- Генетика популяцій

Содержание
- 2. Популяція — одиниця еволюційного процесу Біологічна еволюція — це процес змін і дивергенції біологічних форм у
- 3. Генетики користуються поняттями ідеальних і реальних популяцій. Ідеальна популяція складається із безмежної кількості особин, між якими
- 4. Частоти генотипів та частоти алелів Головна відмінність методології популяційної генетики від уже звичної методології генетичного аналізу
- 5. Закон Харді — Вайнберга Закон Харді-Вайнберга свідчить про те, що спадкування як таке не міняє частоти
- 6. Неважко побачити, що при цьому зберігається вихідна частота алелей: 0,5А и 0,5а. У наступному поколінні одержують
- 7. Модель Харди-Вайнберга, показує, що відбувається з алелем і з генотипом в «ідеальній» популяції (позбавленої всіх складних
- 8. Застосування закону Харди-Вайнберга: розрахунок частоти гетерозигот Одне із застосувань закону Харді-Вайнберга — оцінка частоти гетерозигот у
- 9. Співвідношення частот алелей і генотипів, що випливає з рівняння Харді-Вайнберга
- 10. Проблема генетичної гетерогенності природних популяцій Значення генетичної гетерогенності природних популяцій уперше по достоїнству оцінив С. С.
- 11. В 40— 50-х роках розглядалися в основному дві точки зору на структуру природних популяцій: класична модель
- 12. Методи вивчення структури популяцій Одним із методів, який широко використовується в популяційній генетиці, є електрофоретичний розподіл
- 13. Зімограмми аспартатамінотрансферази (ААТ) 25 проростків з популяції жита Secale cereale. Множинні форми ААТ (фермент - димер,
- 14. Поліморфізм (Р) популяції оцінюється часткою поліморфних локусів із всіх досліджених. Так, з 30 локусів, вивчених у
- 15. Оскільки популяції, безсумнівно, розрізняються по ступені панмиксії (наприклад, перехреснозапильники і самозапильники), а також по селективній цінності
- 17. Елементарна еволюційна подія — зміна частот алелей у популяції Зміна й дивергенція біологічних форм у часі
- 18. Внутривидова дівергенція або утворення видів Процес видоутворення, що пов’язаний із просторовим розєднанням популяції є алопатричним (алос
- 19. Добір Генетична гетерогенність, широко розповсюджена в природних популяціях, становить основу ефективності дарвінівського природного добору. Пряму кореляцію
- 20. Частота алелі А у F1 після добору буде: Зміна частоти алелі А за покоління буде складати:
- 21. При крайніх значеннях частот алелей добір діє найменш ефективно. Найбільше ефективним є добір при середніх значеннях
- 22. Результати дії стабілізуючого, спрямованого й дизруптивного добору. У кожному з показаних випадків х0 - середнє арифметичне
- 23. Як відзначалося, добір діє на фенотипи, а непрямо позначається на зміні частот алелей. Залежно від того
- 24. Мутаційний процес Мутаційний процес — основа виникнення гетерогенності популяції. Через наявність мутаційного процесу важко говорити про
- 25. Потік генів Потік генів, або міграції особин, являє собою обмін генами між популяціями. Якщо популяції мають
- 26. Дрейф генів, або генетико-автоматичні процеси, які впливають на частоти алелей у популяціях Дрейф генів (або просто
- 27. Іноді, особливо в ізолятах чисельність популяції може бути дуже низькою. Зазвичай у нормальних популяціях бувають періоди
- 28. Інбридинг Кінцева чисельність реальних популяцій приводить ще до одного важливого наслідку. Розглянемо його на прикладі людини.
- 29. Інбридинг має кілька наслідків для популяції: Підвищення гомозиготності. Прояв рецесивных алелей. При звичайно негативному ефекті рецесивных
- 31. Скачать презентацию

Популяція — одиниця еволюційного процесу
Біологічна еволюція — це процес змін і
Популяція — одиниця еволюційного процесу
Біологічна еволюція — це процес змін і

Генетики користуються поняттями ідеальних і реальних популяцій. Ідеальна популяція складається із
Генетики користуються поняттями ідеальних і реальних популяцій. Ідеальна популяція складається із

Частоти генотипів та частоти алелів
Головна відмінність методології популяційної генетики від уже
Частоти генотипів та частоти алелів
Головна відмінність методології популяційної генетики від уже

Закон Харді — Вайнберга
Закон Харді-Вайнберга свідчить про те, що спадкування як
Закон Харді — Вайнберга
Закон Харді-Вайнберга свідчить про те, що спадкування як

Неважко побачити, що при цьому зберігається вихідна частота алелей: 0,5А и
Неважко побачити, що при цьому зберігається вихідна частота алелей: 0,5А и

Модель Харди-Вайнберга, показує, що відбувається з алелем і з генотипом в
Модель Харди-Вайнберга, показує, що відбувається з алелем і з генотипом в

Застосування закону Харди-Вайнберга: розрахунок частоти гетерозигот
Одне із застосувань закону Харді-Вайнберга —
Застосування закону Харди-Вайнберга: розрахунок частоти гетерозигот
Одне із застосувань закону Харді-Вайнберга —
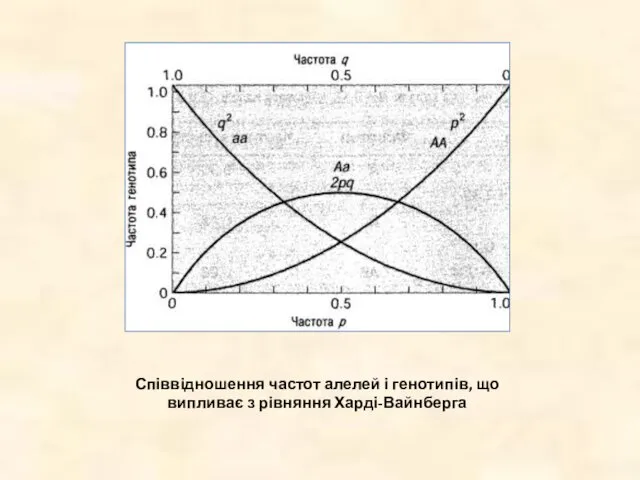
Співвідношення частот алелей і генотипів, що випливає з рівняння Харді-Вайнберга
Співвідношення частот алелей і генотипів, що випливає з рівняння Харді-Вайнберга

Проблема генетичної гетерогенності природних популяцій
Значення генетичної гетерогенності природних популяцій уперше по
Проблема генетичної гетерогенності природних популяцій
Значення генетичної гетерогенності природних популяцій уперше по

В 40— 50-х роках розглядалися в основному дві точки зору на
В 40— 50-х роках розглядалися в основному дві точки зору на

Методи вивчення структури популяцій
Одним із методів, який широко використовується в популяційній
Методи вивчення структури популяцій
Одним із методів, який широко використовується в популяційній

Зімограмми аспартатамінотрансферази (ААТ) 25 проростків з популяції жита Secale cereale. Множинні
Зімограмми аспартатамінотрансферази (ААТ) 25 проростків з популяції жита Secale cereale. Множинні

Поліморфізм (Р) популяції оцінюється часткою поліморфних локусів із всіх досліджених. Так,
Поліморфізм (Р) популяції оцінюється часткою поліморфних локусів із всіх досліджених. Так,

Оскільки популяції, безсумнівно, розрізняються по ступені панмиксії (наприклад, перехреснозапильники і самозапильники),
Оскільки популяції, безсумнівно, розрізняються по ступені панмиксії (наприклад, перехреснозапильники і самозапильники),


Елементарна еволюційна подія — зміна частот алелей у популяції
Зміна й дивергенція
Елементарна еволюційна подія — зміна частот алелей у популяції
Зміна й дивергенція

Внутривидова дівергенція або утворення видів
Процес видоутворення, що пов’язаний із просторовим розєднанням
Внутривидова дівергенція або утворення видів
Процес видоутворення, що пов’язаний із просторовим розєднанням

Добір
Генетична гетерогенність, широко розповсюджена в природних популяціях, становить основу ефективності дарвінівського
Добір
Генетична гетерогенність, широко розповсюджена в природних популяціях, становить основу ефективності дарвінівського

Частота алелі А у F1 після добору буде:
Зміна частоти алелі А
Частота алелі А у F1 після добору буде:
Зміна частоти алелі А

При крайніх значеннях частот алелей добір діє найменш ефективно. Найбільше ефективним
При крайніх значеннях частот алелей добір діє найменш ефективно. Найбільше ефективним

Результати дії стабілізуючого, спрямованого й дизруптивного добору. У кожному з показаних
Результати дії стабілізуючого, спрямованого й дизруптивного добору. У кожному з показаних

Як відзначалося, добір діє на фенотипи, а непрямо позначається на зміні
Як відзначалося, добір діє на фенотипи, а непрямо позначається на зміні

Мутаційний процес
Мутаційний процес — основа виникнення гетерогенності популяції. Через наявність мутаційного
Мутаційний процес
Мутаційний процес — основа виникнення гетерогенності популяції. Через наявність мутаційного

Потік генів
Потік генів, або міграції особин, являє собою обмін генами між
Потік генів
Потік генів, або міграції особин, являє собою обмін генами між

Дрейф генів, або генетико-автоматичні процеси, які впливають на частоти алелей у
Дрейф генів, або генетико-автоматичні процеси, які впливають на частоти алелей у

Іноді, особливо в ізолятах чисельність популяції може бути дуже низькою. Зазвичай
Іноді, особливо в ізолятах чисельність популяції може бути дуже низькою. Зазвичай

Інбридинг
Кінцева чисельність реальних популяцій приводить ще до одного важливого наслідку. Розглянемо
Інбридинг
Кінцева чисельність реальних популяцій приводить ще до одного важливого наслідку. Розглянемо

Інбридинг має кілька наслідків для популяції:
Підвищення гомозиготності.
Прояв рецесивных алелей.
При звичайно негативному
Інбридинг має кілька наслідків для популяції:
Підвищення гомозиготності.
Прояв рецесивных алелей.
При звичайно негативному
 Презентация на тему "Класс паукообразных" - скачать презентации по Биологии
Презентация на тему "Класс паукообразных" - скачать презентации по Биологии Презентация____
Презентация____ Классификация пресмыкающихся
Классификация пресмыкающихся Химический состав клеток
Химический состав клеток Строение, функции и свойства мембраны
Строение, функции и свойства мембраны Физиология мозжечка
Физиология мозжечка Общие признаки хордовых животных. Подтип бесчерепные
Общие признаки хордовых животных. Подтип бесчерепные Нормована годівля коней
Нормована годівля коней Культурные растения
Культурные растения  Презентация на тему "Воробьинообразные" - скачать бесплатно презентации по Биологии
Презентация на тему "Воробьинообразные" - скачать бесплатно презентации по Биологии Биохимия ферментов
Биохимия ферментов Роль биологии в космических исследованиях Чтобы понять какова роль биологии в космических исследованиях мы должны обратиться
Роль биологии в космических исследованиях Чтобы понять какова роль биологии в космических исследованиях мы должны обратиться Филиал МБОУ Бейской ОШИ Новониколаевская ООШ Учитель химии и биологии Годлевская Л.А
Филиал МБОУ Бейской ОШИ Новониколаевская ООШ Учитель химии и биологии Годлевская Л.А Безвременник весёлый
Безвременник весёлый Презентация на тему "МНОГООБРАЗИЕ ПРОСТЕЙШИХ. ПАРАЗИТИЧЕСКИЕ" - скачать бесплатно презентации по Биологии
Презентация на тему "МНОГООБРАЗИЕ ПРОСТЕЙШИХ. ПАРАЗИТИЧЕСКИЕ" - скачать бесплатно презентации по Биологии Мухи и меры борьбы с ними
Мухи и меры борьбы с ними Презентация на тему Строение и многообразие лишайников
Презентация на тему Строение и многообразие лишайников  Мир растений. Наука о растениях
Мир растений. Наука о растениях Великий учёный и хирург Николай Иванович Пирогов
Великий учёный и хирург Николай Иванович Пирогов Презентация на тему "Строение кожи" - скачать презентации по Биологии
Презентация на тему "Строение кожи" - скачать презентации по Биологии Собака - лучший друг человека
Собака - лучший друг человека Презентация на тему Тип Кольчатые черви
Презентация на тему Тип Кольчатые черви  Пути достижения биологического прогресса (главные направления прогрессивной эволюции) Составитель Большаков С. В.
Пути достижения биологического прогресса (главные направления прогрессивной эволюции) Составитель Большаков С. В.  Энергетический обмен в клетке
Энергетический обмен в клетке Гаметогенез у растений и животных. Спорогенез и гаметогенез у растений. Стадии гаметогенеза человека
Гаметогенез у растений и животных. Спорогенез и гаметогенез у растений. Стадии гаметогенеза человека Органы чувств
Органы чувств Когнитивное обучение
Когнитивное обучение Презентация на тему ЦВЕТОК .
Презентация на тему ЦВЕТОК .