холестеролгидроксилазы. Активность
- 15. Обезвреживающая функция печени. В организме в процессе жизнедеятельности образуются токсичные метаболиты как собственных соединений, так и
- 16. Этап химической модификации Этап химической модификации обеспечивает повышение гидрофильности вещества и обязателен для всех гидрофобных соединеий.
- 17. Микросомальное окисление О2 SH НАДФН+ ФАД(ФМН)- цитохром редуктаза Р450 OH Н2О S Fe+2 Fe+3 2H+ 2Н+ê
- 18. Этап конъюгации Коньюгация с с гидрофильными молекулами: УДФ-глюкуроновая кислота, фосфоаденозинфосфосульфат (ФАФС) и др. Примеры: образование глюкуронида
- 20. Скачать презентацию

Строение мышц. Главные компоненты сократительных систем.
Мышцы составляют около половины массы всего
Строение мышц. Главные компоненты сократительных систем.
Мышцы составляют около половины массы всего
Строение мышц. Главные компоненты сократительных систем.
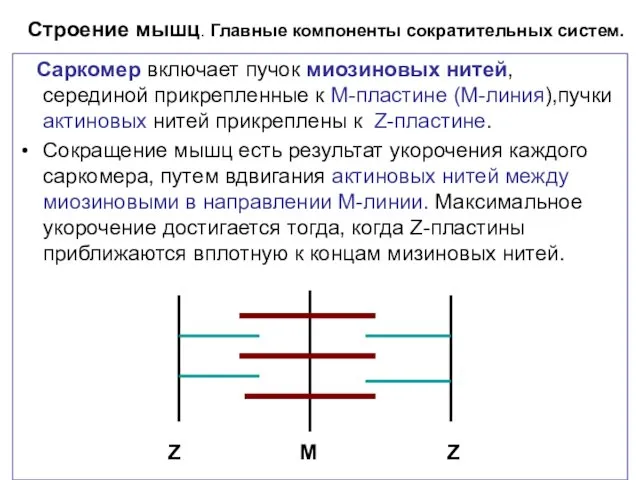
Саркомер включает пучок миозиновых нитей,
Строение мышц. Главные компоненты сократительных систем.
Саркомер включает пучок миозиновых нитей,

Механизм сокращения
Миозин – белок миозиновых нитей содержит две идентичные цепи, скрученные
Механизм сокращения
Миозин – белок миозиновых нитей содержит две идентичные цепи, скрученные
Механизм мышечного сокращения
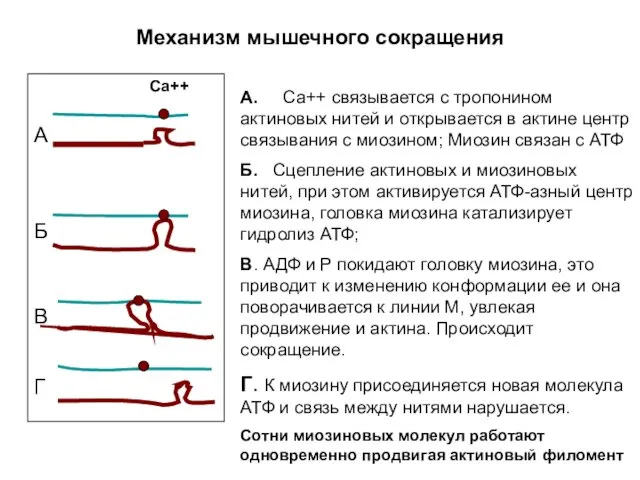
А. Са++ связывается с тропонином актиновых нитей и открывается
Механизм мышечного сокращения
А. Са++ связывается с тропонином актиновых нитей и открывается

Мышечное сокращения. Условия.
Сила сокращения зависит от количества миозиновых головок, включенных в
Мышечное сокращения. Условия.
Сила сокращения зависит от количества миозиновых головок, включенных в

Источники энергии (АТФ)для мышечного сокращения.
Мышца, работающая с максимальной активностью потребляет энергии
Источники энергии (АТФ)для мышечного сокращения.
Мышца, работающая с максимальной активностью потребляет энергии

Источники энергии для мышечного сокращения
Креатинфосфат образуется из креатина и АТФ. Креатин
Источники энергии для мышечного сокращения
Креатинфосфат образуется из креатина и АТФ. Креатин
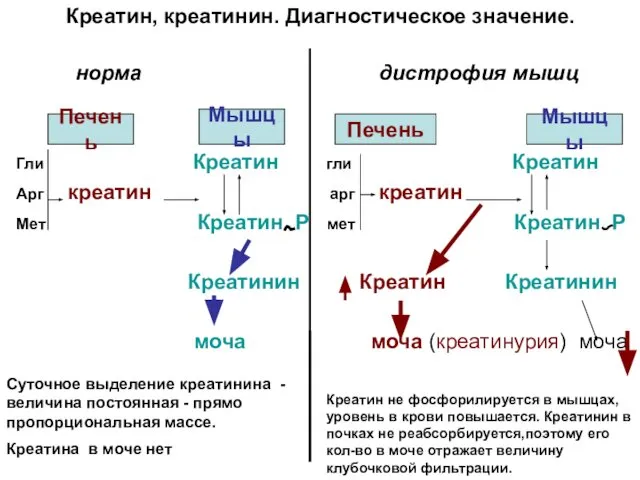
Креатин, креатинин. Диагностическое значение.
норма дистрофия мышц
Мышцы Печень Мышцы
Гли Креатин
Креатин, креатинин. Диагностическое значение.
норма дистрофия мышц
Мышцы Печень Мышцы
Гли Креатин

Функциональная биохимия печени
Печень занимает центральное место в обмене веществ, что определяется
Функциональная биохимия печени
Печень занимает центральное место в обмене веществ, что определяется

Метаболическая и гомеостатическая функция
Выполнение этой функции обусловлено участием печени в обмене
Метаболическая и гомеостатическая функция
Выполнение этой функции обусловлено участием печени в обмене

Роль печени в липидном обмене
Печень участвует во всех этапах обмена липидов,
Роль печени в липидном обмене
Печень участвует во всех этапах обмена липидов,

Роль печени в белковом обмене.
Около половины белков организма синтезируется в печени
Роль печени в белковом обмене.
Около половины белков организма синтезируется в печени

Желчеобразовательная и экскреторная функция.
В печени образуются желчные кислоты из холестерола под
Желчеобразовательная и экскреторная функция.
В печени образуются желчные кислоты из холестерола под

Обезвреживающая функция печени.
В организме в процессе жизнедеятельности образуются токсичные метаболиты как
Обезвреживающая функция печени.
В организме в процессе жизнедеятельности образуются токсичные метаболиты как

Этап химической модификации
Этап химической модификации обеспечивает повышение гидрофильности вещества и обязателен
Этап химической модификации
Этап химической модификации обеспечивает повышение гидрофильности вещества и обязателен
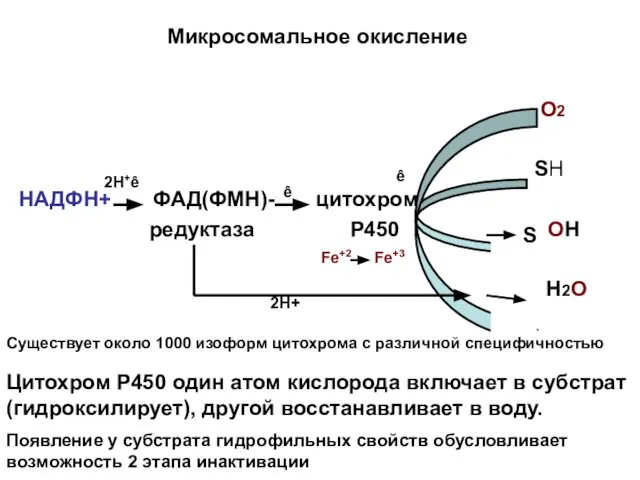
Микросомальное окисление
О2
SH
НАДФН+ ФАД(ФМН)- цитохром
редуктаза Р450 OH
Н2О
S
Fe+2 Fe+3
2H+
2Н+ê
ê
ê
Микросомальное окисление
О2
SH
НАДФН+ ФАД(ФМН)- цитохром
редуктаза Р450 OH
Н2О
S
Fe+2 Fe+3
2H+
2Н+ê
ê
ê

Этап конъюгации
Коньюгация с с гидрофильными молекулами:
УДФ-глюкуроновая кислота, фосфоаденозинфосфосульфат
Этап конъюгации
Коньюгация с с гидрофильными молекулами:
УДФ-глюкуроновая кислота, фосфоаденозинфосфосульфат
 Презентация на тему Естественный отбор – направляющий фактор эволюции.
Презентация на тему Естественный отбор – направляющий фактор эволюции.  Зоология - наука о животных
Зоология - наука о животных Жизненный цикл клетки
Жизненный цикл клетки Каракал
Каракал Игра-путешествие Наши зелёные друзья
Игра-путешествие Наши зелёные друзья Перелетные и зимующие птицы. Конспект занятий в старшей группе
Перелетные и зимующие птицы. Конспект занятий в старшей группе Ткани человека
Ткани человека Қолдан сұрыптау
Қолдан сұрыптау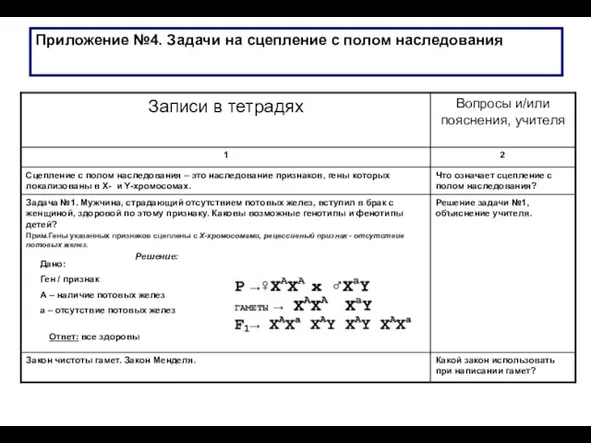 Задачи на сцепление с полом наследования. (Приложение 4)
Задачи на сцепление с полом наследования. (Приложение 4) Нәсілдердің қалыптасуы
Нәсілдердің қалыптасуы Человек
Человек Основные направления эволюции
Основные направления эволюции Плесневые грибы. Дрожжи
Плесневые грибы. Дрожжи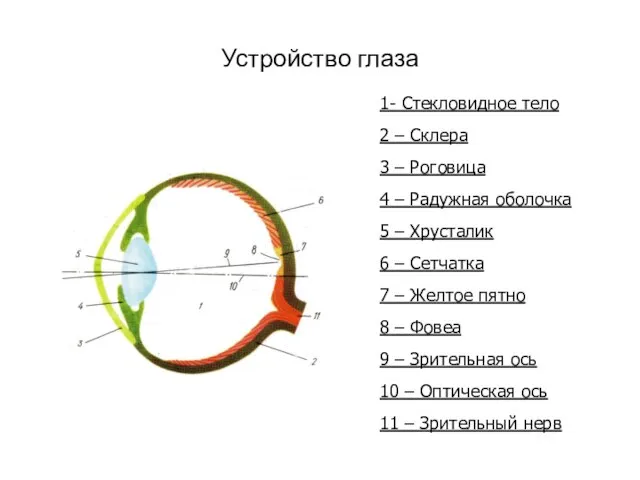 Устройство глаза
Устройство глаза Презентация на тему СТРУКТУРА И ФУНКЦИИ ХРОМОСОМ
Презентация на тему СТРУКТУРА И ФУНКЦИИ ХРОМОСОМ Общая вирусология. Бактериофаги
Общая вирусология. Бактериофаги Бесполое и половое размножение. Мужская и женская половые системы
Бесполое и половое размножение. Мужская и женская половые системы Электрофорез. Введение в теорию электрофореза
Электрофорез. Введение в теорию электрофореза Презентация на тему "Нарушения речи" - скачать бесплатно презентации по Биологии
Презентация на тему "Нарушения речи" - скачать бесплатно презентации по Биологии Презентация Природные ресурсы и охрана природы
Презентация Природные ресурсы и охрана природы Многоклеточные (Metazoa)
Многоклеточные (Metazoa) Викторина Зимушка-зима
Викторина Зимушка-зима Жидкое, а не вода, Белое, а не снег. Выполнила: Никулина Галина Ивановна Учитель начальных классов МОУ «СОШ №4» г.Энгельс
Жидкое, а не вода, Белое, а не снег. Выполнила: Никулина Галина Ивановна Учитель начальных классов МОУ «СОШ №4» г.Энгельс Ботаника. ОГЭ по биологии
Ботаника. ОГЭ по биологии Анатомо-физиологические особенности органа зрения у собак и кошек
Анатомо-физиологические особенности органа зрения у собак и кошек Стандартизация и оценка соответствия малины и ежевики
Стандартизация и оценка соответствия малины и ежевики Бронхи и лёгкие человека
Бронхи и лёгкие человека ГМО Генетически модифицированный организм
ГМО Генетически модифицированный организм