- Молекулярно-генетические механизмы инициации цветения

Содержание
- 2. Гены, контролирующие время цветения Гены идентичности флоральной меристемы Гены идентичности органов цветка вегетативная меристема побега органы
- 3. Генетическая регуляция цветения у A.thaliana Гены, контролитрующие инициацию цветения Blázquez, 2001
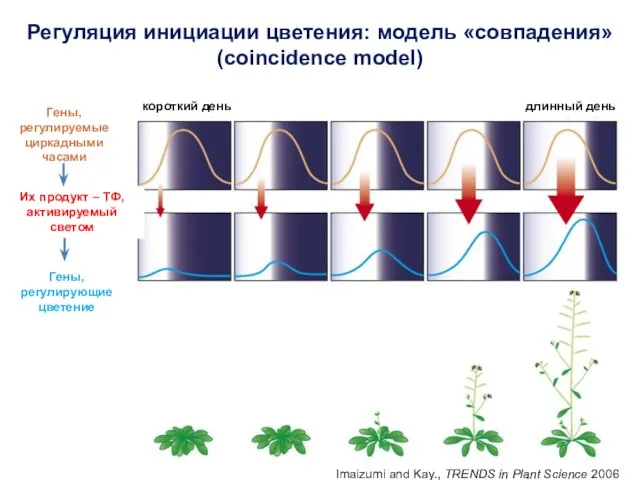
- 4. Инициация цветения Гормоны, сахара Температура Длина светового дня Циркадные ритмы Влияние различных факторов на инициацию цветения
- 5. Открытие фотопериодизма у растений:
- 6. Флориген - гормон цветения, термин предложен М. Х. Чайлахяном, 1936 Гипотеза: флориген синтезируется в листьях и
- 7. Инициация цветения Гормоны, сахара Температура Длина светового дня Циркадные ритмы Влияние различных факторов на инициацию цветения
- 8. 1. Растения короткого дня (кукуруза, рис и др.) 2. Растения длинного дня (арабидопсис, пшеница, овес, рожь
- 9. свет красный свет синий свет фитохромы (PHYB, PHYD, PHYE) криптохромы (CRY1, CRY2) «циркадные часы» Гены, регулирующие
- 10. Основные компоненты «циркадных часов» растений LHY – LATE ELONGATED HYPOCOTYL CCA1 – CIRCADIAN CLOCK ASSOCIATED TOC1
- 11. Активность люциферазы у растений CAB::LUC (CAB - chlorophyll a/b-binding protein) «Циркадные ритмы» растений
- 12. Imaizumi and Kay., TRENDS in Plant Science 2006 Гены, регулируемые циркадными часами Гены, регулирующие цветение Их
- 13. WT 35S::CO Ген CONSTANS (CO) в регуляции инициации цветения Ген CO – кодирует предполагаемый ТФ с
- 14. Экспрессия гена CONSTANS (CO::GUS) в проводящих тканях An et al., Development 2004
- 15. Короткий день день ночь Длинный день Время Цветение день ночь Динамика экспрессии гена CONSTANS в течение
- 16. Фотопериодическая регуляция активности CO
- 17. Фотопериодическая регуляция активности CO
- 18. COP1 синий свет криптохромы (CRY1, CRY2) CO protein цветение Фотопериодическая регуляция активности CO CO mRNA Криптохромоподобный
- 19. COP1 опосредует убиквитин-зависимую деградацию белка CO Liu et al., Plant Cell 2008
- 20. COP1 опосредует убиквитин-зависимую деградацию белка CO: исследование взаимодействия белков COP1 и CO Двугибридная дрожжевая система Анализ
- 21. СОР1 опосредует темновую деградацию белка СО
- 22. Молекулярные механизмы «гипотезы совпадения» Регуляция на уровне мРНК СО: FKF1 (FLAVIN-BINDING KELCH REPEAT FBOX1) – Fbox-содержащий
- 23. Выявление мишеней CONSTANS FT - белок 20 кДа, имеет гомологию с Raf-киназным ингибитором или фосфатидилэтаноламин связывающим
- 24. КОРОТКИЙ ДЕНЬ ДЛИННЫЙ ДЕНЬ мРНК CO мРНК CO белок CO белок CO мРНК FT мРНК FT
- 25. Imaizumi and Kay., TRENDS in Plant Science 2006 Продукт гена FT (мРНК, белок-?) – транспортируется из
- 26. Активация экспрессии FT (HSP::FT) цветение ТШ ТШ Содержание транскриптов FT и GUS (контроль) в апексе побега
- 27. http://4e.plantphys.net/article.php?ch=25&id=398 Jaeger and Wigge Current Biology 2007 Передвижение белка Myc FT в апекс побега Белок FT
- 28. Белок FT (флориген) транспортируется из листа в апекс побега, где инициирует цветение
- 29. http://4e.plantphys.net/article.php?ch=25&id=398 Фотопериодическая регуляции цветения у растений длинного дня и растений короткого дня
- 30. Фотопериодическая регуляция активности CO и FT
- 31. Основные гены, контролирующие фотопериодическую индукцию цветения у длиннодневного растения Arabidopsis thaliana (А) и короткодневного растения Oryza
- 32. Ген TFL (Terminal Flower) арабидопсиса Продукт гена TFL – белок - репрессор цветения, на 60% сходный
- 33. TFL1 – имеет 59% сходства с белком FT FD FD FD FD активация транскрипции репрессия транскрипции
- 34. Белки семейства FT/TFL у разных видов семенных растений
- 35. Дополнительная функция FT – контроль образования клубней картофеля
- 36. Выявление мишеней FT: Abe et al., SCIENCE 2005 Изучение влияния мутаций по различным генам, контролирующих инициацию
- 37. Экспрессия FD (FD::GUS) в апексе побега Abe et al., SCIENCE 2005 Ген FD кодирует транскрипционный фактор
- 38. Передача сигнала флоригена FT: FAC (Florigen Activating Complex) Участники: Белки семейства 14-3-3 Транскрипционный фактор FD Taoka
- 39. Выявление мишеней FT FT SOC1 (SUPPRESSOR OF OVEREXPRESSION OF CONSTANS1)
- 40. Функции ТФ SOC1 SOC1 + AGL24, 71, 72 LFY FT + FD CO FLC гиббереллины Органы
- 41. Мишени ТФ SOC1
- 42. C промотором SOC1 связываются димеры «цветочных» ТФ MADS, подавляя его экспрессию SOC1 SEP3, SEP 1, 2,
- 43. Схема регуляции цветения у растений с участием ТФ SOC1 http://4e.plantphys.net/article.php?ch=25&id=398
- 44. Инициация цветения Гормоны, сахара Температура Длина светового дня Циркадные ритмы Влияние различных факторов на инициацию цветения
- 45. Achard et al., PNAS 2007 Антагонизм этилена и гиббереллинов в регуляции цветения гиббереллины цветение этилен
- 46. Роль гиббереллинов в индукции цветения GA LFY SOC FT цветение
- 47. Инициация цветения Гормоны, сахара Температура Длина светового дня Циркадные ритмы Влияние различных факторов на инициацию цветения
- 48. Вернализация – продолжительное воздействие низких температур (как правило, 1-7 °С в течение 1-3 месяцев), необходимое для
- 49. Молекулярные основы эпигенетических изменений: метилирование ДНК 2. модификация структуры хроматина: изменение уровня метилирования и ацетилирования гистонов
- 50. VRN1 - ДНК-связывающий Myb-белок VRN2 - белок Polycomb-группы, обладающий гистон-метил-трансферазной активностью VIN3 – компонент гистон-деацетилазного комплекса
- 51. Sung, Amasino. Curr Opin Plant Biol 2004 Эпигенетическая регуляция активности гена FLC с участием белков VRN
- 52. Регуляция уровня транскриптов FLC: метилирорвание /де-метилирование гистонов, антисенс-РНК FCA+FY – факторы процессинга РНК, регулируют стабильность COOLAIR
- 53. Drosophila melanigaster Arabidopsis thaliana POLYCOMB - комплекс обладает гистон – метилтрансферазной активностью (метилирует Lys9 и Lys27
- 54. (а) – только PRC2 комплекс ? неполное подавление экспрессии (b) – присоединение белков-кофакторов ? полное подавление
- 55. Регуляция экспрессии FLC с помощью эпигенетических механизмов Jarillo et al., 2009
- 56. Активация экспрессии FLC (в эмбриональном развитии) Jarillo et al., 2009 Гистон H2A.Z ARP6 TRITHORAX 1 (ATX-1)
- 57. Участие некодирующих РНК в подавлении экспрессии FLC COOLAIR (COLD INDUCED LONG ANTISENSE RNA) – антисенс FLC
- 58. Рекрутирует деметилазу H3K4me2/3 Участие некодирующих РНК в подавлении экспрессии FLC Grimanelli and Roudier, 2013 COOLAIR COLDAIR
- 59. COLDAIR рекрутирует компоненты PRC2 (Polycomb repression complex 2) к локусу FLC, опосредуя его стабильную репрессию COLDAIR–
- 60. Регуляция экспрессии с участием комплексов Thrithorax, Polycomb и длинных некодирующих РНК Arabidopsis млекопитающие
- 62. - LEAFY (LFY ) - APETALA1 (AP1) - CAULIFLOWER (CAL ) - UNUSUAL FLORAL ORGANS (UFO)
- 63. Мутации, нарушающие функцию генов, отвечающих за образование цветочной меристемы, приводят к замене цветков на побеги (гомеозисные
- 64. дикий тип lfy Фенотип мутации lfy у арабидопсиса lfy
- 65. Слабый уровень экспрессии гена LFY наблюдается и на вегетативной стадии развития (в АМ и листовых примордиях)
- 66. Фенотип трансгенных растений 35S::LFY служит дополнительным подтверждением роли LFY в формировании ФМ У растений 35S::LFY ген
- 67. Ортологи гена LFY FLORICAULA (FLO) – Antirrhinum majus NFL – Nicotiana tabacum ALF – Petunia hybrida
- 68. Характер экспрессии генов-ортологов LFY/FLO определяет тип соцветия (открытое или закрытое) открытое закрытое LFY
- 69. Ген сосны NEEDLY под промотором гена LFY супрессирует мутацию lfy-26
- 70. Наличие ортологов генов LFY/FLO у голосеменных и покрытосеменных растений позволяет предполагать, что эти гены участвовали в
- 71. Ген TFL1 – антогонист гена LFY. Его экспрессия препятствует экспрессии LFY в АМ. При мутации tfl1
- 72. Связь между структурой соцветия (открытое/закрытое) и особенностями экспрессии генов антогонистов – ортологов LFY и TFL1 в
- 73. Особенности морфологии мутантов ap1 и cal Arabidopsis thaliana APETALA1 (AP1) и CAULIFLOWER (CAL) - гены идентичности
- 74. У мутанта ap1cal меристемы соцветия формируют новые меристемы соцветия, но не цветочные меристемы Соцветие двойного мутанта
- 75. Гомеозисные мутанты арабидопсиса Генетический контроль дифференцировки органов цветка
- 76. ABC-модель развития органов цветка (Coen, Meyerowitz, 1991) Перекрывание доменов экспрессии гомеозисных генов в примордии цветка
- 77. Объяснение фенотипов мутантов с нарушением развития органов цветка с помощью АВС-модели
- 78. 35S::AP3 35S::PI; ag 35S::AP3 35S::PI 35S::AP3 35S::PI 35S::AG AP3/PI AP3/PI AP3/PI AG Подтверждение АВС-модели путем исследования
- 79. Структура белков, содержащих MADS-домены MCM1 – регулятор транскрипции дрожжей AG – гомеозисный ген резушки (класс С)
- 80. Meyerowitz, Yanofsky 1990-1995 Тип I MADS-гены растений Все гены АВС-классов (кроме гена АР2) имеют структуру MADS-бокс
- 81. AP2 1,2,3 - стадии развития цветка ранняя Экспрессия генов, контролирующих развитие органов цветка мРНК AP2 белок
- 82. A. wt B. ap2-9 C. 35S::MIR172a-1 Фенотип трансгенных растений 35S::MIR172a-1 повторяет фенотипическое проявление мутации ap2-9 miR172
- 83. miR172 регулирует экспрессию гена AP2
- 84. (a) Область локализации мРНК AP2 (на стадии 7 развития цветка (b) Область локализации miR172 (на стадии
- 85. sep1 sep2 sep3 Pelaz et al., Nature 2000 wt Роль генов SEPALLATA в развитии органов цветка
- 86. Дополненная модель развития цветка SHATTERPROOF 1, 2, SEEDSTICK SEPALLATA 1,2,3,4
- 87. Дополненная модель развития цветка Lohmann et al., Developmental Cell 2002 «Модель квартета»: продукты генов идентичности органов
- 88. Доказано существование прямой регуляции (связывание с ДНК) Регуляция генов, контролирующих развитие органов цветка
- 89. Lenhard et al., Cell 2001 CLV3 WUS Поддержание запаса стволовых клеток в АМ побега LFY AG
- 90. AG – подавляет экспрессию гена WUS в меристеме цветка сохранение экспрессии WUS в меристеме цветка у
- 91. Кадастровые гены определяют пространственные границы «кругов»: SUPERMAN (SUP) RABBIT EARS (RBE) 2 3 4 Ч Л
- 92. Cubas et al., 2009 Образец пелорического цветка из гербария К. Линнея (1744 г.) Наследуемые «эпимутации»: варианты
- 93. Luo et al., 2005 Мутации в генах CYCLOIDEA и DICHOTOMA у Antirrhínum приводят к появлению пелорических
- 94. Роль ТФ CYCLOIDEA и DICHOTOMA в формировании симметрии цветка Мишени CYC/DICH - RADIALIS и DIVARICATA –
- 95. Анализ метилирования в локусе Lvcyc у пелорических «мутантов»льнянки с помощью чувствительных к метилированию рестриктаз Sau3A и
- 96. Гиперметилирование локуса SUPERMAN у «эпимутантов» clark kent (clk) дикий тип superman clark kent Jacobsen and Meyerowitz,
- 97. Theissen and Melzer, 2007 Эволюция регуляции развития цветка
- 98. Модели, объясняющие формирование в ходе эволюции обоеполого цветка «mostly-male» theory «out-of-male» theory «out-of-female» theory Зависимость экспрессии
- 99. Theissen and Melzer, 2007 Модель, объясняющая распределение областей экспрессии «мужских» и (класс C+B) и«женских» (класс С)
- 100. Chanderbali et al 2010 Эволюция регуляции развития цветка
- 101. Цветки с двойным венчиком (лилейные) Цветки с двойной чашечкой Мутации в АВС- генах приводят к формированию
- 102. Модифицированная модель Формирование цветков однодольных (Лилейные)
- 103. TGDEFA, TGDEFB GLO-подобные гены DEF-подобные гены Tulipa gesneriana Гены класса В TGGLO Lilium regale LRDEF LRGLOA,
- 104. Tulipa gesneriana Экспрессия генов класса В Tricyrtis affinis Asparagales: Phalaenopsis equestris, Dendrobium crumenatum, Agapanthus praecox
- 105. Мутация viridiflora Tulipa gesneriana дикий тип Double-flowered
- 106. Нерешённые вопросы модифицированной АВС модели Muscari armeniacum Asparagus officinalis Tulipa gesneriana
- 108. Скачать презентацию
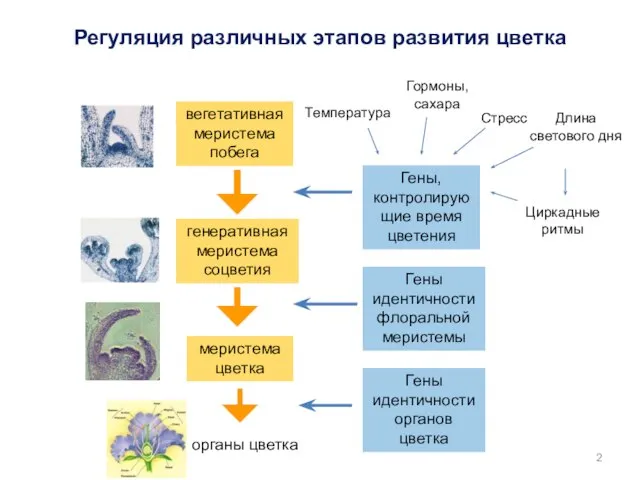
Гены, контролирующие время цветения
Гены идентичности флоральной меристемы
Гены идентичности органов цветка
вегетативная
меристема
Гены, контролирующие время цветения
Гены идентичности флоральной меристемы
Гены идентичности органов цветка
вегетативная
меристема
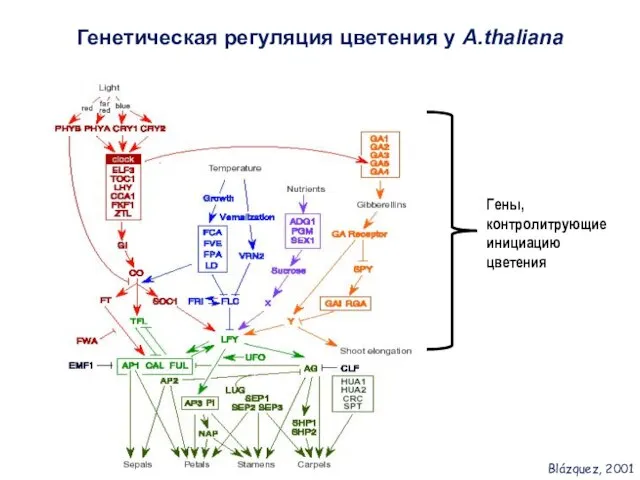
Генетическая регуляция цветения у A.thaliana
Гены, контролитрующие инициацию цветения
Blázquez, 2001
Генетическая регуляция цветения у A.thaliana
Гены, контролитрующие инициацию цветения
Blázquez, 2001
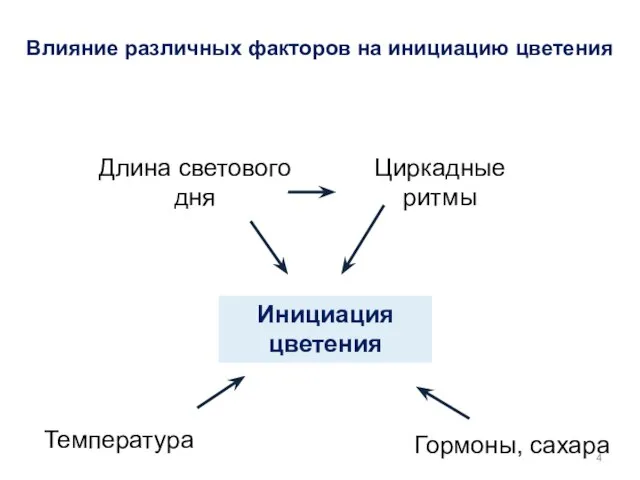
Инициация цветения
Гормоны, сахара
Температура
Длина светового дня
Циркадные ритмы
Влияние различных факторов на инициацию цветения
Инициация цветения
Гормоны, сахара
Температура
Длина светового дня
Циркадные ритмы
Влияние различных факторов на инициацию цветения

Открытие фотопериодизма у растений:
Открытие фотопериодизма у растений:
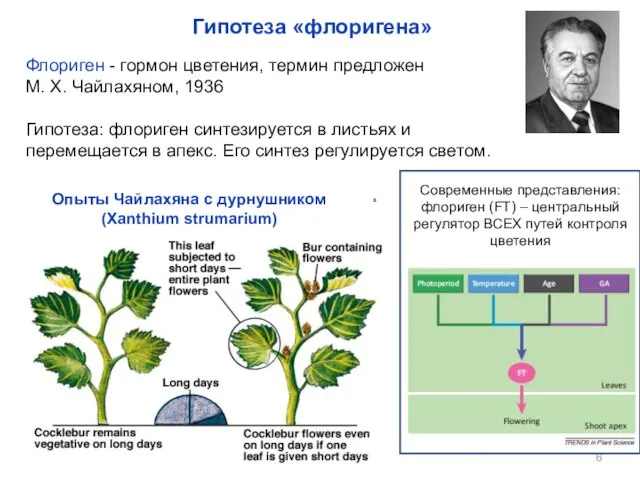
Флориген - гормон цветения, термин предложен
М. Х. Чайлахяном, 1936
Гипотеза:
Флориген - гормон цветения, термин предложен
М. Х. Чайлахяном, 1936
Гипотеза:
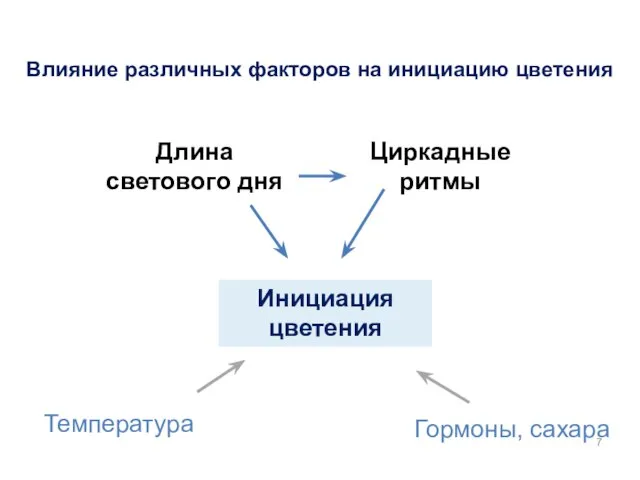
Инициация цветения
Гормоны, сахара
Температура
Длина светового дня
Циркадные ритмы
Влияние различных факторов на инициацию цветения
Инициация цветения
Гормоны, сахара
Температура
Длина светового дня
Циркадные ритмы
Влияние различных факторов на инициацию цветения
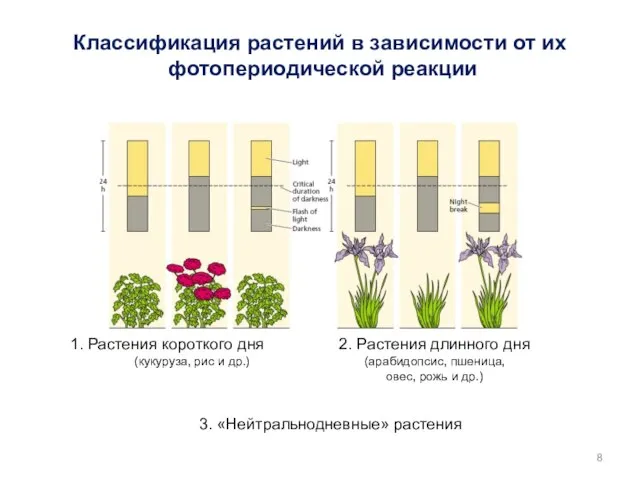
1. Растения короткого дня
(кукуруза, рис и др.)
2. Растения длинного дня (арабидопсис,
1. Растения короткого дня
(кукуруза, рис и др.)
2. Растения длинного дня (арабидопсис,
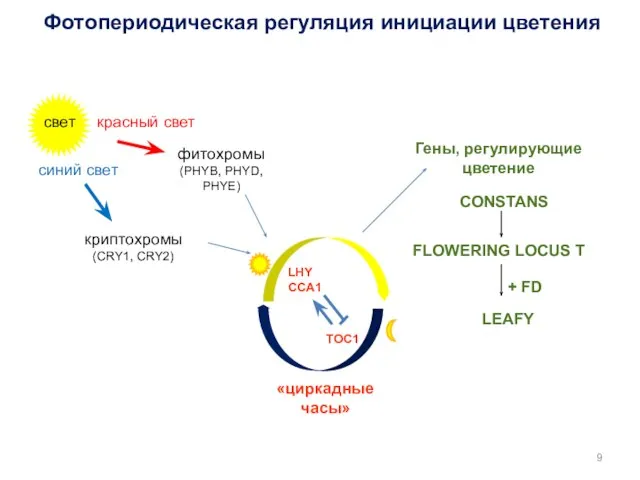
свет
красный свет
синий свет
фитохромы
(PHYB, PHYD, PHYE)
криптохромы
(CRY1, CRY2)
«циркадные часы»
Гены, регулирующие цветение
Фотопериодическая регуляция инициации
свет
красный свет
синий свет
фитохромы
(PHYB, PHYD, PHYE)
криптохромы
(CRY1, CRY2)
«циркадные часы»
Гены, регулирующие цветение
Фотопериодическая регуляция инициации
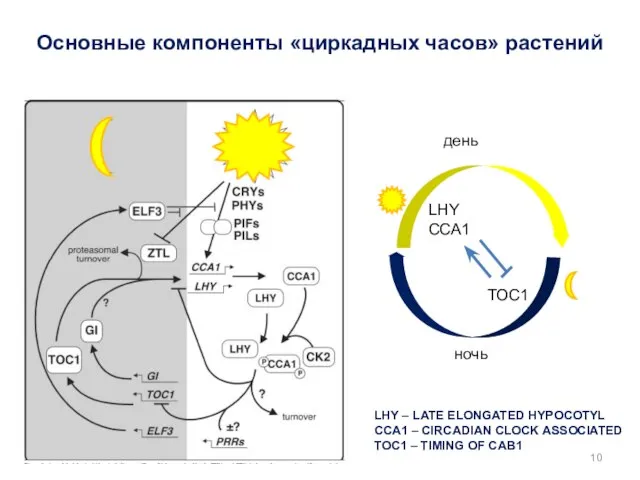
Основные компоненты «циркадных часов» растений
LHY – LATE ELONGATED HYPOCOTYL
CCA1 – CIRCADIAN
Основные компоненты «циркадных часов» растений
LHY – LATE ELONGATED HYPOCOTYL
CCA1 – CIRCADIAN
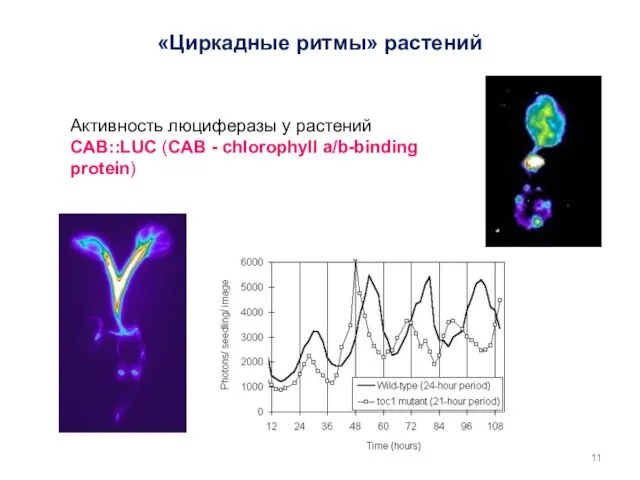
Активность люциферазы у растений CAB::LUC (CAB - chlorophyll a/b-binding protein)
«Циркадные
Активность люциферазы у растений CAB::LUC (CAB - chlorophyll a/b-binding protein)
«Циркадные
Imaizumi and Kay., TRENDS in Plant Science 2006
Гены, регулируемые циркадными
Imaizumi and Kay., TRENDS in Plant Science 2006
Гены, регулируемые циркадными

WT
35S::CO
Ген CONSTANS (CO) в регуляции инициации цветения
Ген CO – кодирует предполагаемый
WT
35S::CO
Ген CONSTANS (CO) в регуляции инициации цветения
Ген CO – кодирует предполагаемый
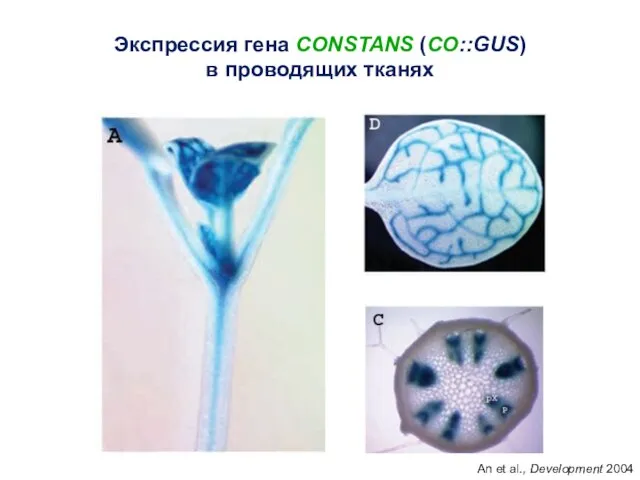
Экспрессия гена CONSTANS (CO::GUS)
в проводящих тканях
An et al., Development 2004
Экспрессия гена CONSTANS (CO::GUS)
в проводящих тканях
An et al., Development 2004

Короткий день
день
ночь
Длинный день
Время
Цветение
день
ночь
Динамика экспрессии гена CONSTANS в течение суток
Нестабильность белка CO
Короткий день
день
ночь
Длинный день
Время
Цветение
день
ночь
Динамика экспрессии гена CONSTANS в течение суток
Нестабильность белка CO
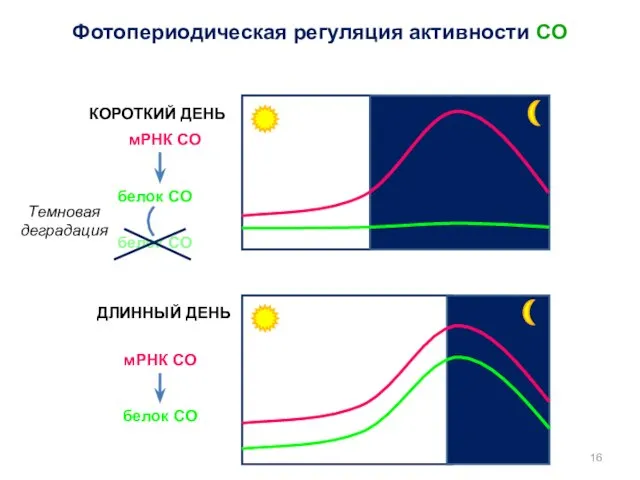
Фотопериодическая регуляция активности CO
Фотопериодическая регуляция активности CO
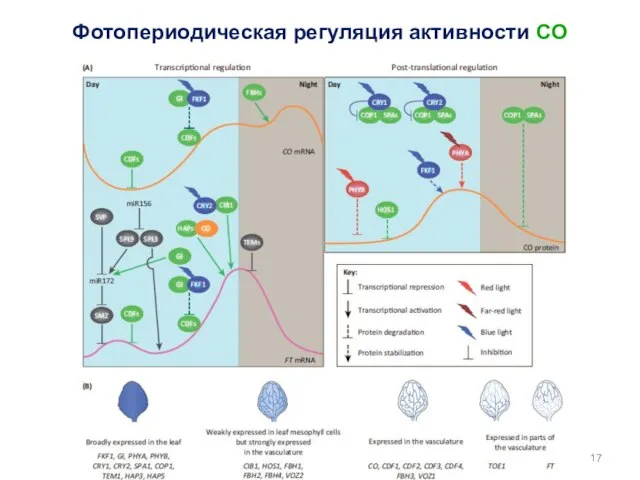
Фотопериодическая регуляция активности CO
Фотопериодическая регуляция активности CO
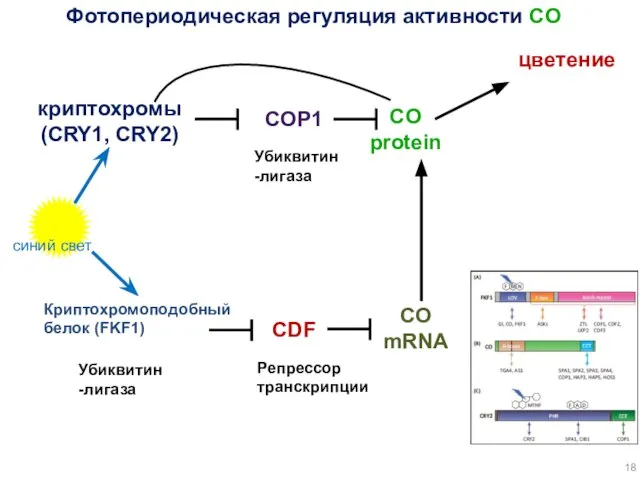
COP1
синий свет
криптохромы
(CRY1, CRY2)
CO
protein
цветение
Фотопериодическая регуляция активности CO
CO
mRNA
Криптохромоподобный белок (FKF1)
Убиквитин-лигаза
Убиквитин-лигаза
CDF
Репрессор транскрипции
COP1
синий свет
криптохромы
(CRY1, CRY2)
CO
protein
цветение
Фотопериодическая регуляция активности CO
CO
mRNA
Криптохромоподобный белок (FKF1)
Убиквитин-лигаза
Убиквитин-лигаза
CDF
Репрессор транскрипции
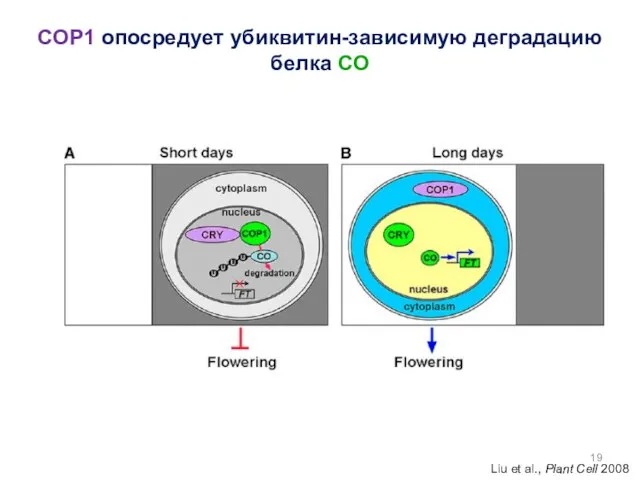
COP1 опосредует убиквитин-зависимую деградацию белка CO
Liu et al., Plant Cell 2008
COP1 опосредует убиквитин-зависимую деградацию белка CO
Liu et al., Plant Cell 2008
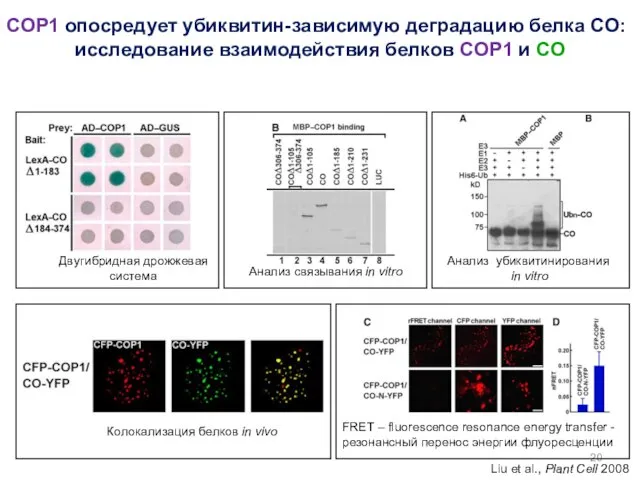
COP1 опосредует убиквитин-зависимую деградацию белка CO:
исследование взаимодействия белков COP1 и
COP1 опосредует убиквитин-зависимую деградацию белка CO:
исследование взаимодействия белков COP1 и
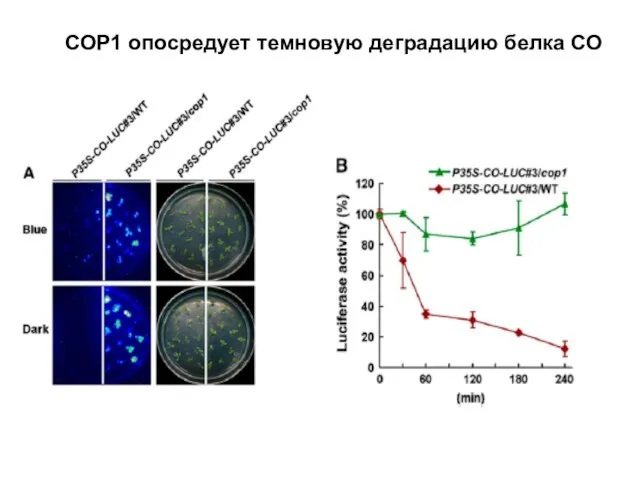
СОР1 опосредует темновую деградацию белка СО
СОР1 опосредует темновую деградацию белка СО
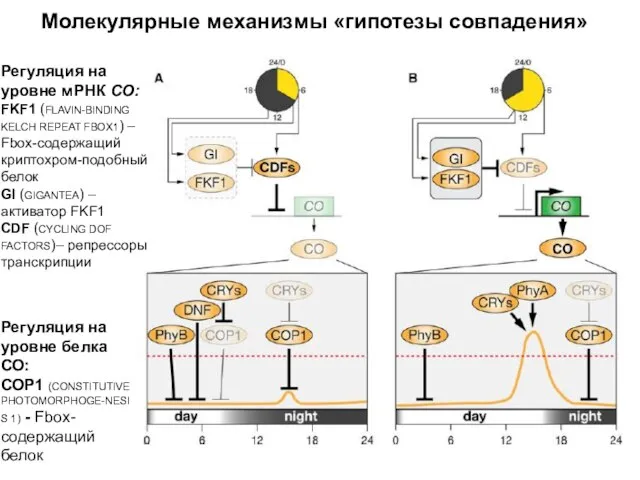
Молекулярные механизмы «гипотезы совпадения»
Регуляция на уровне мРНК СО:
FKF1 (FLAVIN-BINDING KELCH REPEAT
Молекулярные механизмы «гипотезы совпадения»
Регуляция на уровне мРНК СО:
FKF1 (FLAVIN-BINDING KELCH REPEAT
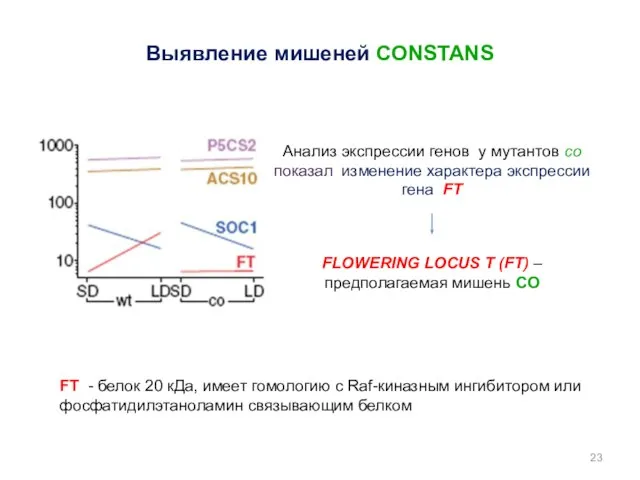
Выявление мишеней CONSTANS
FT - белок 20 кДа, имеет гомологию с Raf-киназным
Выявление мишеней CONSTANS
FT - белок 20 кДа, имеет гомологию с Raf-киназным
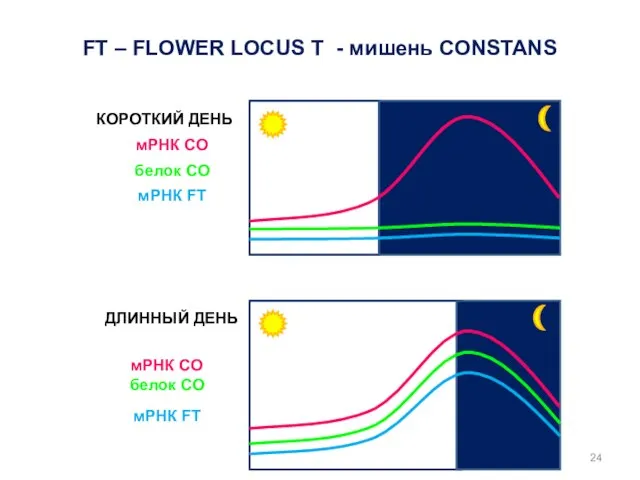
КОРОТКИЙ ДЕНЬ
ДЛИННЫЙ ДЕНЬ
мРНК CO
мРНК CO
белок CO
белок CO
мРНК FT
мРНК FT
FT – FLOWER
КОРОТКИЙ ДЕНЬ
ДЛИННЫЙ ДЕНЬ
мРНК CO
мРНК CO
белок CO
белок CO
мРНК FT
мРНК FT
FT – FLOWER
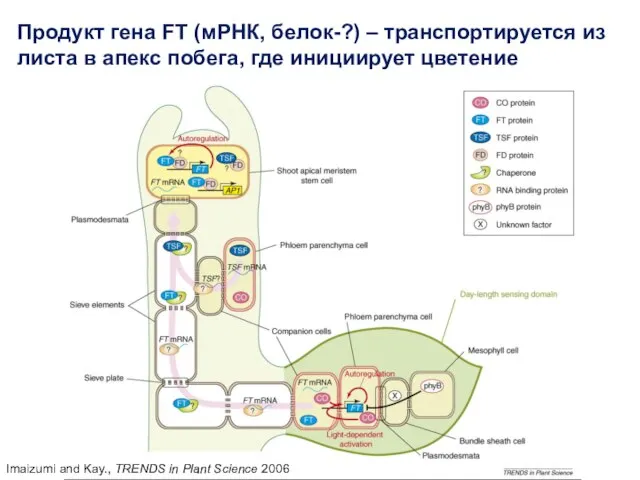
Imaizumi and Kay., TRENDS in Plant Science 2006
Продукт гена FT
Imaizumi and Kay., TRENDS in Plant Science 2006
Продукт гена FT
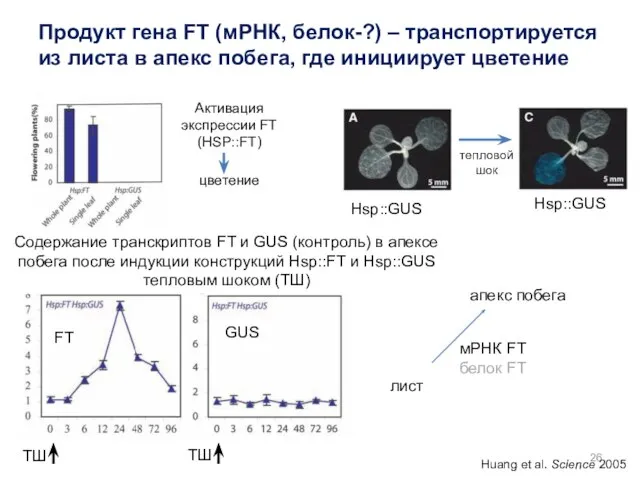
Активация экспрессии FT (HSP::FT)
цветение
ТШ
ТШ
Содержание транскриптов FT и GUS (контроль) в
Активация экспрессии FT (HSP::FT)
цветение
ТШ
ТШ
Содержание транскриптов FT и GUS (контроль) в
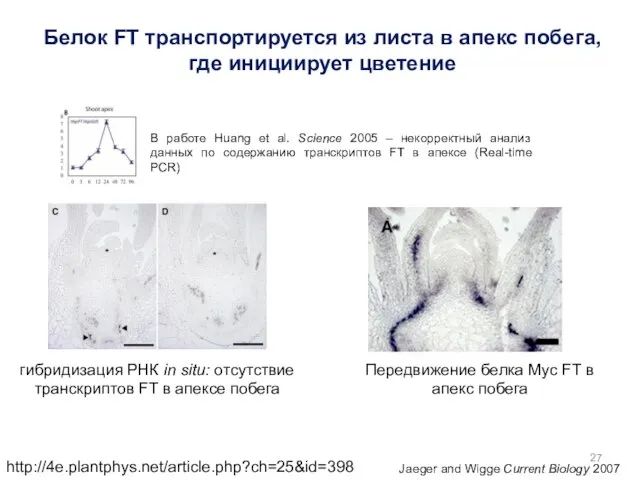
http://4e.plantphys.net/article.php?ch=25&id=398
Jaeger and Wigge Current Biology 2007
Передвижение белка Myc FT в апекс
http://4e.plantphys.net/article.php?ch=25&id=398
Jaeger and Wigge Current Biology 2007
Передвижение белка Myc FT в апекс
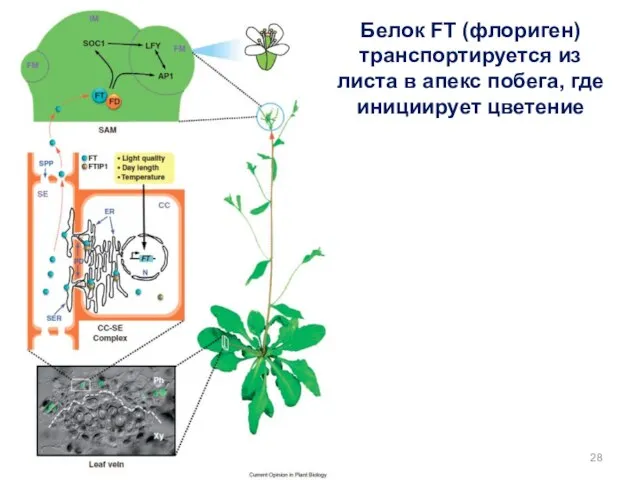
Белок FT (флориген) транспортируется из листа в апекс побега, где инициирует
Белок FT (флориген) транспортируется из листа в апекс побега, где инициирует
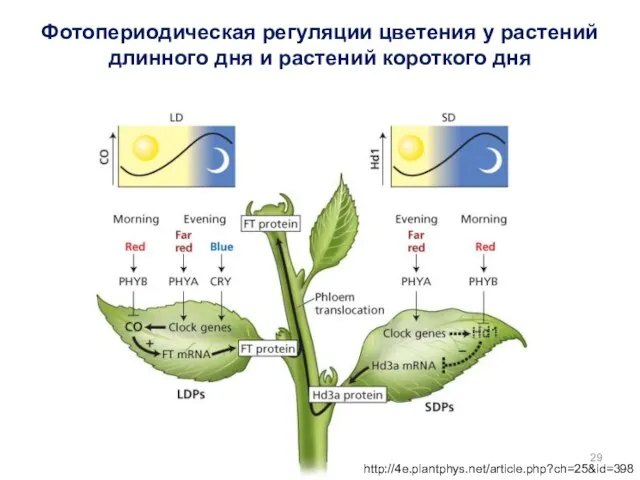
http://4e.plantphys.net/article.php?ch=25&id=398
Фотопериодическая регуляции цветения у растений длинного дня и растений короткого дня
http://4e.plantphys.net/article.php?ch=25&id=398
Фотопериодическая регуляции цветения у растений длинного дня и растений короткого дня
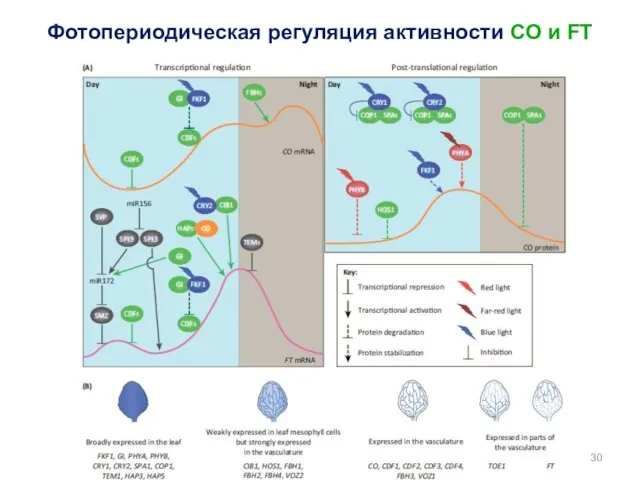
Фотопериодическая регуляция активности CO и FT
Фотопериодическая регуляция активности CO и FT
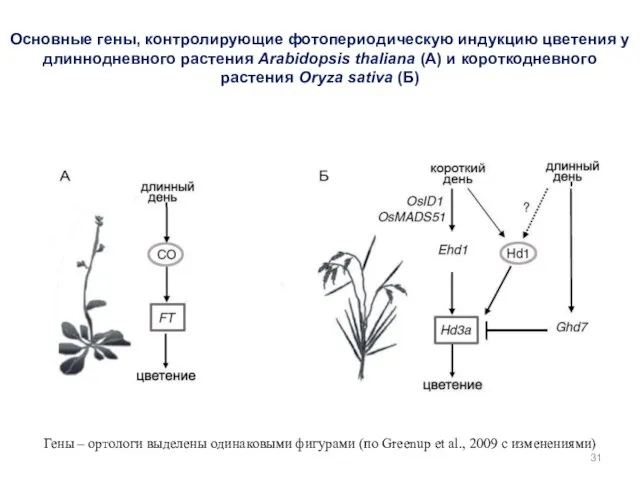
Основные гены, контролирующие фотопериодическую индукцию цветения у длиннодневного растения Arabidopsis thaliana
Основные гены, контролирующие фотопериодическую индукцию цветения у длиннодневного растения Arabidopsis thaliana

Ген TFL (Terminal Flower) арабидопсиса
Продукт гена TFL – белок - репрессор
Ген TFL (Terminal Flower) арабидопсиса
Продукт гена TFL – белок - репрессор
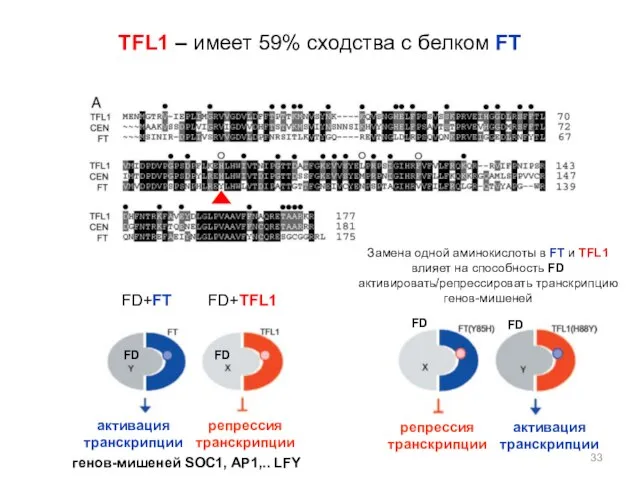
TFL1 – имеет 59% сходства с белком FT
FD
FD
FD
FD
активация транскрипции
репрессия транскрипции
активация транскрипции
репрессия
TFL1 – имеет 59% сходства с белком FT
FD
FD
FD
FD
активация транскрипции
репрессия транскрипции
активация транскрипции
репрессия
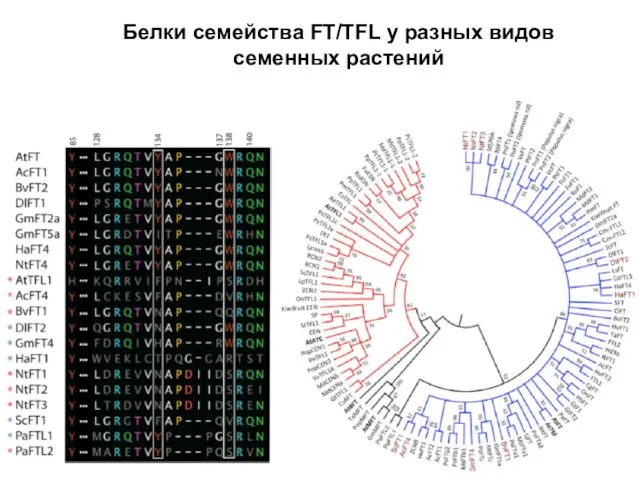
Белки семейства FT/TFL у разных видов семенных растений
Белки семейства FT/TFL у разных видов семенных растений
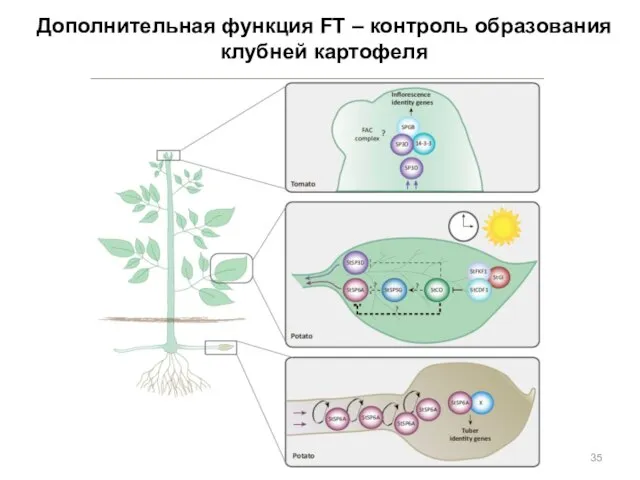
Дополнительная функция FT – контроль образования клубней картофеля
Дополнительная функция FT – контроль образования клубней картофеля

Выявление мишеней FT:
Abe et al., SCIENCE 2005
Изучение влияния мутаций по различным
Выявление мишеней FT:
Abe et al., SCIENCE 2005
Изучение влияния мутаций по различным
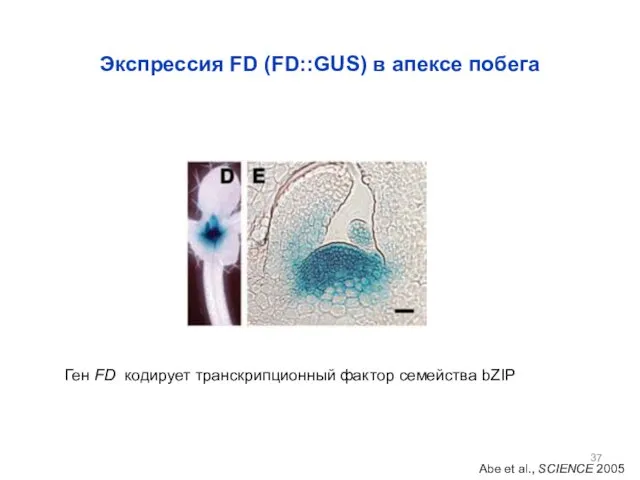
Экспрессия FD (FD::GUS) в апексе побега
Abe et al., SCIENCE 2005
Ген FD
Экспрессия FD (FD::GUS) в апексе побега
Abe et al., SCIENCE 2005
Ген FD
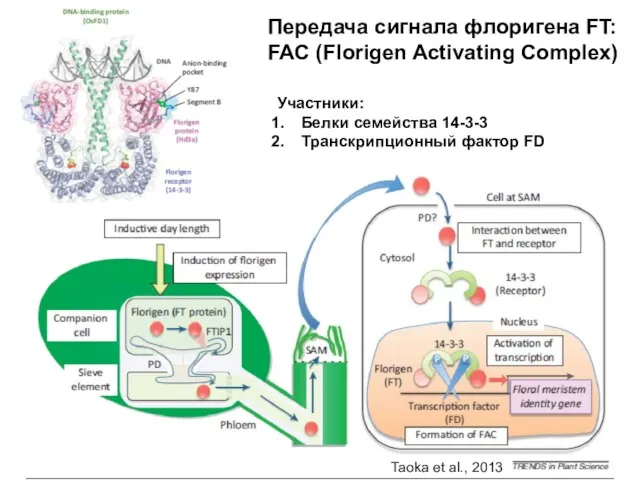
Передача сигнала флоригена FT: FAC (Florigen Activating Complex)
Участники:
Белки семейства 14-3-3
Транскрипционный
Передача сигнала флоригена FT: FAC (Florigen Activating Complex)
Участники:
Белки семейства 14-3-3
Транскрипционный
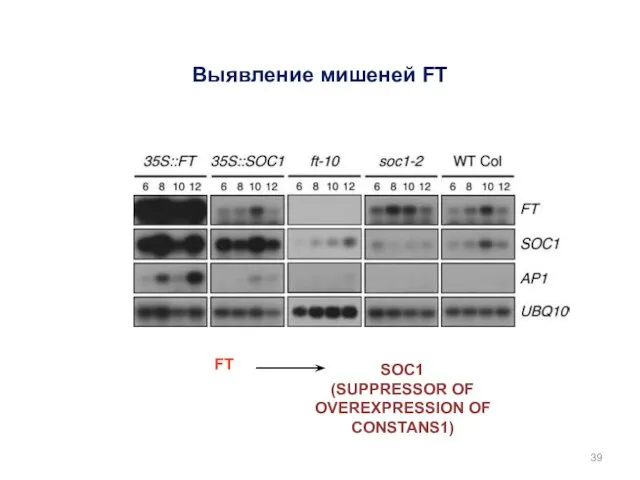
Выявление мишеней FT
FT
SOC1
(SUPPRESSOR OF OVEREXPRESSION OF CONSTANS1)
Выявление мишеней FT
FT
SOC1
(SUPPRESSOR OF OVEREXPRESSION OF CONSTANS1)
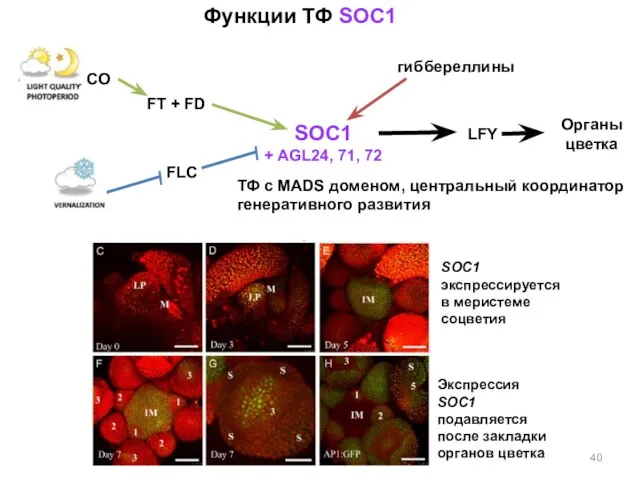
Функции ТФ SOC1
SOC1
+ AGL24, 71, 72
LFY
FT + FD
CO
FLC
гиббереллины
Органы цветка
ТФ с MADS
Функции ТФ SOC1
SOC1
+ AGL24, 71, 72
LFY
FT + FD
CO
FLC
гиббереллины
Органы цветка
ТФ с MADS
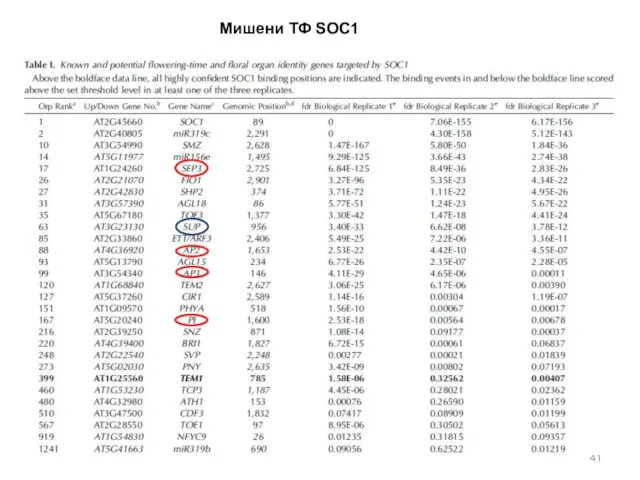
Мишени ТФ SOC1
Мишени ТФ SOC1
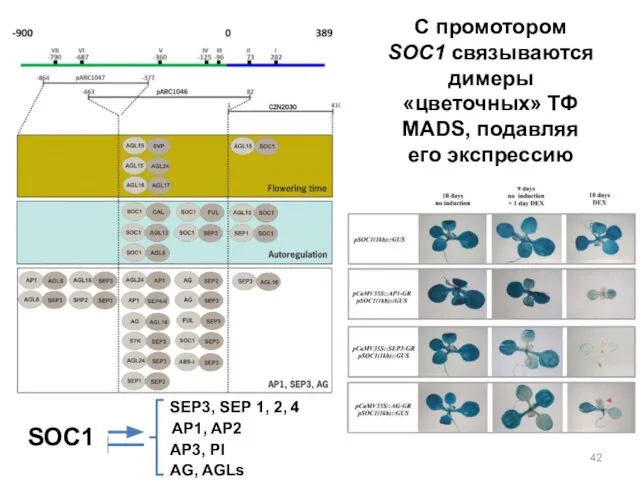
C промотором SOC1 связываются димеры «цветочных» ТФ MADS, подавляя его экспрессию
C промотором SOC1 связываются димеры «цветочных» ТФ MADS, подавляя его экспрессию
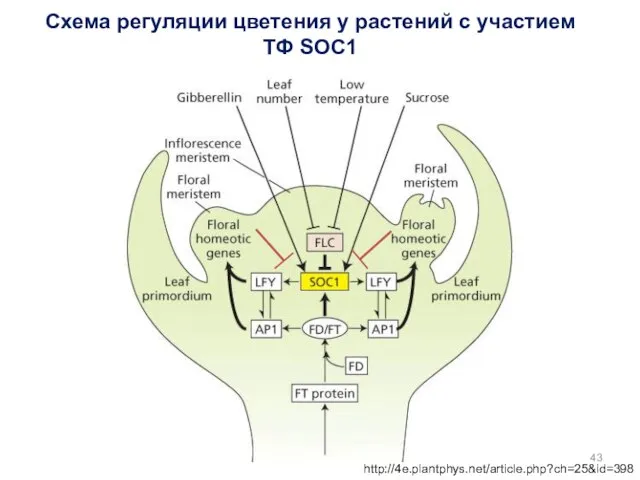
Схема регуляции цветения у растений с участием ТФ SOC1
http://4e.plantphys.net/article.php?ch=25&id=398
Схема регуляции цветения у растений с участием ТФ SOC1
http://4e.plantphys.net/article.php?ch=25&id=398
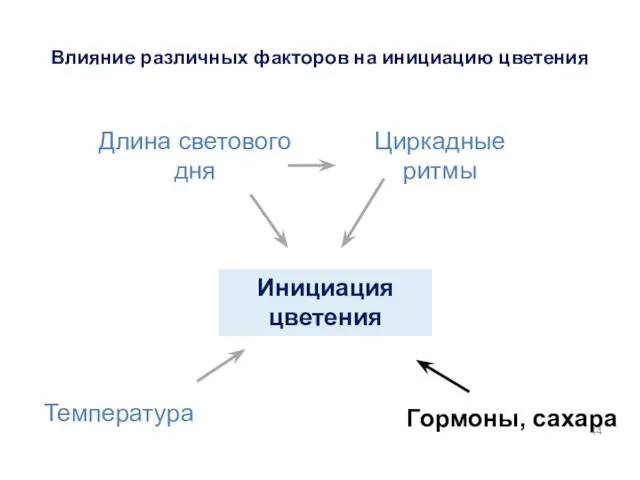
Инициация цветения
Гормоны, сахара
Температура
Длина светового дня
Циркадные ритмы
Влияние различных факторов на инициацию цветения
Инициация цветения
Гормоны, сахара
Температура
Длина светового дня
Циркадные ритмы
Влияние различных факторов на инициацию цветения

Achard et al., PNAS 2007
Антагонизм этилена и гиббереллинов в регуляции цветения
гиббереллины
цветение
этилен
Achard et al., PNAS 2007
Антагонизм этилена и гиббереллинов в регуляции цветения
гиббереллины
цветение
этилен
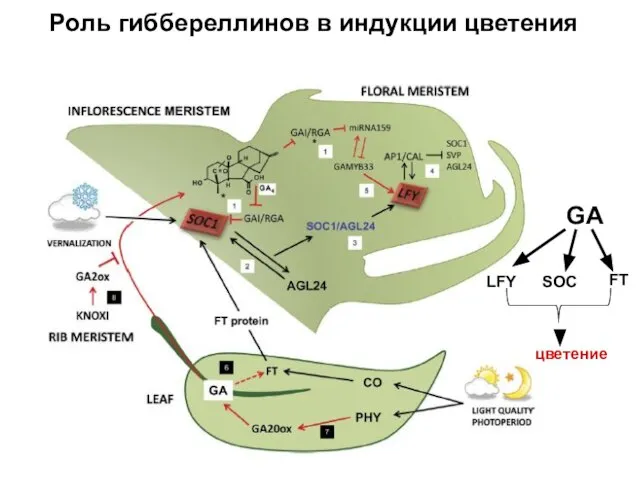
Роль гиббереллинов в индукции цветения
GA
LFY
SOC
FT
цветение
Роль гиббереллинов в индукции цветения
GA
LFY
SOC
FT
цветение
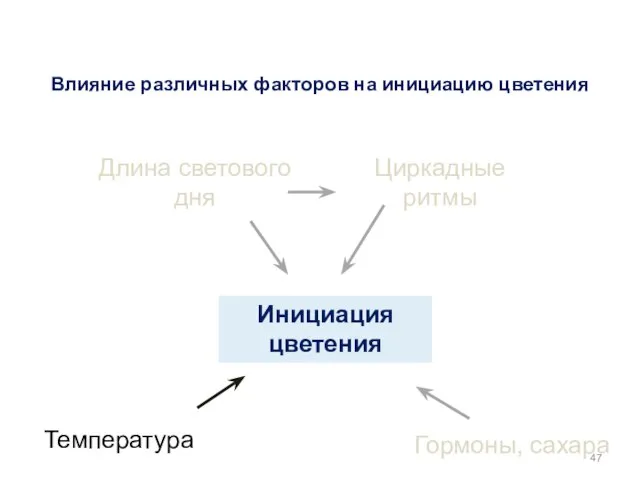
Инициация цветения
Гормоны, сахара
Температура
Длина светового дня
Циркадные ритмы
Влияние различных факторов на инициацию цветения
Инициация цветения
Гормоны, сахара
Температура
Длина светового дня
Циркадные ритмы
Влияние различных факторов на инициацию цветения
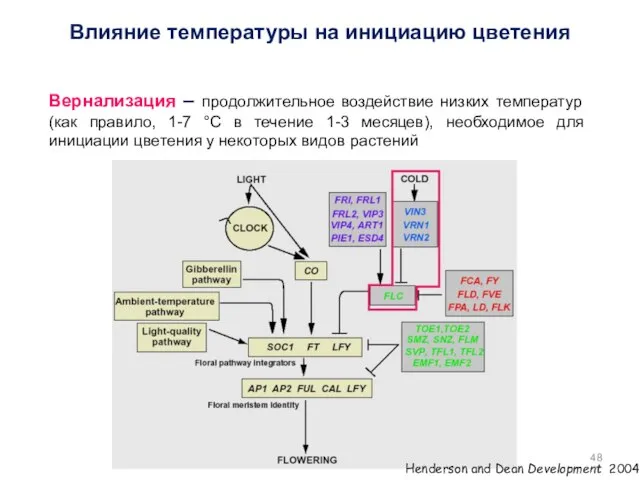
Вернализация – продолжительное воздействие низких температур (как правило, 1-7 °С в
Вернализация – продолжительное воздействие низких температур (как правило, 1-7 °С в

Молекулярные основы эпигенетических изменений:
метилирование ДНК
2. модификация структуры хроматина:
Молекулярные основы эпигенетических изменений:
метилирование ДНК
2. модификация структуры хроматина:
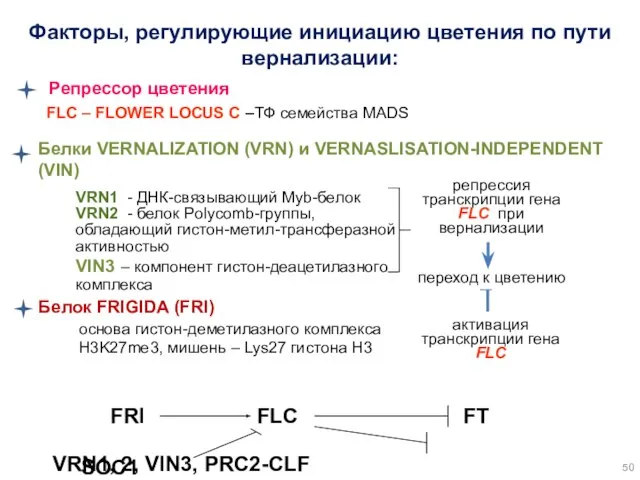
VRN1 - ДНК-связывающий Myb-белок
VRN2 - белок Polycomb-группы, обладающий гистон-метил-трансферазной активностью
VIN3 –
VRN1 - ДНК-связывающий Myb-белок
VRN2 - белок Polycomb-группы, обладающий гистон-метил-трансферазной активностью
VIN3 –
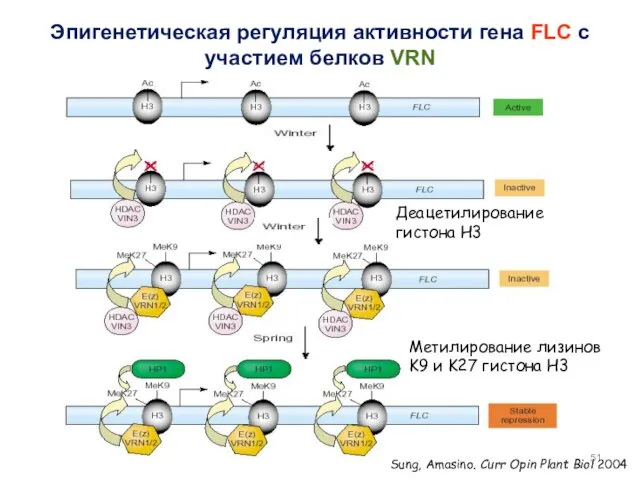
Sung, Amasino. Curr Opin Plant Biol 2004
Эпигенетическая регуляция активности гена FLC
Sung, Amasino. Curr Opin Plant Biol 2004
Эпигенетическая регуляция активности гена FLC
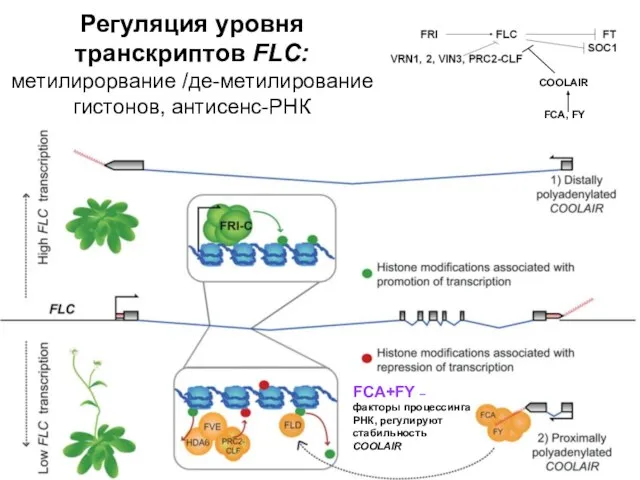
Регуляция уровня транскриптов FLC: метилирорвание /де-метилирование гистонов, антисенс-РНК
FCA+FY – факторы
Регуляция уровня транскриптов FLC: метилирорвание /де-метилирование гистонов, антисенс-РНК
FCA+FY – факторы

Drosophila melanigaster
Arabidopsis thaliana
POLYCOMB - комплекс обладает гистон –
метилтрансферазной активностью (метилирует Lys9
Drosophila melanigaster
Arabidopsis thaliana
POLYCOMB - комплекс обладает гистон –
метилтрансферазной активностью (метилирует Lys9
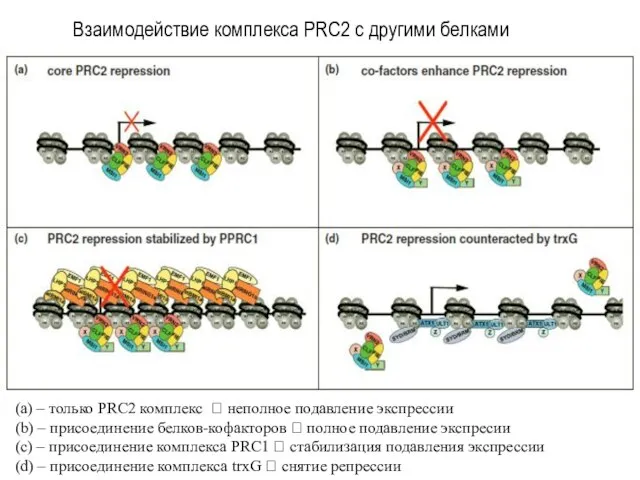
(а) – только PRC2 комплекс ? неполное подавление экспрессии
(b) – присоединение
(а) – только PRC2 комплекс ? неполное подавление экспрессии
(b) – присоединение
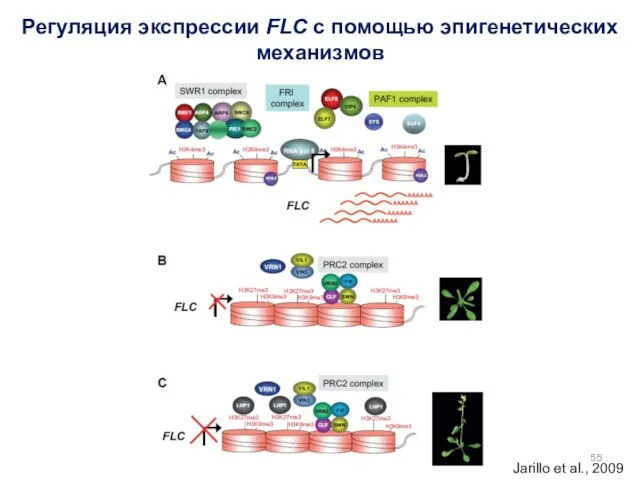
Регуляция экспрессии FLC с помощью эпигенетических механизмов
Jarillo et al., 2009
Регуляция экспрессии FLC с помощью эпигенетических механизмов
Jarillo et al., 2009
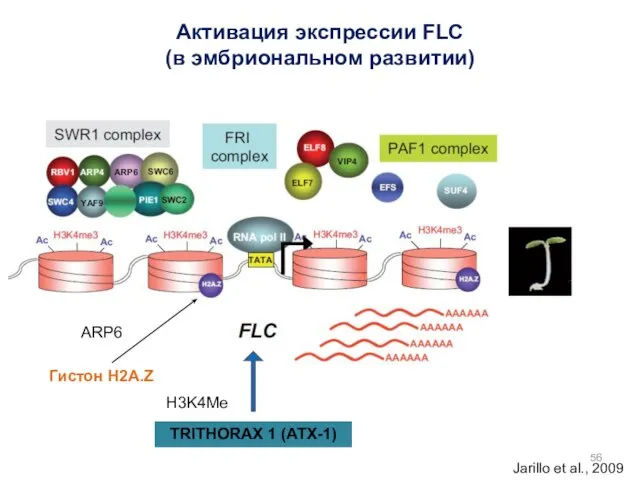
Активация экспрессии FLC
(в эмбриональном развитии)
Jarillo et al., 2009
Гистон H2A.Z
ARP6
TRITHORAX 1
Активация экспрессии FLC
(в эмбриональном развитии)
Jarillo et al., 2009
Гистон H2A.Z
ARP6
TRITHORAX 1
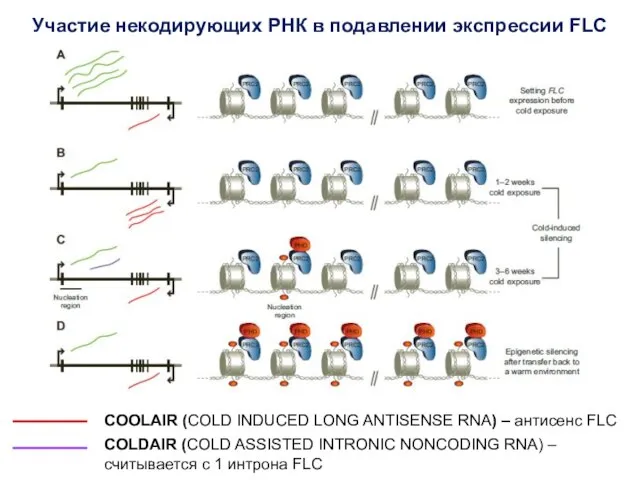
Участие некодирующих РНК в подавлении экспрессии FLC
COOLAIR (COLD INDUCED LONG ANTISENSE
Участие некодирующих РНК в подавлении экспрессии FLC
COOLAIR (COLD INDUCED LONG ANTISENSE
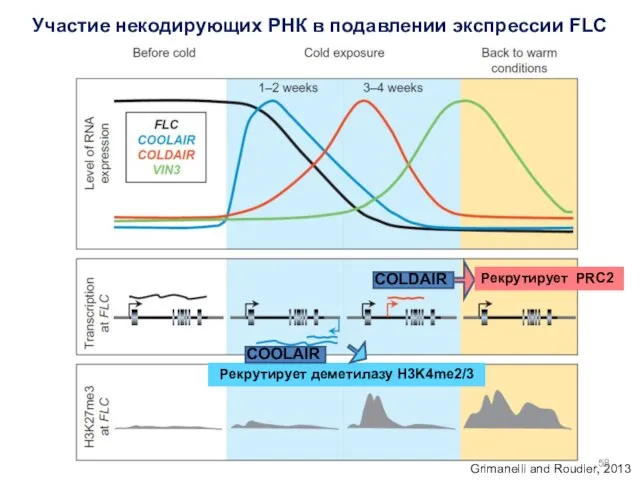
Рекрутирует деметилазу H3K4me2/3
Участие некодирующих РНК в подавлении экспрессии FLC
Grimanelli and Roudier,
Рекрутирует деметилазу H3K4me2/3
Участие некодирующих РНК в подавлении экспрессии FLC
Grimanelli and Roudier,
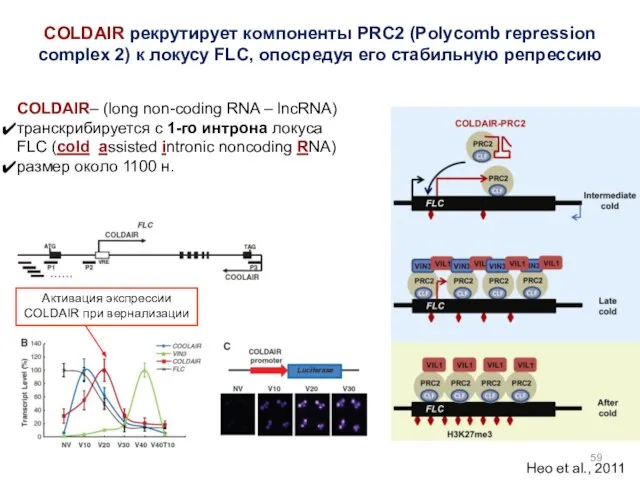
COLDAIR рекрутирует компоненты PRC2 (Polycomb repression complex 2) к локусу FLC,
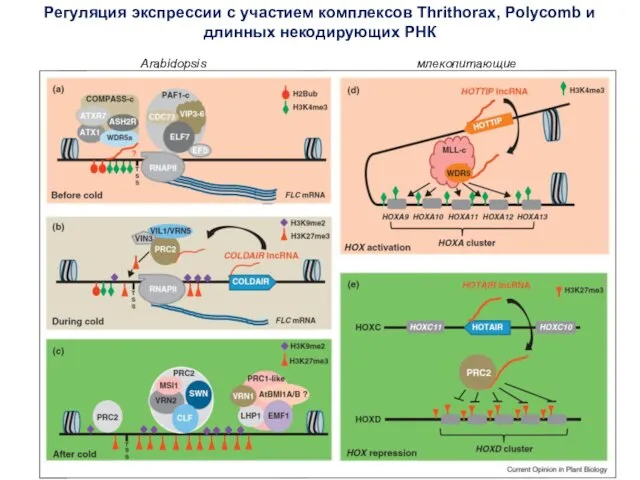
Регуляция экспрессии с участием комплексов Thrithorax, Polycomb и длинных некодирующих РНК
Arabidopsis
млекопитающие
Регуляция экспрессии с участием комплексов Thrithorax, Polycomb и длинных некодирующих РНК
Arabidopsis
млекопитающие


- LEAFY (LFY )
- APETALA1 (AP1)
- CAULIFLOWER (CAL )
- UNUSUAL FLORAL
- LEAFY (LFY )
- APETALA1 (AP1)
- CAULIFLOWER (CAL )
- UNUSUAL FLORAL
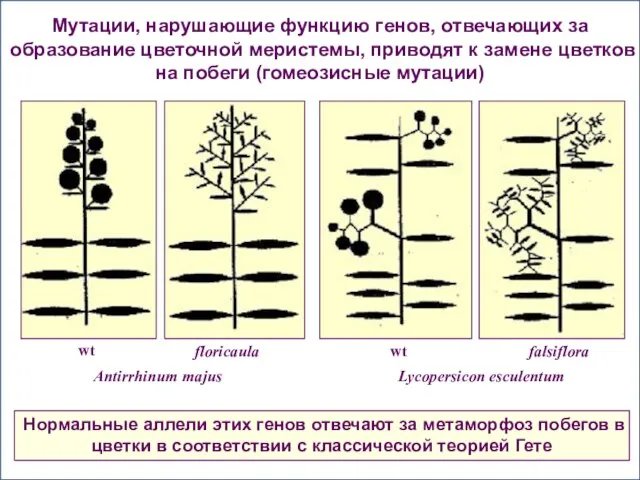
Мутации, нарушающие функцию генов, отвечающих за образование цветочной меристемы, приводят к
Мутации, нарушающие функцию генов, отвечающих за образование цветочной меристемы, приводят к

дикий тип
lfy
Фенотип мутации lfy у арабидопсиса
lfy
дикий тип
lfy
Фенотип мутации lfy у арабидопсиса
lfy
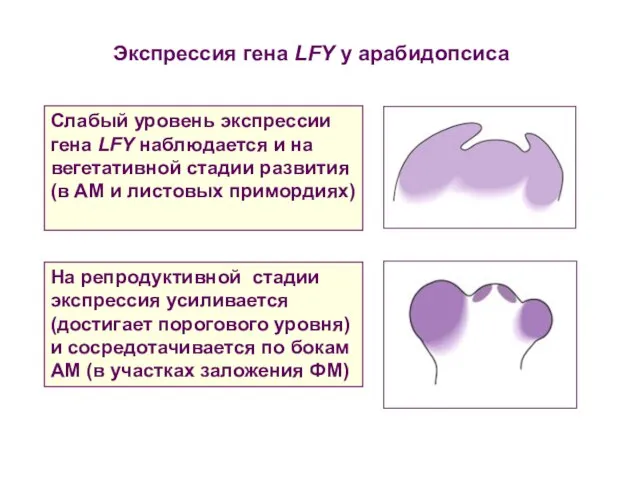
Слабый уровень экспрессии гена LFY наблюдается и на вегетативной стадии развития
Слабый уровень экспрессии гена LFY наблюдается и на вегетативной стадии развития
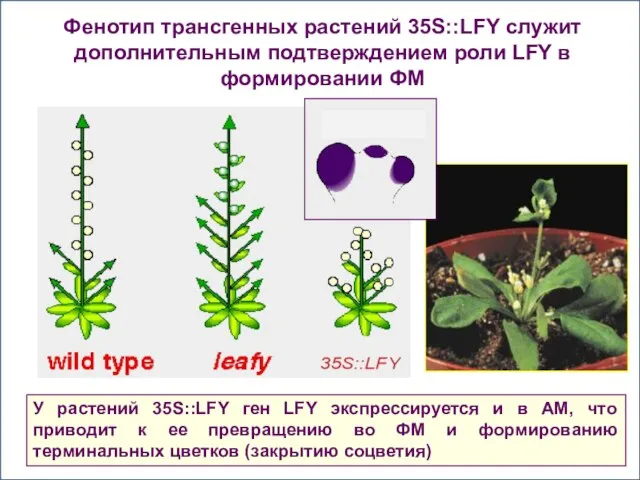
Фенотип трансгенных растений 35S::LFY служит дополнительным подтверждением роли LFY в формировании
Фенотип трансгенных растений 35S::LFY служит дополнительным подтверждением роли LFY в формировании

Ортологи гена LFY
FLORICAULA (FLO) – Antirrhinum majus
NFL – Nicotiana tabacum
ALF –
Ортологи гена LFY
FLORICAULA (FLO) – Antirrhinum majus NFL – Nicotiana tabacum ALF –
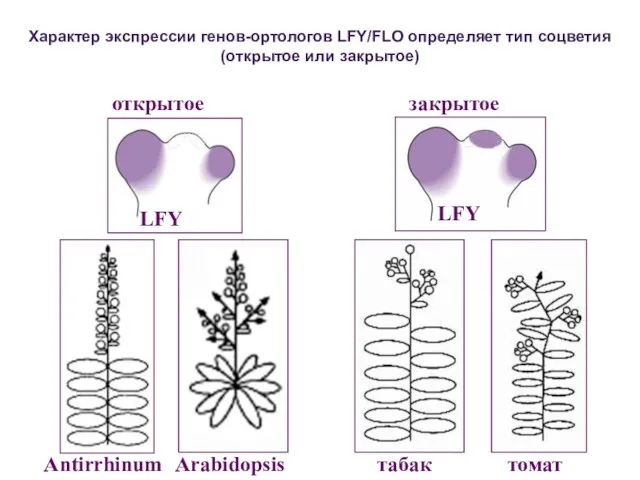
Характер экспрессии генов-ортологов LFY/FLO определяет тип соцветия (открытое или закрытое)
открытое
закрытое
LFY
Характер экспрессии генов-ортологов LFY/FLO определяет тип соцветия (открытое или закрытое)
открытое
закрытое
LFY

Ген сосны NEEDLY под промотором гена LFY супрессирует мутацию lfy-26
Ген сосны NEEDLY под промотором гена LFY супрессирует мутацию lfy-26

Наличие ортологов генов LFY/FLO у голосеменных и покрытосеменных растений позволяет предполагать,
Наличие ортологов генов LFY/FLO у голосеменных и покрытосеменных растений позволяет предполагать,
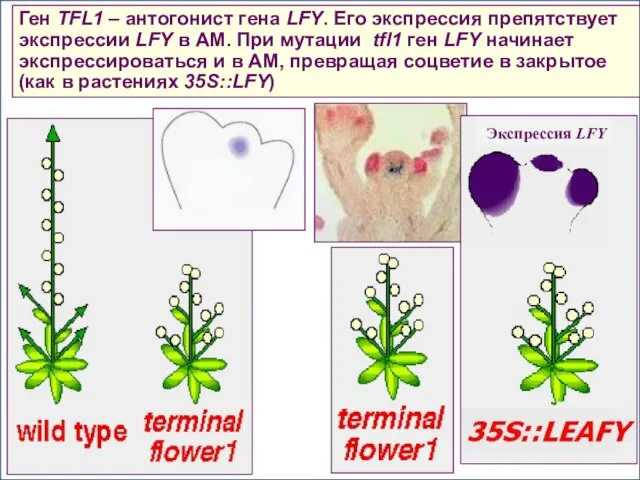
Ген TFL1 – антогонист гена LFY. Его экспрессия препятствует экспрессии LFY
Ген TFL1 – антогонист гена LFY. Его экспрессия препятствует экспрессии LFY
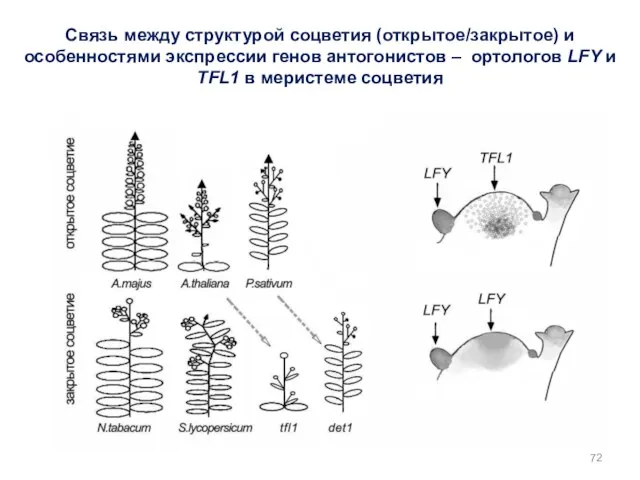
Связь между структурой соцветия (открытое/закрытое) и особенностями экспрессии генов антогонистов –
Связь между структурой соцветия (открытое/закрытое) и особенностями экспрессии генов антогонистов –
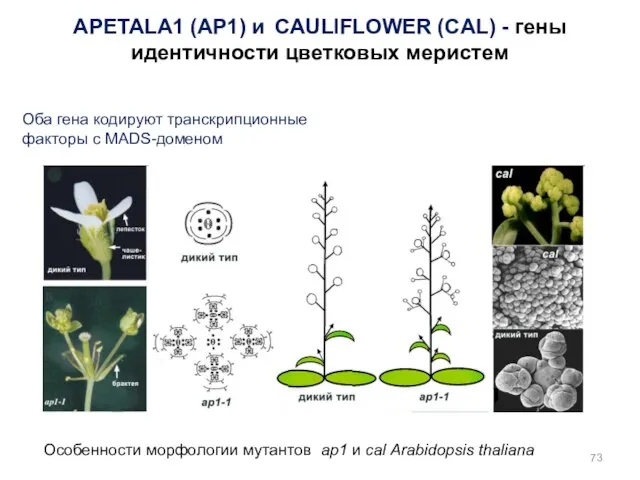
Особенности морфологии мутантов ap1 и cal Arabidopsis thaliana
APETALA1 (AP1) и CAULIFLOWER
Особенности морфологии мутантов ap1 и cal Arabidopsis thaliana
APETALA1 (AP1) и CAULIFLOWER

У мутанта ap1cal меристемы соцветия формируют новые меристемы соцветия, но не
У мутанта ap1cal меристемы соцветия формируют новые меристемы соцветия, но не

Гомеозисные
мутанты
арабидопсиса
Генетический контроль дифференцировки органов цветка
Гомеозисные
мутанты
арабидопсиса
Генетический контроль дифференцировки органов цветка
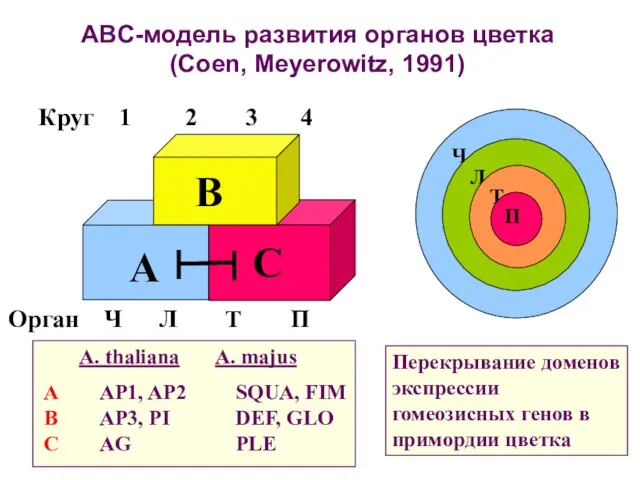
ABC-модель развития органов цветка (Coen, Meyerowitz, 1991)
Перекрывание доменов экспрессии гомеозисных генов
ABC-модель развития органов цветка (Coen, Meyerowitz, 1991)
Перекрывание доменов экспрессии гомеозисных генов
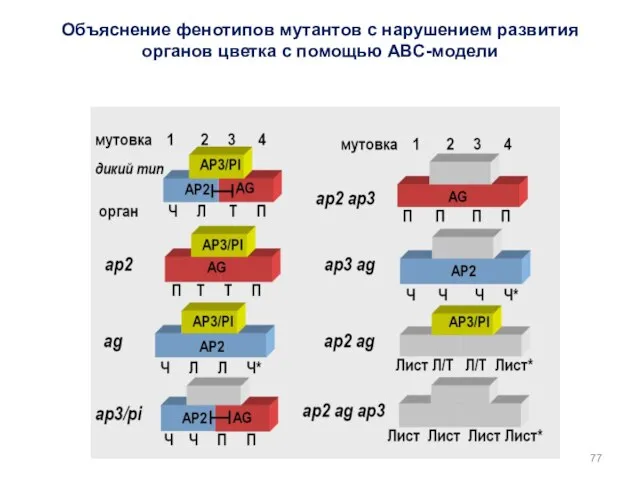
Объяснение фенотипов мутантов с нарушением развития органов цветка с помощью АВС-модели
Объяснение фенотипов мутантов с нарушением развития органов цветка с помощью АВС-модели
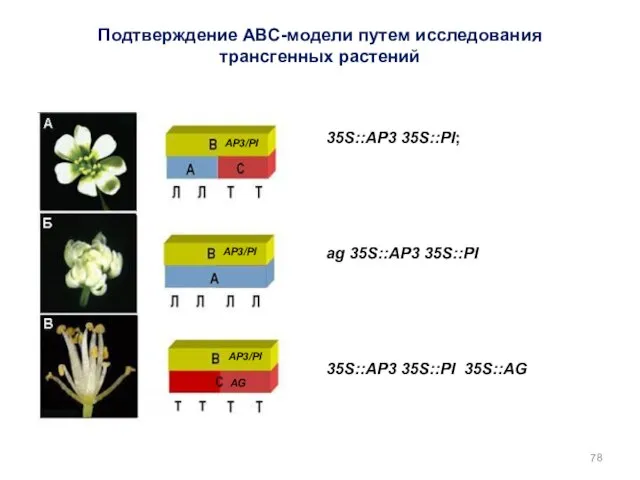
35S::AP3 35S::PI;
ag 35S::AP3 35S::PI
35S::AP3 35S::PI 35S::AG
AP3/PI
AP3/PI
AP3/PI
AG
Подтверждение АВС-модели путем исследования трансгенных
35S::AP3 35S::PI;
ag 35S::AP3 35S::PI
35S::AP3 35S::PI 35S::AG
AP3/PI
AP3/PI
AP3/PI
AG
Подтверждение АВС-модели путем исследования трансгенных
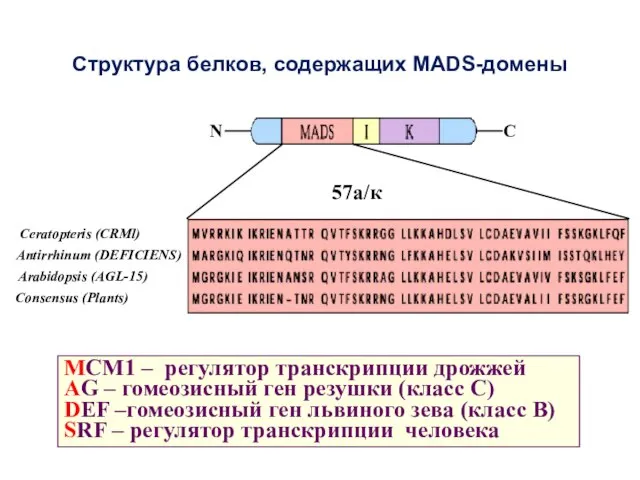
Структура белков, содержащих MADS-домены
MCM1 – регулятор транскрипции дрожжей
AG – гомеозисный ген
Структура белков, содержащих MADS-домены
MCM1 – регулятор транскрипции дрожжей
AG – гомеозисный ген
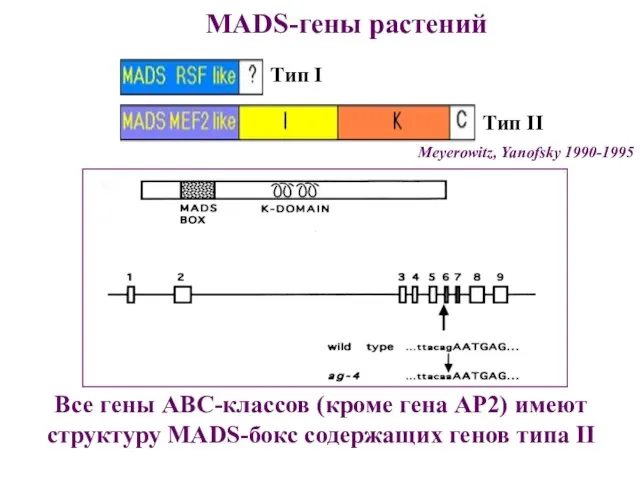
Meyerowitz, Yanofsky 1990-1995
Тип I
MADS-гены растений
Все гены АВС-классов (кроме гена АР2) имеют
Meyerowitz, Yanofsky 1990-1995
Тип I
MADS-гены растений
Все гены АВС-классов (кроме гена АР2) имеют
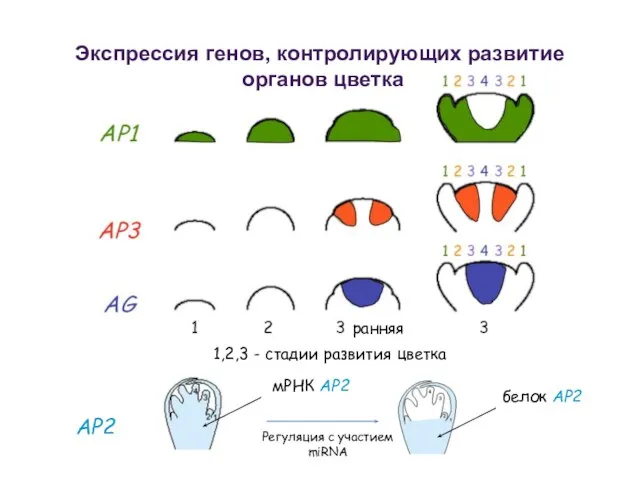
AP2
1,2,3 - стадии развития цветка
ранняя
Экспрессия генов, контролирующих развитие
органов
AP2
1,2,3 - стадии развития цветка
ранняя
Экспрессия генов, контролирующих развитие
органов

A. wt B. ap2-9 C. 35S::MIR172a-1
Фенотип трансгенных растений 35S::MIR172a-1 повторяет
A. wt B. ap2-9 C. 35S::MIR172a-1
Фенотип трансгенных растений 35S::MIR172a-1 повторяет
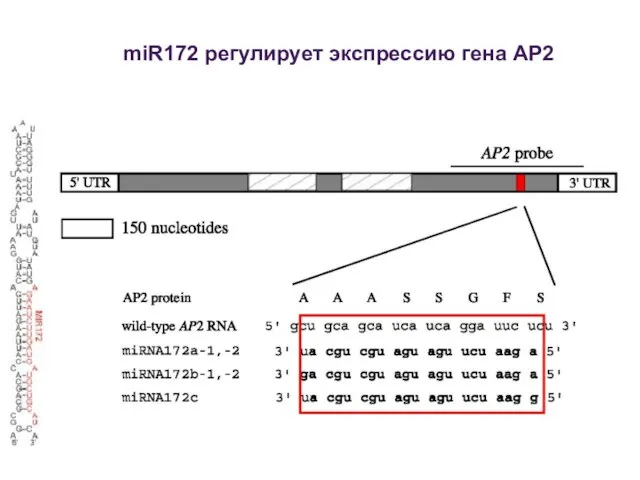
miR172 регулирует экспрессию гена AP2
miR172 регулирует экспрессию гена AP2
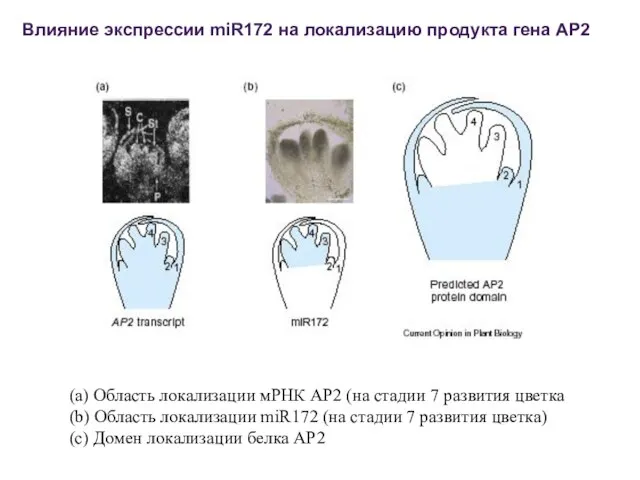
(a) Область локализации мРНК AP2 (на стадии 7 развития цветка
(b) Область
(b) Область
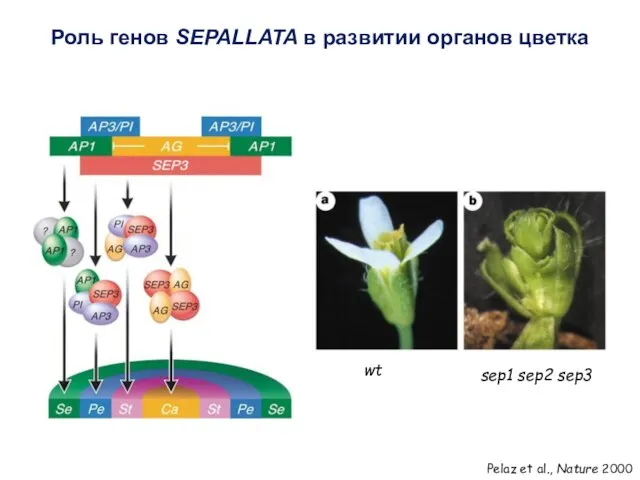
sep1 sep2 sep3
Pelaz et al., Nature 2000
wt
Роль генов SEPALLATA в развитии
sep1 sep2 sep3
Pelaz et al., Nature 2000
wt
Роль генов SEPALLATA в развитии
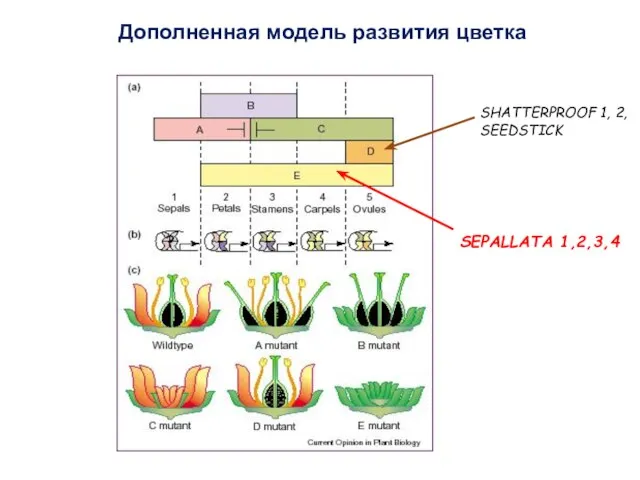
Дополненная модель развития цветка
SHATTERPROOF 1, 2,
SEEDSTICK
SEPALLATA 1,2,3,4
Дополненная модель развития цветка
SHATTERPROOF 1, 2,
SEEDSTICK
SEPALLATA 1,2,3,4
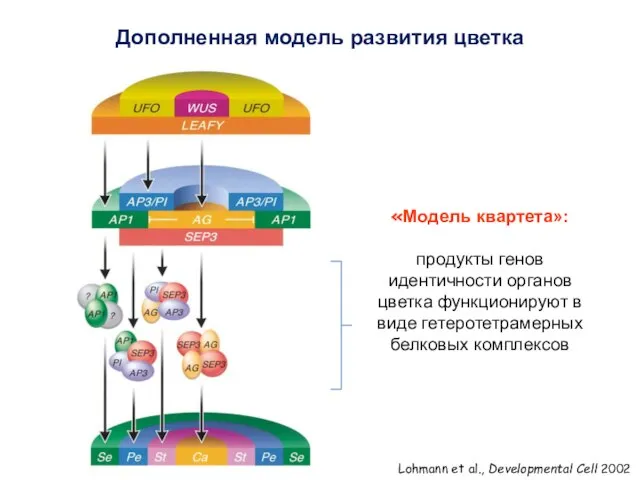
Дополненная модель развития цветка
Lohmann et al., Developmental Cell 2002
«Модель квартета»:
продукты генов
Дополненная модель развития цветка
Lohmann et al., Developmental Cell 2002
«Модель квартета»:
продукты генов
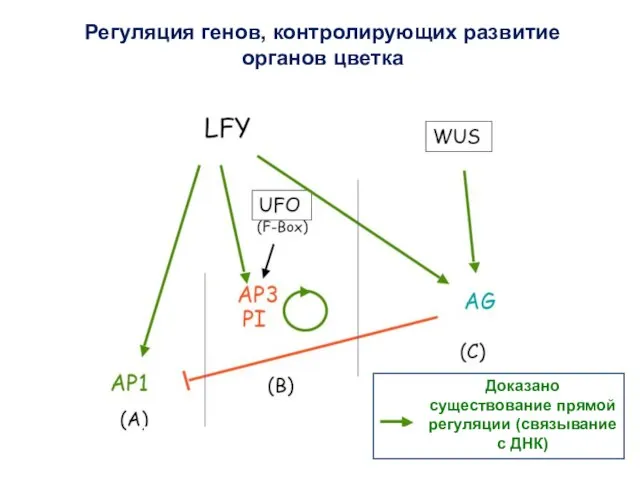
Доказано существование прямой регуляции (связывание с ДНК)
Регуляция генов, контролирующих развитие органов
Доказано существование прямой регуляции (связывание с ДНК)
Регуляция генов, контролирующих развитие органов
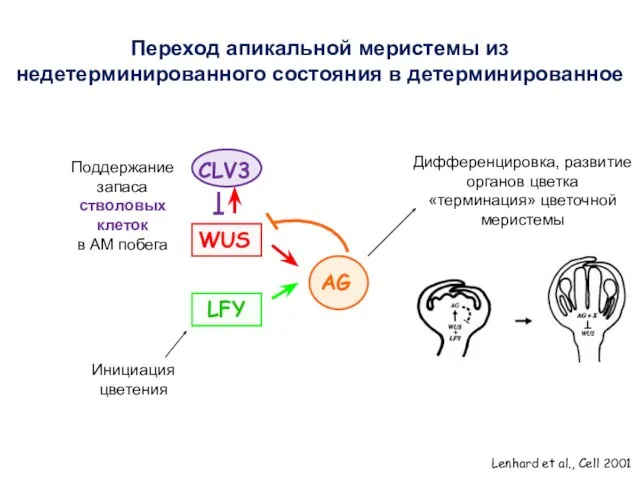
Lenhard et al., Cell 2001
CLV3
WUS
Поддержание запаса стволовых клеток
в АМ побега
Lenhard et al., Cell 2001
CLV3
WUS
Поддержание запаса стволовых клеток
в АМ побега
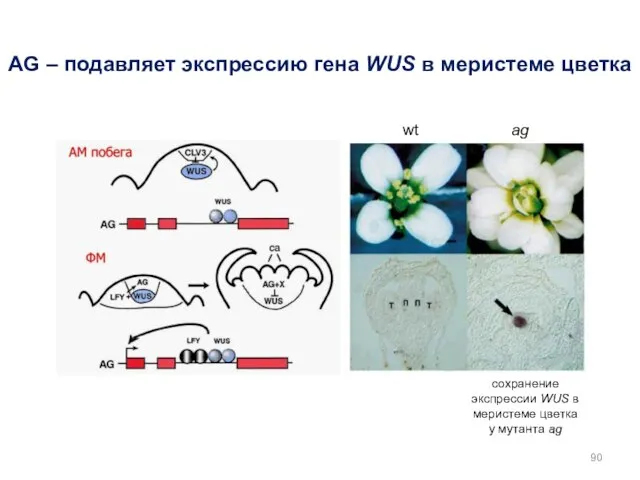
AG – подавляет экспрессию гена WUS в меристеме цветка
сохранение экспрессии WUS
AG – подавляет экспрессию гена WUS в меристеме цветка
сохранение экспрессии WUS
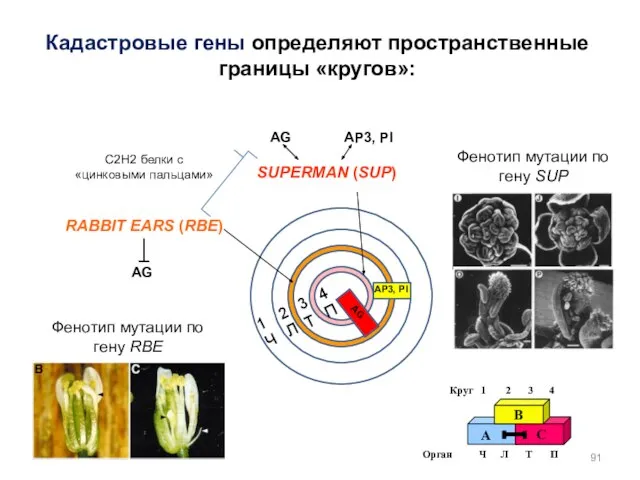
Кадастровые гены определяют пространственные границы «кругов»:
SUPERMAN (SUP)
RABBIT EARS (RBE)
2 3 4
Ч
Кадастровые гены определяют пространственные границы «кругов»:
SUPERMAN (SUP)
RABBIT EARS (RBE)
2 3 4
Ч
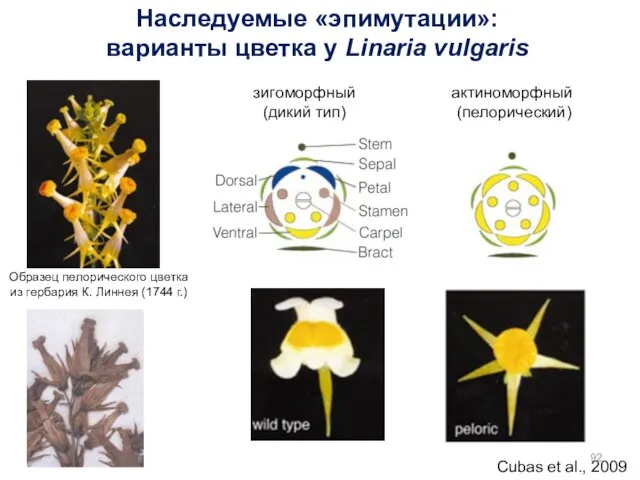
Cubas et al., 2009
Образец пелорического цветка из гербария К. Линнея (1744
Cubas et al., 2009
Образец пелорического цветка из гербария К. Линнея (1744
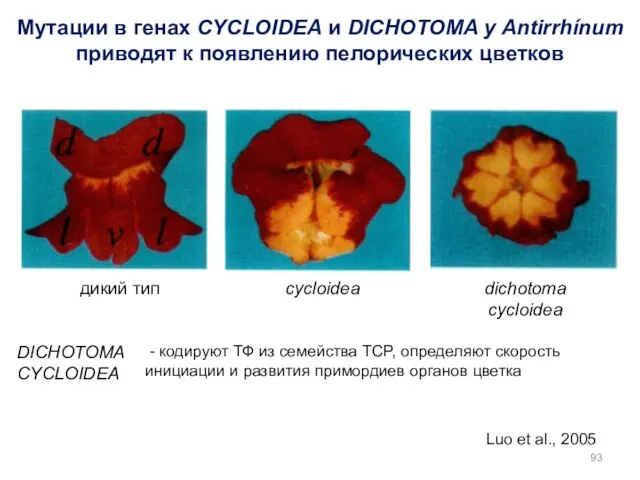
Luo et al., 2005
Мутации в генах CYCLOIDEA и DICHOTOMA у Antirrhínum
Luo et al., 2005
Мутации в генах CYCLOIDEA и DICHOTOMA у Antirrhínum
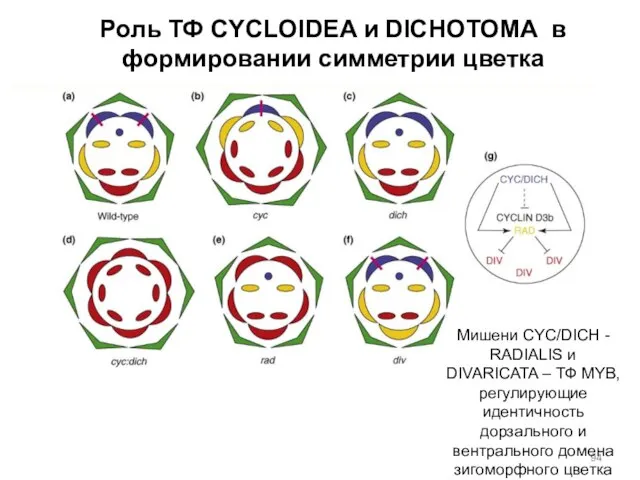
Роль ТФ CYCLOIDEA и DICHOTOMA в формировании симметрии цветка
Мишени CYC/DICH
Роль ТФ CYCLOIDEA и DICHOTOMA в формировании симметрии цветка
Мишени CYC/DICH

Анализ метилирования в локусе Lvcyc у пелорических «мутантов»льнянки с помощью чувствительных
Анализ метилирования в локусе Lvcyc у пелорических «мутантов»льнянки с помощью чувствительных

Гиперметилирование локуса SUPERMAN у «эпимутантов» clark kent (clk)
дикий тип superman clark
Гиперметилирование локуса SUPERMAN у «эпимутантов» clark kent (clk)
дикий тип superman clark
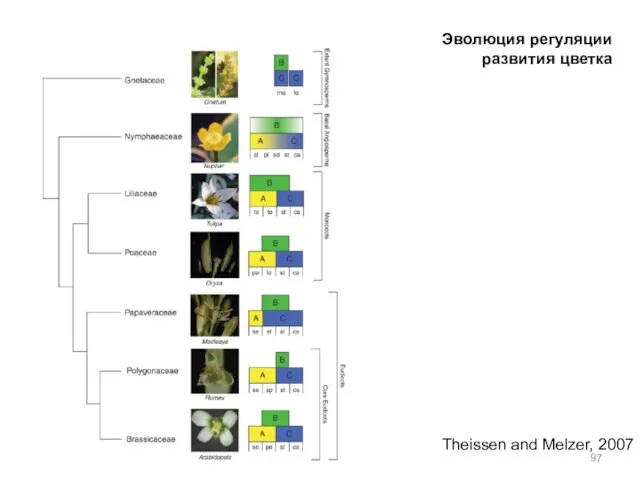
Theissen and Melzer, 2007
Эволюция регуляции развития цветка
Theissen and Melzer, 2007
Эволюция регуляции развития цветка
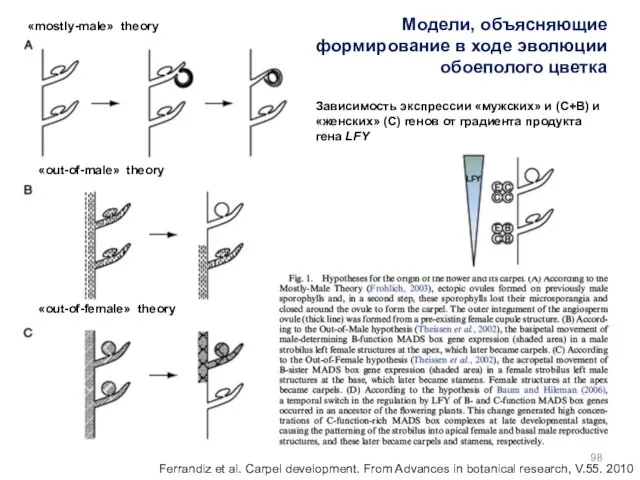
Модели, объясняющие формирование в ходе эволюции обоеполого цветка
«mostly-male» theory
«out-of-male» theory
Модели, объясняющие формирование в ходе эволюции обоеполого цветка
«mostly-male» theory
«out-of-male» theory
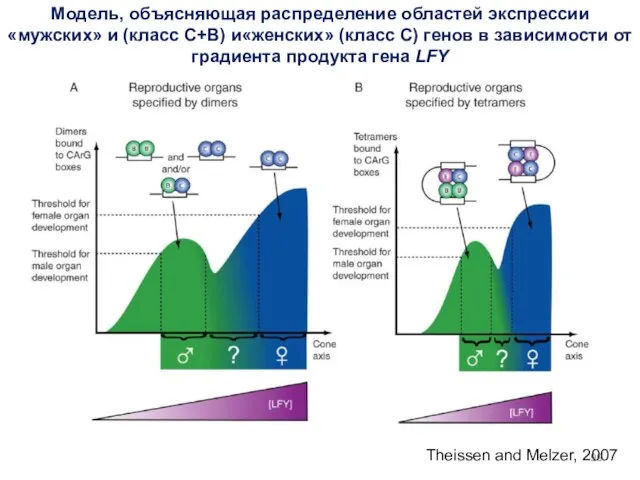
Theissen and Melzer, 2007
Модель, объясняющая распределение областей экспрессии «мужских» и (класс
Theissen and Melzer, 2007
Модель, объясняющая распределение областей экспрессии «мужских» и (класс
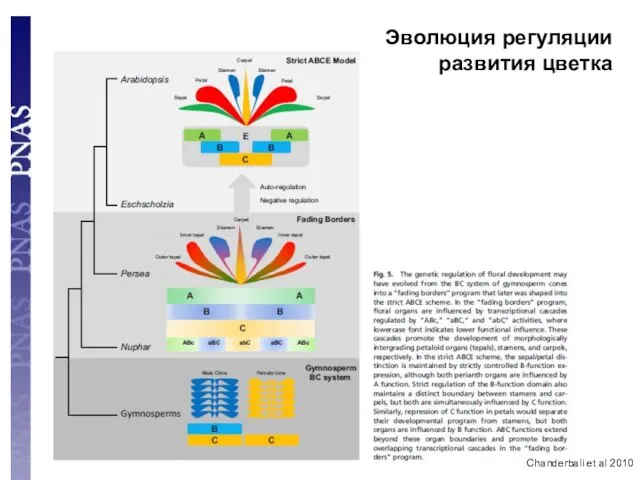
Chanderbali et al 2010
Эволюция регуляции развития цветка
Chanderbali et al 2010
Эволюция регуляции развития цветка
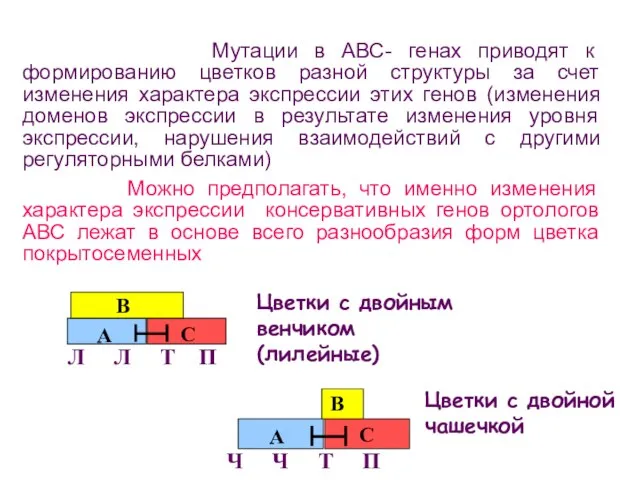
Цветки с двойным венчиком (лилейные)
Цветки с двойной чашечкой
Мутации в АВС-
Цветки с двойным венчиком (лилейные)
Цветки с двойной чашечкой
Мутации в АВС-

Модифицированная модель
Формирование цветков однодольных (Лилейные)
Модифицированная модель
Формирование цветков однодольных (Лилейные)

TGDEFA, TGDEFB
GLO-подобные гены
DEF-подобные гены
Tulipa gesneriana
Гены класса В
TGGLO
Lilium regale
LRDEF
LRGLOA, LRGLOB
Lilium longiflorum
LMADS1
?
Tricyrtis affinis
TriaDEF
TriaGLO
GLO-подобные гены
DEF-подобные гены
Tulipa gesneriana
Гены класса В
TGGLO
Lilium regale
LRDEF
LRGLOA, LRGLOB
Lilium longiflorum
LMADS1
?
Tricyrtis affinis
TriaDEF
TriaGLO
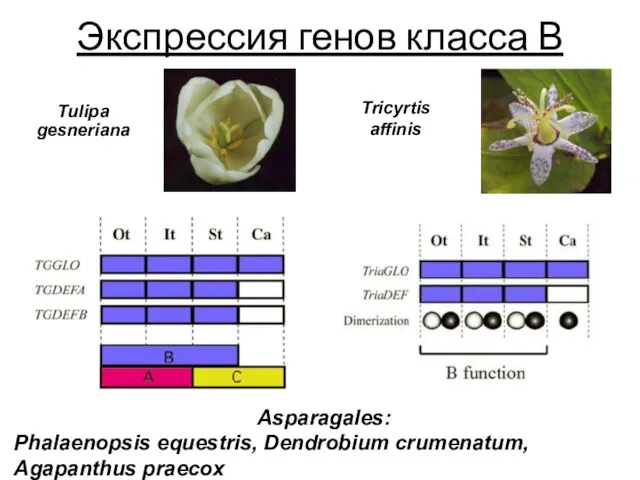
Tulipa gesneriana
Экспрессия генов класса В
Tricyrtis affinis
Asparagales:
Phalaenopsis equestris, Dendrobium crumenatum, Agapanthus praecox
Tulipa gesneriana
Экспрессия генов класса В
Tricyrtis affinis
Asparagales:
Phalaenopsis equestris, Dendrobium crumenatum, Agapanthus praecox
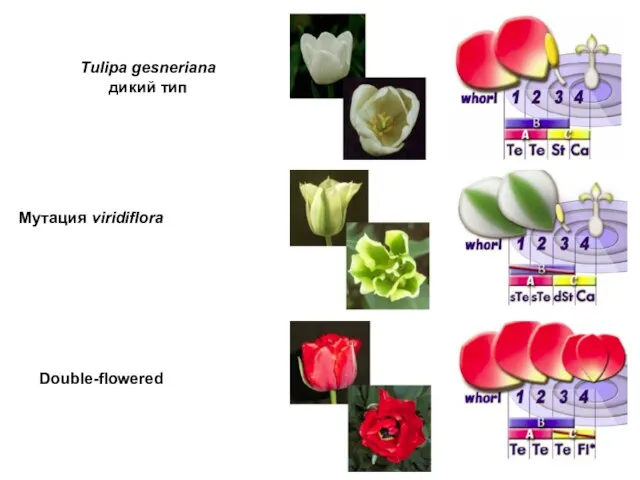
Мутация viridiflora
Tulipa gesneriana
дикий тип
Double-flowered
Мутация viridiflora
Tulipa gesneriana
дикий тип
Double-flowered
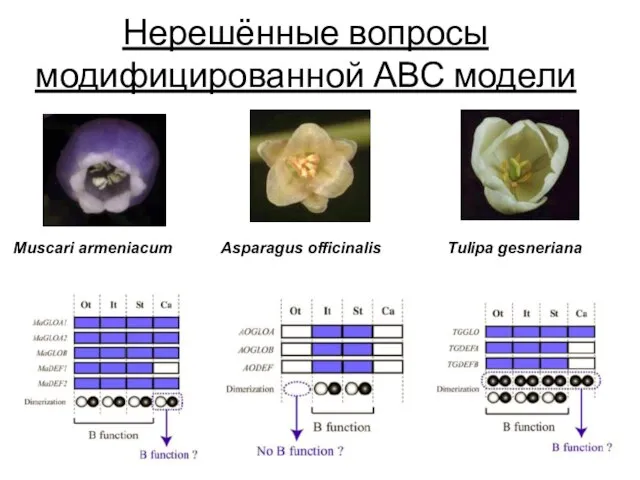
Нерешённые вопросы модифицированной АВС модели
Muscari armeniacum
Asparagus officinalis
Tulipa gesneriana
Нерешённые вопросы модифицированной АВС модели
Muscari armeniacum
Asparagus officinalis
Tulipa gesneriana
 Генетика – прошлое, настоящее, будущее Проект юных биологов Руководитель Караваева Н.М
Генетика – прошлое, настоящее, будущее Проект юных биологов Руководитель Караваева Н.М Головной мозг
Головной мозг Презентация на тему "ЗЕМНОВОДНЫЕ" - скачать бесплатно презентации по Биологии
Презентация на тему "ЗЕМНОВОДНЫЕ" - скачать бесплатно презентации по Биологии Микробиологическое получение белковых продуктов
Микробиологическое получение белковых продуктов Организация и биология надкласса рыб. Эволюция водных позвоночных
Организация и биология надкласса рыб. Эволюция водных позвоночных Биология, как наука
Биология, как наука Природоохранительная акция Покормите птиц зимой
Природоохранительная акция Покормите птиц зимой Витамин B5
Витамин B5 Влияние радиации на живые организмы Ионизирующая радиация называется «ионизирующей» потому, что при прохождении через лю
Влияние радиации на живые организмы Ионизирующая радиация называется «ионизирующей» потому, что при прохождении через лю Презентация на тему "Размножение ползучими побегами" - скачать презентации по Биологии
Презентация на тему "Размножение ползучими побегами" - скачать презентации по Биологии Подтип Tracheata (трахейнодышащие)
Подтип Tracheata (трахейнодышащие) Презентация Богомол
Презентация Богомол Мышцы нижней конечности
Мышцы нижней конечности Эволюция пищеварительной системы
Эволюция пищеварительной системы Презентация на тему "Первые представители рода Homo" - скачать презентации по Биологии
Презентация на тему "Первые представители рода Homo" - скачать презентации по Биологии Дыхательная система
Дыхательная система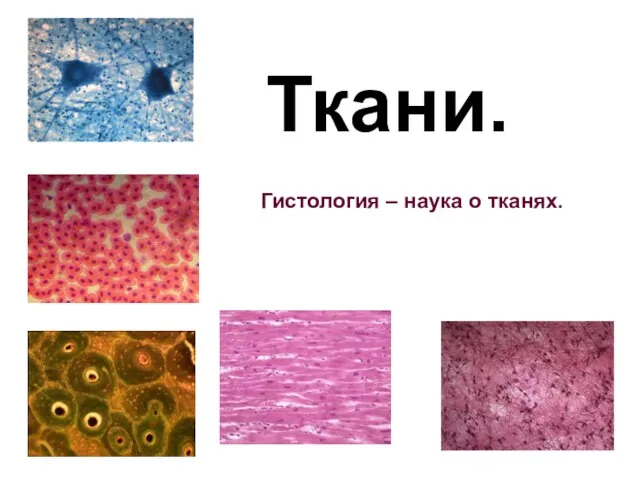 Ткани. Гистология – наука о тканях
Ткани. Гистология – наука о тканях Физиология нервов и нервно-мышечных синапсов
Физиология нервов и нервно-мышечных синапсов Строение корня
Строение корня Презентация на тему "Невероятные факты о животных" - скачать бесплатно презентации по Биологии
Презентация на тему "Невероятные факты о животных" - скачать бесплатно презентации по Биологии Среды жизни на Земле
Среды жизни на Земле Витамины. Ферменты. Прочие вещества продуктов
Витамины. Ферменты. Прочие вещества продуктов Семейство злаковые
Семейство злаковые Л- вирусы
Л- вирусы Гигиена органов пищеварения. Желудочно-кишечные инфекции.
Гигиена органов пищеварения. Желудочно-кишечные инфекции. Жизненный цикл клетки. Деление клетки. Митоз. Мейоз. Амитоз. Эндомитоз
Жизненный цикл клетки. Деление клетки. Митоз. Мейоз. Амитоз. Эндомитоз Насекомые
Насекомые Презентация на тему "Размножение споровых растений" - скачать презентации по Биологии
Презентация на тему "Размножение споровых растений" - скачать презентации по Биологии