- Гендік экспрессияны бақылау

Содержание
- 2. Жасушалардың жетілуі негізінен жасуша геномындағы нуклеотид тізбегінің өзгеруімен емес, ген экспрессиясының өзгеруімен анықталады. Сүтқоректілердің нейрондары мен
- 3. Көпжасушалы ағзалардың жасушаларының әртүрлі типтерінің бір бірінен айырмашылықтары болады. Себебі белоктар мен РНҚ молекуласының әртүрлі жинағын
- 4. Дифференциацияланған жасушада тұтас ағзаның дамуына қажетті барлық генетикалық ақпараттың бар екендігін дәлелдейтін тәжірибелер
- 5. Өсімдіктің дифференциацияланған ұлпасының бөлігін жасанды ортада культивирлеген, содан кейін бұл ұлпаны жеке жасушаларға бөлген. Көптеген жағдайда
- 6. Дәл осы принцип ірі қара мал, қой, ешкі, ит, тышқандар сияқты сүтқоректілерге жасалынды. Бұл жағдайда соматикалық
- 7. Әртүрлі типтегі жасушаларда әртүрлі белок жинағы синтезделеді 1. Барлық жасушаға ортақ көптеген процестер бар және сәйкесінше
- 8. Адамның ісік жасушасының әртүрлі типтерінің арасындағы мРНҚ экспрессиясы кескініндегі айырмашылықтар
- 9. Сыртқы сигналдар жасушадағы ген экспрессиясының өзгеруін тудыруы мүмкін Егер бауыр жасушасына глюкокортикоидты(ГК), гормон әсер ететін болса,
- 10. Спектор бойынша экспрессияланатын белоктардың екі ұлпасының айырмашылығы
- 11. Ген экспрессиясы ДНҚ-дан РНҚ-ға және белокқа дейінгі жолдағы көптеген сатыларда реттеліне алады 1. Осы геннің транскрипция
- 12. Эукариот генінің экспрессиясын бақылаудың алты деңгейі
- 13. Ген транскрипциясының реттелуі мыналардың есебінен жүзеге асады 1) Белгілі бір тізбектің ДНҚ-сының қысқаучастоктарының есебінен; 2) осы
- 14. Ген экспрессиясын реттеуші белоктар спецификалық нуклеотидтік тізбектерді тануы тиіс. ДНҚ-ның бір тізбегін басқасынан айыру үшін қос
- 15. ДНҚ-ның қос спиралі
- 16. ДНҚ-дағы негіздердің әртүрлі жұптары ішкі жағы бойынша-қос спиральдың үзілуінсіз қалай танылуы мүмкін
- 17. ДНҚ-ны тану коды
- 18. Реттеуші белок ДНҚ-ның спецификалық тізбегін таниды, себебі белок беткейі белгілі бір дәрежеде қос спиральдың осы аймағының
- 19. Кейбір реттеуші белоктар және олар танитын ДНҚ тізбегі
- 20. Реттеуші белоктың ДНҚ-ның үлкен жырашығымен байланысуы
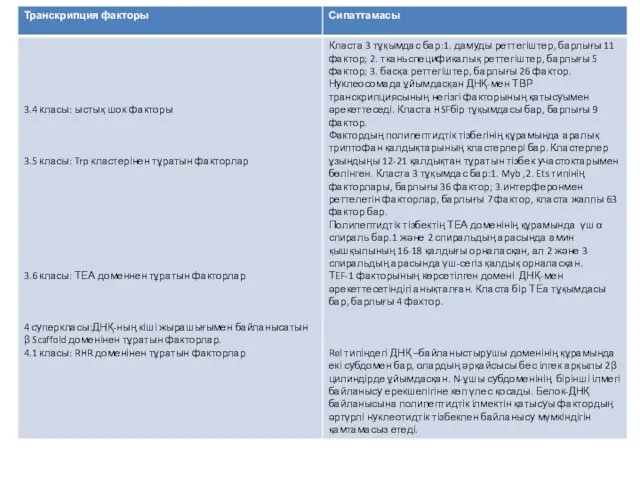
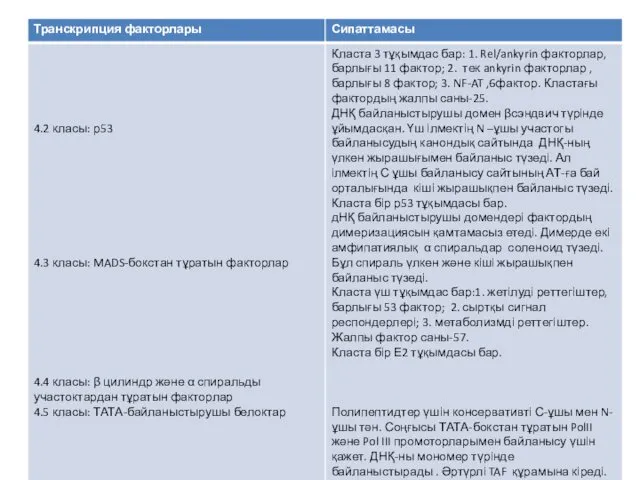
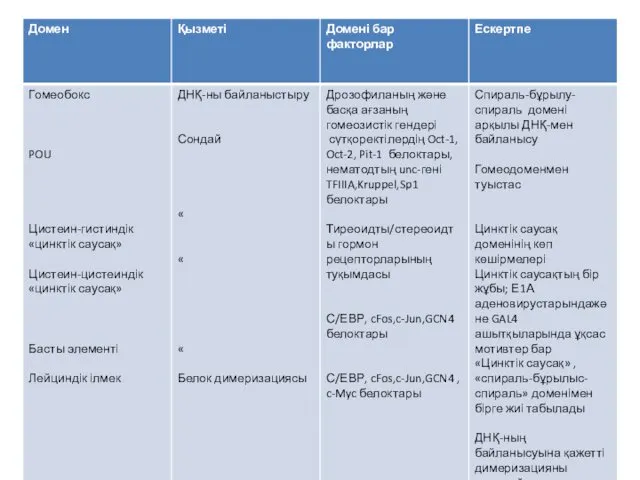
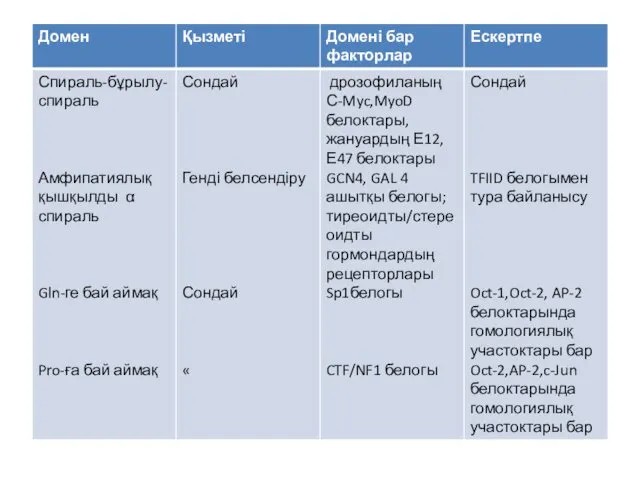
- 21. Бірнеше мыңдаған транскрипция факторлары белгілі. Олардың классификациясы гомологтарының бірнші реттік және екінші реттік құрылымын негізге алады.
- 22. Транскрипция факторының классификациясы
- 31. Транскрипция факторларының қызметі, механизмі үш саты арқылы жүреді; ДНҚ мен байланысу, Транскрипция барысына әсерету, Транскрипциялық факторлардың
- 32. Қазіргі уақытта, транскрипция факторларын жіктеудің негізінде, олардың әртүрлі типтерінен, бәріне ортақ полипептидтік тізбектерінің бірнеше құрылымдық элементтері
- 33. «мыршты саусақ» деп аталатын полипептидтік домендердің құрылымдық ерекшелігі және олардың ДНҚ молекуласымен әсерлесуі а – схемасы
- 34. «Мырышты саусақ» типінің бірі
- 35. «Мырышты саусақтан» тұратын белоктың ДНҚ-мен байланысуы
- 36. Жасушаішілік рецептор туыстығына жататын, ДНҚ-ның спецификалық тізбегімен байланысқан «мырышты саусақ» типті доменнің димері
- 37. Р53 белогы арқылы ДНҚ-ны тану
- 38. «мырышты саусақ» типті домендер көпеген транскрипця факторларында табылған; РНҚ-полимераза ІІ қызметін қамтамасыз ететін, сонымен қатар Sp1
- 39. Әртүрлі стероидтық кормондар рецепторларының ДНҚ-мен байланысушы орындарын зерттеу арқылы, олардың өзара гомологты екендігі, бірақ бірдей емес
- 40. Клюкокортикоидтар рецепторының мырышты саусағының N- соңының не бәрі екі аминқышқылының орын алмасуынан қалыптасқан (бұндай алмасу эстрогендер
- 41. “спираль-бұрылыс-спираль” мотивті транскрипция факторлары. ДНҚ молекуласындағы реттеуші арнаулы бөлігін танитын, басқаша пептидтік домендер, гомеотикалық (гомеозистық) гендердің
- 42. "спираль–бұрылыс–спираль" полипептидтық домендер типі құрылымы (а) және "лейцинды ілгек" (б) L – қалдық Leu
- 43. «Лейцинді ілгек» типті ДНК-байланысушы домендер. Leu қалдығының бұл құрылымында, фактордың α-спираль ды полипеитидті тізбегінде әрбір алты
- 44. ДНҚ-мен байланысатын спираль-бұрылыс-спираль мотиві
- 45. Спираль-бұрылыс-спираль мотивінен тұратын ДНҚ байланыстырушы белоктар
- 46. ДНҚ-ның спецификалық тізбегімен байланысқан гомеодомен
- 47. ДНҚ-мен байланысқан «лейциндік найзағай» димері
- 48. «Лейциндік молниядан» тұратын белоктардың гетеродимеризациясы ДНҚ-мен байланысудың спецификалық қасиетін өзгерте алады
- 49. Екі ДНҚ-байланыстырушы домен икемді полипептидпен ковалентті байланысқан
- 50. ДНҚ-мен байланысқан спираль-ілгек мотивінің димері
- 51. HLH-белокпен қиылған ингибиторлы реттелу
- 52. Белок-ДНҚ байланысының кең таралған титерінің бірі
- 53. Алты әртүрлі «цинктік саусақ» және ДНқ тізбегі арасындағы сайт-спецификалық байланыстардың жинақталған схемасы
- 54. Глюкокортикоидтар рецепторларының полипеитидті тізбегінің доменді құрылымы Қызыметті домендердің шек арасындағы цифрлар аминқышқылы қалдығы санын көрсетеді, олар
- 58. Транскрипцияның белсенуін генетикалық бақылау. Транскрипцияның көптеген факторларының біріншілік рөлі белгілі бір ұлпада гендердің белгілі бір тобының
- 59. Эукариодтарда транскрипция факторлары активтілігінің реттелу ерекшелігі (а–г)
- 60. Эукариот генінің транскрипциясын бақылау Эукариоттық РНҚ полимераза II үшін транскрипцияның 5 ортақ факторы қажет. Эукариоттық промотордағы
- 61. Геннің бақылаушы аймағы -ген транскрипциясының и нициациясын реттеуге қатысатын ДНҚ-ның бүкіл кесіндісі. Реттеуші тізбек – промотордағы
- 62. Әдеттегі эукариоттық геннің бақылаушы аймағы
- 63. Эукариоттық белок-активаторлар байланысқан ДНҚ участоктары бірінші энхансер деп аталған болатын. Алайда бұл белок активаторлар промоторға он
- 64. Белок активаторының модульдік құрылымы
- 65. Транскрипцияның ортақ факторлары, медиатор және РНҚ полимераза стандарттық нуклеосомаға жинақталған промоторда өздігінен жинала алмайды. Белок-активаторлар сонымен
- 66. Эукариоттық белок-активаторлары әртүрлі төрт әдістің көмегімен транскрипция инициациясын стимулдау үшін хроматин құрылымының Локальдық өзгрістер арқылы басқара
- 67. Алайда транскрипция кезіндегі гсистондардың коваленттік модификациясының өте маңызды рөлі хроматиннің құрылымының тура өзгермеуімен байланысты: бұл модификациялар
- 68. Транскрипция инициациясының басталуы кезінде гистондық кодты оқу және жазу
- 69. Реакцияның жылдамдығын арттыру үшін бірлесіп бірнеше факторлар әрекет еткен жерлердегі олардың бірлескен әсері оң әсерлердің қосындысы
- 70. Транскрипциялық синергизм
- 71. Белок-активаторлар транскрипцияның активациясына әкелетін жолдардың әртүрлі кезеңдеріне әсер ете алатын болғандықтан бұл кезеңдер үнемі тағайындалған тәртіп
- 72. Спецификалық геннің транскрипциясының инициациясына әкелетін Жағдайының жүру тәртібі
- 73. Жеке реттеуші белок реттеуші кешеннің бірнеше типіне қатыса алады. Мысалы, бір жағдайда транскрипцияны белсендіретін белок кешенінің
- 74. Эукариоттық белок-репрессорының мүмкін болатын алты әрекет ету әдісі
- 75. Эукариоттардың реттеуші белоктары үнемі ДНҚ-дағы кешенге жиналады
- 77. Скачать презентацию
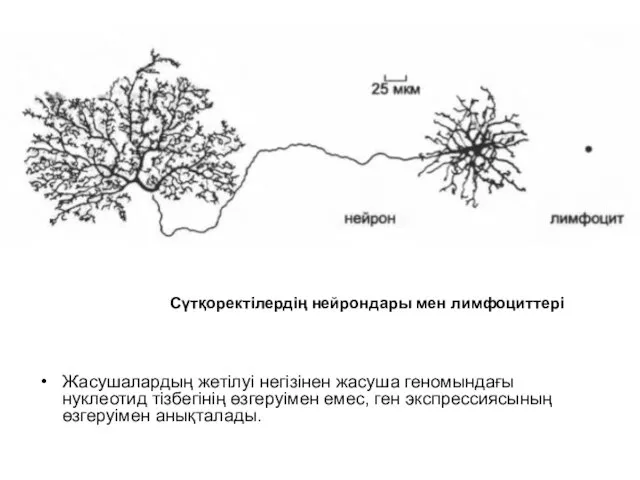
Жасушалардың жетілуі негізінен жасуша геномындағы нуклеотид тізбегінің өзгеруімен емес, ген экспрессиясының
Жасушалардың жетілуі негізінен жасуша геномындағы нуклеотид тізбегінің өзгеруімен емес, ген экспрессиясының

Көпжасушалы ағзалардың жасушаларының әртүрлі типтерінің бір бірінен айырмашылықтары болады. Себебі белоктар
Көпжасушалы ағзалардың жасушаларының әртүрлі типтерінің бір бірінен айырмашылықтары болады. Себебі белоктар
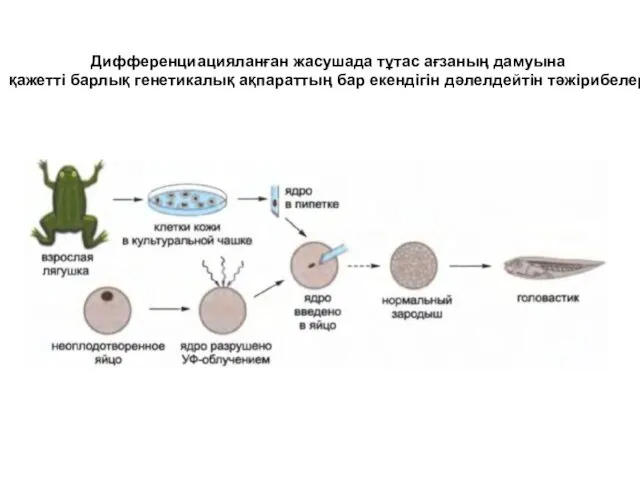
Дифференциацияланған жасушада тұтас ағзаның дамуына
қажетті барлық генетикалық ақпараттың бар екендігін
Дифференциацияланған жасушада тұтас ағзаның дамуына
қажетті барлық генетикалық ақпараттың бар екендігін
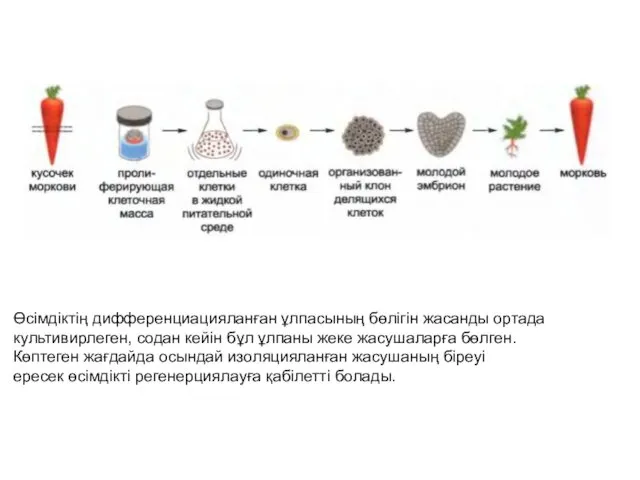
Өсімдіктің дифференциацияланған ұлпасының бөлігін жасанды ортада
культивирлеген, содан кейін бұл ұлпаны
Өсімдіктің дифференциацияланған ұлпасының бөлігін жасанды ортада
культивирлеген, содан кейін бұл ұлпаны
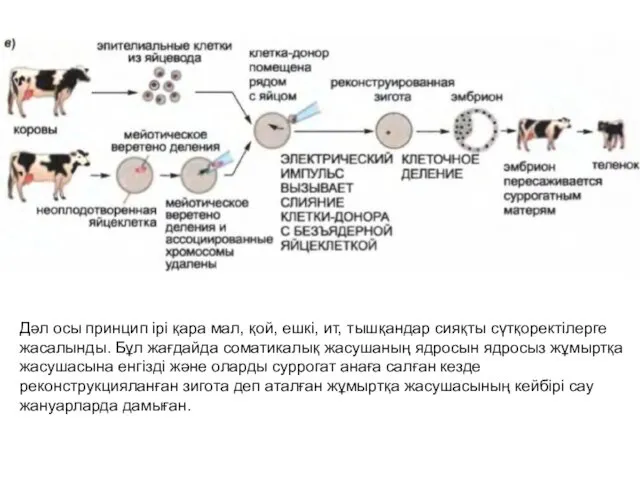
Дәл осы принцип ірі қара мал, қой, ешкі, ит, тышқандар сияқты
Дәл осы принцип ірі қара мал, қой, ешкі, ит, тышқандар сияқты

Әртүрлі типтегі жасушаларда әртүрлі белок жинағы синтезделеді
1. Барлық жасушаға ортақ көптеген
Әртүрлі типтегі жасушаларда әртүрлі белок жинағы синтезделеді
1. Барлық жасушаға ортақ көптеген
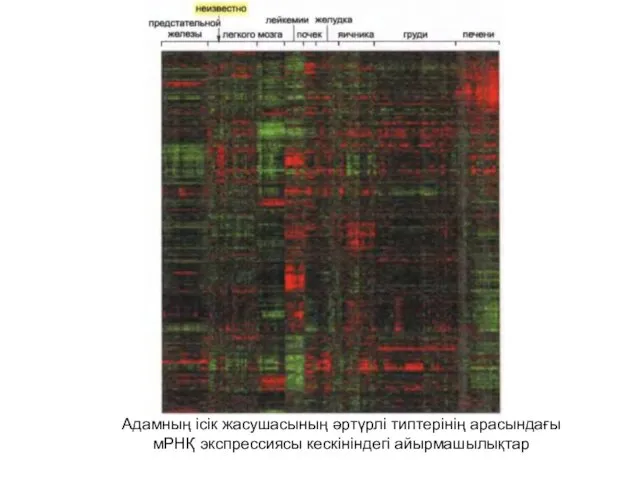
Адамның ісік жасушасының әртүрлі типтерінің арасындағы
мРНҚ экспрессиясы кескініндегі айырмашылықтар
Адамның ісік жасушасының әртүрлі типтерінің арасындағы
мРНҚ экспрессиясы кескініндегі айырмашылықтар

Сыртқы сигналдар жасушадағы ген экспрессиясының өзгеруін тудыруы мүмкін
Егер бауыр жасушасына глюкокортикоидты(ГК),
Сыртқы сигналдар жасушадағы ген экспрессиясының өзгеруін тудыруы мүмкін
Егер бауыр жасушасына глюкокортикоидты(ГК),

Спектор бойынша экспрессияланатын белоктардың
екі ұлпасының айырмашылығы
Спектор бойынша экспрессияланатын белоктардың
екі ұлпасының айырмашылығы

Ген экспрессиясы ДНҚ-дан РНҚ-ға және белокқа дейінгі жолдағы көптеген сатыларда реттеліне
Ген экспрессиясы ДНҚ-дан РНҚ-ға және белокқа дейінгі жолдағы көптеген сатыларда реттеліне
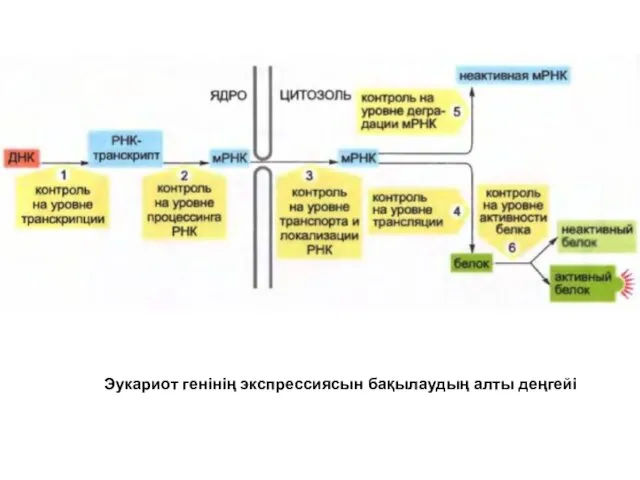
Эукариот генінің экспрессиясын бақылаудың алты деңгейі
Эукариот генінің экспрессиясын бақылаудың алты деңгейі

Ген транскрипциясының реттелуі мыналардың есебінен жүзеге асады
1) Белгілі бір тізбектің ДНҚ-сының
Ген транскрипциясының реттелуі мыналардың есебінен жүзеге асады
1) Белгілі бір тізбектің ДНҚ-сының

Ген экспрессиясын реттеуші белоктар спецификалық нуклеотидтік тізбектерді тануы тиіс.
ДНҚ-ның бір
Ген экспрессиясын реттеуші белоктар спецификалық нуклеотидтік тізбектерді тануы тиіс.
ДНҚ-ның бір
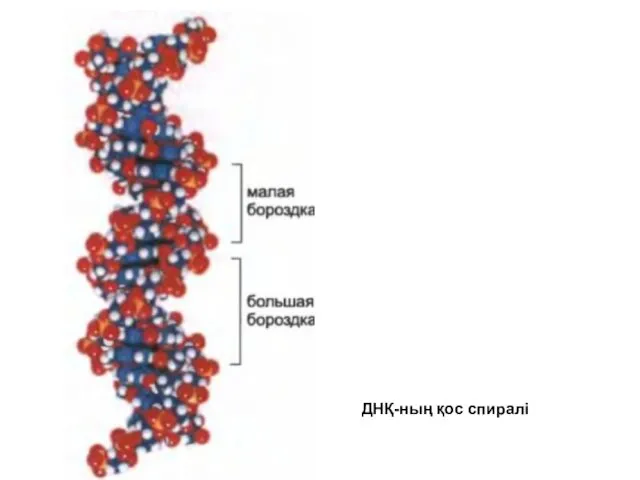
ДНҚ-ның қос спиралі
ДНҚ-ның қос спиралі
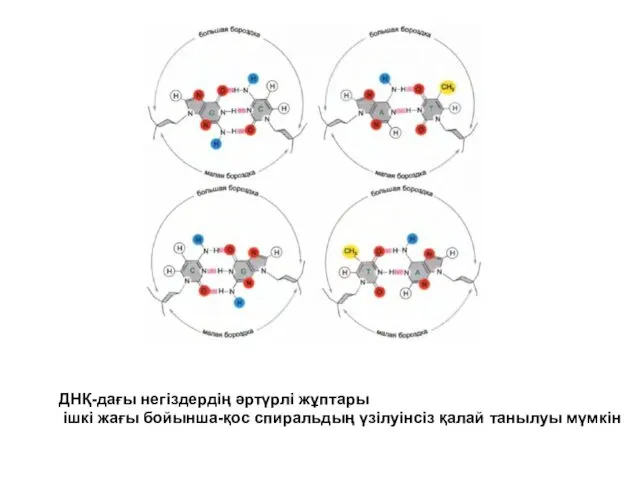
ДНҚ-дағы негіздердің әртүрлі жұптары
ішкі жағы бойынша-қос спиральдың үзілуінсіз қалай танылуы
ДНҚ-дағы негіздердің әртүрлі жұптары
ішкі жағы бойынша-қос спиральдың үзілуінсіз қалай танылуы
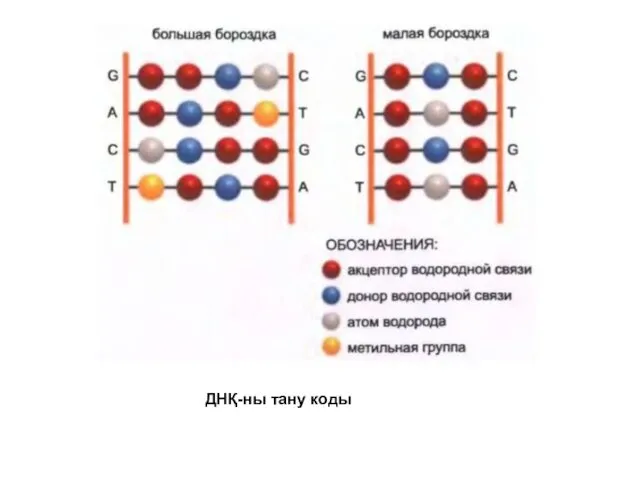
ДНҚ-ны тану коды
ДНҚ-ны тану коды

Реттеуші белок ДНҚ-ның спецификалық тізбегін таниды, себебі белок беткейі белгілі бір
Реттеуші белок ДНҚ-ның спецификалық тізбегін таниды, себебі белок беткейі белгілі бір

Кейбір реттеуші белоктар және олар танитын ДНҚ тізбегі
Кейбір реттеуші белоктар және олар танитын ДНҚ тізбегі

Реттеуші белоктың ДНҚ-ның үлкен жырашығымен байланысуы
Реттеуші белоктың ДНҚ-ның үлкен жырашығымен байланысуы

Бірнеше мыңдаған транскрипция факторлары белгілі. Олардың классификациясы гомологтарының бірнші реттік және
Бірнеше мыңдаған транскрипция факторлары белгілі. Олардың классификациясы гомологтарының бірнші реттік және

Транскрипция факторының классификациясы
Транскрипция факторының классификациясы




Транскрипция факторларының қызметі, механизмі үш саты арқылы жүреді;
ДНҚ мен байланысу,
Транскрипция барысына
Транскрипция факторларының қызметі, механизмі үш саты арқылы жүреді;
ДНҚ мен байланысу,
Транскрипция барысына

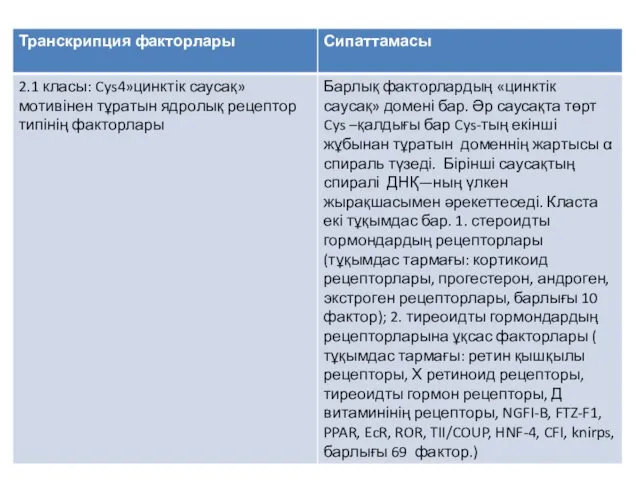
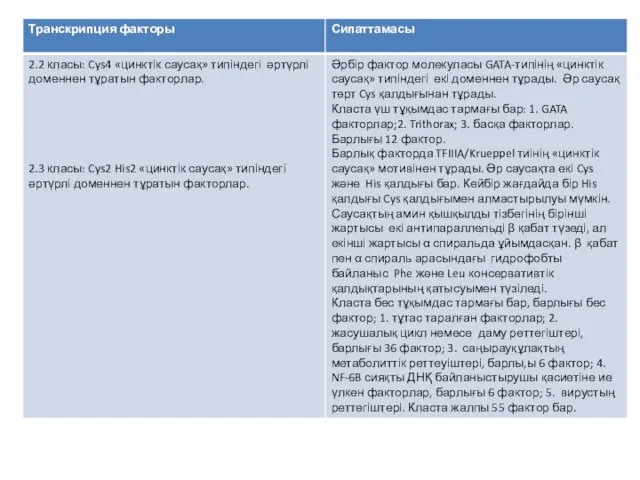
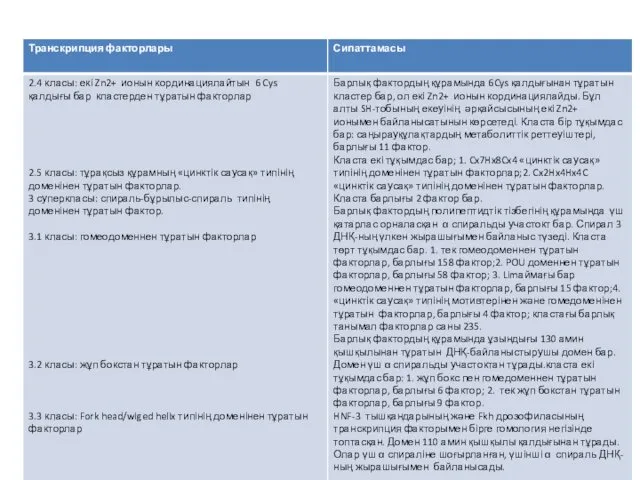
Қазіргі уақытта, транскрипция факторларын жіктеудің негізінде, олардың әртүрлі типтерінен, бәріне ортақ
Қазіргі уақытта, транскрипция факторларын жіктеудің негізінде, олардың әртүрлі типтерінен, бәріне ортақ
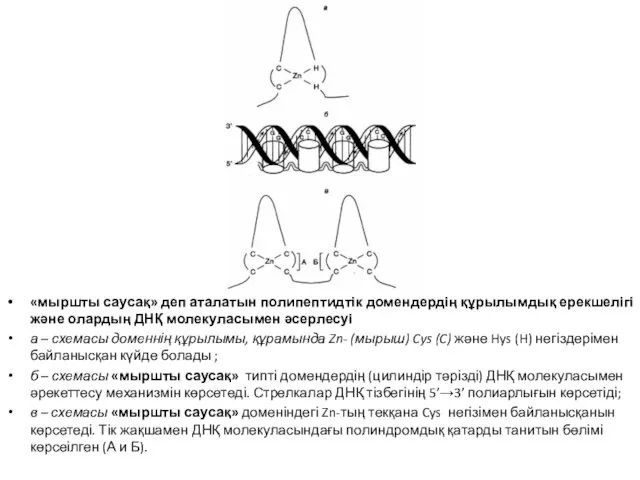
«мыршты саусақ» деп аталатын полипептидтік домендердің құрылымдық ерекшелігі және олардың ДНҚ
«мыршты саусақ» деп аталатын полипептидтік домендердің құрылымдық ерекшелігі және олардың ДНҚ
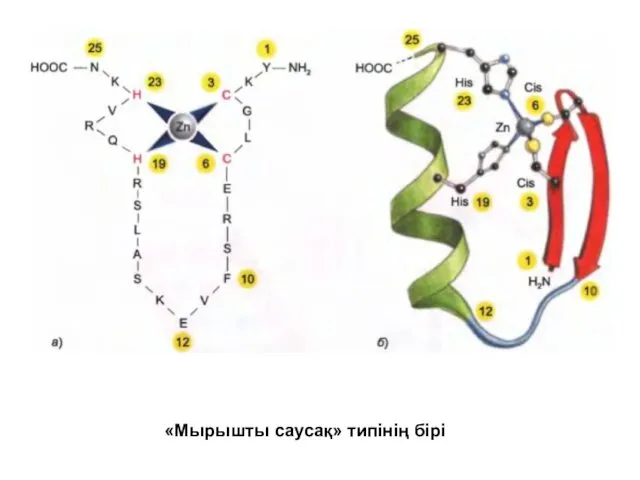
«Мырышты саусақ» типінің бірі
«Мырышты саусақ» типінің бірі
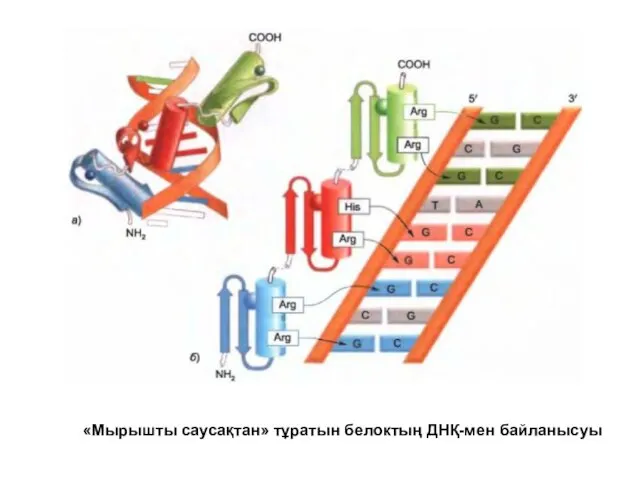
«Мырышты саусақтан» тұратын белоктың ДНҚ-мен байланысуы
«Мырышты саусақтан» тұратын белоктың ДНҚ-мен байланысуы
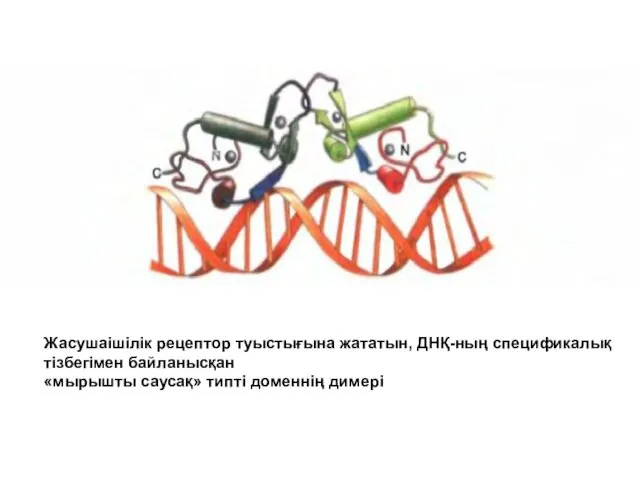
Жасушаішілік рецептор туыстығына жататын, ДНҚ-ның спецификалық
тізбегімен байланысқан
«мырышты саусақ» типті
Жасушаішілік рецептор туыстығына жататын, ДНҚ-ның спецификалық
тізбегімен байланысқан
«мырышты саусақ» типті
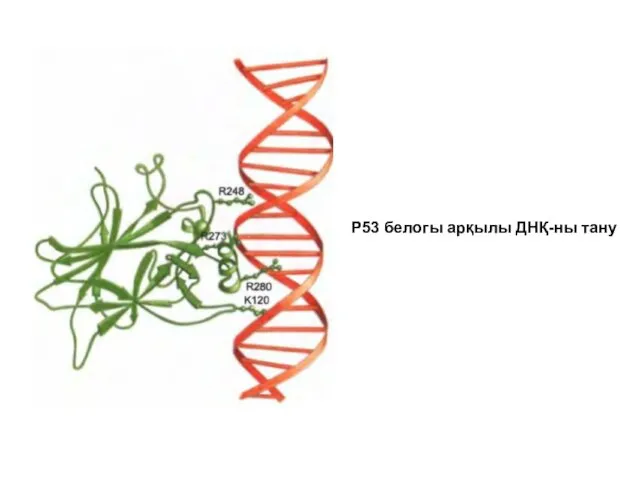
Р53 белогы арқылы ДНҚ-ны тану
Р53 белогы арқылы ДНҚ-ны тану

«мырышты саусақ» типті домендер көпеген транскрипця факторларында табылған; РНҚ-полимераза ІІ қызметін
«мырышты саусақ» типті домендер көпеген транскрипця факторларында табылған; РНҚ-полимераза ІІ қызметін

Әртүрлі стероидтық кормондар рецепторларының ДНҚ-мен байланысушы орындарын зерттеу арқылы, олардың өзара
Әртүрлі стероидтық кормондар рецепторларының ДНҚ-мен байланысушы орындарын зерттеу арқылы, олардың өзара

Клюкокортикоидтар рецепторының мырышты саусағының N- соңының не бәрі екі аминқышқылының орын
Клюкокортикоидтар рецепторының мырышты саусағының N- соңының не бәрі екі аминқышқылының орын

“спираль-бұрылыс-спираль” мотивті транскрипция факторлары.
ДНҚ молекуласындағы реттеуші арнаулы бөлігін танитын, басқаша пептидтік
“спираль-бұрылыс-спираль” мотивті транскрипция факторлары.
ДНҚ молекуласындағы реттеуші арнаулы бөлігін танитын, басқаша пептидтік
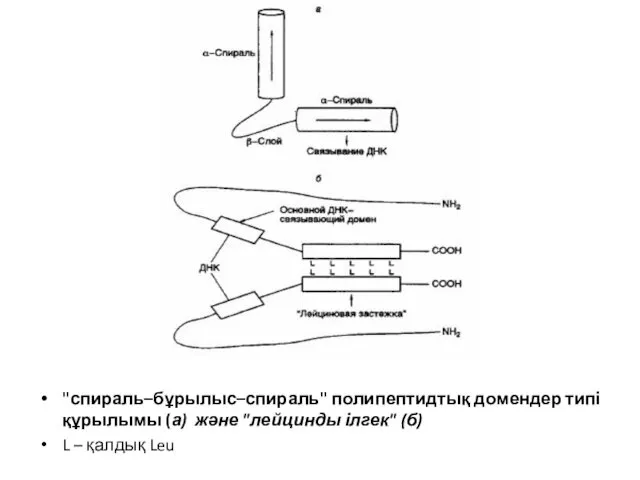
"спираль–бұрылыс–спираль" полипептидтық домендер типі құрылымы (а) және "лейцинды ілгек" (б)
L
"спираль–бұрылыс–спираль" полипептидтық домендер типі құрылымы (а) және "лейцинды ілгек" (б)
L

«Лейцинді ілгек» типті ДНК-байланысушы домендер.
Leu қалдығының бұл құрылымында, фактордың α-спираль
«Лейцинді ілгек» типті ДНК-байланысушы домендер.
Leu қалдығының бұл құрылымында, фактордың α-спираль
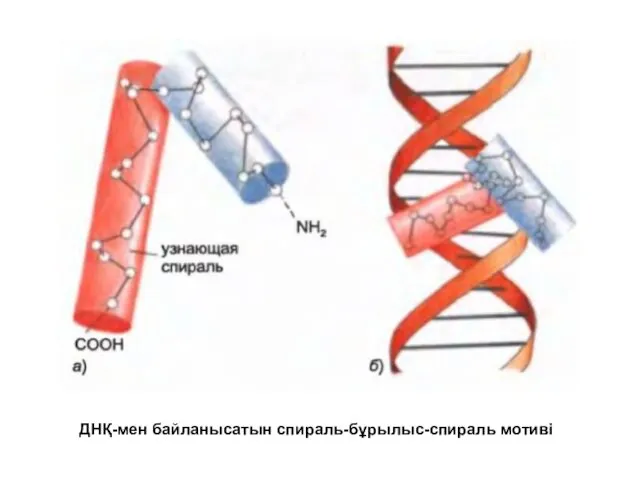
ДНҚ-мен байланысатын спираль-бұрылыс-спираль мотиві
ДНҚ-мен байланысатын спираль-бұрылыс-спираль мотиві
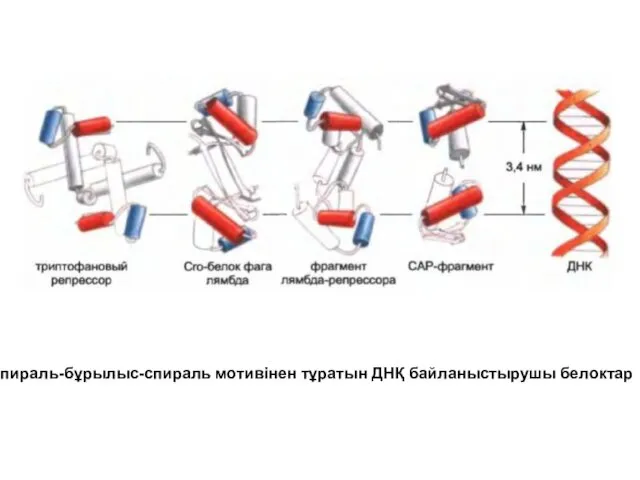
Спираль-бұрылыс-спираль мотивінен тұратын ДНҚ байланыстырушы белоктар
Спираль-бұрылыс-спираль мотивінен тұратын ДНҚ байланыстырушы белоктар
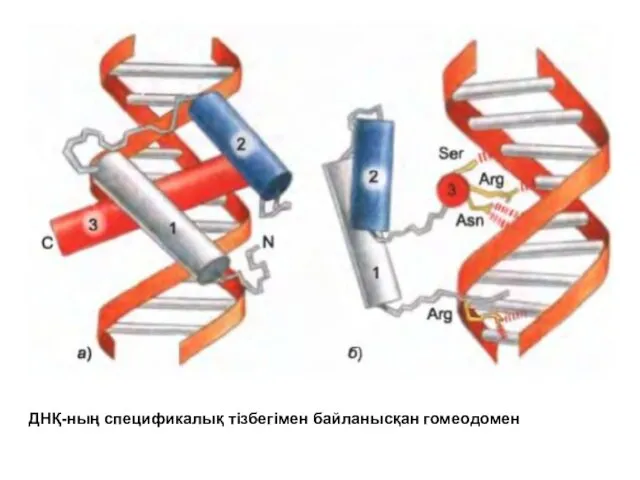
ДНҚ-ның спецификалық тізбегімен байланысқан гомеодомен
ДНҚ-ның спецификалық тізбегімен байланысқан гомеодомен
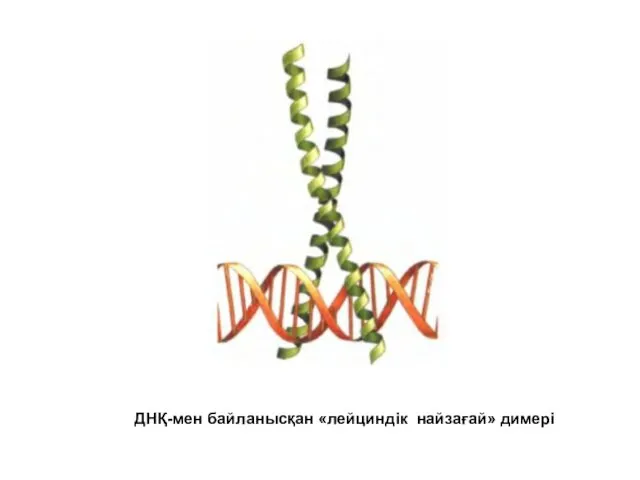
ДНҚ-мен байланысқан «лейциндік найзағай» димері
ДНҚ-мен байланысқан «лейциндік найзағай» димері

«Лейциндік молниядан» тұратын белоктардың гетеродимеризациясы
ДНҚ-мен байланысудың спецификалық қасиетін өзгерте алады
«Лейциндік молниядан» тұратын белоктардың гетеродимеризациясы
ДНҚ-мен байланысудың спецификалық қасиетін өзгерте алады
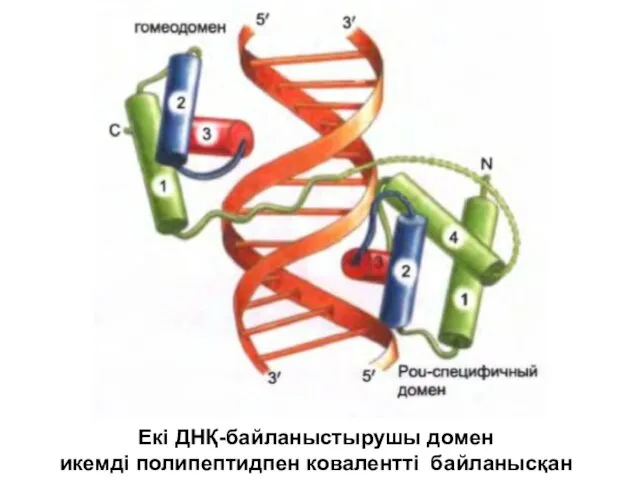
Екі ДНҚ-байланыстырушы домен
икемді полипептидпен ковалентті байланысқан
Екі ДНҚ-байланыстырушы домен
икемді полипептидпен ковалентті байланысқан
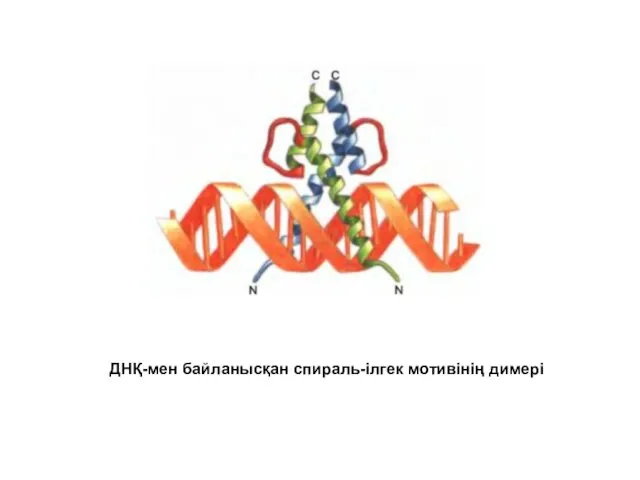
ДНҚ-мен байланысқан спираль-ілгек мотивінің димері
ДНҚ-мен байланысқан спираль-ілгек мотивінің димері
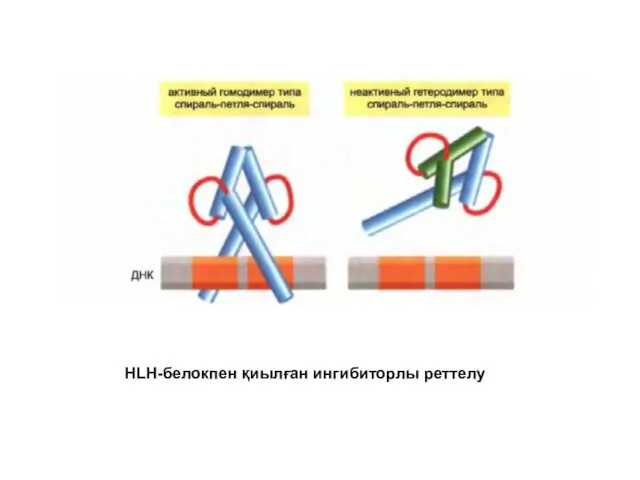
HLH-белокпен қиылған ингибиторлы реттелу
HLH-белокпен қиылған ингибиторлы реттелу
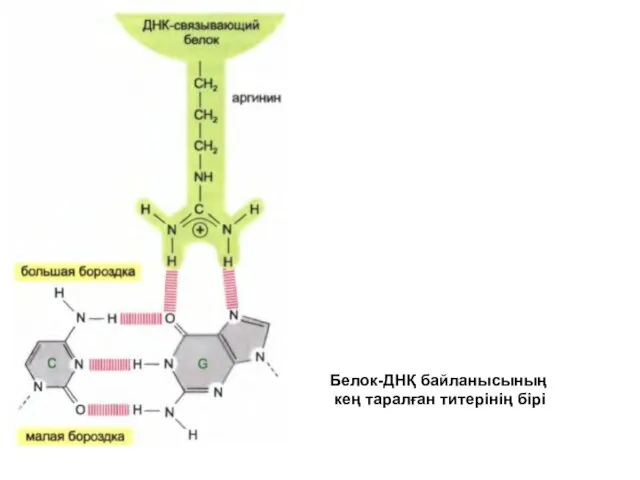
Белок-ДНҚ байланысының
кең таралған титерінің бірі
Белок-ДНҚ байланысының
кең таралған титерінің бірі
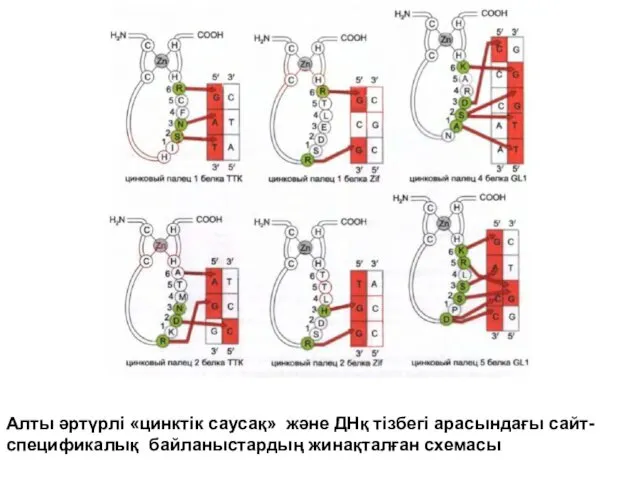
Алты әртүрлі «цинктік саусақ» және ДНқ тізбегі арасындағы сайт-спецификалық байланыстардың жинақталған
Алты әртүрлі «цинктік саусақ» және ДНқ тізбегі арасындағы сайт-спецификалық байланыстардың жинақталған
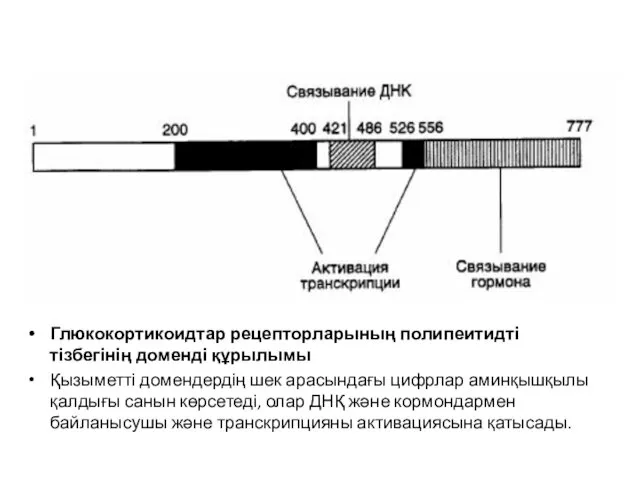
Глюкокортикоидтар рецепторларының полипеитидті тізбегінің доменді құрылымы
Қызыметті домендердің шек арасындағы цифрлар аминқышқылы
Глюкокортикоидтар рецепторларының полипеитидті тізбегінің доменді құрылымы
Қызыметті домендердің шек арасындағы цифрлар аминқышқылы


Транскрипцияның белсенуін генетикалық бақылау.
Транскрипцияның көптеген факторларының біріншілік рөлі белгілі бір ұлпада
Транскрипцияның белсенуін генетикалық бақылау.
Транскрипцияның көптеген факторларының біріншілік рөлі белгілі бір ұлпада
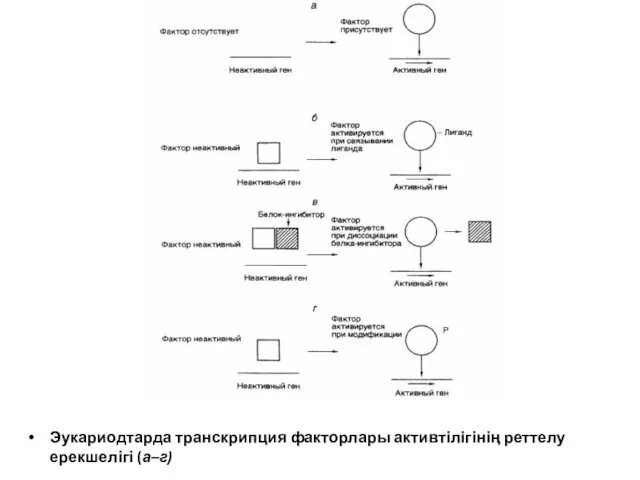
Эукариодтарда транскрипция факторлары активтілігінің реттелу ерекшелігі (а–г)
Эукариодтарда транскрипция факторлары активтілігінің реттелу ерекшелігі (а–г)

Эукариот генінің транскрипциясын бақылау
Эукариоттық РНҚ полимераза II үшін транскрипцияның 5 ортақ
Эукариот генінің транскрипциясын бақылау
Эукариоттық РНҚ полимераза II үшін транскрипцияның 5 ортақ

Геннің бақылаушы аймағы -ген транскрипциясының и нициациясын реттеуге қатысатын ДНҚ-ның бүкіл
Геннің бақылаушы аймағы -ген транскрипциясының и нициациясын реттеуге қатысатын ДНҚ-ның бүкіл
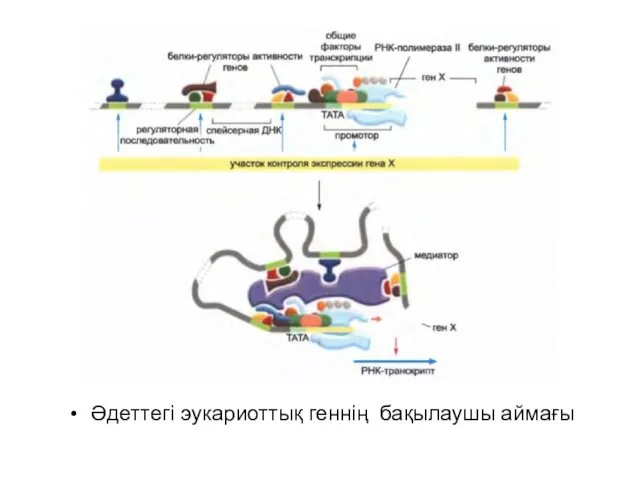
Әдеттегі эукариоттық геннің бақылаушы аймағы
Әдеттегі эукариоттық геннің бақылаушы аймағы

Эукариоттық белок-активаторлар байланысқан ДНҚ участоктары бірінші энхансер деп аталған болатын. Алайда
Эукариоттық белок-активаторлар байланысқан ДНҚ участоктары бірінші энхансер деп аталған болатын. Алайда
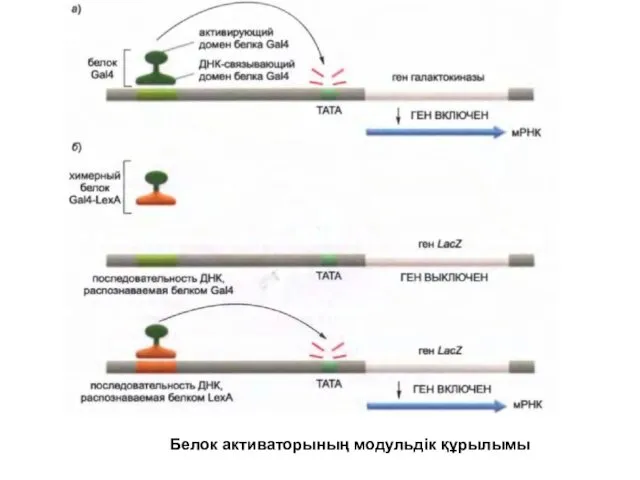
Белок активаторының модульдік құрылымы
Белок активаторының модульдік құрылымы

Транскрипцияның ортақ факторлары, медиатор және РНҚ полимераза стандарттық нуклеосомаға жинақталған промоторда
Транскрипцияның ортақ факторлары, медиатор және РНҚ полимераза стандарттық нуклеосомаға жинақталған промоторда
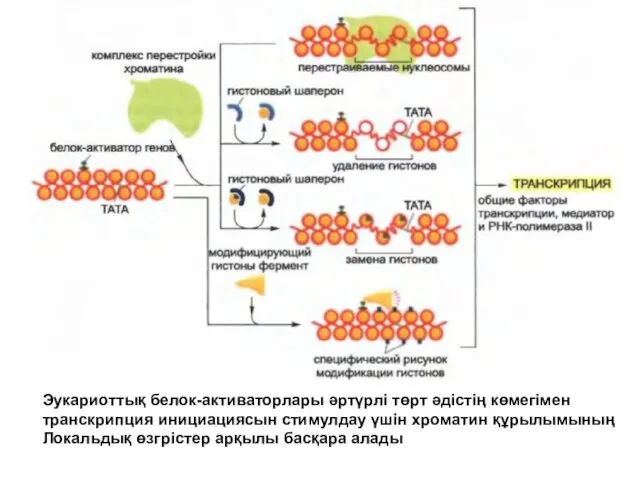
Эукариоттық белок-активаторлары әртүрлі төрт әдістің көмегімен
транскрипция инициациясын стимулдау үшін хроматин
Эукариоттық белок-активаторлары әртүрлі төрт әдістің көмегімен
транскрипция инициациясын стимулдау үшін хроматин

Алайда транскрипция кезіндегі гсистондардың коваленттік модификациясының өте маңызды рөлі хроматиннің құрылымының
Алайда транскрипция кезіндегі гсистондардың коваленттік модификациясының өте маңызды рөлі хроматиннің құрылымының
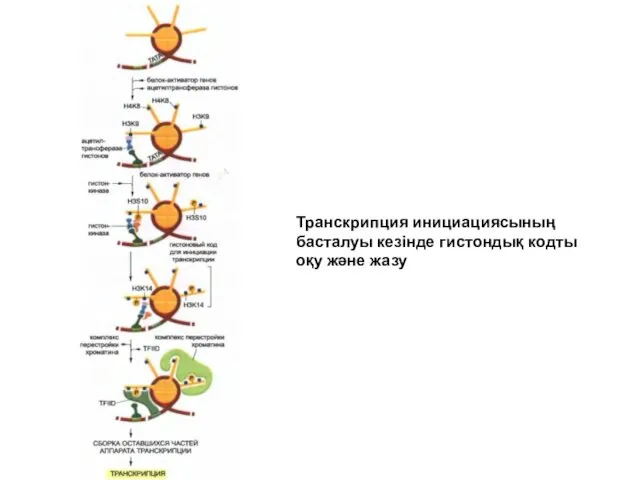
Транскрипция инициациясының
басталуы кезінде гистондық кодты
оқу және жазу
Транскрипция инициациясының
басталуы кезінде гистондық кодты
оқу және жазу

Реакцияның жылдамдығын арттыру үшін бірлесіп бірнеше факторлар әрекет еткен жерлердегі олардың
Реакцияның жылдамдығын арттыру үшін бірлесіп бірнеше факторлар әрекет еткен жерлердегі олардың
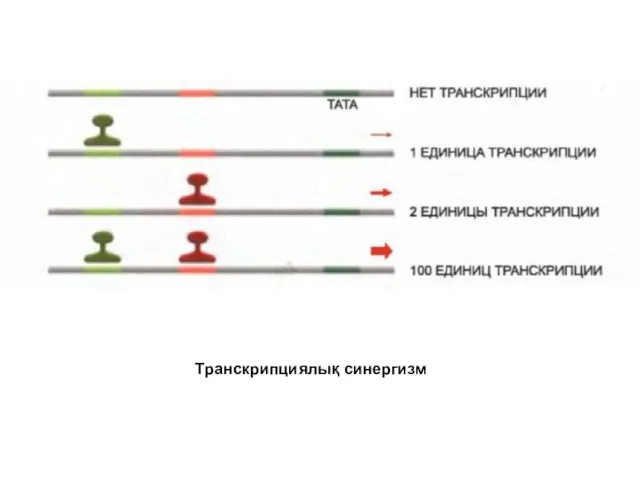
Транскрипциялық синергизм
Транскрипциялық синергизм

Белок-активаторлар транскрипцияның активациясына әкелетін жолдардың әртүрлі кезеңдеріне әсер ете алатын болғандықтан
Белок-активаторлар транскрипцияның активациясына әкелетін жолдардың әртүрлі кезеңдеріне әсер ете алатын болғандықтан
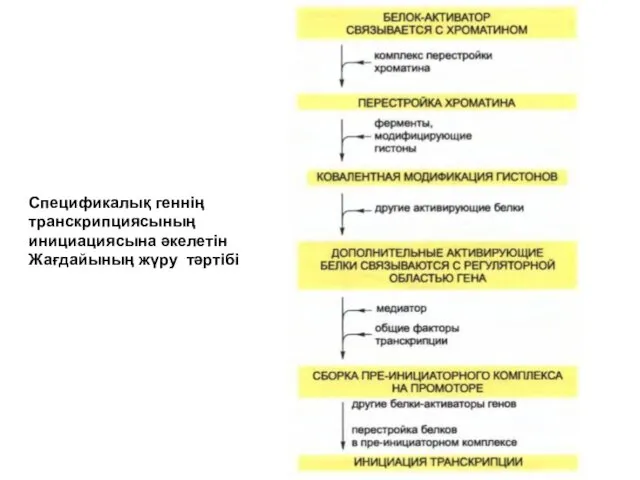
Спецификалық геннің
транскрипциясының
инициациясына әкелетін
Жағдайының жүру тәртібі
Спецификалық геннің
транскрипциясының
инициациясына әкелетін
Жағдайының жүру тәртібі

Жеке реттеуші белок реттеуші кешеннің бірнеше типіне қатыса алады.
Мысалы, бір
Жеке реттеуші белок реттеуші кешеннің бірнеше типіне қатыса алады.
Мысалы, бір
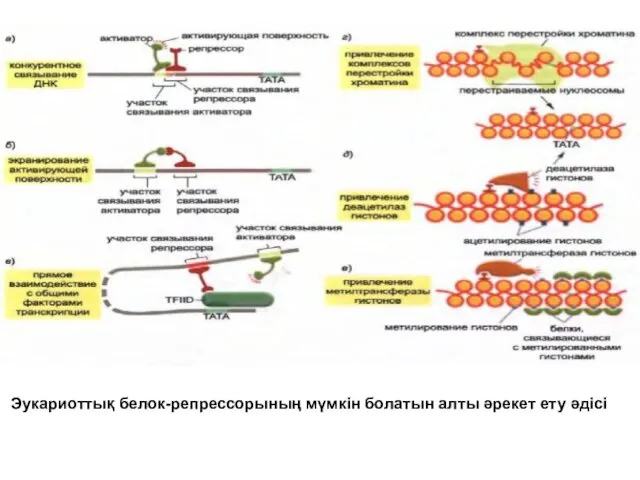
Эукариоттық белок-репрессорының мүмкін болатын алты әрекет ету әдісі
Эукариоттық белок-репрессорының мүмкін болатын алты әрекет ету әдісі
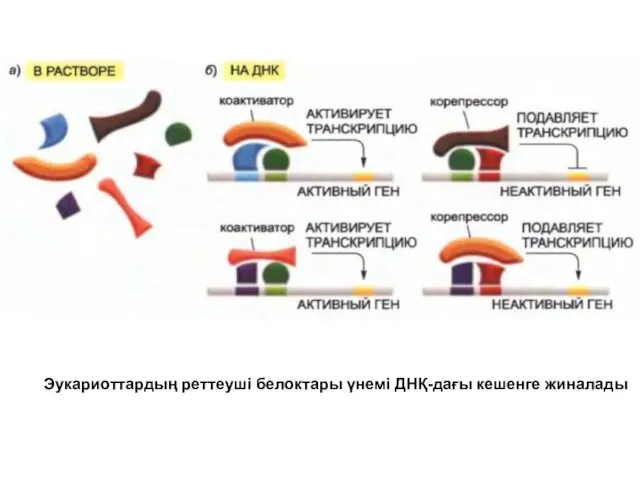
Эукариоттардың реттеуші белоктары үнемі ДНҚ-дағы кешенге жиналады
Эукариоттардың реттеуші белоктары үнемі ДНҚ-дағы кешенге жиналады
 Солнечный цветок.
Солнечный цветок.  Історія вивчення клітини. Методи цитологічних досліджень
Історія вивчення клітини. Методи цитологічних досліджень Презентация на тему "Дыхание" - скачать презентации по Биологии
Презентация на тему "Дыхание" - скачать презентации по Биологии Биохимическая гипотеза возникновения и развития жизни на Земле
Биохимическая гипотеза возникновения и развития жизни на Земле Презентация на тему "Табакокурение" - скачать презентации по Биологии
Презентация на тему "Табакокурение" - скачать презентации по Биологии Информация и эволюция живого мира Работа студента группы 212-псо Кузнецова Даниила.
Информация и эволюция живого мира Работа студента группы 212-псо Кузнецова Даниила. Интересные факты об экзотических фруктах и овощах
Интересные факты об экзотических фруктах и овощах Характеристика микрофлоры почвы, воды и воздуха
Характеристика микрофлоры почвы, воды и воздуха Физиология дыхательной системы
Физиология дыхательной системы Презентация Химические загрязнения среды промышленностью
Презентация Химические загрязнения среды промышленностью  Презентация на тему "Наше здоровье" - скачать презентации по Биологии
Презентация на тему "Наше здоровье" - скачать презентации по Биологии Ядовитые животные Автор Самойленко Э.А. учитель биологии МБОУСОШ № 14 пгт Ильского
Ядовитые животные Автор Самойленко Э.А. учитель биологии МБОУСОШ № 14 пгт Ильского Обмен веществ и энергии. Характеристика метаболизма
Обмен веществ и энергии. Характеристика метаболизма Мозг необъятен как Вселенная
Мозг необъятен как Вселенная Полезное - рядом. Лечебные свойства лука
Полезное - рядом. Лечебные свойства лука Дыхание и кровообращение
Дыхание и кровообращение Скелет человека
Скелет человека Презентация на тему "Размножение и развитие Земноводных. Происхождение Земноводных" - скачать презентации по Биологии
Презентация на тему "Размножение и развитие Земноводных. Происхождение Земноводных" - скачать презентации по Биологии Презентация на тему "Классификация неорганических веществ" - скачать презентации по Биологии
Презентация на тему "Классификация неорганических веществ" - скачать презентации по Биологии Строение и работа скелетных мышц Презентация для 8 класса Проценко Л.В.
Строение и работа скелетных мышц Презентация для 8 класса Проценко Л.В.  Ёж обыкновенный
Ёж обыкновенный Тема работы Изменчивость качественных и количественных признаков бородатых ирисов степной зоны Кабардино-Балкарии
Тема работы Изменчивость качественных и количественных признаков бородатых ирисов степной зоны Кабардино-Балкарии  Сравнительная характеристика гончих в СССР и в настоящее время
Сравнительная характеристика гончих в СССР и в настоящее время Презентация на тему Осанка и ее коррекция
Презентация на тему Осанка и ее коррекция  Презентация на тему "Основоположник учения об условных рефлексах" - скачать презентации по Биологии
Презентация на тему "Основоположник учения об условных рефлексах" - скачать презентации по Биологии Рыбы. Где живут рыбы?
Рыбы. Где живут рыбы? Отряд Двукрылые Составила: учитель биологии и химии высшей квалификационной категории Бочкова Ирина Анатольевна
Отряд Двукрылые Составила: учитель биологии и химии высшей квалификационной категории Бочкова Ирина Анатольевна  Столберова Галина
Столберова Галина