- клеточные мембраны

Содержание
- 2. План лекции: 1. Ультраструктура клеточных мембран. 2. Молекулярное строение цитоплазматических мембран. 3. Физическая природа сил взаимодействия
- 3. Важнейшими условиями существования клетки является: 1. Автономность по отношению к окружающей среде. 2. Связь с окружающей
- 4. Клеточная мембра́на (также цитолемма, плазмолемма, или плазматическая мембрана) — эластическая молекулярная структура, состоящая из белков и
- 5. История исследования В 1925 году Гортер и Грендель с помощью осмотического удара получили так называемые «тени»
- 6. Молекулярное строение клеточных мембран - В 1902 г. - Овертон предположил, что мембрана состоит из тонкого
- 7. Молекулярное строение клеточных мембран В 1935 Даниэлли и Доусон предложили модель строения мембраны, получившей название «сэндвич».
- 8. Молекулярное строение клеточных мембран В 1959 г. Дж. Робертсоном была выдвинута теория унитарной мембраны, состоящей из
- 9. Все это привело к созданию в 1972 году Сингером (S. Jonathan Singer) и Николсоном (Garth L.
- 10. В основе биомембраны – двойной слой: 1. Амфифильные липиды (липидный слой). Молекула мембранного липида имеет гидрофильную
- 11. Структура и состав биомембран Мембраны состоят из липидов трёх классов: фосфолипиды, гликолипиды и холестерол. Фосфолипиды и
- 12. Липидный слой – это основная структура мембраны, которая создает относительно непроницаемый барьер для большинства водорастворимых молекул.
- 13. У фосфолипидов (ФЛ) в состав «головки» входят остатки азотистого основания (холина, коламина или серина), фосфатной группы
- 14. Строение фосфолипида
- 15. Сфинголипиды (СЛ) – вместо глицерина и одной из жирной кислот, включают сфингозин (сфингенин) – 2-х атомный
- 16. Гликолипиды - сложные липиды, образующиеся в результате соединения липидов с углеводами. В молекулах гликолипидов есть полярные
- 17. Холестери́н (др.-греч. χολή — желчь и στερεός — твёрдый) — органическое соединение, природный жирный (липофильный) спирт,
- 18. Липосомы – это нано частицы с пузырьковой структурой. Липосомы самопроизвольно образуются в смесях фосфолипидов (сложных липидов)
- 19. Мембранные липиды – амфипатические молекулы, и в водной среде большинство из них самопроизвольно образует бислои. Именно
- 20. Важную часть мембраны составляют белки, пронизывающие её и отвечающие за разнообразные свойства мембран. Их состав и
- 21. Белки мембраны клеток представлены, в основном, гликопротеинами. Различают: •интегральные белки, проникающие через всю толщу мембраны и
- 22. Периферические белки почти все функционируют как энзимы (ацетилхолинестераза, кислая и шелочная фосфатазы и др.). Но некоторые
- 23. Схема расположения белков в примембранном цитоскелете эритроцитов. 1 — спектрин; 2 — анкирин; 3 — белок
- 24. Спектрин является основным белком цитоскелета, составляющим двумерную сеть, к которой прикрепляется актин. Актин образует микрофиламенты, представляющие
- 25. Гликофорин – интегральный белок. Он пронизывает всю мембрану и с обей сторон выступает над ее поверхностью.
- 26. Виды мембранных белков Структурные Транспортные Белки, обеспечивающие межклеточное взаимодействие Белки, участвующие в передачи сигналов
- 27. Структурные белки Придают клетке и органеллам определенную форму Придают мембране механические свойства (эластичность и т.д.) Обеспечивают
- 28. Транспортные белки Создают устойчивые транспортные потоки определенных веществ через мембраны Транспорт ионов приводит к возникновению трансмембранного
- 29. Белки, обеспечивающие межклеточное взаимодействие Адгезивные белки – для связывания клеток друг с другом или неклеточными структурами
- 30. Белки, участвующие в передачи сигналов Рецепторные белки Белки эффекторного устройства Фермент инактивации медиатора.
- 31. Клеточные мембраны часто асимметричны, то есть слои отличаются по составу липидов, в наружном содержатся преимущественно фосфатидилинозитол,
- 32. Гидрофобные и гидрофильные силы взаимодействия В подержание структуры мембраны большую роль играют гидрофобные и гидрофильные силы
- 33. Гидрофильные силы взаимодействия способствуют удержанию белков на поверхности бислоя фосфолипидов. Таким образом, гидрофильные и гидрофобные силы
- 34. Основные свойства мембран Замкнутость Латеральная подвижность Асимметрия
- 35. Замкнутость Липидные бислои (и мембраны) всегда самостоятельно замыкаются на себе с образованием полностью отграниченных отсеков. При
- 36. Латеральная подвижность Компоненты мембраны могут перемещаться в пределах своего слоя. Мембраны обладают свойствами двумерных жидкостей. По
- 37. Асимметрия Наружная и внутренняя поверхности мембраны различаются по своему составу: Углеводные компоненты находятся с внешней поверхности
- 38. Свойства мембраны 1. Проницаемость. 2. Полупроницаемость. 3. Избирательная проницаемость. 4. Активная проницаемость. 5. Управляемая проницаемость. Как
- 39. 10. Раздражимость. 11. Каталитическая ферментативная активность. Ферменты могут быть встроены в мембрану или связаны с её
- 40. Основные функции биологических мембран: 1. Барьерная – обеспечивает селективный регулируемый, пассивный и активный обмен веществ 2.
- 41. Основные функции биологических мембран: 3. Механическая – обеспечивает прочность и автономность клетки, внутриклеточных структур 4. Энергетическая
- 42. Основные функции биологических мембран: 5. Рецепторная (механическая, акустическая, обонятельная, зрительная, химическая, терморецепция и тд.) 6. Генерация
- 43. Существуют 4 способа ассоциации белков с липидным слоем: интегральные белки, пронизывающие мембрану насквозь поверхностные белки, погруженные
- 44. Методы изучения и исследования биомембран Метод рентгеноструктурного анализа – основан на явлении дифракции. Дифракция наблюдается в
- 45. Методы изучения и исследования биомембран 1. Метод рентгеновского рассеивания нейтронов. Метод рассеивания нейтронов под малыми углами
- 46. Метод моделирования мембран В 1917 г. Лэнгмюр разработал технику получения модели мономолекулярного слоя фосфолипидов – модель
- 47. Метод моделирования мембран В 1962 Мюллер и Рудин разработали способ получения больших двуслойных мембран (d до
- 48. Метод моделирования мембран Получение липосом - замкнутые мембранные пузырьки, содержащие водную фазу внутри и находящиеся в
- 49. Перенос веществ через мембраны Низкомолекулярные соединения: А) простая диффузия Б) облегченная диффузия В) активный транспорт
- 50. Облегченная диффузия Молекула глюкозы связывается переносчиком на наружной поверхности плазматической мембраны. Происходит конформационное изменение, и центр
- 51. Типы (виды) облегчённой диффузии с участием переносчиков (транслоказ). S1, S2- разные молекулы. Некоторые транслоказы могут переносить
- 52. Примером транслоказы, работающей по механизму пассивного антипорта, может служить анионный переносчик мембраны эритроцитов. Пассивный антипорт анионов
- 53. Строение и функционирование Nа+,К+-АТФ-азы плазматической мембраны. 1 - три иона натрия связываются специфическим центром транслоказы; 2
- 54. Вторичный активный транспорт Перенос некоторых растворимых веществ против градиента концентрации зависит от одновременного или последовательного переноса
- 55. Перенос через мембрану макромолекул и частиц: эндоцитоз и экзоцитоз Перенос вещества из среды в клетку вместе
- 56. Последовательность событий при образовании окаймлённого пузырька из окаймлённой ямки
- 57. Положение рецепторов ЛПНП в цитоплазматической мембране А - положение рецепторов ЛПНП в окаймлённой ямке; Б -
- 58. G-белки Сигнальные G-белки являются универсальными посредниками при передаче гормональных сигналов от рецепторов клеточной мембраны к эффекторным
- 60. Патологии клеточных мембран нарушения мембранного транспорта, изменения проницаемости мембран, изменения коммуникации клеток и их "узнавания", изменения
- 62. Скачать презентацию

План лекции:
1. Ультраструктура клеточных мембран.
2. Молекулярное строение цитоплазматических мембран.
3. Физическая природа
План лекции:
1. Ультраструктура клеточных мембран.
2. Молекулярное строение цитоплазматических мембран.
3. Физическая природа

Важнейшими условиями существования клетки является:
1. Автономность по отношению к окружающей
Важнейшими условиями существования клетки является:
1. Автономность по отношению к окружающей

Клеточная мембра́на (также цитолемма, плазмолемма, или плазматическая мембрана) — эластическая
Клеточная мембра́на (также цитолемма, плазмолемма, или плазматическая мембрана) — эластическая

История исследования
В 1925 году Гортер и Грендель с помощью осмотического удара
История исследования
В 1925 году Гортер и Грендель с помощью осмотического удара

Молекулярное строение клеточных мембран
- В 1902 г. - Овертон
Молекулярное строение клеточных мембран
- В 1902 г. - Овертон

Молекулярное строение клеточных мембран
В 1935 Даниэлли и Доусон предложили модель строения
Молекулярное строение клеточных мембран
В 1935 Даниэлли и Доусон предложили модель строения

Молекулярное строение клеточных мембран
В 1959 г. Дж. Робертсоном была выдвинута теория
Молекулярное строение клеточных мембран
В 1959 г. Дж. Робертсоном была выдвинута теория

Все это привело к созданию в 1972 году Сингером (S. Jonathan
Все это привело к созданию в 1972 году Сингером (S. Jonathan
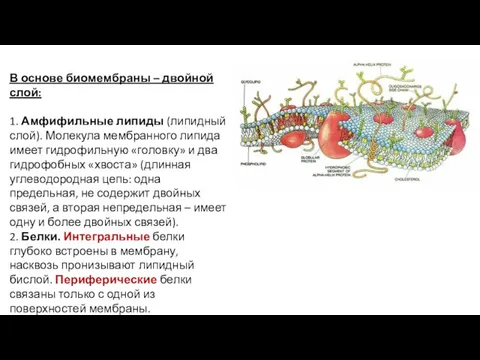
В основе биомембраны – двойной слой:
1. Амфифильные липиды (липидный слой). Молекула
В основе биомембраны – двойной слой:
1. Амфифильные липиды (липидный слой). Молекула

Структура и состав биомембран
Мембраны состоят из липидов трёх классов: фосфолипиды, гликолипиды
Структура и состав биомембран
Мембраны состоят из липидов трёх классов: фосфолипиды, гликолипиды

Липидный слой – это основная структура мембраны, которая создает относительно непроницаемый
Липидный слой – это основная структура мембраны, которая создает относительно непроницаемый

У фосфолипидов (ФЛ) в состав «головки» входят остатки азотистого основания (холина,
У фосфолипидов (ФЛ) в состав «головки» входят остатки азотистого основания (холина,
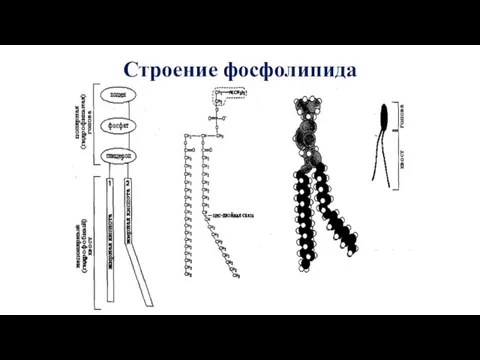
Строение фосфолипида
Строение фосфолипида

Сфинголипиды (СЛ) – вместо глицерина и одной из жирной кислот, включают
Сфинголипиды (СЛ) – вместо глицерина и одной из жирной кислот, включают

Гликолипиды - сложные липиды, образующиеся в результате соединения липидов с углеводами.
Гликолипиды - сложные липиды, образующиеся в результате соединения липидов с углеводами.

Холестери́н (др.-греч. χολή — желчь и στερεός — твёрдый) — органическое
Холестери́н (др.-греч. χολή — желчь и στερεός — твёрдый) — органическое

Липосомы – это нано частицы с пузырьковой структурой. Липосомы самопроизвольно образуются
Липосомы – это нано частицы с пузырьковой структурой. Липосомы самопроизвольно образуются

Мембранные липиды – амфипатические молекулы, и в водной среде большинство из
Мембранные липиды – амфипатические молекулы, и в водной среде большинство из

Важную часть мембраны составляют белки, пронизывающие её и отвечающие за разнообразные
Важную часть мембраны составляют белки, пронизывающие её и отвечающие за разнообразные

Белки мембраны клеток представлены, в основном, гликопротеинами.
Различают:
•интегральные белки, проникающие через
Белки мембраны клеток представлены, в основном, гликопротеинами.
Различают:
•интегральные белки, проникающие через

Периферические белки почти все функционируют как энзимы (ацетилхолинестераза, кислая и шелочная
Периферические белки почти все функционируют как энзимы (ацетилхолинестераза, кислая и шелочная

Схема расположения белков в примембранном цитоскелете эритроцитов.
1 — спектрин; 2 —
Схема расположения белков в примембранном цитоскелете эритроцитов.
1 — спектрин; 2 —

Спектрин является основным белком цитоскелета, составляющим двумерную сеть, к которой прикрепляется
Спектрин является основным белком цитоскелета, составляющим двумерную сеть, к которой прикрепляется

Гликофорин – интегральный белок. Он пронизывает всю мембрану и с обей
Гликофорин – интегральный белок. Он пронизывает всю мембрану и с обей

Виды мембранных белков
Структурные
Транспортные
Белки, обеспечивающие межклеточное взаимодействие
Белки, участвующие в передачи сигналов
Виды мембранных белков
Структурные
Транспортные
Белки, обеспечивающие межклеточное взаимодействие
Белки, участвующие в передачи сигналов

Структурные белки
Придают клетке и органеллам определенную форму
Придают мембране механические свойства (эластичность
Структурные белки
Придают клетке и органеллам определенную форму
Придают мембране механические свойства (эластичность

Транспортные белки
Создают устойчивые транспортные потоки определенных веществ через мембраны
Транспорт ионов приводит
Транспортные белки
Создают устойчивые транспортные потоки определенных веществ через мембраны
Транспорт ионов приводит

Белки, обеспечивающие межклеточное взаимодействие
Адгезивные белки – для связывания клеток друг с
Белки, обеспечивающие межклеточное взаимодействие
Адгезивные белки – для связывания клеток друг с

Белки, участвующие в передачи сигналов
Рецепторные белки
Белки эффекторного устройства
Фермент инактивации медиатора.
Белки, участвующие в передачи сигналов
Рецепторные белки
Белки эффекторного устройства
Фермент инактивации медиатора.

Клеточные мембраны часто асимметричны, то есть слои отличаются по составу липидов,
Клеточные мембраны часто асимметричны, то есть слои отличаются по составу липидов,

Гидрофобные и гидрофильные силы взаимодействия
В подержание структуры мембраны большую роль играют
Гидрофобные и гидрофильные силы взаимодействия
В подержание структуры мембраны большую роль играют

Гидрофильные силы взаимодействия способствуют удержанию белков на поверхности бислоя фосфолипидов. Таким
Гидрофильные силы взаимодействия способствуют удержанию белков на поверхности бислоя фосфолипидов. Таким

Основные свойства мембран
Замкнутость
Латеральная подвижность
Асимметрия
Основные свойства мембран
Замкнутость
Латеральная подвижность
Асимметрия

Замкнутость
Липидные бислои (и мембраны) всегда самостоятельно замыкаются на себе с образованием
Замкнутость
Липидные бислои (и мембраны) всегда самостоятельно замыкаются на себе с образованием

Латеральная подвижность
Компоненты мембраны могут перемещаться в пределах своего слоя.
Мембраны обладают свойствами
Латеральная подвижность
Компоненты мембраны могут перемещаться в пределах своего слоя.
Мембраны обладают свойствами

Асимметрия
Наружная и внутренняя поверхности мембраны различаются по своему составу:
Углеводные компоненты находятся
Асимметрия
Наружная и внутренняя поверхности мембраны различаются по своему составу:
Углеводные компоненты находятся

Свойства мембраны
1. Проницаемость.
2. Полупроницаемость.
3. Избирательная проницаемость.
4. Активная проницаемость.
5. Управляемая проницаемость.
Как видим,
Свойства мембраны
1. Проницаемость.
2. Полупроницаемость.
3. Избирательная проницаемость.
4. Активная проницаемость.
5. Управляемая проницаемость.
Как видим,

10. Раздражимость.
11. Каталитическая ферментативная активность. Ферменты могут быть встроены в мембрану
10. Раздражимость.
11. Каталитическая ферментативная активность. Ферменты могут быть встроены в мембрану

Основные функции биологических мембран:
1. Барьерная – обеспечивает селективный регулируемый, пассивный и
Основные функции биологических мембран:
1. Барьерная – обеспечивает селективный регулируемый, пассивный и

Основные функции биологических мембран:
3. Механическая – обеспечивает прочность и автономность клетки,
Основные функции биологических мембран:
3. Механическая – обеспечивает прочность и автономность клетки,

Основные функции биологических мембран:
5. Рецепторная (механическая, акустическая, обонятельная, зрительная, химическая, терморецепция
Основные функции биологических мембран:
5. Рецепторная (механическая, акустическая, обонятельная, зрительная, химическая, терморецепция

Существуют 4 способа ассоциации белков с липидным слоем:
интегральные белки, пронизывающие мембрану
Существуют 4 способа ассоциации белков с липидным слоем:
интегральные белки, пронизывающие мембрану

Методы изучения и исследования биомембран
Метод рентгеноструктурного анализа – основан на явлении
Методы изучения и исследования биомембран
Метод рентгеноструктурного анализа – основан на явлении

Методы изучения и исследования биомембран
1. Метод рентгеновского рассеивания нейтронов.
Метод
Методы изучения и исследования биомембран
1. Метод рентгеновского рассеивания нейтронов.
Метод

Метод моделирования мембран
В 1917 г. Лэнгмюр разработал технику получения модели
Метод моделирования мембран
В 1917 г. Лэнгмюр разработал технику получения модели
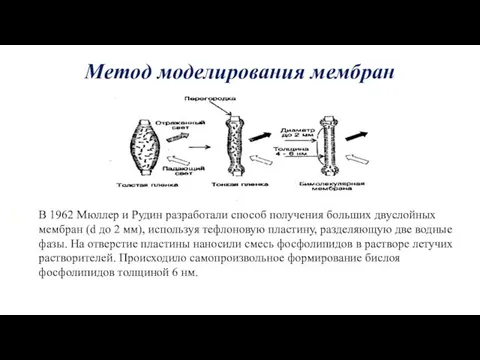
Метод моделирования мембран
В 1962 Мюллер и Рудин разработали способ получения больших
Метод моделирования мембран
В 1962 Мюллер и Рудин разработали способ получения больших

Метод моделирования мембран
Получение липосом - замкнутые мембранные пузырьки, содержащие водную фазу
Метод моделирования мембран
Получение липосом - замкнутые мембранные пузырьки, содержащие водную фазу

Перенос веществ через мембраны
Низкомолекулярные соединения:
А) простая диффузия
Б) облегченная диффузия
В) активный транспорт
Перенос веществ через мембраны
Низкомолекулярные соединения:
А) простая диффузия
Б) облегченная диффузия
В) активный транспорт

Облегченная диффузия
Молекула глюкозы связывается переносчиком на наружной поверхности плазматической мембраны. Происходит
Облегченная диффузия
Молекула глюкозы связывается переносчиком на наружной поверхности плазматической мембраны. Происходит
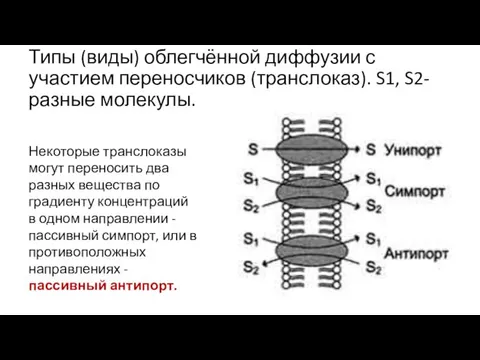
Типы (виды) облегчённой диффузии с участием переносчиков (транслоказ). S1, S2- разные
Типы (виды) облегчённой диффузии с участием переносчиков (транслоказ). S1, S2- разные

Примером транслоказы, работающей по механизму пассивного антипорта, может служить анионный переносчик
Примером транслоказы, работающей по механизму пассивного антипорта, может служить анионный переносчик

Строение и функционирование Nа+,К+-АТФ-азы плазматической мембраны.
1 - три иона натрия связываются
Строение и функционирование Nа+,К+-АТФ-азы плазматической мембраны.
1 - три иона натрия связываются

Вторичный активный транспорт
Перенос некоторых растворимых веществ против градиента концентрации зависит от
Вторичный активный транспорт
Перенос некоторых растворимых веществ против градиента концентрации зависит от

Перенос через мембрану макромолекул и частиц: эндоцитоз и экзоцитоз
Перенос вещества из
Перенос через мембрану макромолекул и частиц: эндоцитоз и экзоцитоз
Перенос вещества из
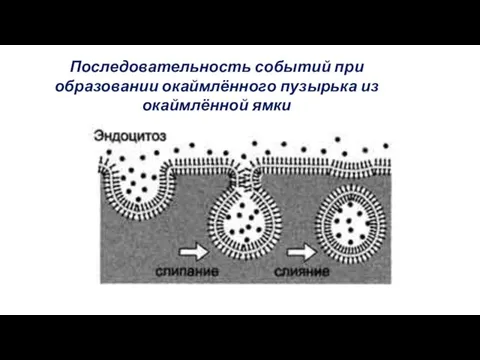
Последовательность событий при образовании окаймлённого пузырька из окаймлённой ямки
Последовательность событий при образовании окаймлённого пузырька из окаймлённой ямки
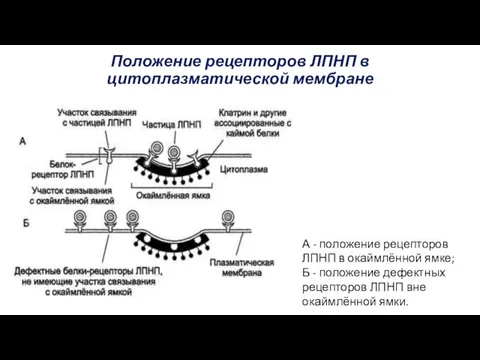
Положение рецепторов ЛПНП в цитоплазматической мембране
А - положение рецепторов ЛПНП в
Положение рецепторов ЛПНП в цитоплазматической мембране
А - положение рецепторов ЛПНП в

G-белки
Сигнальные G-белки являются универсальными посредниками при передаче гормональных сигналов от рецепторов
G-белки
Сигнальные G-белки являются универсальными посредниками при передаче гормональных сигналов от рецепторов


Патологии клеточных мембран
нарушения мембранного транспорта,
изменения проницаемости мембран,
изменения коммуникации клеток
Патологии клеточных мембран
нарушения мембранного транспорта,
изменения проницаемости мембран,
изменения коммуникации клеток
 Земноводные - амфибии
Земноводные - амфибии  Биологические особенности развития риса в Кызылординской области
Биологические особенности развития риса в Кызылординской области Дыхание, его значение. Строение и функции органов дыхания. Урок биологии 8 класс Л.К.Юшкова
Дыхание, его значение. Строение и функции органов дыхания. Урок биологии 8 класс Л.К.Юшкова Нервная система. Общие принципы организации нервной системы
Нервная система. Общие принципы организации нервной системы Лекция 8 Классификация экосистем
Лекция 8 Классификация экосистем Учитель биологии: Галковская И.И 7 класс (По учебнику Константинова В.М., Бабенко В.Г., Кучменко В.С.) 2010 год
Учитель биологии: Галковская И.И 7 класс (По учебнику Константинова В.М., Бабенко В.Г., Кучменко В.С.) 2010 год Животный мир Челябинской области
Животный мир Челябинской области Бесполое размножение организмов – формы и значение в природе
Бесполое размножение организмов – формы и значение в природе Презентация на тему "Основні небезпеки навколишнього середовища" - скачать бесплатно презентации по Биологии
Презентация на тему "Основні небезпеки навколишнього середовища" - скачать бесплатно презентации по Биологии Бөлме өсімдіктері
Бөлме өсімдіктері Черты биологического прогресса насекомых Автор: Елисеева Марина Александровна учитель биологии МБОУ СОШ № 200 г. Екатеринбург
Черты биологического прогресса насекомых Автор: Елисеева Марина Александровна учитель биологии МБОУ СОШ № 200 г. Екатеринбург Фрактальные свойства биоэлектрической активности мозга человека
Фрактальные свойства биоэлектрической активности мозга человека Рослини хижаки
Рослини хижаки Семейство крестоцветные
Семейство крестоцветные Кость как орган. Соединения костей
Кость как орган. Соединения костей Презентация на тему Клеточная инженерия
Презентация на тему Клеточная инженерия  Горячее или холодное
Горячее или холодное Значение нервной системы. Строение нервной системы. Спинной мозг.
Значение нервной системы. Строение нервной системы. Спинной мозг.  Опорно-двигательный аппарат. Мышечная система. Осанка
Опорно-двигательный аппарат. Мышечная система. Осанка Происхождение человека
Происхождение человека Моногибридное скрещивание1
Моногибридное скрещивание1 Движение. Где можно наблюдать движение
Движение. Где можно наблюдать движение Подготовила Лейн Ирина 5В
Подготовила Лейн Ирина 5В Лист и видоизменения листа
Лист и видоизменения листа Соматотропин - гормон роста
Соматотропин - гормон роста Кулинарная обработка и сохранение витаминов!
Кулинарная обработка и сохранение витаминов! Методы исследования генетики человека
Методы исследования генетики человека Изучение жирафов
Изучение жирафов