- Молекулярные механизмы регуляции поведения. Молекулы и поведение

Содержание
- 2. Измерение поведения Правильный выбор признака обеспечивает успех выяснения его молекулярной организации. Определение признака должно быть конструктивным
- 3. Двигательная активность и время нахождения в областях арены Измерение двигательной активности и времени нахождения животного в
- 4. EthoStudio Установка включает арену, видеокамеру, компьютер и клавиатуру. Изображение арены захватывается видеокамерой с частотой 10-25 к/с,
- 5. Тест открытое поле Лабораторные грызуны (мыши и крысы) имеют врожденный страх перед открытым и ярко освещенным
- 6. Измерение тревожности Тест «приподнятый крестообразный лабиринт» - мыши и крысы избегают открытых и небезопасных рукавов. Тест
- 7. Тест принудительного плавания Предложенный в 1977 г. этот тест является самым распространенным методом оценки антидепрессантной активности
- 8. Омический подход к исследованию молекулярного механизма поведения Основан на логически обоснованной и очевидной идеи, что поведение
- 9. Доктрина полигенной регуляции Предложена в 1949 Kenneth Mather и John Jinks. Постулирует, что 1) сложный количественный
- 11. Классификация мутаций По структуре: По действию: 1. функциональные 2. нейтральные
- 12. Forward genetics: 1. GWAS 2. QTL analysis Reversed genetics: 1. Null mutations 2. Point mutations Стратегии
- 13. Техника GWAS
- 14. Потерянная наследственность (Aulchenko et al., 2009)
- 15. QTL анализ поведения Самый распространенный и мощный метод генетического картирования сложных признаков. Основан на измерении сцепления
- 16. QTL анализ. Двигательная активность Моторная активность лежит в основе любого поведенческого акта. Два вида нарушений моторной
- 17. Гены, определяющие двигательную активность мыши в открытом поле
- 18. Каталепсия Каталепсия – длительная неподвижность с повышенным тонусом гравитационной мускулатуры, способность сохранять приданную неудобную позу длительное
- 19. Галоперидоловая каталепсия Галоперидол – наиболее мощный антагонист D2 рецепторов. Препарат вызывает глубокую каталепсию. Галоперидоловая каталепсия –
- 20. Геномное картирование чувствительности к галоперидолу у мышей
- 21. Интервальное картирование чувствительности к галоперидолу на хромосомах 1 и 9
- 22. QTL analysis каталепсии у мышей (Куликов и др., ДАН, 2003) (Kulikov et al., 2008)
- 23. (Daniel MacArthur, Nature, 2012, 487:427-428) Forward genetics. Заключение 1. Позволяет получить список возможных генов-кандидатов. 2. Не
- 24. Научный подход Основой современного естествознания служит экспериментальный метод. С древних времен и до XVI века в
- 25. Связь между поведением и молекулами Между поведением и молекулами имеется сигнальная связь, сформированная в ходе процесса
- 26. Разнообразие методов и подходов нейрогеномики поведения Фармакологические методы: Изменение концентрации медиаторов (снижение или повышение с помощью
- 27. Фармакологические методы: Уменьшение концентрации медиатора Разрушение окончаний (нейротоксины). Опустошение везикул (моноамины). Разрушение тел нейронов. (не применимо
- 28. Увеличение концентрации медиатора Ингибирование ферментов разрушения. Введение релизеров, усиливающих секрецию медиатора (амфетамин, d-фенфлурамин). Ингибирование обратного захвата
- 29. Фармакология рецепторов Основная информация о роли медиаторов в регуляции поведения получена при введении веществ действующих на
- 30. Введение препаратов Острое (один раз). Повторное (до 5 раз). Хроническое (более 7 дней). Через рот в
- 31. Измерение концентрации медиатора In situ на срезах мозга с помощью химических соединений или моноклональных антител. Используется
- 32. Reversed genetics Запрос на сайте NCBI словосочетания “Animal Model” Ʃ= 600275 Годы Первая трансгенная мышь (1982)
- 33. Получение нокаутных мышей Инактивирует ген. Получение нокаута: Введение в бластоцисты ДНК гена, инактивированного вставкой устойчивости к
- 34. Генетический нокаут: за и против Позволяет исследовать значение отдельного гена. Влияние нокаута на поведение в общем
- 35. Основное противоречие полигенной концепции Один полиген добавляет мало информации о молекулярной структуре поведения. Одновременный анализ всех
- 36. Гипотеза главного гена корректирует недостатки полигенной гипотезы Гипотеза главного гена возникла как компромисс на главное противоречие
- 37. Концепция генов-кандидатов Согласуется с концепцией главных генов и является ее реализацией; постулирует, что наследственные изменения поведения
- 38. Возможные пути от гена к поведению
- 39. Гены кандидаты, ассоциированные с биполярными психозами БП часто связывают с дисфункцией 5-HT системы мозга
- 40. Гены кандидаты в психиатрической генетики Главный подход в психиатрической генетике включает изучение ассоциаций между мутациями генов-кандидатов
- 41. Прогресс, который приносит концепция генов-кандидатов в психиатрию Распространение идей и методов молекулярной биологии в психиатрию, нейробиологию
- 42. Первая проблема: выбор гена-кандидата из тысяч потенциальных генов Около 10 000 генов экспрессируются в мозге и
- 43. Вторая проблема: выбор одного полиморфизма из большого числа возможных Результат работы часто зависит от выбора полиморфизма.
- 44. Протокол исследования генов-кандидатов Идеальный протокол включает: 1. Выбор системы, вовлеченной в регуляцию исследуемого поведения. 2. Выбор
- 45. 5-HT система мозге SERT and MAOA are targets for antidepressant drugs
- 46. Функциональные полиморфизмы в генах, регулирующих 5-HT систему человека
- 47. Ассоциация между геном hTPH2 и психопатологиями Мета анализ с использование большого числа маркеров демонстрирует ассоциацию гена
- 48. Мышиные модели функциональных мутаций в генах системы человека
- 49. 1Alenina et al., 2009; 2Angoa-Perez et al., 2012; 3Beaulieu et al., 2008; 4Lesch et al., 2012;
- 51. Действие антидепрессантов на неподвижность мышей Slc6a4+/+, Slc6a4+/- and Slc6a4-/- в TST
- 52. Спонтанная межсамцовая агрессия
- 53. Гены-кандидаты межсамцовой агрессии (Maxson, 1999) Аденозин 2а рецептор Андроген рецептор Кальций/кальмодулин киназа Андроген рецептор Кальций/кальмодулин киназа
- 54. Выбор полиморфизма C1473G Полиморфизм C1473G приводит к замене Pro447Arg в молекуле ТПГ2 (Zhang et al., 2004)
- 55. Распределение полиморфизмов C1473G и С1477T в популяциях диких мышей (Osipova et al., Genes Brain Behav, 2010).
- 56. Ассоциация полиморфизма C1473G с интенсивностью межсамцовой агрессии мышей Самцы 5 линий гомозиготных по C аллелю атаковали
- 57. Доказательство сцепления полиморфизма c1473g с интенсивностью межсамцовой агрессии
- 58. Фармакологическое доказательство участия ТПГ2 в регуляции агрессии у мышей Активация синтеза 5-HT L-триптофаном значительно усиливает интенсивность
- 59. Наследственная каталепсия у крыс ГК В популяции крыс Вистар около 10% животных сохраняют приданную им вертикальную
- 60. Наследственная каталепсия у мышей и крыс сопровождается локальным увеличением активности ТПГ-2 в стриатуме CBA AKR DD
- 61. Ингибиторы ТПГ-2 уменьшают время замирания мышей и крыс МЫШИ CBA КРЫСЫ ГК КОНТРОЛЬ p-CMA *** КОНТРОЛЬ
- 63. Скачать презентацию

Измерение поведения
Правильный выбор признака обеспечивает успех выяснения его молекулярной организации.
Определение признака
Измерение поведения
Правильный выбор признака обеспечивает успех выяснения его молекулярной организации.
Определение признака

Двигательная активность и время нахождения в областях арены
Измерение двигательной активности и
Двигательная активность и время нахождения в областях арены
Измерение двигательной активности и
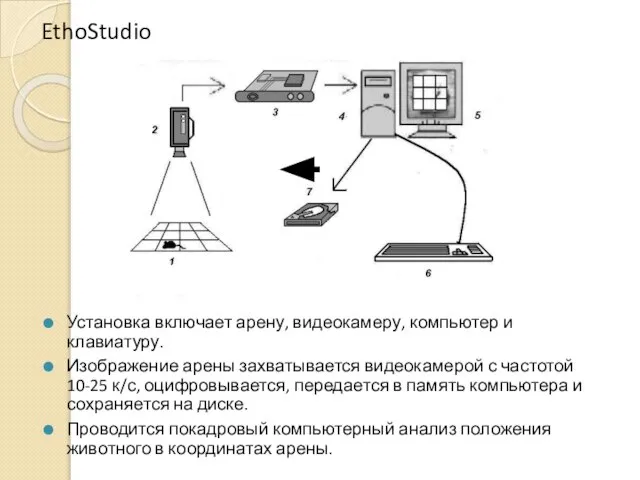
EthoStudio
Установка включает арену, видеокамеру, компьютер и клавиатуру.
Изображение арены захватывается видеокамерой с
EthoStudio
Установка включает арену, видеокамеру, компьютер и клавиатуру.
Изображение арены захватывается видеокамерой с

Тест открытое поле
Лабораторные грызуны (мыши и крысы) имеют врожденный страх перед
Тест открытое поле
Лабораторные грызуны (мыши и крысы) имеют врожденный страх перед

Измерение тревожности
Тест «приподнятый крестообразный лабиринт» - мыши и крысы избегают открытых
Измерение тревожности
Тест «приподнятый крестообразный лабиринт» - мыши и крысы избегают открытых

Тест принудительного плавания
Предложенный в 1977 г. этот тест является самым распространенным
Тест принудительного плавания
Предложенный в 1977 г. этот тест является самым распространенным

Омический подход к исследованию молекулярного механизма поведения
Основан на логически обоснованной и
Омический подход к исследованию молекулярного механизма поведения
Основан на логически обоснованной и

Доктрина полигенной регуляции
Предложена в 1949 Kenneth Mather и John Jinks. Постулирует,
Доктрина полигенной регуляции
Предложена в 1949 Kenneth Mather и John Jinks. Постулирует,

Классификация мутаций
По структуре:
По действию:
1. функциональные
2. нейтральные
Классификация мутаций
По структуре:
По действию:
1. функциональные
2. нейтральные
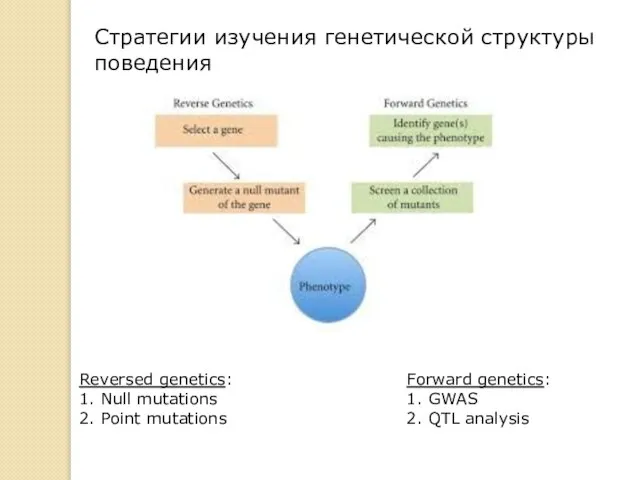
Forward genetics:
1. GWAS
2. QTL analysis
Reversed genetics:
1. Null mutations
2. Point mutations
Стратегии
Forward genetics:
1. GWAS
2. QTL analysis
Reversed genetics:
1. Null mutations
2. Point mutations
Стратегии
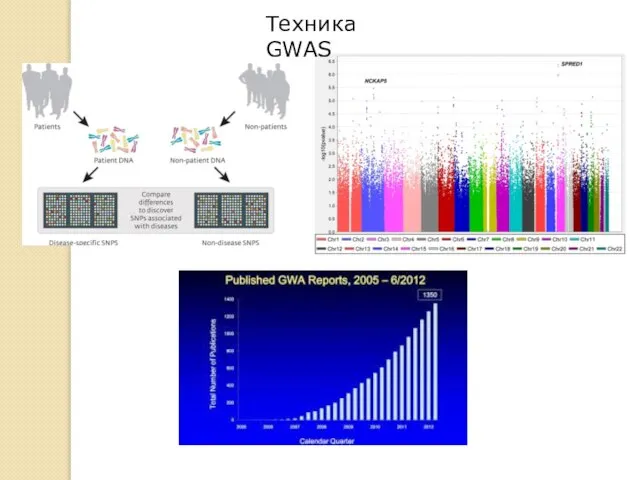
Техника GWAS
Техника GWAS
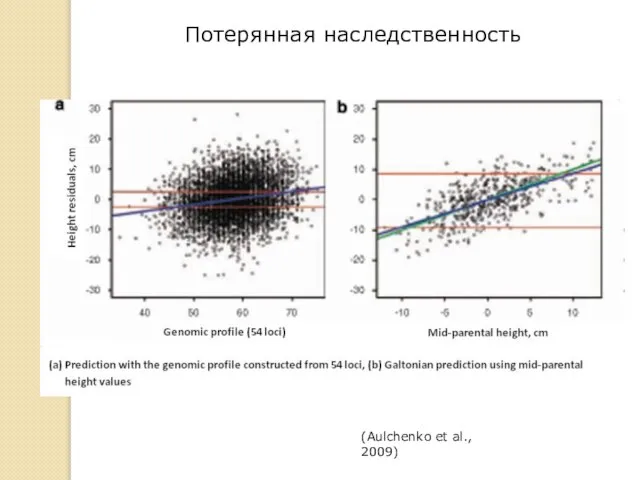
Потерянная наследственность
(Aulchenko et al., 2009)
Потерянная наследственность
(Aulchenko et al., 2009)

QTL анализ поведения
Самый распространенный и мощный метод генетического картирования сложных признаков.
Основан
QTL анализ поведения
Самый распространенный и мощный метод генетического картирования сложных признаков.
Основан

QTL анализ. Двигательная активность
Моторная активность лежит в основе любого поведенческого акта.
Два
QTL анализ. Двигательная активность
Моторная активность лежит в основе любого поведенческого акта.
Два
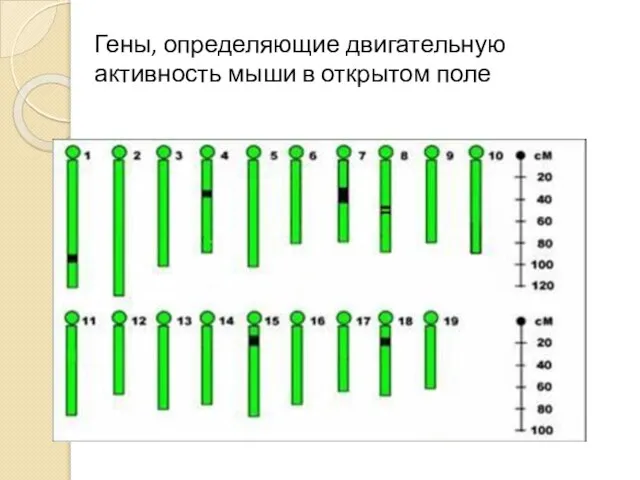
Гены, определяющие двигательную активность мыши в открытом поле
Гены, определяющие двигательную активность мыши в открытом поле
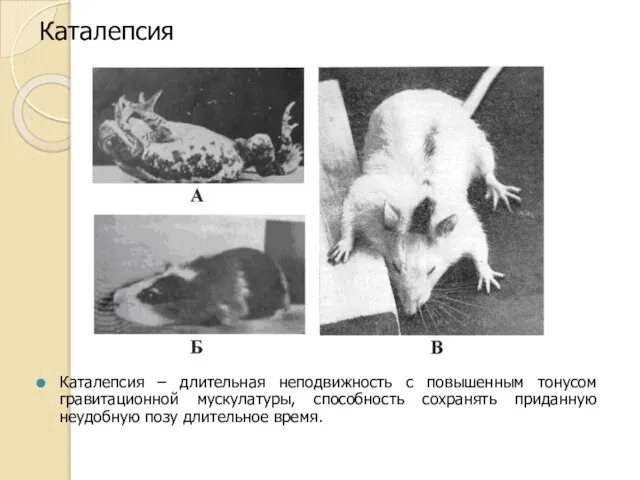
Каталепсия
Каталепсия – длительная неподвижность с повышенным тонусом гравитационной мускулатуры, способность сохранять
Каталепсия
Каталепсия – длительная неподвижность с повышенным тонусом гравитационной мускулатуры, способность сохранять

Галоперидоловая каталепсия
Галоперидол – наиболее мощный антагонист D2 рецепторов.
Препарат вызывает глубокую каталепсию.
Галоперидоловая
Галоперидоловая каталепсия
Галоперидол – наиболее мощный антагонист D2 рецепторов.
Препарат вызывает глубокую каталепсию.
Галоперидоловая
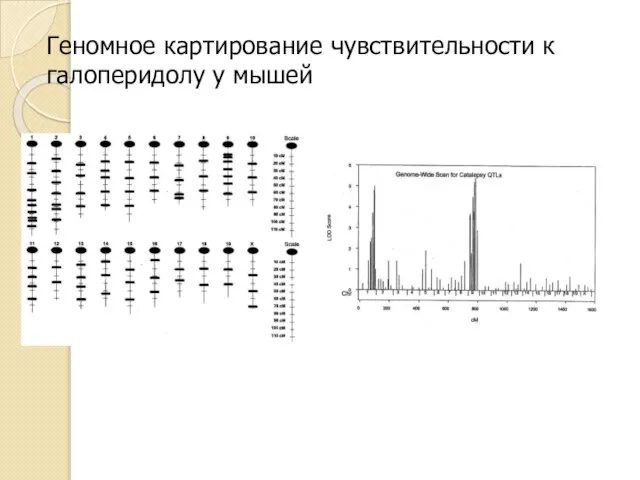
Геномное картирование чувствительности к галоперидолу у мышей
Геномное картирование чувствительности к галоперидолу у мышей
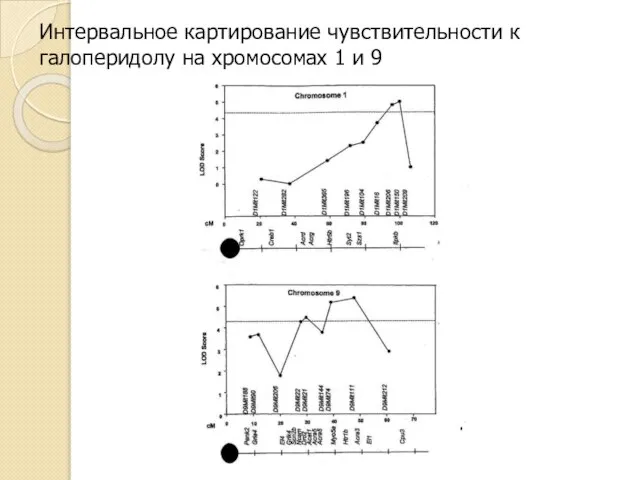
Интервальное картирование чувствительности к галоперидолу на хромосомах 1 и 9
Интервальное картирование чувствительности к галоперидолу на хромосомах 1 и 9
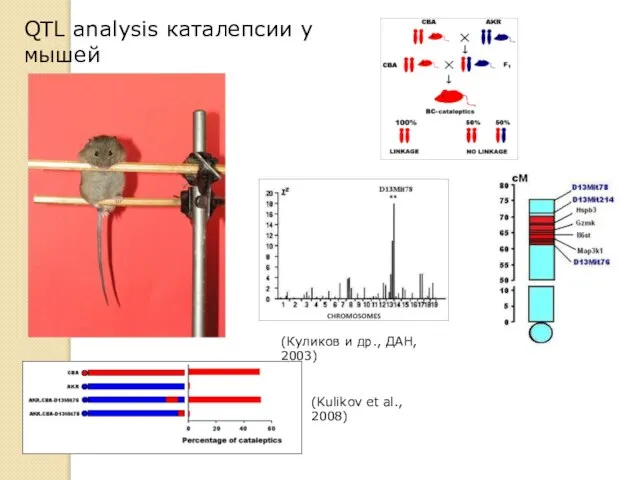
QTL analysis каталепсии у мышей
(Куликов и др., ДАН, 2003)
(Kulikov et al.,
QTL analysis каталепсии у мышей
(Куликов и др., ДАН, 2003)
(Kulikov et al.,

(Daniel MacArthur, Nature, 2012, 487:427-428)
Forward genetics. Заключение
1. Позволяет получить список возможных
(Daniel MacArthur, Nature, 2012, 487:427-428)
Forward genetics. Заключение
1. Позволяет получить список возможных

Научный подход
Основой современного естествознания служит экспериментальный метод.
С древних времен и до
Научный подход
Основой современного естествознания служит экспериментальный метод.
С древних времен и до

Связь между поведением и молекулами
Между поведением и молекулами имеется сигнальная связь,
Связь между поведением и молекулами
Между поведением и молекулами имеется сигнальная связь,

Разнообразие методов и подходов нейрогеномики поведения
Фармакологические методы:
Изменение концентрации медиаторов (снижение или
Разнообразие методов и подходов нейрогеномики поведения
Фармакологические методы:
Изменение концентрации медиаторов (снижение или
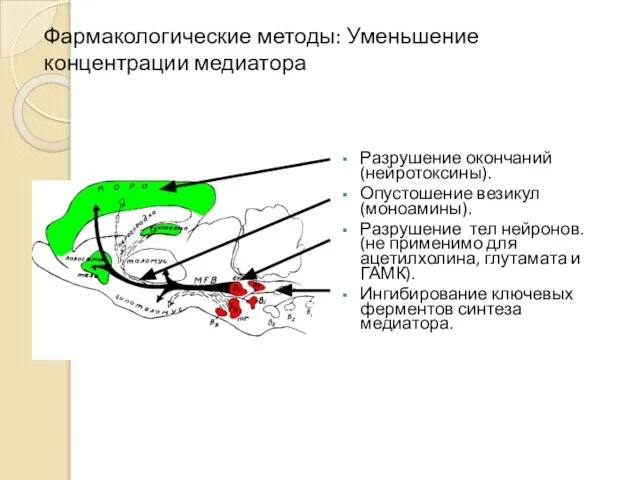
Фармакологические методы: Уменьшение концентрации медиатора
Разрушение окончаний (нейротоксины).
Опустошение везикул (моноамины).
Разрушение тел нейронов.
Фармакологические методы: Уменьшение концентрации медиатора
Разрушение окончаний (нейротоксины).
Опустошение везикул (моноамины).
Разрушение тел нейронов.
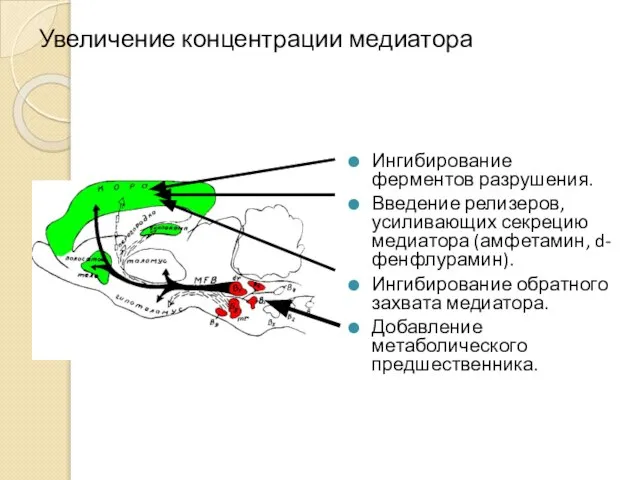
Увеличение концентрации медиатора
Ингибирование ферментов разрушения.
Введение релизеров, усиливающих секрецию медиатора (амфетамин, d-фенфлурамин).
Ингибирование
Увеличение концентрации медиатора
Ингибирование ферментов разрушения.
Введение релизеров, усиливающих секрецию медиатора (амфетамин, d-фенфлурамин).
Ингибирование

Фармакология рецепторов
Основная информация о роли медиаторов в регуляции поведения получена при
Фармакология рецепторов
Основная информация о роли медиаторов в регуляции поведения получена при

Введение препаратов
Острое (один раз).
Повторное (до 5 раз).
Хроническое (более 7 дней).
Через рот
Введение препаратов
Острое (один раз).
Повторное (до 5 раз).
Хроническое (более 7 дней).
Через рот

Измерение концентрации медиатора
In situ на срезах мозга с помощью химических соединений
Измерение концентрации медиатора
In situ на срезах мозга с помощью химических соединений
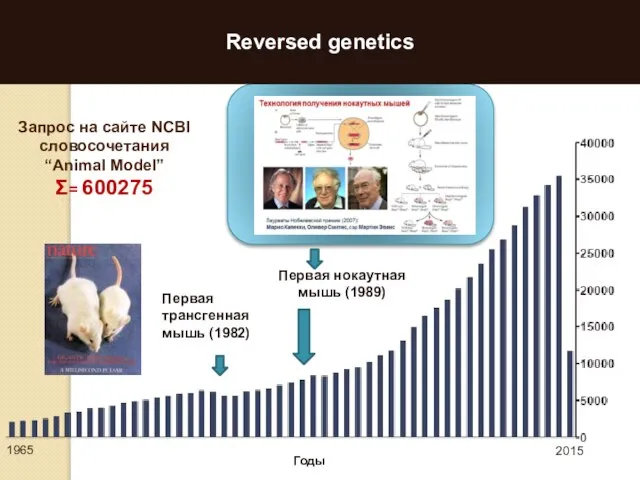
Reversed genetics
Запрос на сайте NCBI словосочетания “Animal Model”
Ʃ= 600275
Годы
Первая трансгенная
Запрос на сайте NCBI словосочетания “Animal Model”
Ʃ= 600275
Годы
Первая трансгенная
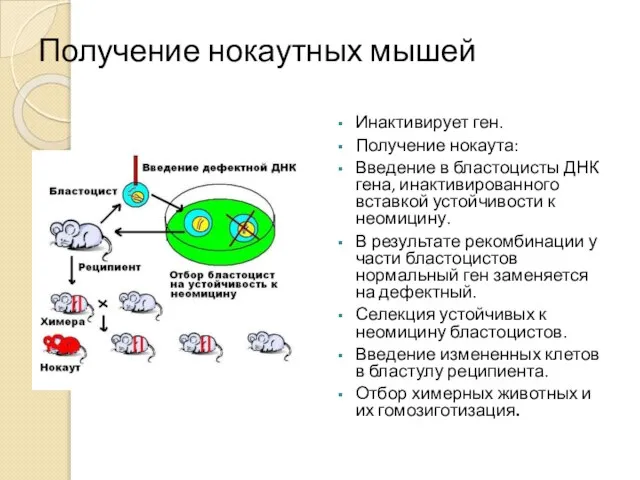
Получение нокаутных мышей
Инактивирует ген.
Получение нокаута:
Введение в бластоцисты ДНК гена, инактивированного вставкой
Получение нокаутных мышей
Инактивирует ген.
Получение нокаута:
Введение в бластоцисты ДНК гена, инактивированного вставкой

Генетический нокаут: за и против
Позволяет исследовать значение отдельного гена.
Влияние нокаута на
Генетический нокаут: за и против
Позволяет исследовать значение отдельного гена.
Влияние нокаута на

Основное противоречие полигенной концепции
Один полиген добавляет мало информации о молекулярной структуре
Основное противоречие полигенной концепции
Один полиген добавляет мало информации о молекулярной структуре

Гипотеза главного гена корректирует недостатки полигенной гипотезы
Гипотеза главного гена возникла как
Гипотеза главного гена корректирует недостатки полигенной гипотезы
Гипотеза главного гена возникла как
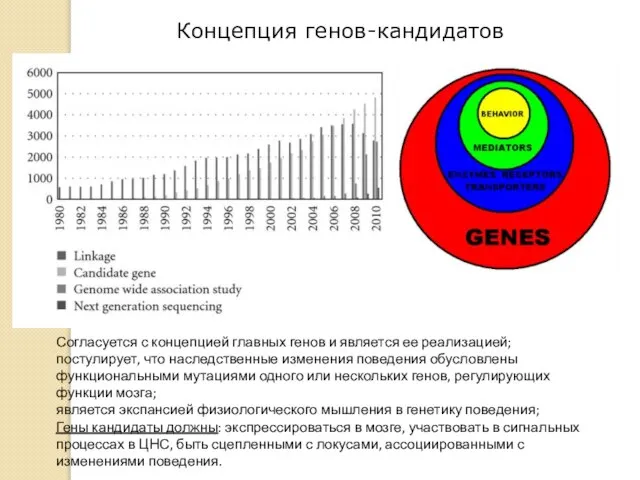
Концепция генов-кандидатов
Согласуется с концепцией главных генов и является ее реализацией;
постулирует, что
Концепция генов-кандидатов
Согласуется с концепцией главных генов и является ее реализацией;
постулирует, что
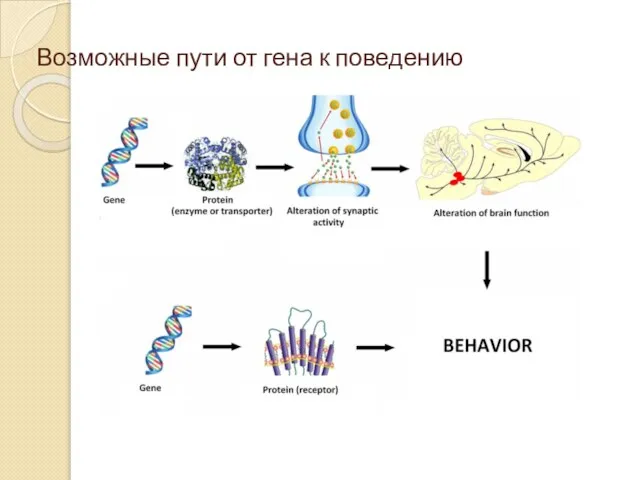
Возможные пути от гена к поведению
Возможные пути от гена к поведению

Гены кандидаты, ассоциированные с
биполярными психозами
БП часто связывают с дисфункцией
5-HT
Гены кандидаты, ассоциированные с
биполярными психозами
БП часто связывают с дисфункцией
5-HT

Гены кандидаты в психиатрической генетики
Главный подход в психиатрической генетике включает изучение
Гены кандидаты в психиатрической генетики
Главный подход в психиатрической генетике включает изучение

Прогресс, который приносит концепция генов-кандидатов в психиатрию
Распространение идей и методов молекулярной
Прогресс, который приносит концепция генов-кандидатов в психиатрию
Распространение идей и методов молекулярной
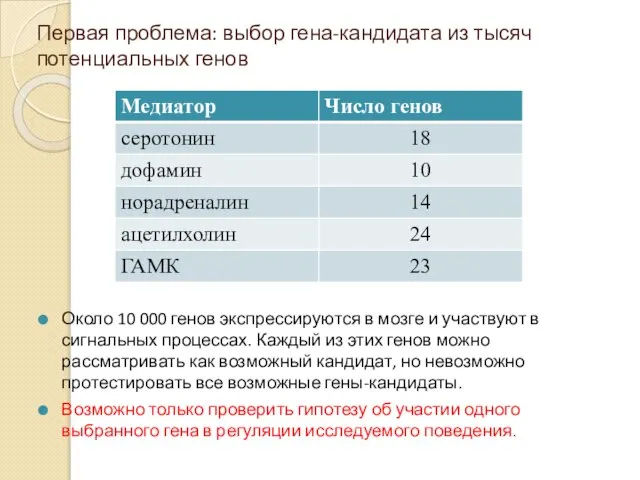
Первая проблема: выбор гена-кандидата из тысяч потенциальных генов
Около 10 000 генов
Первая проблема: выбор гена-кандидата из тысяч потенциальных генов
Около 10 000 генов
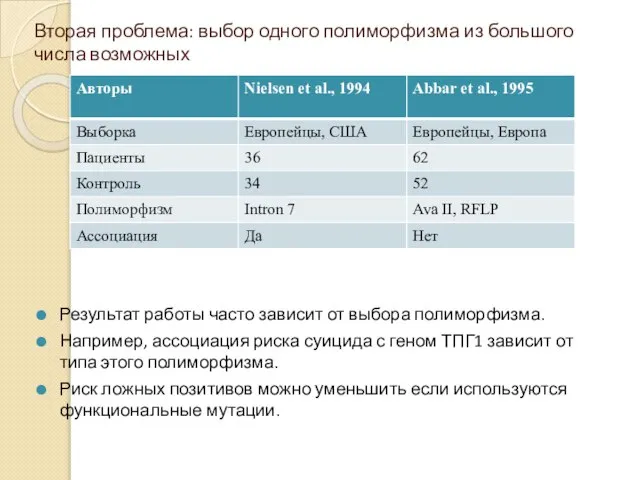
Вторая проблема: выбор одного полиморфизма из большого числа возможных
Результат работы часто
Вторая проблема: выбор одного полиморфизма из большого числа возможных
Результат работы часто

Протокол исследования генов-кандидатов
Идеальный протокол включает:
1. Выбор системы, вовлеченной в регуляцию исследуемого
Протокол исследования генов-кандидатов
Идеальный протокол включает:
1. Выбор системы, вовлеченной в регуляцию исследуемого
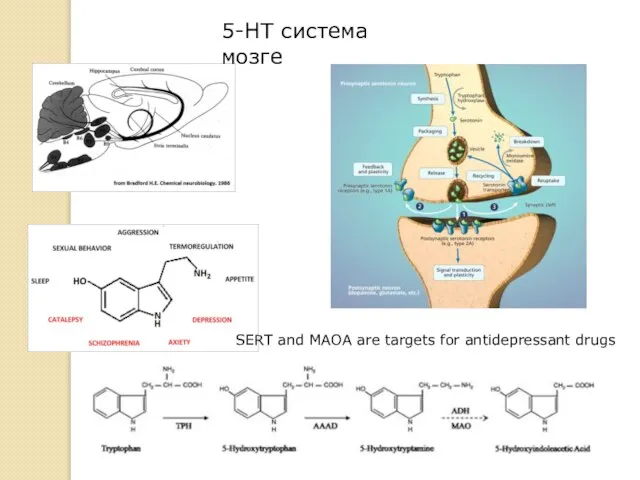
5-HT система мозге
SERT and MAOA are targets for antidepressant drugs
5-HT система мозге
SERT and MAOA are targets for antidepressant drugs
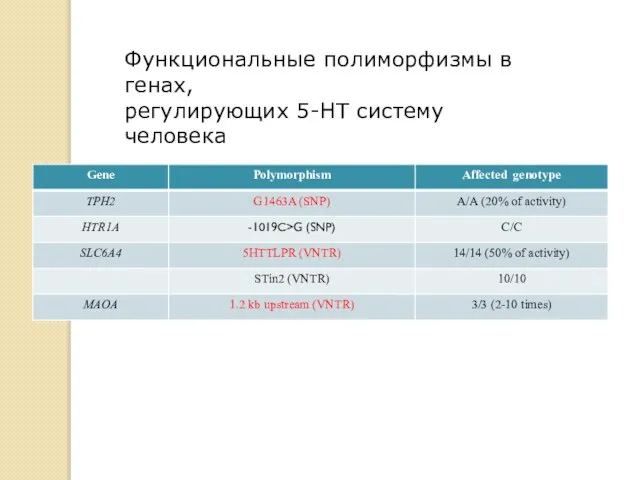
Функциональные полиморфизмы в генах,
регулирующих 5-HT систему человека
Функциональные полиморфизмы в генах,
регулирующих 5-HT систему человека
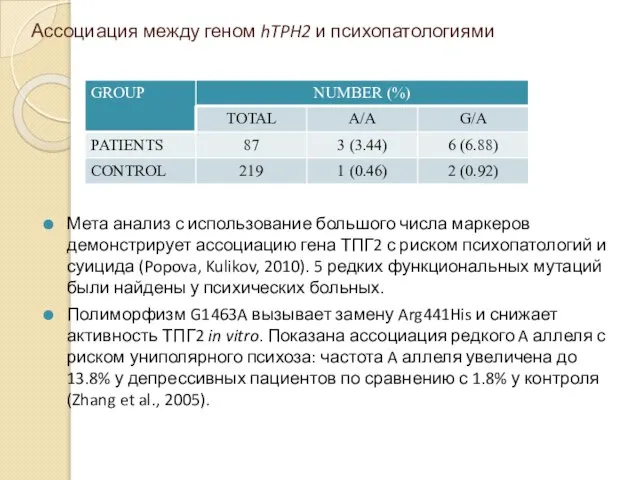
Ассоциация между геном hTPH2 и психопатологиями
Мета анализ с использование большого числа
Ассоциация между геном hTPH2 и психопатологиями
Мета анализ с использование большого числа

Мышиные модели функциональных мутаций в генах системы человека
Мышиные модели функциональных мутаций в генах системы человека

1Alenina et al., 2009; 2Angoa-Perez et al., 2012; 3Beaulieu et al.,
1Alenina et al., 2009; 2Angoa-Perez et al., 2012; 3Beaulieu et al.,
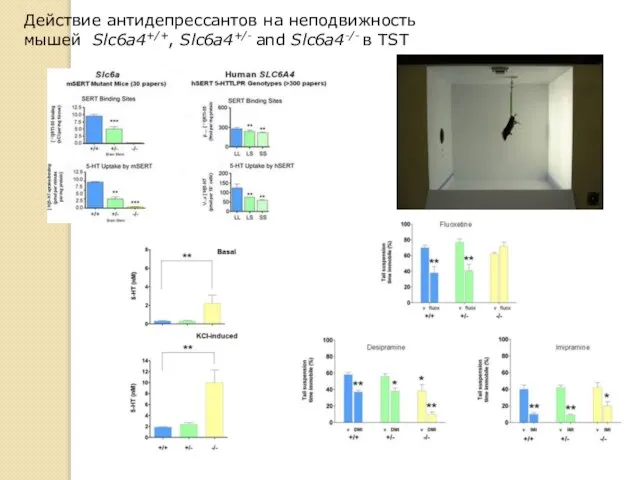
Действие антидепрессантов на неподвижность
мышей Slc6a4+/+, Slc6a4+/- and Slc6a4-/- в TST
Действие антидепрессантов на неподвижность
мышей Slc6a4+/+, Slc6a4+/- and Slc6a4-/- в TST

Спонтанная межсамцовая агрессия
Спонтанная межсамцовая агрессия

Гены-кандидаты межсамцовой агрессии
(Maxson, 1999)
Аденозин 2а рецептор
Андроген рецептор
Кальций/кальмодулин киназа
Андроген рецептор
Кальций/кальмодулин киназа II
Энкефалин
Рецептор
Гены-кандидаты межсамцовой агрессии
(Maxson, 1999)
Аденозин 2а рецептор
Андроген рецептор
Кальций/кальмодулин киназа
Андроген рецептор
Кальций/кальмодулин киназа II
Энкефалин
Рецептор
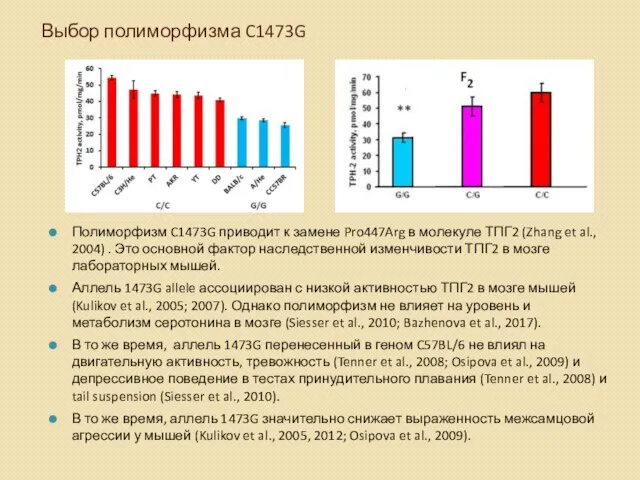
Выбор полиморфизма C1473G
Полиморфизм C1473G приводит к замене Pro447Arg в молекуле ТПГ2
Выбор полиморфизма C1473G
Полиморфизм C1473G приводит к замене Pro447Arg в молекуле ТПГ2
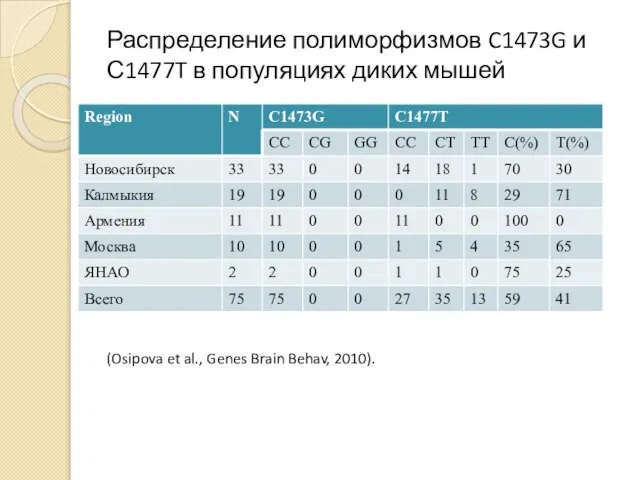
Распределение полиморфизмов C1473G и С1477T в популяциях диких мышей
(Osipova et al.,
Распределение полиморфизмов C1473G и С1477T в популяциях диких мышей
(Osipova et al.,
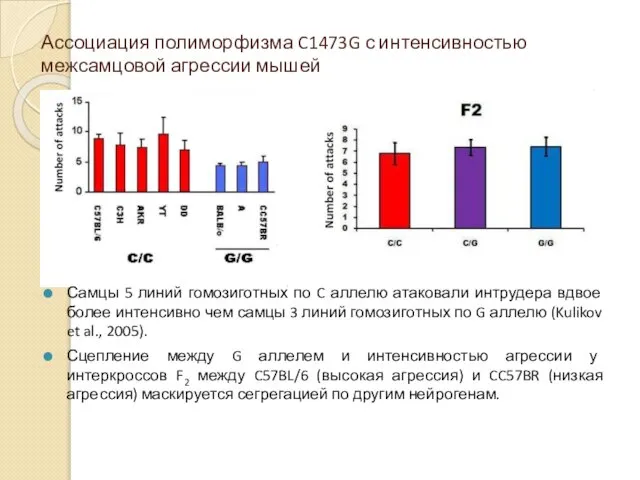
Ассоциация полиморфизма C1473G с интенсивностью межсамцовой агрессии мышей
Самцы 5 линий гомозиготных
Ассоциация полиморфизма C1473G с интенсивностью межсамцовой агрессии мышей
Самцы 5 линий гомозиготных
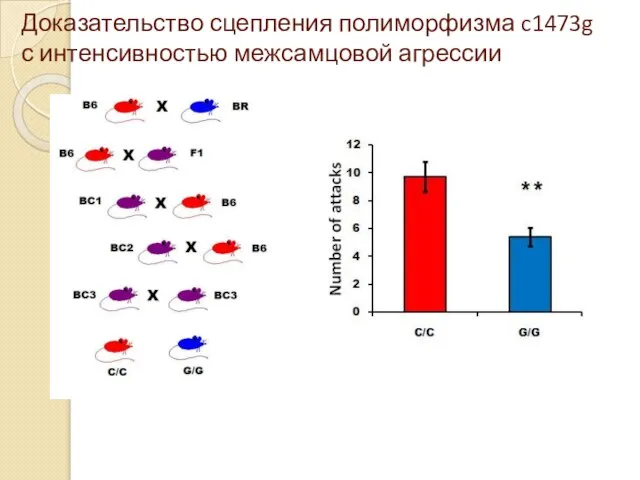
Доказательство сцепления полиморфизма c1473g с интенсивностью межсамцовой агрессии
Доказательство сцепления полиморфизма c1473g с интенсивностью межсамцовой агрессии
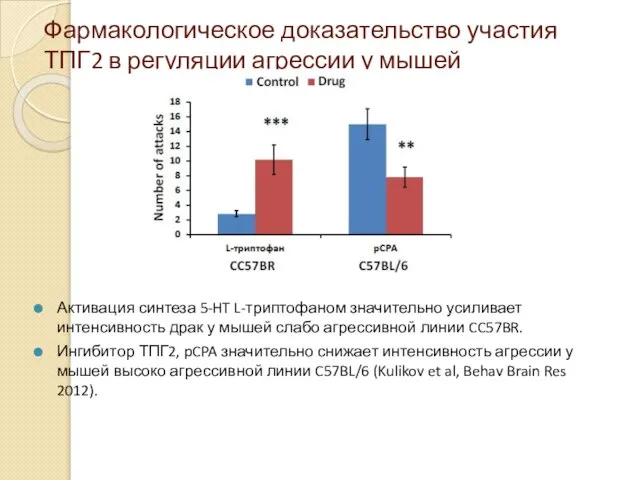
Фармакологическое доказательство участия ТПГ2 в регуляции агрессии у мышей
Активация синтеза 5-HT
Фармакологическое доказательство участия ТПГ2 в регуляции агрессии у мышей
Активация синтеза 5-HT

Наследственная каталепсия у крыс ГК
В популяции крыс Вистар около 10% животных
Наследственная каталепсия у крыс ГК
В популяции крыс Вистар около 10% животных
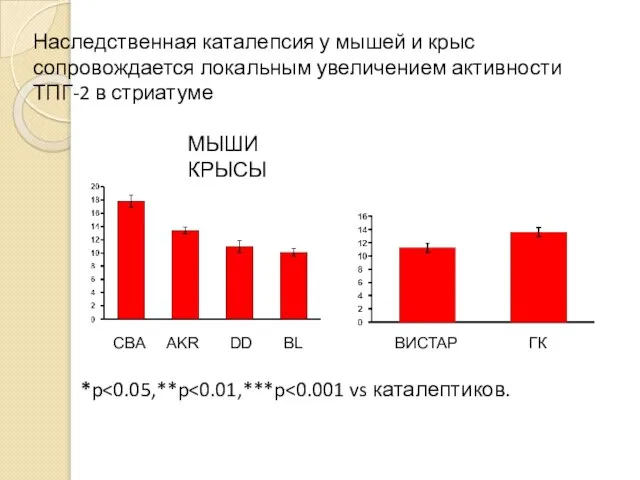
Наследственная каталепсия у мышей и крыс сопровождается локальным увеличением активности ТПГ-2
Наследственная каталепсия у мышей и крыс сопровождается локальным увеличением активности ТПГ-2
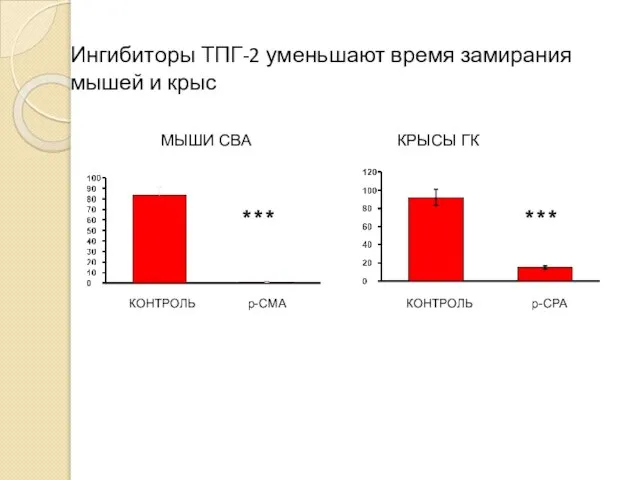
Ингибиторы ТПГ-2 уменьшают время замирания мышей и крыс
МЫШИ CBA КРЫСЫ
Ингибиторы ТПГ-2 уменьшают время замирания мышей и крыс
МЫШИ CBA КРЫСЫ
 Селекция как наука
Селекция как наука Ученые-ботаники, исследователи Донского и Азово-Черноморского региона
Ученые-ботаники, исследователи Донского и Азово-Черноморского региона Жизнь пресного водоема
Жизнь пресного водоема Кровь и кровообращение
Кровь и кровообращение Презентация на тему "Грибы" - скачать презентации по Биологии
Презентация на тему "Грибы" - скачать презентации по Биологии Презентация на тему "Удивительные факты из жизни животных. Рыбы" - скачать презентации по Биологии
Презентация на тему "Удивительные факты из жизни животных. Рыбы" - скачать презентации по Биологии Раздельное питание
Раздельное питание Сочинение-рассуждение «Береги и охраняй родную природу» Природа всё учла и взвесила - Не нарушайте равновесия! (В.Шефнер)
Сочинение-рассуждение «Береги и охраняй родную природу» Природа всё учла и взвесила - Не нарушайте равновесия! (В.Шефнер)  Закономерность эволюции живых организмов
Закономерность эволюции живых организмов Питание растений
Питание растений Презентация Царство растений
Презентация Царство растений Синтетическая теория эволюции
Синтетическая теория эволюции Перелетные птицы. Скворец
Перелетные птицы. Скворец Презентация на тему "Спинной мозг" - скачать бесплатно презентации по Биологии
Презентация на тему "Спинной мозг" - скачать бесплатно презентации по Биологии Систематика эукариотов и прокариотов. Морфология микроорганизмов (лекции №1)
Систематика эукариотов и прокариотов. Морфология микроорганизмов (лекции №1) Тема 12.Наваз, воздух, вода
Тема 12.Наваз, воздух, вода Лишайники: строение и практическое значение Биология 7 класс «Выступает ли из волн океана водный утес, оторвется ли обломок ск
Лишайники: строение и практическое значение Биология 7 класс «Выступает ли из волн океана водный утес, оторвется ли обломок ск Рациональное планирование режима дня в соответствии с хронотипами школьников
Рациональное планирование режима дня в соответствии с хронотипами школьников Высота и долговечность древесных растений
Высота и долговечность древесных растений Матриксные металлопротеиназы
Матриксные металлопротеиназы Эндокринная система
Эндокринная система Андре́й Андре́евич Громы́ко
Андре́й Андре́евич Громы́ко Ветеринарные препараты в кролиководстве
Ветеринарные препараты в кролиководстве Pet the coo
Pet the coo Подготовка к ЕГЭ по темам Человек
Подготовка к ЕГЭ по темам Человек Гигиена питания Урок составил учитель биологии МОУ СОШ №2 г Искитима НСО Белич Ю.П.
Гигиена питания Урок составил учитель биологии МОУ СОШ №2 г Искитима НСО Белич Ю.П. Развитие психики в онтогенезе. Постнатальный период. Ранний постнатальный период
Развитие психики в онтогенезе. Постнатальный период. Ранний постнатальный период Возникновение жизни на земле
Возникновение жизни на земле