- Матричный синтез информационных макромолекул. Типы переноса генетической информации. Репликация ДНК

Содержание
- 2. Основным свойством нуклеиновых кислот является - хранение и реализация генетической ин-формации в процессе размножения и функциони-рования
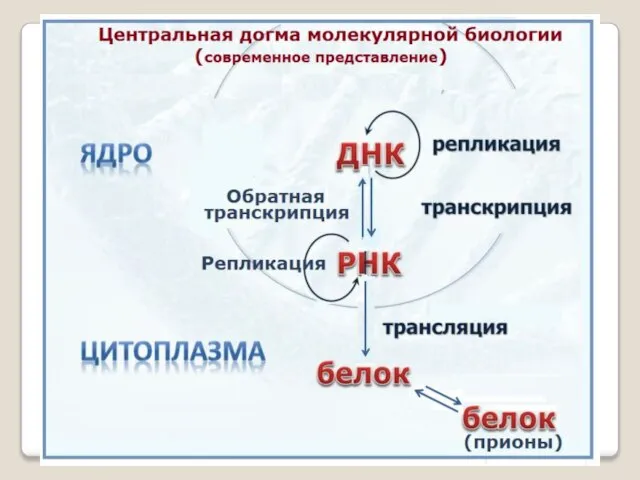
- 4. В клетке имеется направленный поток информации от ДНК, кото-рая представляет собой исходный генетический текст, состоящий из
- 5. По мнению Ф.Крика никогда не может быть потока информации от белка к нуклеи-новым кислотам, а внутри
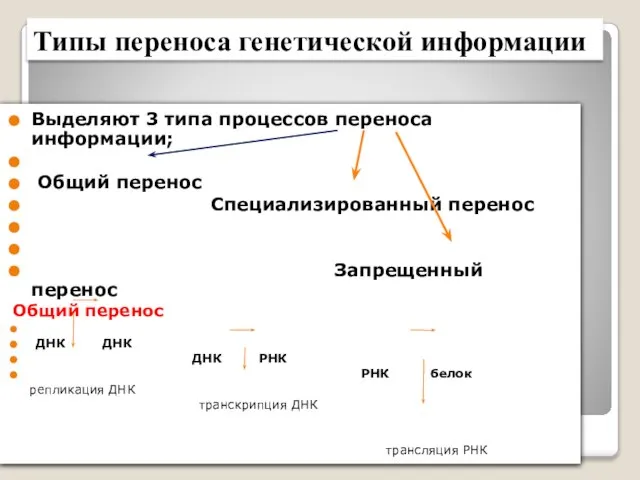
- 8. Типы переноса генетической информации Выделяют 3 типа процессов переноса информации; Общий перенос Специализированный перенос Запрещенный перенос
- 9. Специализированный перенос происходит в клетках при особых обстоятельствах и включает следующие процессы: (РНК→РНК); ДНК → белок
- 10. Запрещённый перенос информации Подразумевает перевод информации с аминокислотной последовательности белка на последовательность нуклеотидов белок→ДНК белок→РНК белок→белок
- 11. Прио́ны (англ. prion от protein — «белок» и infection — «инфекция», слово предложено в 1982 году
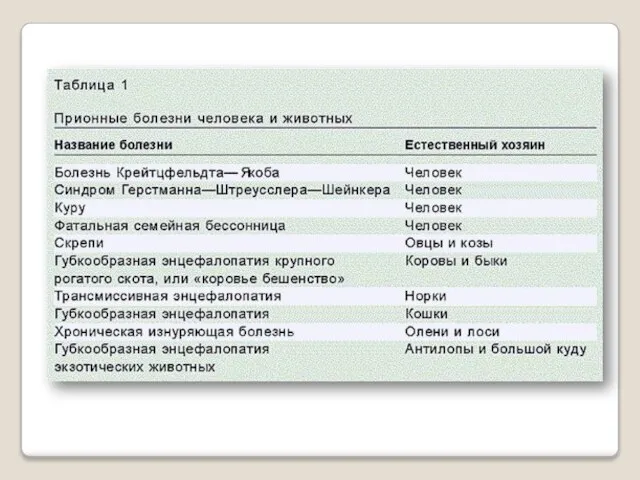
- 12. Все известные прионные заболевания млекопитающих вызываются белком PrP. Его форма с нормальной третичной струк-турой называется PrPC
- 14. Пути возникновения : в случае прямого заражения, наследственно спорадически (спонтанно) комбинация этих факторов.
- 15. Репликация ДНК, важнейший из внутриядерных процессов, обеспечивающий максимальную точность передачи информации от родительских молекул ДНК к
- 16. а - Полуконсервативная репликация б - Консервативная репликация. в - Дисперсная репликация. Модели репликации ДНК
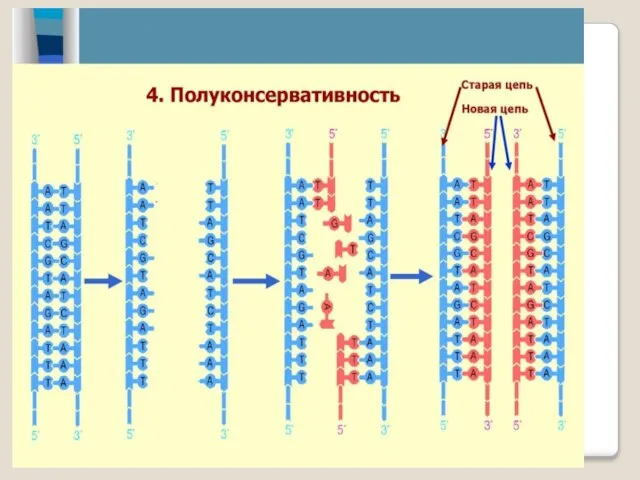
- 17. В полуконсервативной модели репликации ДНК две ро-дительские нити отделяются друг от друга и каждая из них
- 18. Каждая молекула ДНК состоит из одной цепи исходной родительской молекулы и одной вновь синтезированной цепи. Такой
- 19. Полуконсервативная репликация ДНК При полуконсерва-тивной репликации на материнских цепях родительской моле-кулы ДНК син тези-руются дочерние нити.
- 30. Репликация может начинаться не с любого участка ДНК, а со строго определённого, называемого точка начала репликации
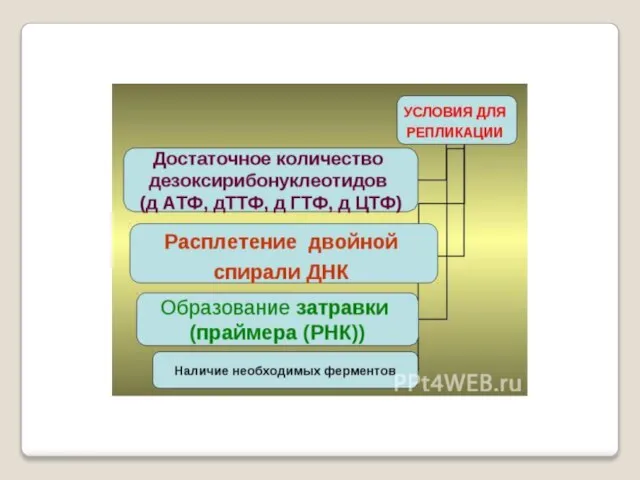
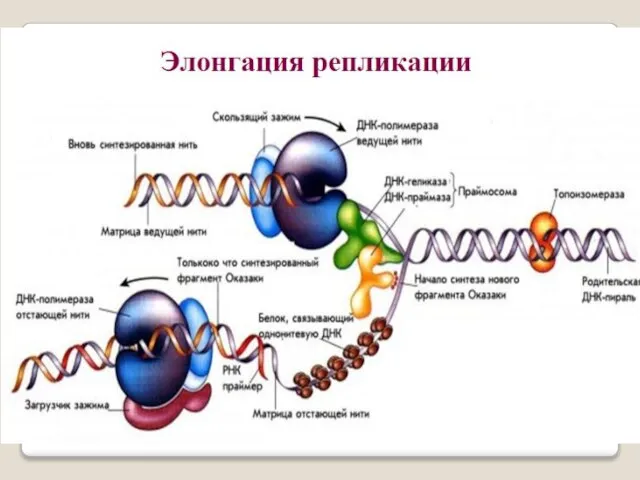
- 32. Репликацию можно разделить на 3 этапа: инициация - образование реп-ликативной вилки , элонгация - синтез новых
- 33. Инициация репликации
- 34. Фермент хеликаза разделяет цепи двухцепочечной молекулы ДНК или внутримолекулярные связи в молекулах РНК, используя энергию гидролиза
- 35. Топоизомера́зы - ферменты, которые расплетают спирализо- ванные молекулы ДНК путём внесения одно- или двуцепочеч-ных разрывов с
- 39. Белки, связывающие одноцепочечную ДНК (англ. Single-strand binding pro-tein, SSB) — связывают од-ноцепочечные фрагменты ДНК, и предотвращают
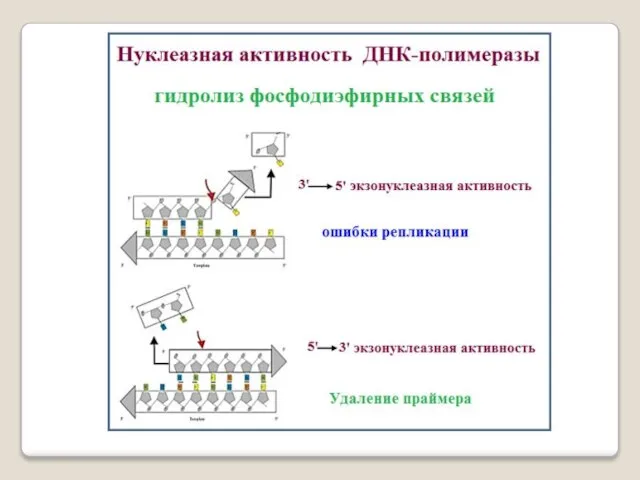
- 41. Репликация ДНК осуществляется ДНК-зависимыми ДНК-полимеразами. Субстратами и источниками энергии для синтеза продукта служат 4 макроэргических соединения
- 42. ДНК-полимераза — фермент, участвующий в репликации ДНК. Ферменты этого класса катализ-руют полимеризацию дезоксирибонуклеотидов вдоль цепочки нуклеотидов
- 46. У прокариот элонгацию осу- ществляют ДНК-полимеразы I,II,III, обозначаемые как pol I; pol II; pol III. ДНК-полимераза
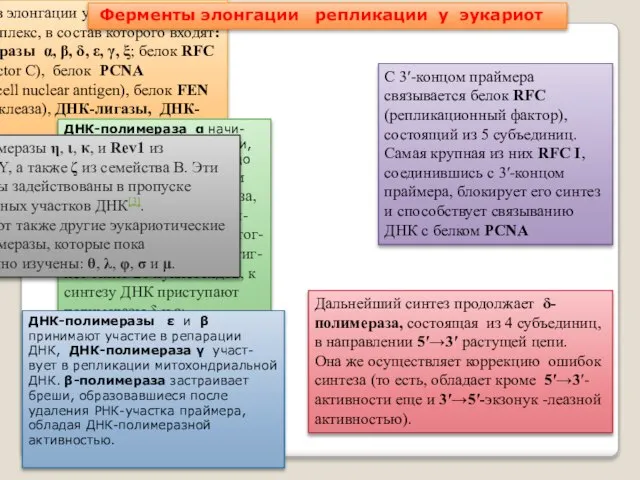
- 47. У эукариот в элонгации участвует сложный белковый комплекс, в состав которого входят: ДНК-полимеразы α, β, δ,
- 48. Ни одна эукариотическая полимераза не может отщеплять праймеры, то есть не обладает 5’-3’-экзонуклеазным действием. Эту функцию
- 49. Прокариотические ДНК-полимеразы У бактерий обнаружено пять ДНК-полимераз: ДНК-полимераза I задействована в восстановлении ДНК, обладает и 5'-3',
- 50. Три полимеразы (pol I, pol II и pol III) обладают полимеризующей 5 ′ 3′ активностью и
- 51. Белки́ скользя́щего зажима, DNA clamp) — белки, которые выполняют функцию усилителя действия ферментов при репликации ДНК.
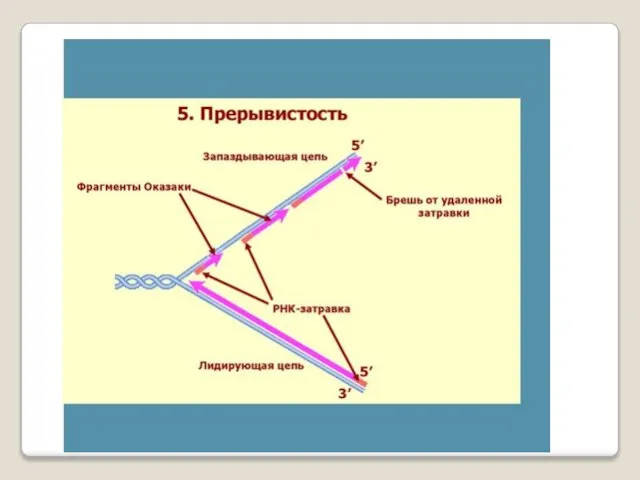
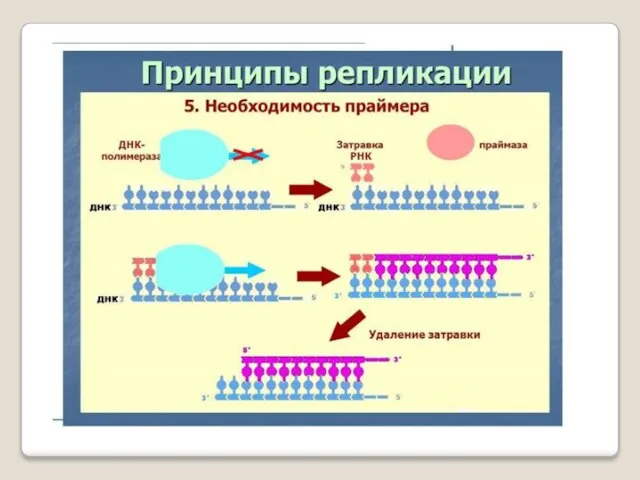
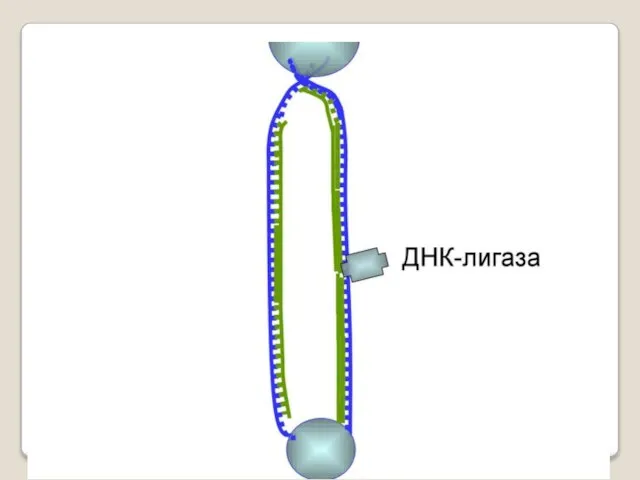
- 54. ДНК-праймаза — это фермент РНК-полимераза, который принимает участие в репликации ДНК. Праймаза синтезирует короткий фрагмент РНК,
- 71. Теломераза — фермент, добавляющий особые повторяющиеся последовательности ДНК (TTAGGG у позвоночных) к 3'-концу цепи ДНК на
- 72. А.М. Оловников в 1971 году высказал гипотезу о том, что потеря концевых последовательностей ДНК вследс-твие их
- 73. ДНК-полимеразы, синтезируя дочернюю цепь ДНК, прочитывают родительскую цепь в направлении от ее 3конца → к 5'-концу.
- 79. Скачать презентацию

Основным свойством нуклеиновых кислот является - хранение и реализация генетической
Основным свойством нуклеиновых кислот является - хранение и реализация генетической


В клетке имеется направленный поток информации от ДНК, кото-рая представляет
В клетке имеется направленный поток информации от ДНК, кото-рая представляет

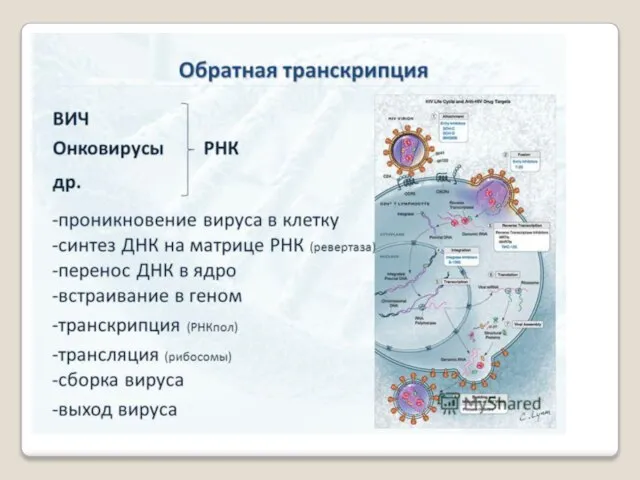
По мнению Ф.Крика никогда не может быть потока информации от
По мнению Ф.Крика никогда не может быть потока информации от
Типы переноса генетической информации
Выделяют 3 типа процессов переноса информации;
Общий перенос
Типы переноса генетической информации
Выделяют 3 типа процессов переноса информации;
Общий перенос

Специализированный перенос происходит в клетках при особых обстоятельствах и включает следующие
Специализированный перенос происходит в клетках при особых обстоятельствах и включает следующие

Запрещённый перенос информации
Подразумевает перевод информации с аминокислотной последовательности белка
Запрещённый перенос информации
Подразумевает перевод информации с аминокислотной последовательности белка

Прио́ны (англ. prion от protein — «белок» и infection — «инфекция», слово предложено в 1982 году Стенли Прузинером) —
Прио́ны (англ. prion от protein — «белок» и infection — «инфекция», слово предложено в 1982 году Стенли Прузинером) —

Все известные прионные заболевания млекопитающих вызываются белком PrP. Его форма с
Все известные прионные заболевания млекопитающих вызываются белком PrP. Его форма с

Пути возникновения :
в случае прямого заражения,
наследственно
спорадически (спонтанно)
комбинация этих
Пути возникновения :
в случае прямого заражения,
наследственно
спорадически (спонтанно)
комбинация этих
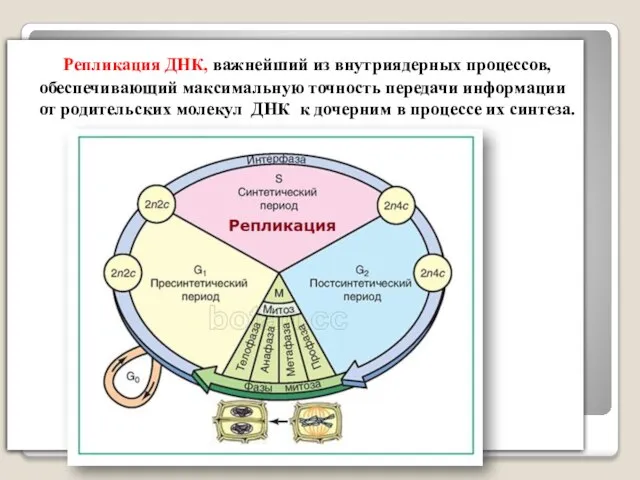
Репликация ДНК, важнейший из внутриядерных процессов, обеспечивающий максимальную точность передачи
Репликация ДНК, важнейший из внутриядерных процессов, обеспечивающий максимальную точность передачи
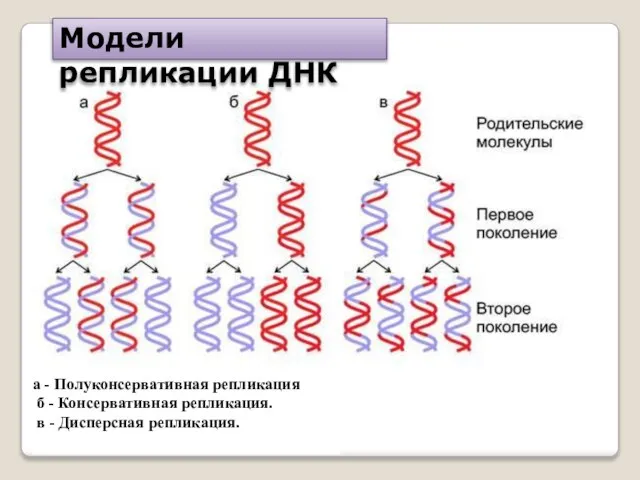
а - Полуконсервативная репликация
б - Консервативная репликация.
в - Дисперсная
а - Полуконсервативная репликация
б - Консервативная репликация.
в - Дисперсная

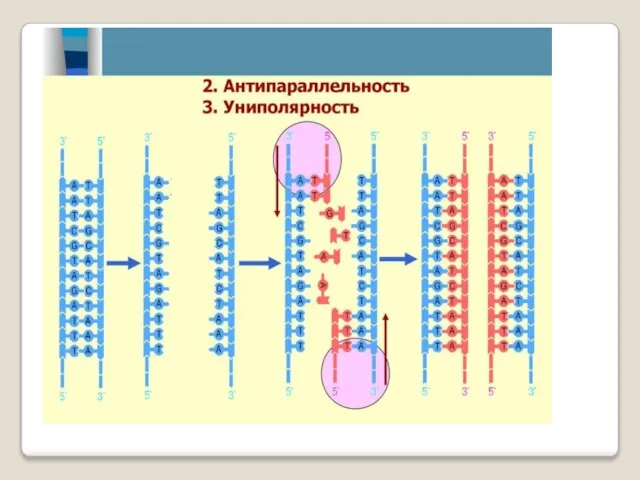
В полуконсервативной модели репликации ДНК две ро-дительские нити отделяются друг от
В полуконсервативной модели репликации ДНК две ро-дительские нити отделяются друг от

Каждая молекула ДНК состоит из одной цепи исходной родительской молекулы и
Каждая молекула ДНК состоит из одной цепи исходной родительской молекулы и
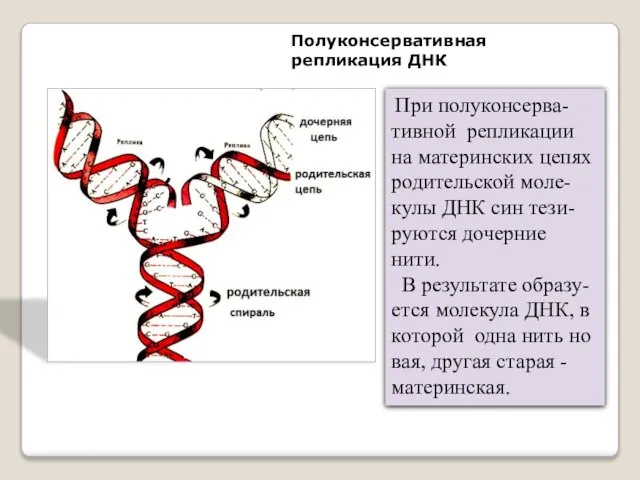
Полуконсервативная репликация ДНК
При полуконсерва-тивной репликации на материнских цепях родительской моле-кулы
Полуконсервативная репликация ДНК
При полуконсерва-тивной репликации на материнских цепях родительской моле-кулы

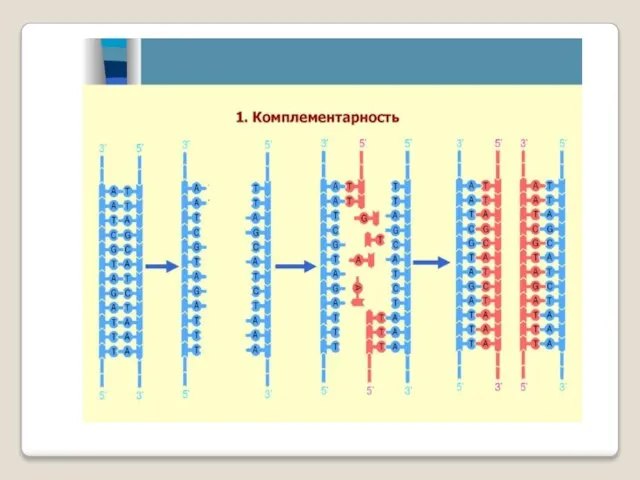



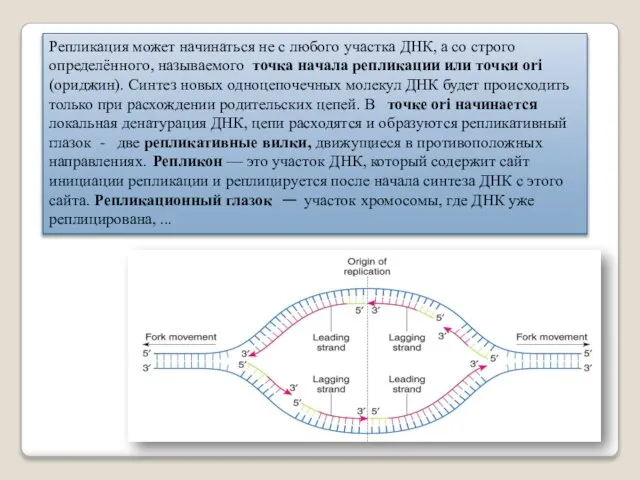
Репликация может начинаться не с любого участка ДНК, а со строго
Репликация может начинаться не с любого участка ДНК, а со строго
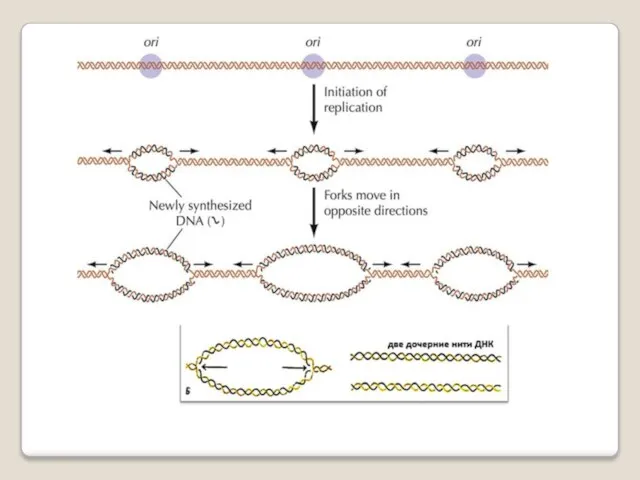

Репликацию можно разделить на 3 этапа:
инициация - образование реп-ликативной
Репликацию можно разделить на 3 этапа:
инициация - образование реп-ликативной
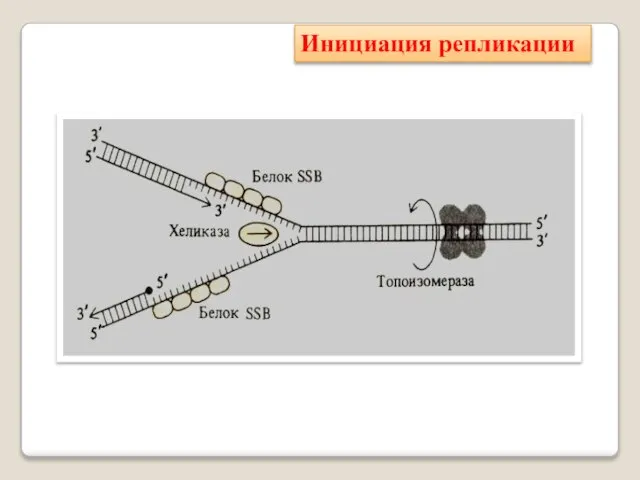
Инициация репликации
Инициация репликации
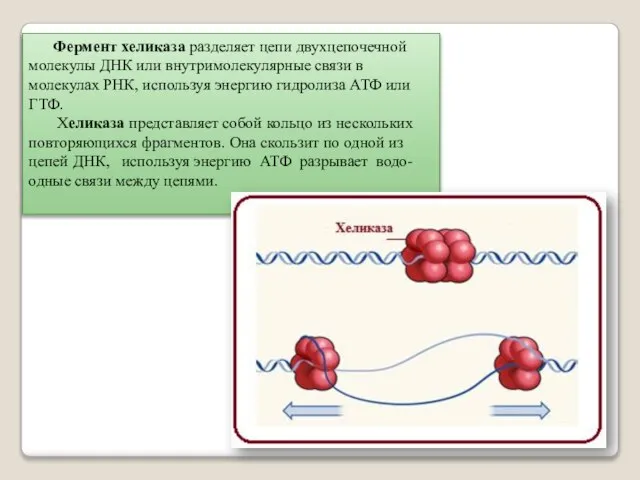
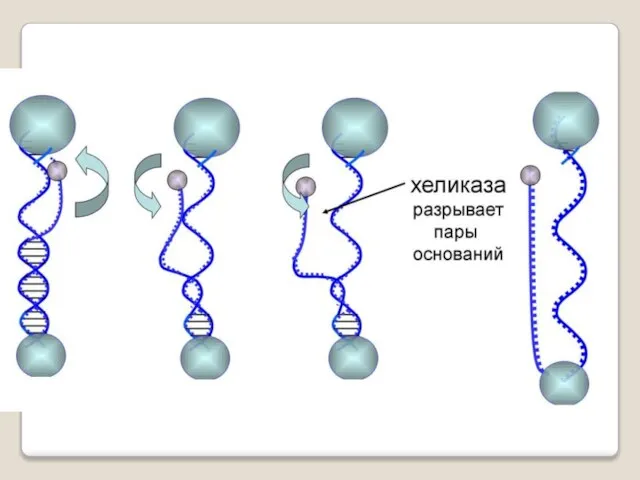
Фермент хеликаза разделяет цепи двухцепочечной молекулы ДНК или внутримолекулярные связи
Фермент хеликаза разделяет цепи двухцепочечной молекулы ДНК или внутримолекулярные связи
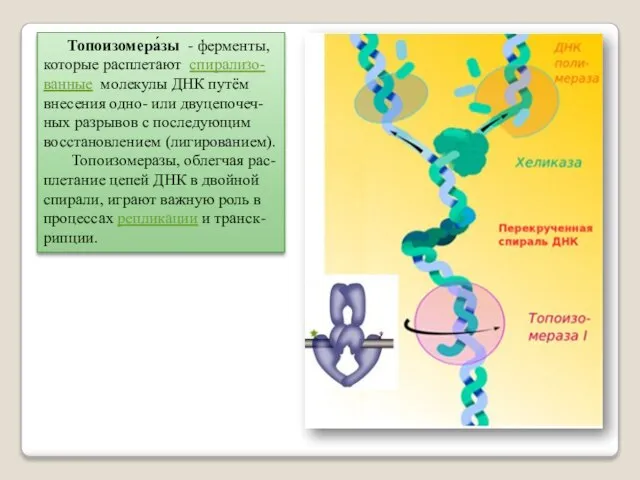
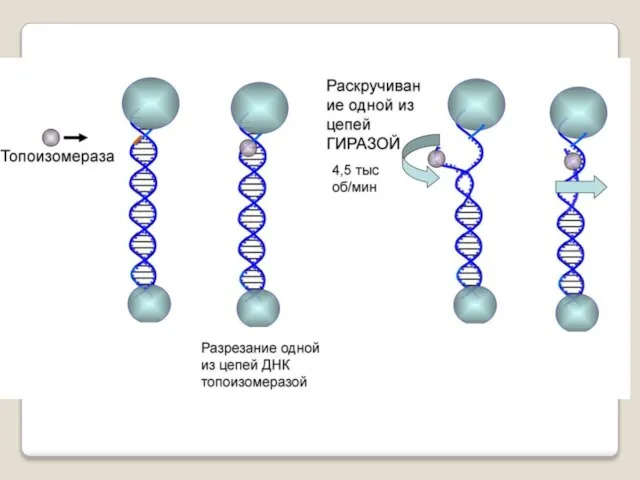
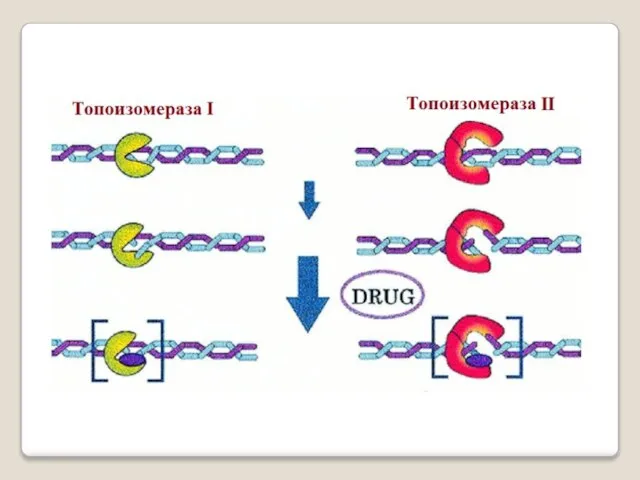
Топоизомера́зы - ферменты, которые расплетают спирализо-
ванные молекулы ДНК путём внесения
Топоизомера́зы - ферменты, которые расплетают спирализо-
ванные молекулы ДНК путём внесения
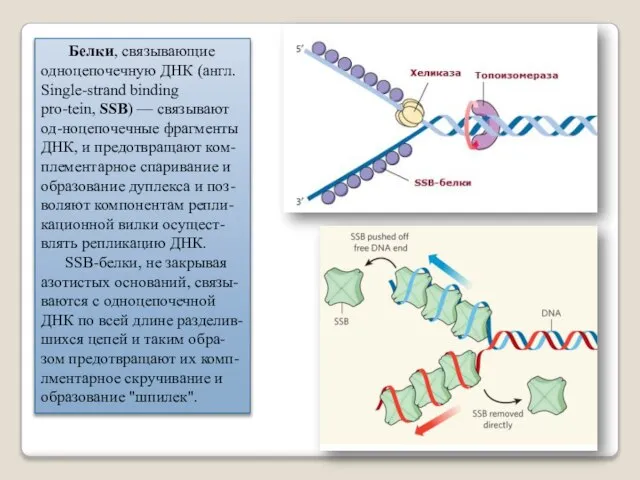
Белки, связывающие одноцепочечную ДНК (англ. Single-strand binding pro-tein, SSB) — связывают
Белки, связывающие одноцепочечную ДНК (англ. Single-strand binding pro-tein, SSB) — связывают
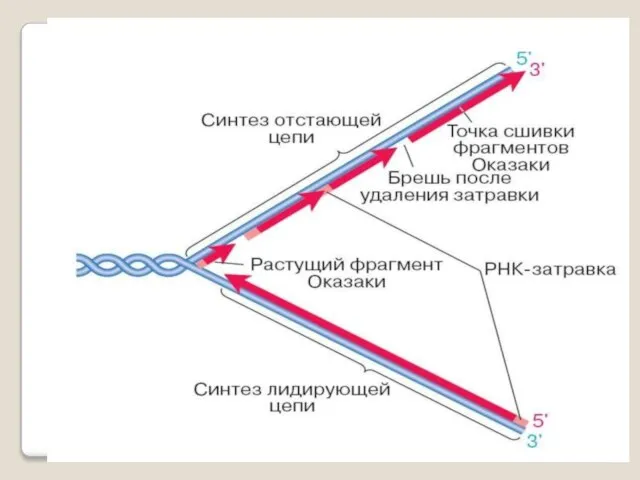

Репликация ДНК осуществляется ДНК-зависимыми ДНК-полимеразами.
Субстратами и источниками энергии для
Репликация ДНК осуществляется ДНК-зависимыми ДНК-полимеразами.
Субстратами и источниками энергии для

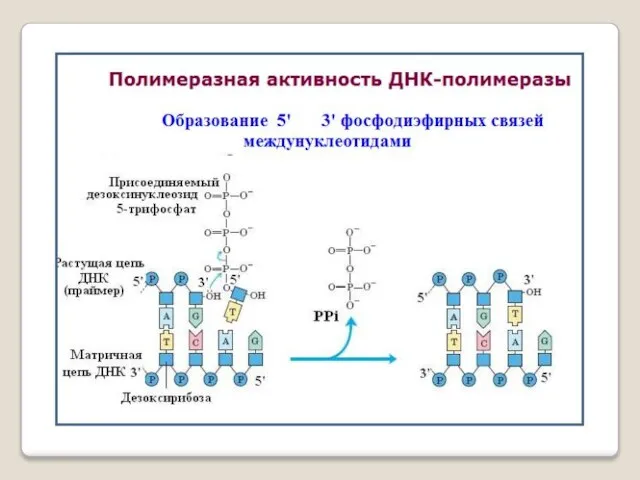
ДНК-полимераза — фермент, участвующий в репликации ДНК. Ферменты этого класса катализ-руют полимеризацию
ДНК-полимераза — фермент, участвующий в репликации ДНК. Ферменты этого класса катализ-руют полимеризацию


У прокариот элонгацию осу- ществляют ДНК-полимеразы I,II,III, обозначаемые как pol
У прокариот элонгацию осу- ществляют ДНК-полимеразы I,II,III, обозначаемые как pol
У эукариот в элонгации участвует сложный белковый комплекс, в состав которого
У эукариот в элонгации участвует сложный белковый комплекс, в состав которого
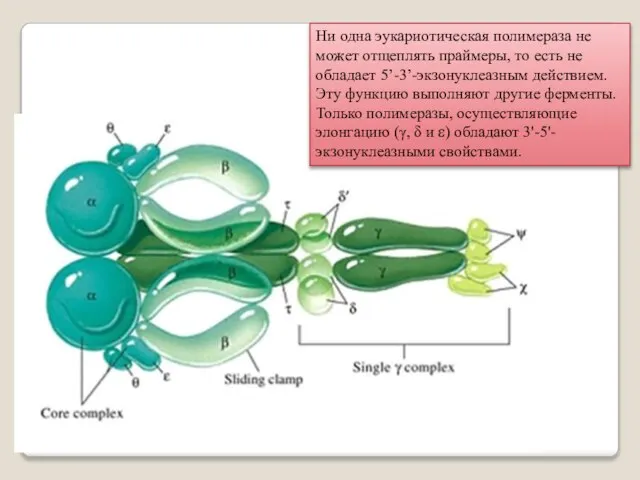
Ни одна эукариотическая полимераза не может отщеплять праймеры, то есть не
Ни одна эукариотическая полимераза не может отщеплять праймеры, то есть не

Прокариотические ДНК-полимеразы
У бактерий обнаружено пять ДНК-полимераз:
ДНК-полимераза I задействована в восстановлении ДНК, обладает
Прокариотические ДНК-полимеразы
У бактерий обнаружено пять ДНК-полимераз:
ДНК-полимераза I задействована в восстановлении ДНК, обладает

Три полимеразы (pol I, pol II и pol III) обладают полимеризующей
Три полимеразы (pol I, pol II и pol III) обладают полимеризующей
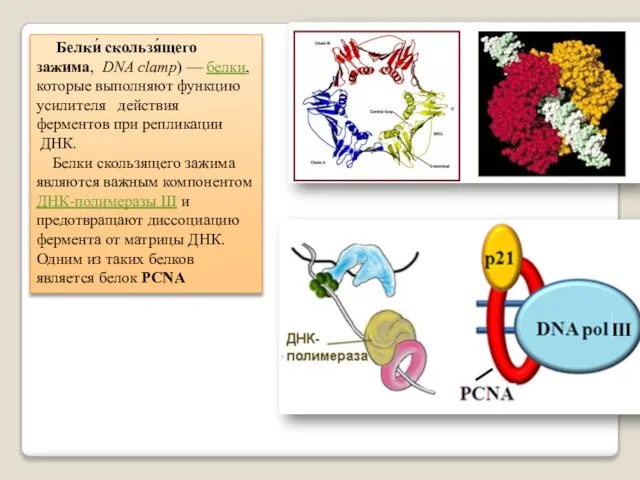
Белки́ скользя́щего зажима, DNA clamp) — белки, которые выполняют функцию усилителя действия
Белки́ скользя́щего зажима, DNA clamp) — белки, которые выполняют функцию усилителя действия

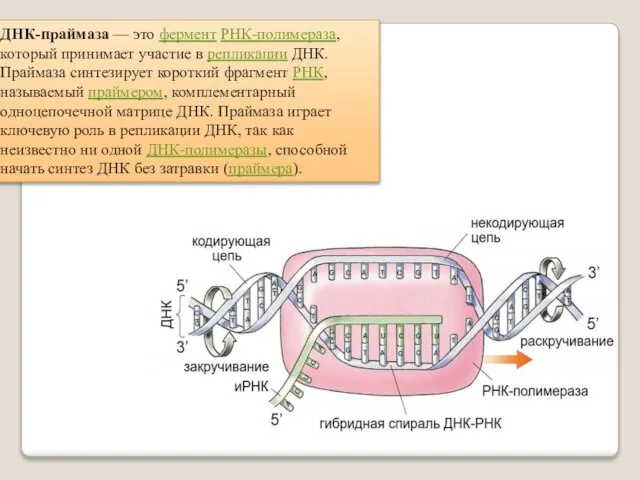
ДНК-праймаза — это фермент РНК-полимераза, который принимает участие в репликации ДНК.
Праймаза синтезирует короткий фрагмент РНК, называемый праймером, комплементарный
ДНК-праймаза — это фермент РНК-полимераза, который принимает участие в репликации ДНК.
Праймаза синтезирует короткий фрагмент РНК, называемый праймером, комплементарный
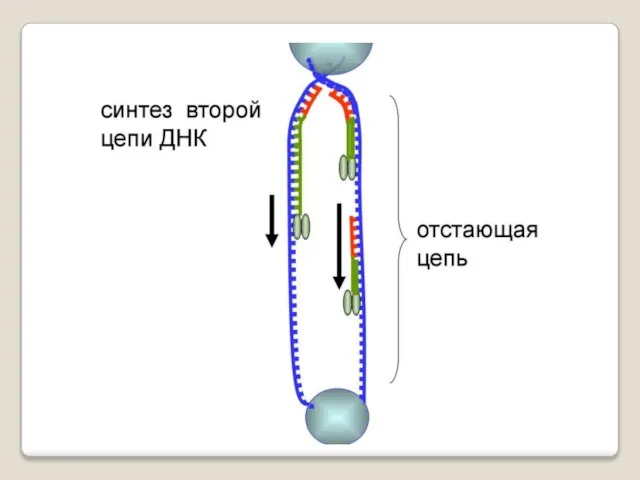
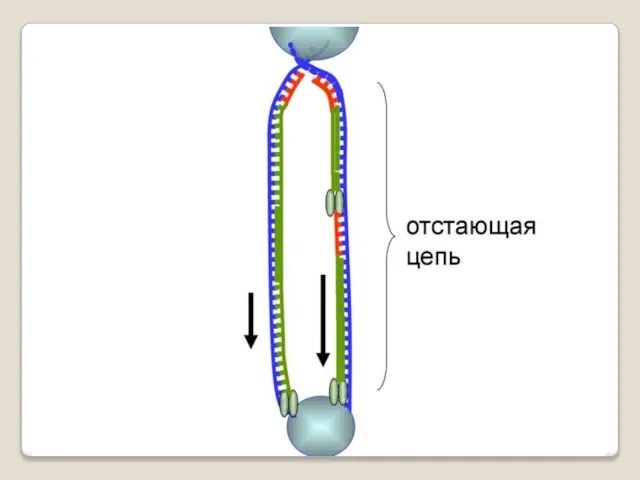
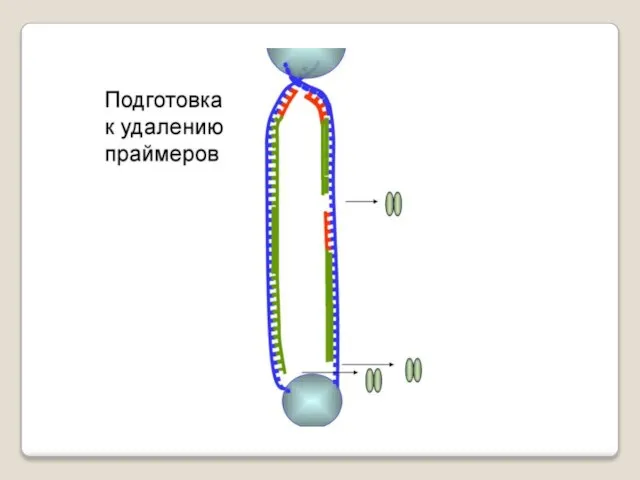
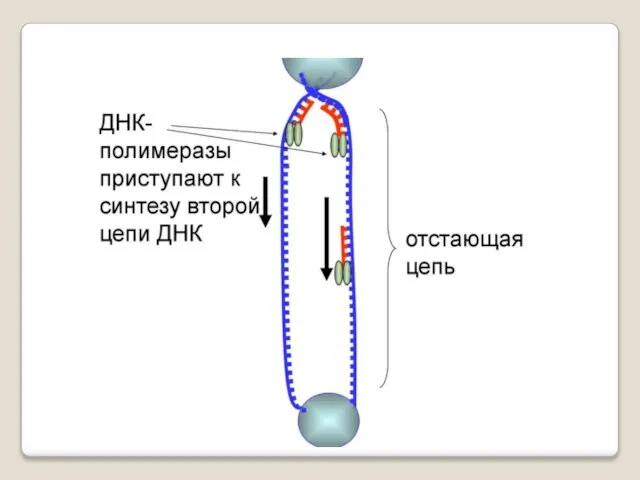

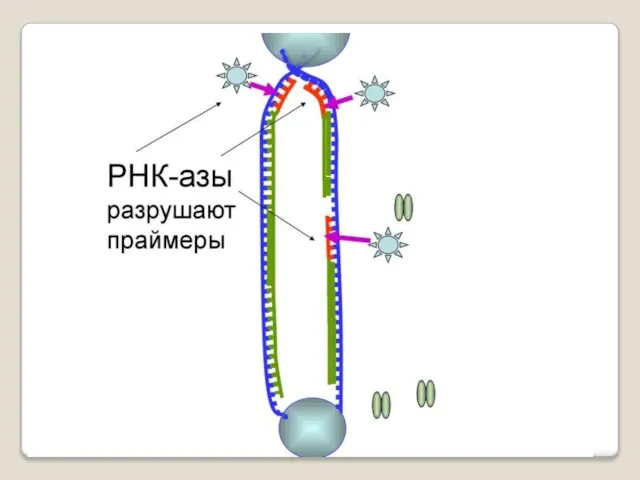
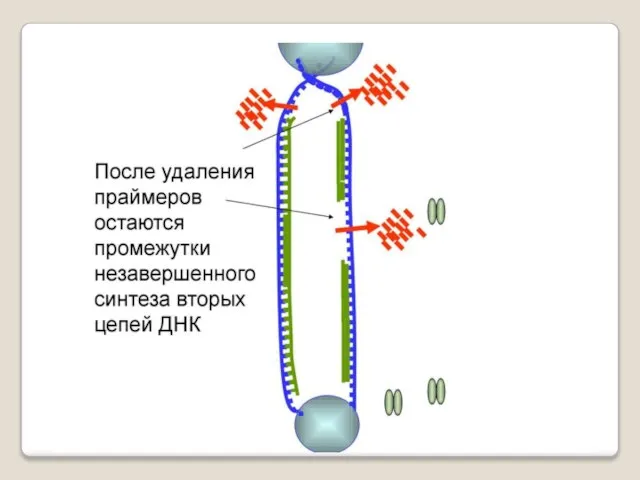

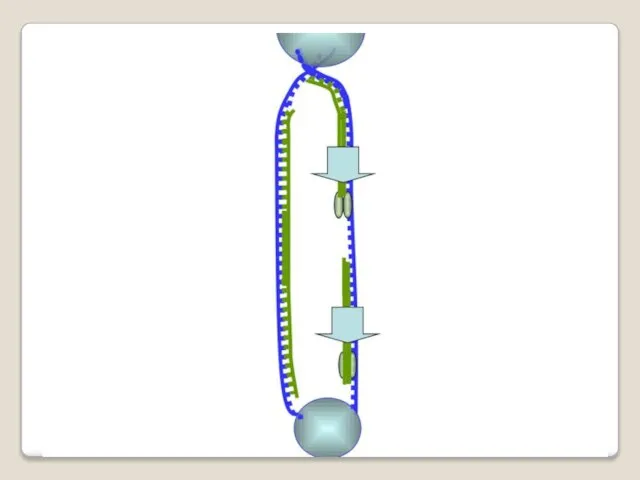
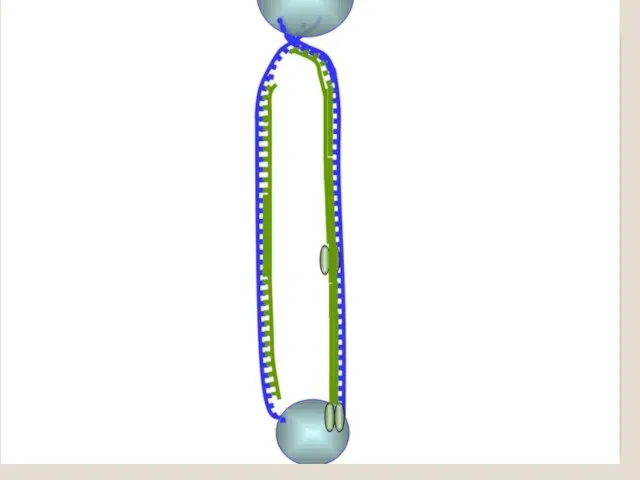



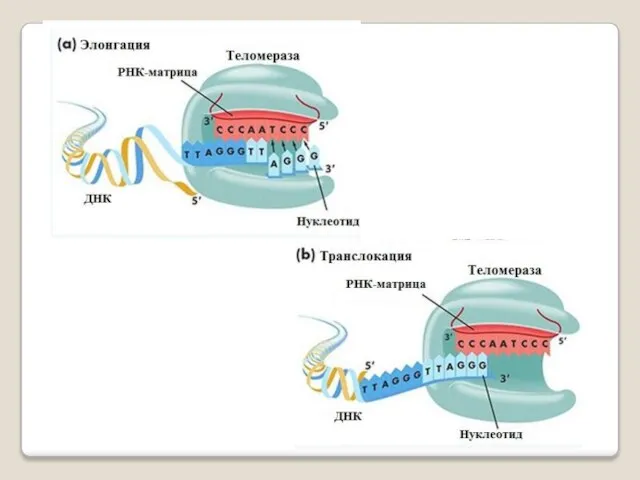
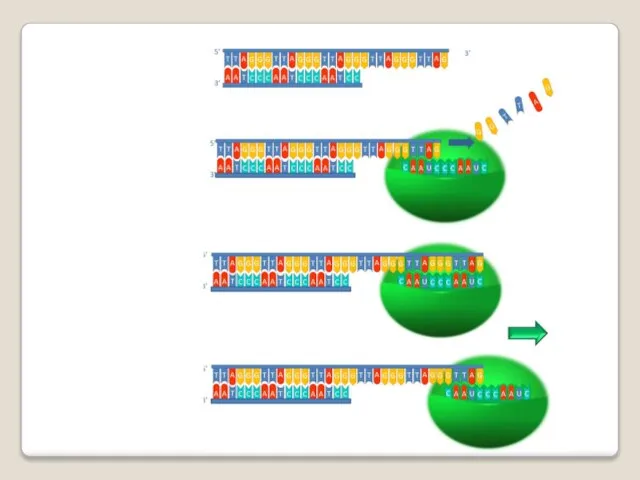
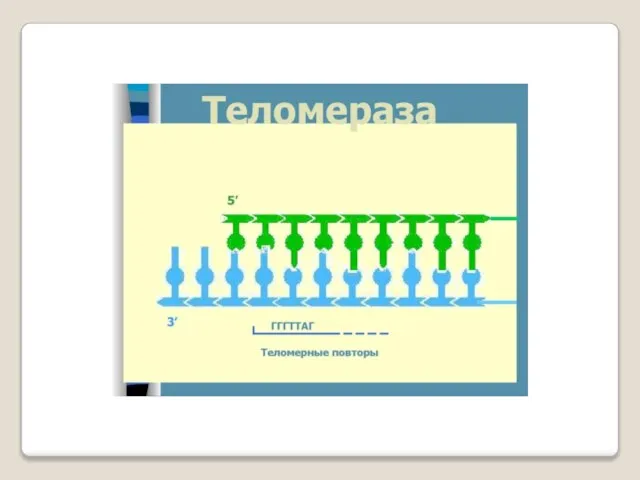
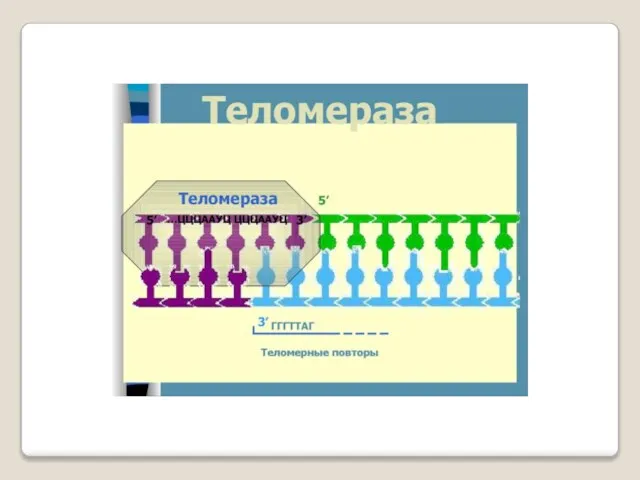
Теломераза — фермент, добавляющий особые повторяющиеся последовательности ДНК (TTAGGG у позвоночных) к 3'-концу цепи ДНК
Теломераза — фермент, добавляющий особые повторяющиеся последовательности ДНК (TTAGGG у позвоночных) к 3'-концу цепи ДНК

А.М. Оловников в 1971 году высказал гипотезу о том, что
А.М. Оловников в 1971 году высказал гипотезу о том, что

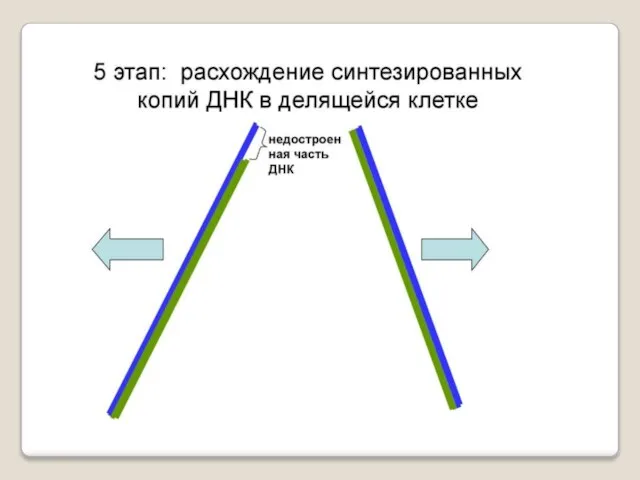
ДНК-полимеразы, синтезируя дочернюю цепь ДНК, прочитывают родительскую цепь в направлении
ДНК-полимеразы, синтезируя дочернюю цепь ДНК, прочитывают родительскую цепь в направлении
 Презентация для класса Паразитические грибы и бактерии
Презентация для класса Паразитические грибы и бактерии  Организация проектной деятельности учащихся на уроках биологии и во внеклассной работе по предмету
Организация проектной деятельности учащихся на уроках биологии и во внеклассной работе по предмету Брожения. Типы жизни, основанные на субстратном фосфорилировании А. Общая характеристика процессов брожения Брожение – это проц
Брожения. Типы жизни, основанные на субстратном фосфорилировании А. Общая характеристика процессов брожения Брожение – это проц Ферменты
Ферменты Презентация на тему "Общая Характеристика водорослей" - скачать презентации по Биологии
Презентация на тему "Общая Характеристика водорослей" - скачать презентации по Биологии Соколоподібні
Соколоподібні Презентация Вода-источник жизни!
Презентация Вода-источник жизни! Гигиена сердечно – сосудистой системы.
Гигиена сердечно – сосудистой системы. Бобр-строитель
Бобр-строитель Функции крови и лимфы
Функции крови и лимфы Субкультуры. История
Субкультуры. История Удивительная Плесень
Удивительная Плесень Органы цветковых растений
Органы цветковых растений Устойчивость растений к внешним факторам среды
Устойчивость растений к внешним факторам среды Многообразие Кольчатых червей Малощетинковые черви Многощетинковые черви Пиявки
Многообразие Кольчатых червей Малощетинковые черви Многощетинковые черви Пиявки  Презентация на тему Семейство Тыквенные – бахчевые культуры. Семейство. Крестоцветные (Капустные)
Презентация на тему Семейство Тыквенные – бахчевые культуры. Семейство. Крестоцветные (Капустные) Презентация на тему "Глаз и зрение" - скачать презентации по Биологии
Презентация на тему "Глаз и зрение" - скачать презентации по Биологии Страусообразные
Страусообразные Зорова сенсорна система. Будова ока
Зорова сенсорна система. Будова ока Внутрішнє середовище організму. Склад крові
Внутрішнє середовище організму. Склад крові Туя западная
Туя западная Дыхательная система
Дыхательная система Двоякодышащие рыбы
Двоякодышащие рыбы Орхідеї
Орхідеї Репликация. Теломеры. Теломераза
Репликация. Теломеры. Теломераза Презентация на тему Лишайники
Презентация на тему Лишайники Презентация по биологии на тему: Палеозой
Презентация по биологии на тему: Палеозой Вирусы
Вирусы