- Биохимия ферментов

Содержание
- 2. 07/25/2023 Коваль А. Н. (C), 2006 Содержание Строение, размер и форма белковой молекулы, функции белков. Денатурация,
- 3. 07/25/2023 Коваль А. Н. (C), 2006 Пептиды и белки: общие сведения Белки При соединении аминокислот в
- 4. 07/25/2023 Коваль А. Н. (C), 2006 Примеры белков Чтобы дать представление о многообразии белков, на схеме
- 5. 07/25/2023 Коваль А. Н. (C), 2006 Ферменты Ферменты (от лат. fermentum - закваска), или энзимы (от
- 6. 07/25/2023 Коваль А. Н. (C), 2006 Ферменты являются биокатализаторами Ускоряют химические реакции Составляют функциональный аппарат клетки,
- 7. 07/25/2023 Коваль А. Н. (C), 2006 История энзимологии На заре истории развития человечества, уже использовались ряд
- 8. 07/25/2023 Коваль А. Н. (C), 2006 История энзимологии (прод.) Позже процесс ферментации был более подробно изучен
- 9. 07/25/2023 Коваль А. Н. (C), 2006 История энзимологии (прод.) В 1814 г. русский ученый (немец по
- 10. 07/25/2023 Коваль А. Н. (C), 2006 История энзимологии (прод.) Либих и Вёлер открыли эмульсин, расщепляющий амигдалин
- 11. 07/25/2023 Коваль А. Н. (C), 2006 История энзимологии. Спор Либиха и Пастера Важную роль в развитии
- 12. 07/25/2023 Коваль А. Н. (C), 2006 История энзимологии (прод.) В 1878 г. Манассеина получила бесклеточный препарат
- 13. 07/25/2023 Коваль А. Н. (C), 2006 История энзимологии (прод.) В 1894 году Э. Фишер предложил модель
- 14. 07/25/2023 Коваль А. Н. (C), 2006 История энзимологии (прод.) В 1963 году исследована аминокислотная последовательность РНКазы
- 15. 07/25/2023 Коваль А. Н. (C), 2006 Доказательства белковой природы ферментов Все ферменты действуют в мягких условиях:
- 16. 07/25/2023 Коваль А. Н. (C), 2006 Доказательства белковой природы ферментов (прод.) Ферменты как и белки обладают
- 17. 07/25/2023 Коваль А. Н. (C), 2006 Особенности ферментативного катализа Исключительно высокая эффективность. Эффективность биологического катализа превышает
- 18. 07/25/2023 Коваль А. Н. (C), 2006 Особенности ферментативного катализа (прод.) Кооперативность: ферменты способны организовывать ассоциации -
- 19. 07/25/2023 Коваль А. Н. (C), 2006 Строение ферментов Ферменты как и белки делятся на две группы:
- 20. 07/25/2023 Коваль А. Н. (C), 2006 Кофакторы Если константа диссоциации сложного фермента настолько мала, что в
- 21. 07/25/2023 Коваль А. Н. (C), 2006 Примеры коферментов Между апоферментом и коферментом существуют нековалентные связи (водородные
- 22. 07/25/2023 Коваль А. Н. (C), 2006 Простетическая группа Примером простетической группы является витамин Н (биотин) биотин
- 23. 07/25/2023 Коваль А. Н. (C), 2006 Тиаминпирофосфат, ТПФ Тиаминпирофосфат, ТПФ (производное витамина В1, тиамин) – кофермент
- 24. 07/25/2023 Коваль А. Н. (C), 2006 Витамин PP Витамин PP (никотиновая кислота, ниацин) в форме никотинамида
- 25. 07/25/2023 Коваль А. Н. (C), 2006 Строение NAD+ и NADP+
- 26. 07/25/2023 Коваль А. Н. (C), 2006 Рибофлавин, витамин B2. Строение ФАД Образует ФАД и ФМН –
- 27. 07/25/2023 Коваль А. Н. (C), 2006 Пантотеновая кислота Пантотеновая кислота – входит в состав коэнзима А
- 28. 07/25/2023 Коваль А. Н. (C), 2006 Металлы как кофакторы Многие двухвалентные металлы (Сu2+, Zn2+, Fe2+, Fe3+,
- 29. 07/25/2023 Коваль А. Н. (C), 2006 Металлы как кофакторы (прод.)
- 30. 07/25/2023 Коваль А. Н. (C), 2006 Структурно-функциональная организация ферментов. Активный центр Активный (субстратный) центр - это
- 31. 07/25/2023 Коваль А. Н. (C), 2006 Структурно-функциональная организация ферментов. Строение активного центра фермента Активный центр включает
- 32. 07/25/2023 Коваль А. Н. (C), 2006 Структурно-функциональная организация ферментов. Схема Активные центры Якорные площадки Субстратный субстрат
- 33. 07/25/2023 Коваль А. Н. (C), 2006 Формирование активного центра фермента Предполагается, что формирование активного центра фермента
- 34. 07/25/2023 Коваль А. Н. (C), 2006 Центры аллостерической регуляции У олигомерных ферментов (имеющих четвертичную структуру) имеются
- 35. 07/25/2023 Коваль А. Н. (C), 2006 Аллостерические ферменты Присоединение эффектора к аллостерическому центру приводит к изменению
- 36. 07/25/2023 Коваль А. Н. (C), 2006 Спасибо за внимание! Следующая лекция – Биохимия ферментов-2. Механизм действия
- 38. Скачать презентацию

07/25/2023
Коваль А. Н. (C), 2006
Содержание
Строение, размер и форма белковой молекулы, функции
07/25/2023
Коваль А. Н. (C), 2006
Содержание
Строение, размер и форма белковой молекулы, функции

07/25/2023
Коваль А. Н. (C), 2006
Пептиды и белки: общие сведения
Белки
При соединении аминокислот
07/25/2023
Коваль А. Н. (C), 2006
Пептиды и белки: общие сведения
Белки
При соединении аминокислот
07/25/2023
Коваль А. Н. (C), 2006
Примеры белков
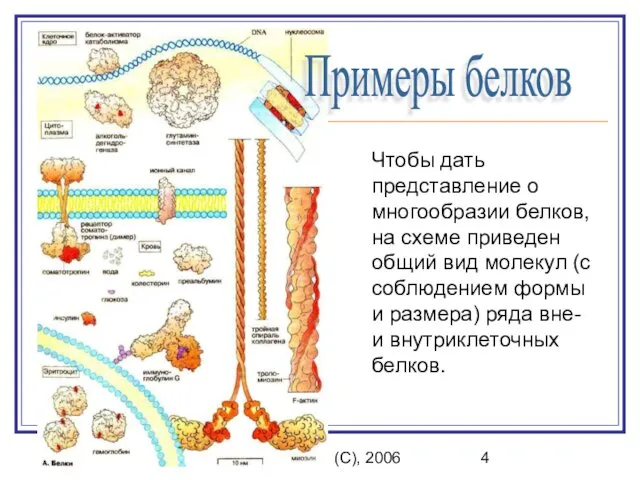
Чтобы дать представление о многообразии белков,
07/25/2023
Коваль А. Н. (C), 2006
Примеры белков
Чтобы дать представление о многообразии белков,

07/25/2023
Коваль А. Н. (C), 2006
Ферменты
Ферменты (от лат. fermentum - закваска), или
07/25/2023
Коваль А. Н. (C), 2006
Ферменты
Ферменты (от лат. fermentum - закваска), или

07/25/2023
Коваль А. Н. (C), 2006
Ферменты являются биокатализаторами
Ускоряют химические реакции
Составляют функциональный аппарат
07/25/2023
Коваль А. Н. (C), 2006
Ферменты являются биокатализаторами
Ускоряют химические реакции
Составляют функциональный аппарат

07/25/2023
Коваль А. Н. (C), 2006
История энзимологии
На заре истории развития человечества, уже
07/25/2023
Коваль А. Н. (C), 2006
История энзимологии
На заре истории развития человечества, уже

07/25/2023
Коваль А. Н. (C), 2006
История энзимологии (прод.)
Позже процесс ферментации был более
07/25/2023
Коваль А. Н. (C), 2006
История энзимологии (прод.)
Позже процесс ферментации был более

07/25/2023
Коваль А. Н. (C), 2006
История энзимологии (прод.)
В 1814 г. русский ученый
07/25/2023
Коваль А. Н. (C), 2006
История энзимологии (прод.)
В 1814 г. русский ученый

07/25/2023
Коваль А. Н. (C), 2006
История энзимологии (прод.)
Либих и Вёлер открыли эмульсин,
07/25/2023
Коваль А. Н. (C), 2006
История энзимологии (прод.)
Либих и Вёлер открыли эмульсин,

07/25/2023
Коваль А. Н. (C), 2006
История энзимологии.
Спор Либиха и Пастера
Важную роль
07/25/2023
Коваль А. Н. (C), 2006
История энзимологии.
Спор Либиха и Пастера
Важную роль

07/25/2023
Коваль А. Н. (C), 2006
История энзимологии (прод.)
В 1878 г. Манассеина получила
07/25/2023
Коваль А. Н. (C), 2006
История энзимологии (прод.)
В 1878 г. Манассеина получила

07/25/2023
Коваль А. Н. (C), 2006
История энзимологии (прод.)
В 1894 году Э. Фишер
07/25/2023
Коваль А. Н. (C), 2006
История энзимологии (прод.)
В 1894 году Э. Фишер

07/25/2023
Коваль А. Н. (C), 2006
История энзимологии (прод.)
В 1963 году исследована аминокислотная
07/25/2023
Коваль А. Н. (C), 2006
История энзимологии (прод.)
В 1963 году исследована аминокислотная

07/25/2023
Коваль А. Н. (C), 2006
Доказательства белковой природы ферментов
Все ферменты действуют в
07/25/2023
Коваль А. Н. (C), 2006
Доказательства белковой природы ферментов
Все ферменты действуют в

07/25/2023
Коваль А. Н. (C), 2006
Доказательства белковой природы ферментов (прод.)
Ферменты как и
07/25/2023
Коваль А. Н. (C), 2006
Доказательства белковой природы ферментов (прод.)
Ферменты как и

07/25/2023
Коваль А. Н. (C), 2006
Особенности ферментативного катализа
Исключительно высокая эффективность.
Эффективность биологического
07/25/2023
Коваль А. Н. (C), 2006
Особенности ферментативного катализа
Исключительно высокая эффективность.
Эффективность биологического

07/25/2023
Коваль А. Н. (C), 2006
Особенности ферментативного катализа (прод.)
Кооперативность: ферменты способны организовывать
07/25/2023
Коваль А. Н. (C), 2006
Особенности ферментативного катализа (прод.)
Кооперативность: ферменты способны организовывать

07/25/2023
Коваль А. Н. (C), 2006
Строение ферментов
Ферменты как и белки делятся на
07/25/2023
Коваль А. Н. (C), 2006
Строение ферментов
Ферменты как и белки делятся на

07/25/2023
Коваль А. Н. (C), 2006
Кофакторы
Если константа диссоциации сложного фермента настолько мала,
07/25/2023
Коваль А. Н. (C), 2006
Кофакторы
Если константа диссоциации сложного фермента настолько мала,

07/25/2023
Коваль А. Н. (C), 2006
Примеры коферментов
Между апоферментом и коферментом существуют нековалентные
07/25/2023
Коваль А. Н. (C), 2006
Примеры коферментов
Между апоферментом и коферментом существуют нековалентные

07/25/2023
Коваль А. Н. (C), 2006
Простетическая группа
Примером простетической группы является витамин Н
07/25/2023
Коваль А. Н. (C), 2006
Простетическая группа
Примером простетической группы является витамин Н
07/25/2023
Коваль А. Н. (C), 2006
Тиаминпирофосфат, ТПФ
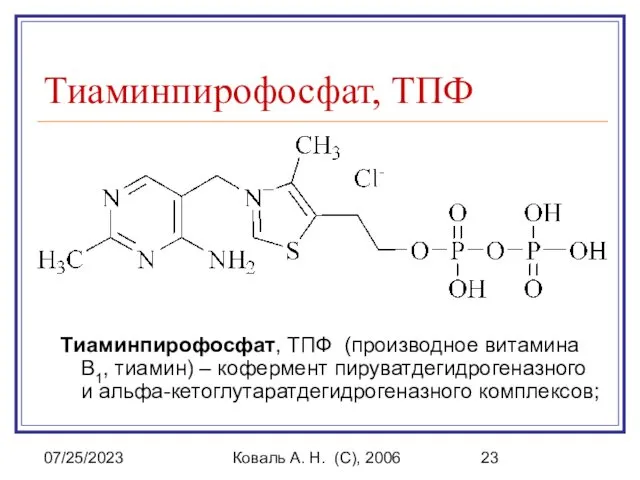
Тиаминпирофосфат, ТПФ (производное витамина В1, тиамин)
07/25/2023
Коваль А. Н. (C), 2006
Тиаминпирофосфат, ТПФ
Тиаминпирофосфат, ТПФ (производное витамина В1, тиамин)
07/25/2023
Коваль А. Н. (C), 2006
Витамин PP
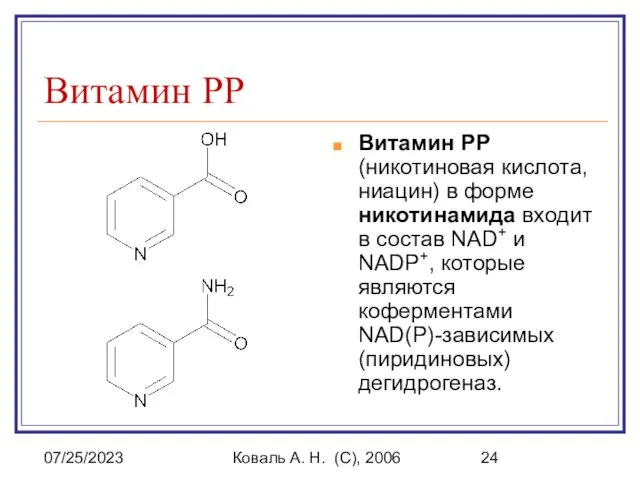
Витамин PP (никотиновая кислота, ниацин) в
07/25/2023
Коваль А. Н. (C), 2006
Витамин PP
Витамин PP (никотиновая кислота, ниацин) в
07/25/2023
Коваль А. Н. (C), 2006
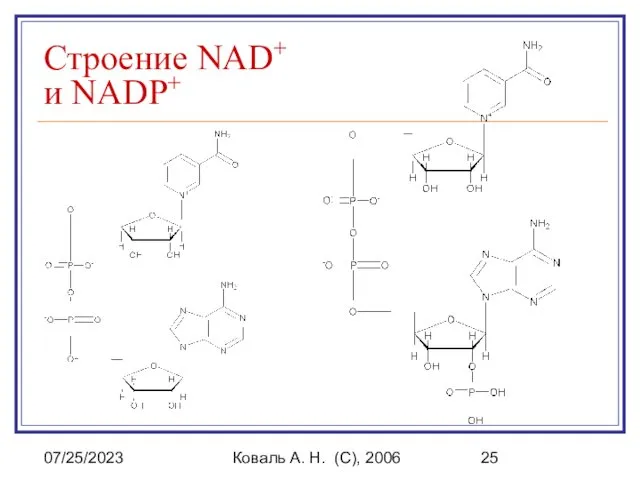
Строение NAD+ и NADP+
07/25/2023
Коваль А. Н. (C), 2006
Строение NAD+ и NADP+
07/25/2023
Коваль А. Н. (C), 2006
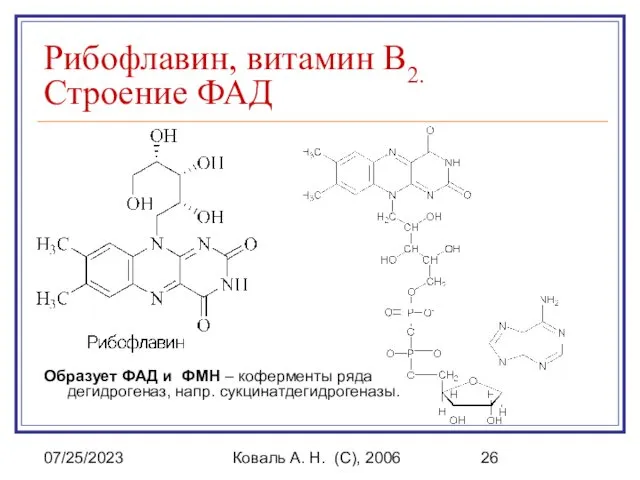
Рибофлавин, витамин B2.
Строение ФАД
Образует ФАД и
07/25/2023
Коваль А. Н. (C), 2006
Рибофлавин, витамин B2.
Строение ФАД
Образует ФАД и

07/25/2023
Коваль А. Н. (C), 2006
Пантотеновая кислота
Пантотеновая кислота – входит в состав
07/25/2023
Коваль А. Н. (C), 2006
Пантотеновая кислота
Пантотеновая кислота – входит в состав

07/25/2023
Коваль А. Н. (C), 2006
Металлы как кофакторы
Многие двухвалентные металлы (Сu2+, Zn2+,
07/25/2023
Коваль А. Н. (C), 2006
Металлы как кофакторы
Многие двухвалентные металлы (Сu2+, Zn2+,

07/25/2023
Коваль А. Н. (C), 2006
Металлы как кофакторы (прод.)
07/25/2023
Коваль А. Н. (C), 2006
Металлы как кофакторы (прод.)

07/25/2023
Коваль А. Н. (C), 2006
Структурно-функциональная организация ферментов.
Активный центр
Активный (субстратный) центр
07/25/2023
Коваль А. Н. (C), 2006
Структурно-функциональная организация ферментов.
Активный центр
Активный (субстратный) центр

07/25/2023
Коваль А. Н. (C), 2006
Структурно-функциональная организация ферментов. Строение активного центра фермента
Активный
07/25/2023
Коваль А. Н. (C), 2006
Структурно-функциональная организация ферментов. Строение активного центра фермента
Активный
07/25/2023
Коваль А. Н. (C), 2006
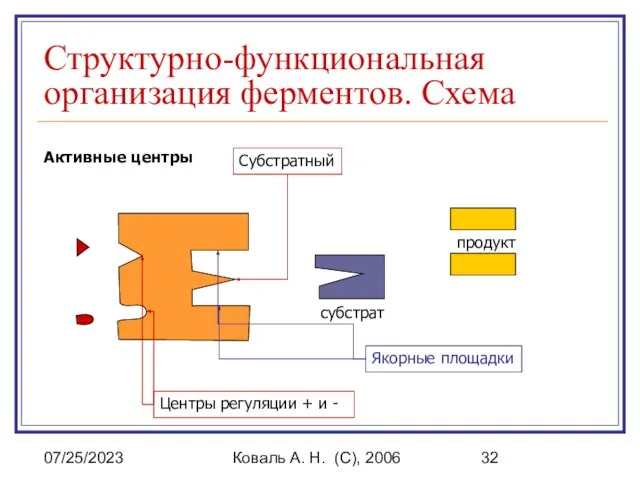
Структурно-функциональная организация ферментов. Схема
Активные центры
Якорные площадки
Субстратный
субстрат
продукт
Центры регуляции
07/25/2023
Коваль А. Н. (C), 2006
Структурно-функциональная организация ферментов. Схема
Активные центры
Якорные площадки
Субстратный
субстрат
продукт
Центры регуляции

07/25/2023
Коваль А. Н. (C), 2006
Формирование активного центра фермента
Предполагается, что формирование активного
07/25/2023
Коваль А. Н. (C), 2006
Формирование активного центра фермента
Предполагается, что формирование активного

07/25/2023
Коваль А. Н. (C), 2006
Центры аллостерической регуляции
У олигомерных ферментов (имеющих четвертичную
07/25/2023
Коваль А. Н. (C), 2006
Центры аллостерической регуляции
У олигомерных ферментов (имеющих четвертичную

07/25/2023
Коваль А. Н. (C), 2006
Аллостерические ферменты
Присоединение эффектора к аллостерическому центру приводит
07/25/2023
Коваль А. Н. (C), 2006
Аллостерические ферменты
Присоединение эффектора к аллостерическому центру приводит

07/25/2023
Коваль А. Н. (C), 2006
Спасибо за внимание!
Следующая лекция – Биохимия ферментов-2.
Механизм
07/25/2023
Коваль А. Н. (C), 2006
Спасибо за внимание!
Следующая лекция – Биохимия ферментов-2.
Механизм
 Великий учёный и хирург Николай Иванович Пирогов
Великий учёный и хирург Николай Иванович Пирогов Этот вкусный и полезный мед
Этот вкусный и полезный мед Скопа. Дрофа
Скопа. Дрофа Тема: «Класс Насекомые»
Тема: «Класс Насекомые» Дефицит элементов и внешность
Дефицит элементов и внешность Функции тонкого и толстого кишечника. Всасывание. Барьерная роль печени. Аппендицит.
Функции тонкого и толстого кишечника. Всасывание. Барьерная роль печени. Аппендицит.  Животные красной книги россии, обитающие в ивановской области
Животные красной книги россии, обитающие в ивановской области Типы взаимодействия популяций
Типы взаимодействия популяций Проект з біології на тему “Астма” учениці 9-Г класу Луцького НВК №9 Карпось Катерини
Проект з біології на тему “Астма” учениці 9-Г класу Луцького НВК №9 Карпось Катерини 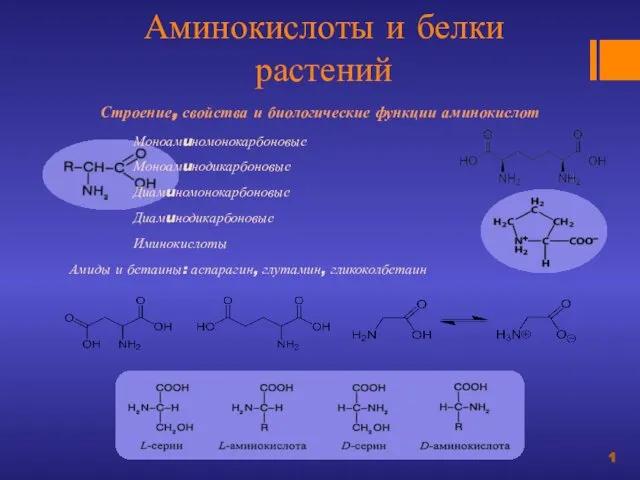 Аминокислоты и белки растений
Аминокислоты и белки растений Брачное поведение птиц
Брачное поведение птиц Біологія наука про живу природу
Біологія наука про живу природу  Тема №2 «Размножение растений»
Тема №2 «Размножение растений»  Тип Членистоногие
Тип Членистоногие Как растут овощи
Как растут овощи Класс Млекопитающие. Общая характеристика и особенности внешнего строения млекопитающих
Класс Млекопитающие. Общая характеристика и особенности внешнего строения млекопитающих Нервная система. Черепные нервы. Продолговатый мозг и мост
Нервная система. Черепные нервы. Продолговатый мозг и мост Кустарники леса
Кустарники леса Презентация на тему "Почвенная среда обитания" - скачать презентации по Биологии
Презентация на тему "Почвенная среда обитания" - скачать презентации по Биологии Тема: Выполнила: Масленникова Юлия, МКОУ Новоперуновская СОШ, ученица 6 класса Руководитель: Филяева Ольга Николаевна, МКОУ Нов
Тема: Выполнила: Масленникова Юлия, МКОУ Новоперуновская СОШ, ученица 6 класса Руководитель: Филяева Ольга Николаевна, МКОУ Нов Членистоногие. Строение паукообразных, насекомых, ракообразных
Членистоногие. Строение паукообразных, насекомых, ракообразных Механізми видоутворення
Механізми видоутворення Членистоногие (Arthropoda) – паразиты человека и животных
Членистоногие (Arthropoda) – паразиты человека и животных Выращивание огурца в весенней пленочной теплице. (6 класс, технология)
Выращивание огурца в весенней пленочной теплице. (6 класс, технология) Аппарат Гольджи (комплекс Гольджи)
Аппарат Гольджи (комплекс Гольджи) Лети, лети, лепесток
Лети, лети, лепесток Строение и структура нуклеиновых кислот. Лекция №3А
Строение и структура нуклеиновых кислот. Лекция №3А Этапы эволюции человека
Этапы эволюции человека