- Метаболизм. Взаимосвязь различных путей обмена веществ

Содержание
- 4. Основные характеристики метаболизма клеток млекопитающих Гетеротрофы Аэробы (факультативные или облигатные). Акцептором электронов являются органические вещества и
- 5. Функции метаболизма Аккумулирование энергии в макроэргических связях АТФ. Использование энергии АТФ для биосинтеза de novo молекул
- 6. Все механизмы регуляции направлены на поддержание гомеостаза и адаптацию к изменяющимся условиям внешней и внутренней среды
- 7. Взаимосвязь обменов: Углеводы – липиды: «Лишние» углеводы обеспечивают синтез липидов (образование глицерол-3- фосфата, ацетил-КоА, НАДФН) глицерин
- 8. Взаимосвязь обменов: Белки – липиды: Запас нейтральных липидов предотвращает использование белков на энергетические нужды. Аминокислоты ?
- 9. Взаимосвязь обменов Белки – углеводы: Гликогенные аминокислоты (кроме лиз и лей) Основной путь синтеза заменимых аминокислот
- 10. Роль нуклеотидов в обмене веществ Адениловые нуклеотиды обеспечивают энергетический потенциал клеток УТФ участвует в обмене углеводов
- 11. Этапы энергетического обмена 1. Крупные «пищевые» молекулы гидролизуются до мономеров: аминокислот, жирных кислот, моносахаров. 2. Специфические
- 12. Унификация клеточного «топлива» В процессе основных путей катаболизма - гликолиза, окислительного дезаминирования, β – окисления, ЦТК
- 13. Основные этапы адаптации организма к голоданию Суточная потребность в энергетических ресурсах в зависимости от физической нагрузки
- 14. Адаптация к голоданию Голодание (равно как любой стресс или диабет) приводит к повышению уровня катехоламинов, глюкагона,
- 15. 1- ая фаза голодания (1сутки) На начальном этапе уровень глюкозы крови (особенно важный для нейронов и
- 16. 2- ая фаза голодания (1 –ая неделя) Гормональная ситуация приводит к адаптивному переключению энергетического обмена с
- 17. 3 – я фаза голодания (1 – 3 месяца) Усиление кетогенеза предотвращает использование аминокислот (Но! метаболический
- 18. Биоэнергетика изучает процессы превращения энергии в живом организме. Биологическое окисление (тканевое дыхание) – процессы, протекающие с
- 19. История учения о биоокислении 18 век, А.Лавуазье: « Дыхание – медленное горение» 1920 - е г.,
- 20. История учения о биоокислении Биологическое окисление – отнятие водорода от субстратов и передача его через серию
- 21. Аккумулирование энергии в клетке: НАДН, ФАДН2, НАДФН АТФ (реже другие нуклеозидтрифосфаты) Внутренняя мембрана митохондрий, функционирующая как
- 22. НАДН, НАДФН, ФАДН2 Восстановленные эквиваленты образуются при окислении органических субстратов; НАДН и ФАДН2 реокисляются в дыхательной
- 23. Роль НАДФН в метаболизме 1. Восстановительные синтезы аминокислот, нуклеотидов, жирных кислот, холестерола, стероидов, желчных кислот. 2.
- 24. Челночные механизмы транспорта Восстановленные эквиваленты, образующиеся в цитозоле, например при окислении глицеральдегид- 3 фосфата попадают в
- 28. Почему именно адениловые нуклеотиды составляют энергетический потенциал клеток? Адениловые нуклеотиды более устойчивы к химическим и физическим
- 29. Содержание адениловых нуклеотидов к клетке АДФ + Ф? [АТФ]?АДФ + Фн (АМФ +ФФ) Синтез АТФ: субстратное
- 36. Гипотезы сопряжения окисления и фосфорилирования Липман, 1946г., Слэйтер, 1953г. Гипотеза химического сопряжения Перенос ē сопряжен с
- 37. Гипотеза химического сопряжения АН2 + В + Х ? А + ВН2 ∞Х ВН2 ∞ Х
- 38. Гипотеза конформационного сопряжения Бойер, 1964: Ленинджер, 1966; Грин, 1970г. Транспорт ē по внутренней мембране МХ вызывает
- 39. Хемиоосмотическая гипотеза Митчелл,1961 Перенос ē вдоль внутренней мембраны сопровождается выкачиванием Н+ из матрикса МХ в наружную
- 40. Доказательства хемиоосмотической гипотезы 1. В МХ нет высокоэнергетических посредников. связывающих перенос ē с биосинтезом АТФ. 2.
- 42. Дыхательная цепь ферментов осуществляет окислительно- восстановительные реакции, перенося ē от одной молекулы к другой и в
- 43. Структура дыхательной цепи МХ I комплекс: НАДН –КоQ дегидрогеназа: кофакторы FMN и Fe-S белки II комплекс:
- 44. Никотинамидные дегидрогеназы Более сотни субстратов окисляются через НАД. Основное количество НАДН образуется при окислении изоцитрата, малата,
- 45. Флавинзависимые дегидрогеназы Коферментную функцию выполняют ФМН или ФАД (производные рибофлавина) Прочно связаны с белковой частью ферментов.
- 46. Убихинон (КоQ) Повсеместно распространенное производное бензохинона. Гидрофобное, низкомолекулярное вещество, не связанное с белком (кофермент), способное мигрировать
- 47. Цитохромы Гемсодержащие белки (15-45 кДа), простетические группы - гем b или a, отличающиеся строением боковых радикалов
- 48. Железосерные белки Низкомолекулярные белки, содержащие негемовое железо и неорганическую серу (2Fe -2S; 4Fe- 4S). Атом железа
- 49. Создание градиента ионов на внутренней мембране МХ Перенос ē по дыхательной цепи сопровождается закислением наружной среды
- 50. Дыхательная цепь ферментов Полная цепь системы МХ окисления: SH2 (изоцитрат, малат, гидроксиацилКоА, гидроксибутират, глутамат) ? никотинамидные
- 51. Транслоказы Кроме оксидоредуктаз дыхательной цепи во внутреннюю мембрану МХ встроена система переноса «клеточного топлива»: кетокислот, аминокислот.
- 57. Строение АТФ – синтазы Н+ - АТФ –синтаза (АТФ-АЗА) V – комплекс дыхательной цепи Сложный белковый
- 59. Протонный градиент и пункты фосфорилирования Атомы водорода от окисляемых субстратов отдают ē в дыхательную цепь и
- 61. Соотношение дыхания и фосфорилирования 4ē + 4Н+ + О2 ? 2Н2О АДФ +Фн ? АТФ Эффективность
- 62. Дыхательный контроль ДК (дыхательный контроль) отражает зависимость интенсивности дыхания от концентрации АДФ. Измеряется полярографически по поглощению
- 63. Энергетический заряд клеток АТФ: АДФ:АМФ Энергетический заряд изменяется от 1 (если все адениловые нуклеотиды максимально фосфорилированы
- 64. Ингибиторы дыхания На различных участках цепи передача ē блокируется специфическими веществами: барбитуратами, некоторыми гликозидами, антибиотиками, цианидами.
- 66. Ингибирование фосфорилирования Олигомицин блокирует Fo – субъединицу АТФ – синтетазы. Электрохимический градиент при этом не снижается,
- 67. Разобщители дыхания и фосфорилирования Липофильные вещества, способные переносить протоны (протонофоры) и катионы (ионофоры) через внутреннюю мембрану,
- 69. Митохондриальная цепь окисления – источник активных форм кислорода 4Н+ + 4ē + О2 ? 2 Н2О
- 71. Скачать презентацию



Основные характеристики метаболизма клеток млекопитающих
Гетеротрофы
Аэробы (факультативные или облигатные). Акцептором электронов являются
Основные характеристики метаболизма клеток млекопитающих
Гетеротрофы
Аэробы (факультативные или облигатные). Акцептором электронов являются

Функции метаболизма
Аккумулирование энергии в макроэргических связях АТФ.
Использование энергии АТФ для биосинтеза
Функции метаболизма
Аккумулирование энергии в макроэргических связях АТФ.
Использование энергии АТФ для биосинтеза

Все механизмы регуляции направлены на поддержание гомеостаза и адаптацию к изменяющимся
Все механизмы регуляции направлены на поддержание гомеостаза и адаптацию к изменяющимся

Взаимосвязь обменов:
Углеводы – липиды:
«Лишние» углеводы обеспечивают синтез липидов (образование глицерол-3-
Взаимосвязь обменов:
Углеводы – липиды:
«Лишние» углеводы обеспечивают синтез липидов (образование глицерол-3-

Взаимосвязь обменов:
Белки – липиды:
Запас нейтральных липидов предотвращает использование белков на энергетические
Взаимосвязь обменов:
Белки – липиды:
Запас нейтральных липидов предотвращает использование белков на энергетические

Взаимосвязь обменов
Белки – углеводы:
Гликогенные аминокислоты (кроме лиз и лей)
Основной путь синтеза
Взаимосвязь обменов
Белки – углеводы:
Гликогенные аминокислоты (кроме лиз и лей)
Основной путь синтеза

Роль нуклеотидов в обмене веществ
Адениловые нуклеотиды обеспечивают энергетический потенциал клеток
УТФ участвует
Роль нуклеотидов в обмене веществ
Адениловые нуклеотиды обеспечивают энергетический потенциал клеток
УТФ участвует

Этапы энергетического обмена
1. Крупные «пищевые» молекулы гидролизуются до мономеров: аминокислот, жирных
Этапы энергетического обмена
1. Крупные «пищевые» молекулы гидролизуются до мономеров: аминокислот, жирных

Унификация клеточного «топлива»
В процессе основных путей катаболизма - гликолиза, окислительного дезаминирования,
Унификация клеточного «топлива»
В процессе основных путей катаболизма - гликолиза, окислительного дезаминирования,

Основные этапы адаптации организма к голоданию
Суточная потребность в энергетических ресурсах в
Основные этапы адаптации организма к голоданию
Суточная потребность в энергетических ресурсах в

Адаптация к голоданию
Голодание (равно как любой стресс или диабет) приводит к
Адаптация к голоданию
Голодание (равно как любой стресс или диабет) приводит к

1- ая фаза голодания (1сутки)
На начальном этапе уровень глюкозы крови (особенно
1- ая фаза голодания (1сутки)
На начальном этапе уровень глюкозы крови (особенно

2- ая фаза голодания (1 –ая неделя)
Гормональная ситуация приводит к адаптивному
2- ая фаза голодания (1 –ая неделя)
Гормональная ситуация приводит к адаптивному

3 – я фаза голодания (1 – 3 месяца)
Усиление кетогенеза предотвращает
3 – я фаза голодания (1 – 3 месяца)
Усиление кетогенеза предотвращает

Биоэнергетика изучает процессы превращения энергии в живом организме.
Биологическое окисление (тканевое дыхание)
Биоэнергетика изучает процессы превращения энергии в живом организме.
Биологическое окисление (тканевое дыхание)

История учения о биоокислении
18 век, А.Лавуазье: « Дыхание – медленное горение»
1920
История учения о биоокислении
18 век, А.Лавуазье: « Дыхание – медленное горение»
1920

История учения о биоокислении
Биологическое окисление – отнятие водорода от субстратов и
История учения о биоокислении
Биологическое окисление – отнятие водорода от субстратов и

Аккумулирование энергии в клетке:
НАДН, ФАДН2, НАДФН
АТФ (реже другие нуклеозидтрифосфаты)
Внутренняя мембрана митохондрий,
Аккумулирование энергии в клетке:
НАДН, ФАДН2, НАДФН
АТФ (реже другие нуклеозидтрифосфаты)
Внутренняя мембрана митохондрий,

НАДН, НАДФН, ФАДН2
Восстановленные эквиваленты образуются при окислении органических субстратов; НАДН и
НАДН, НАДФН, ФАДН2
Восстановленные эквиваленты образуются при окислении органических субстратов; НАДН и

Роль НАДФН в метаболизме
1. Восстановительные синтезы аминокислот, нуклеотидов, жирных кислот, холестерола,
Роль НАДФН в метаболизме
1. Восстановительные синтезы аминокислот, нуклеотидов, жирных кислот, холестерола,

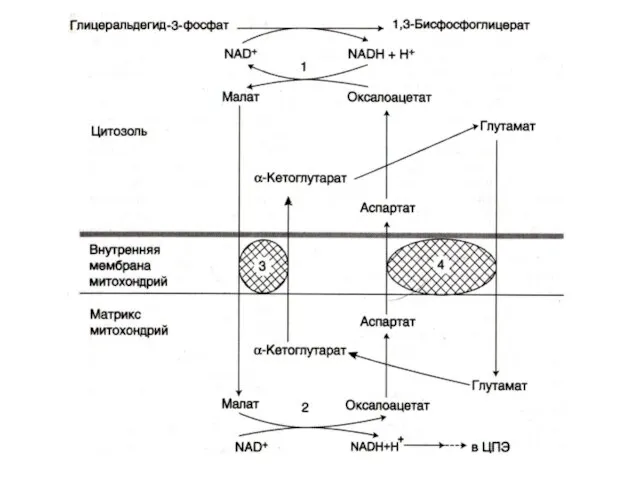
Челночные механизмы транспорта
Восстановленные эквиваленты, образующиеся в цитозоле, например при окислении глицеральдегид-
Челночные механизмы транспорта
Восстановленные эквиваленты, образующиеся в цитозоле, например при окислении глицеральдегид-



Почему именно адениловые нуклеотиды составляют энергетический потенциал клеток?
Адениловые нуклеотиды более устойчивы
Почему именно адениловые нуклеотиды составляют энергетический потенциал клеток?
Адениловые нуклеотиды более устойчивы
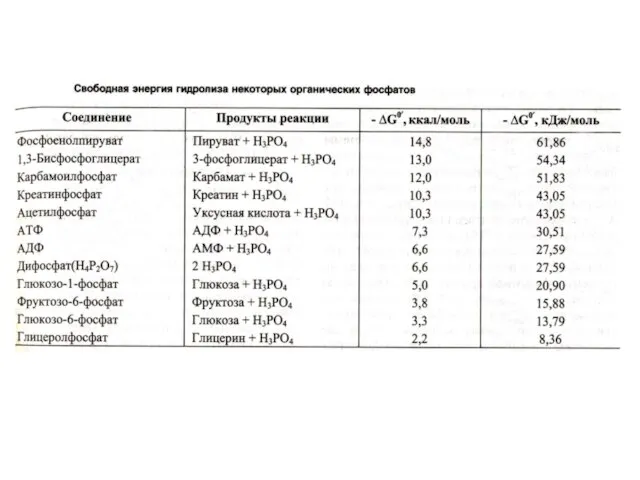
![Содержание адениловых нуклеотидов к клетке АДФ + Ф? [АТФ]?АДФ + Фн](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/512908/slide-28.jpg)
Содержание адениловых нуклеотидов к клетке
АДФ + Ф? [АТФ]?АДФ + Фн (АМФ
Содержание адениловых нуклеотидов к клетке
АДФ + Ф? [АТФ]?АДФ + Фн (АМФ




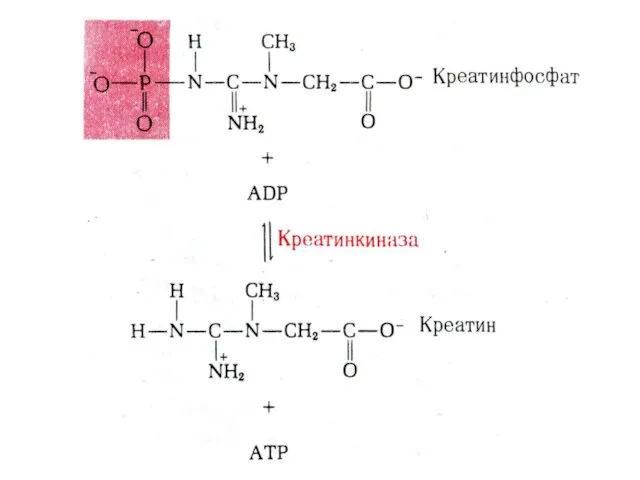

Гипотезы сопряжения окисления и фосфорилирования
Липман, 1946г., Слэйтер, 1953г.
Гипотеза химического сопряжения
Перенос ē
Гипотезы сопряжения окисления и фосфорилирования
Липман, 1946г., Слэйтер, 1953г.
Гипотеза химического сопряжения
Перенос ē

Гипотеза химического сопряжения
АН2 + В + Х ? А + ВН2
Гипотеза химического сопряжения
АН2 + В + Х ? А + ВН2

Гипотеза конформационного сопряжения
Бойер, 1964: Ленинджер, 1966; Грин, 1970г.
Транспорт ē по внутренней
Гипотеза конформационного сопряжения
Бойер, 1964: Ленинджер, 1966; Грин, 1970г.
Транспорт ē по внутренней

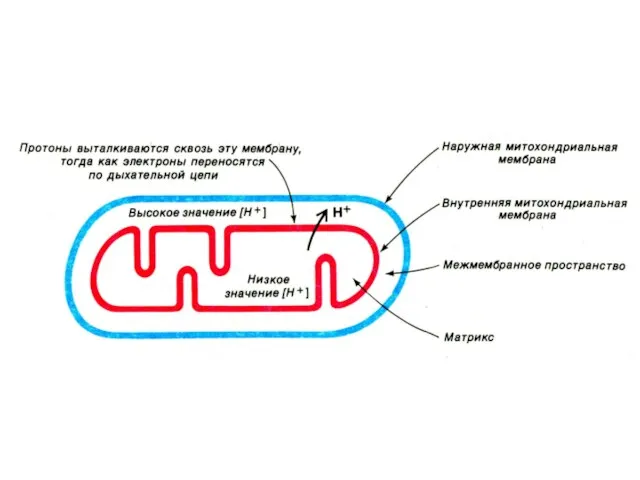
Хемиоосмотическая гипотеза
Митчелл,1961
Перенос ē вдоль внутренней мембраны сопровождается выкачиванием Н+ из матрикса
Хемиоосмотическая гипотеза
Митчелл,1961
Перенос ē вдоль внутренней мембраны сопровождается выкачиванием Н+ из матрикса

Доказательства хемиоосмотической гипотезы
1. В МХ нет высокоэнергетических посредников. связывающих перенос ē
Доказательства хемиоосмотической гипотезы
1. В МХ нет высокоэнергетических посредников. связывающих перенос ē

Дыхательная цепь ферментов осуществляет окислительно- восстановительные реакции, перенося ē от одной
Дыхательная цепь ферментов осуществляет окислительно- восстановительные реакции, перенося ē от одной

Структура дыхательной цепи МХ
I комплекс: НАДН –КоQ дегидрогеназа: кофакторы FMN и
Структура дыхательной цепи МХ
I комплекс: НАДН –КоQ дегидрогеназа: кофакторы FMN и

Никотинамидные дегидрогеназы
Более сотни субстратов окисляются через НАД. Основное количество НАДН образуется
Никотинамидные дегидрогеназы
Более сотни субстратов окисляются через НАД. Основное количество НАДН образуется

Флавинзависимые дегидрогеназы
Коферментную функцию выполняют ФМН или ФАД (производные рибофлавина)
Прочно связаны с
Флавинзависимые дегидрогеназы
Коферментную функцию выполняют ФМН или ФАД (производные рибофлавина)
Прочно связаны с

Убихинон (КоQ)
Повсеместно распространенное производное бензохинона.
Гидрофобное, низкомолекулярное вещество, не
Убихинон (КоQ)
Повсеместно распространенное производное бензохинона.
Гидрофобное, низкомолекулярное вещество, не

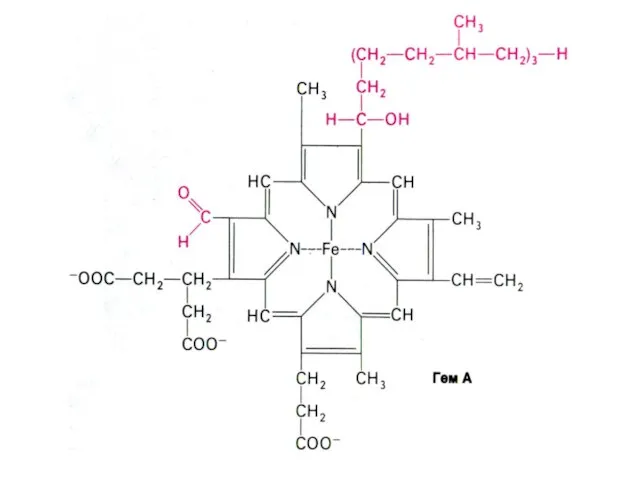
Цитохромы
Гемсодержащие белки (15-45 кДа), простетические группы - гем b или
Цитохромы
Гемсодержащие белки (15-45 кДа), простетические группы - гем b или

Железосерные белки
Низкомолекулярные белки, содержащие негемовое железо и неорганическую серу (2Fe -2S;
Железосерные белки
Низкомолекулярные белки, содержащие негемовое железо и неорганическую серу (2Fe -2S;

Создание градиента ионов на внутренней мембране МХ
Перенос ē по дыхательной
Создание градиента ионов на внутренней мембране МХ
Перенос ē по дыхательной

Дыхательная цепь ферментов
Полная цепь системы МХ окисления:
SH2 (изоцитрат, малат, гидроксиацилКоА, гидроксибутират,
Дыхательная цепь ферментов
Полная цепь системы МХ окисления:
SH2 (изоцитрат, малат, гидроксиацилКоА, гидроксибутират,

Транслоказы
Кроме оксидоредуктаз дыхательной цепи во внутреннюю мембрану МХ встроена система переноса
Транслоказы
Кроме оксидоредуктаз дыхательной цепи во внутреннюю мембрану МХ встроена система переноса





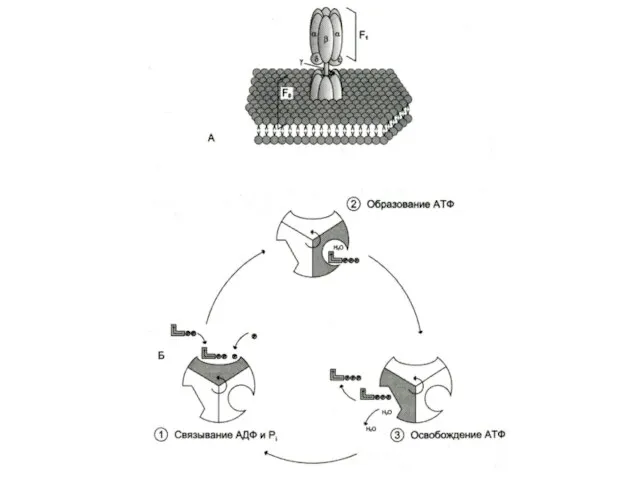
Строение АТФ – синтазы
Н+ - АТФ –синтаза (АТФ-АЗА)
V – комплекс дыхательной
Строение АТФ – синтазы
Н+ - АТФ –синтаза (АТФ-АЗА)
V – комплекс дыхательной

Протонный градиент и пункты фосфорилирования
Атомы водорода от окисляемых субстратов отдают ē
Протонный градиент и пункты фосфорилирования
Атомы водорода от окисляемых субстратов отдают ē


Соотношение дыхания и фосфорилирования
4ē + 4Н+ + О2 ? 2Н2О
АДФ +Фн
Соотношение дыхания и фосфорилирования
4ē + 4Н+ + О2 ? 2Н2О
АДФ +Фн

Дыхательный контроль
ДК (дыхательный контроль) отражает зависимость интенсивности дыхания от концентрации АДФ.
Измеряется
Дыхательный контроль
ДК (дыхательный контроль) отражает зависимость интенсивности дыхания от концентрации АДФ.
Измеряется

Энергетический заряд клеток
АТФ: АДФ:АМФ
Энергетический заряд изменяется от 1 (если все адениловые
Энергетический заряд клеток
АТФ: АДФ:АМФ
Энергетический заряд изменяется от 1 (если все адениловые

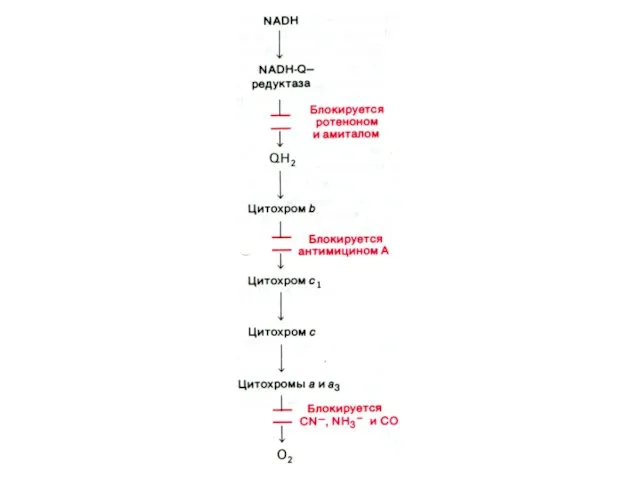
Ингибиторы дыхания
На различных участках цепи передача ē блокируется специфическими веществами:
Ингибиторы дыхания
На различных участках цепи передача ē блокируется специфическими веществами:

Ингибирование фосфорилирования
Олигомицин блокирует Fo – субъединицу АТФ – синтетазы.
Электрохимический градиент
Ингибирование фосфорилирования
Олигомицин блокирует Fo – субъединицу АТФ – синтетазы.
Электрохимический градиент

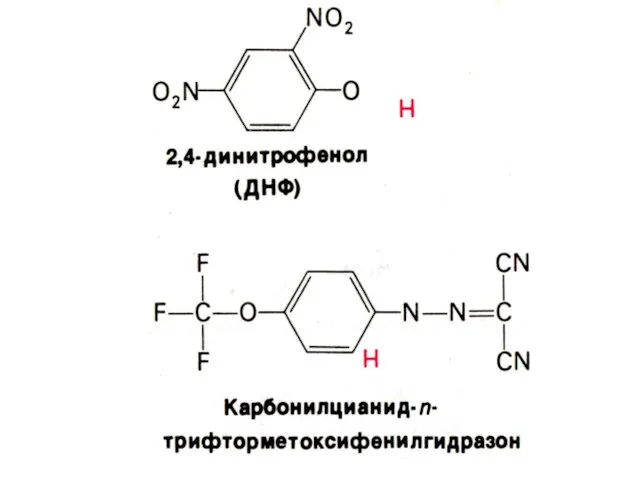
Разобщители дыхания и фосфорилирования
Липофильные вещества, способные переносить протоны (протонофоры) и катионы
Разобщители дыхания и фосфорилирования
Липофильные вещества, способные переносить протоны (протонофоры) и катионы

Митохондриальная цепь окисления – источник активных форм кислорода
4Н+ + 4ē +
Митохондриальная цепь окисления – источник активных форм кислорода
4Н+ + 4ē +
 Пространственная структура глобулярных белков
Пространственная структура глобулярных белков Непрямое постэмбриональное развитие
Непрямое постэмбриональное развитие Аминокислоты и белки Строение и свойства.
Аминокислоты и белки Строение и свойства. Клеточное строение организма
Клеточное строение организма Переходные формы в эволюции Составила учитель биологии Логунцова Ю.В. Школа № 14 и гимназия № 2 г.Кимры, Тверская обл.
Переходные формы в эволюции Составила учитель биологии Логунцова Ю.В. Школа № 14 и гимназия № 2 г.Кимры, Тверская обл.  ТКАНИ И ОРГАНЫ ЧЕЛОВЕКА Урок по биологии 8 класс Учитель Жданова Оксана Викторовна
ТКАНИ И ОРГАНЫ ЧЕЛОВЕКА Урок по биологии 8 класс Учитель Жданова Оксана Викторовна  Ұлулар
Ұлулар Жираф
Жираф Млекопитающие. Летучая мышь
Млекопитающие. Летучая мышь Тепе - теңдік талдағышы. Тактильді талдағышы. Ауырсыну
Тепе - теңдік талдағышы. Тактильді талдағышы. Ауырсыну Презентация на тему "«ПИЩЕВАРЕНИЕ»" - скачать бесплатно презентации по Биологии
Презентация на тему "«ПИЩЕВАРЕНИЕ»" - скачать бесплатно презентации по Биологии «Овсяное «молочко» Мука овсяная для детского и диабетического питания.
«Овсяное «молочко» Мука овсяная для детского и диабетического питания. Презентация на тему Зимующие птицы
Презентация на тему Зимующие птицы  Сущность жизни и свойства живого вещества
Сущность жизни и свойства живого вещества Чувство кворума и альтруизм у бактерий
Чувство кворума и альтруизм у бактерий Тема: Редкие растения
Тема: Редкие растения  Общие представления о жизни
Общие представления о жизни Презентация на тему "Губки 7 класс" - скачать презентации по Биологии
Презентация на тему "Губки 7 класс" - скачать презентации по Биологии Генетическая безопасность агробиоценозов Выполнила студентка 4 курса Факультета естествознания Селянкина Юлия
Генетическая безопасность агробиоценозов Выполнила студентка 4 курса Факультета естествознания Селянкина Юлия  Биоценозы озер
Биоценозы озер АТФ и другие органические соединения клетки
АТФ и другие органические соединения клетки Рептилії. Чому плаче крокодил?
Рептилії. Чому плаче крокодил? Imunitātes veidi
Imunitātes veidi Презентация на тему "Отряд Воробьинообразные" - скачать бесплатно презентации по Биологии
Презентация на тему "Отряд Воробьинообразные" - скачать бесплатно презентации по Биологии Анализаторы. Органы чувств
Анализаторы. Органы чувств ВОЗНИКНОВЕНИЕ АДАПТАЦИЙ И ИХ ОТНОСИТЕЛЬНЫЙ ХАРАКТЕР ВЫПОЛНИЛА УЧИТЕЛЬ БИОЛОГИИ МОУ «МИРНЕНСКАЯ СОШ» УЙСКОГО РАЙОНА ЧЕЛЯБИНСКОЙ
ВОЗНИКНОВЕНИЕ АДАПТАЦИЙ И ИХ ОТНОСИТЕЛЬНЫЙ ХАРАКТЕР ВЫПОЛНИЛА УЧИТЕЛЬ БИОЛОГИИ МОУ «МИРНЕНСКАЯ СОШ» УЙСКОГО РАЙОНА ЧЕЛЯБИНСКОЙ Птички-синички
Птички-синички Презентация на тему
Презентация на тему