- Мозг, слух и равновесие. Лекция 9

Содержание
- 2. Три составляющие всякой сенсорной системы: Рецепторы (чувствительные клетки или их отростки) Проводящие нервы (спинномозговые и черепные)
- 3. стимул Вторично-чувствующие рецепторы: специализиро- ванные клетки (не нервные). Нерв образован отростками особых проводящих нейронов: слух и
- 4. Стимул, как правило, вызывает открывание белков-каналов для положительно заряженных ионов (Na+, К+) на мембране рецептора Вход
- 5. При передаче сигнала от рецепторов в ЦНС используется топический принцип: каждый рецептор передает сигнал «своей» нервной
- 6. Топическая организация позволяет закодировать «качество» сенсорного сигнала (= место настроенного на этот сигнал рецептора на рецепторной
- 7. Таким образом, каждый сенсорный сигнал характеризуется определенным количеством (=энергия стимула) и качеством. Нос Ладонь Пятка 1
- 8. Общий «орган чувства» – внутреннее ухо. Состоит из улитки (слуховая часть), а также вестибулярных мешочков и
- 9. Вестибулярная часть – полукружные каналы, саккулюс, утрикулюс (круглый и овальный мешочки). Слуховая часть – лагена («озеро»),
- 10. Появляются волосковые рецепторы в органе боковой линии рыб (система подкожных каналов, соединенных с внешней средой); дополнение
- 11. Рецепторы несут чувствительные волоски-цилии. Киноцилия – одиночный бывший жгутик с микротрубочками; только в вестиб. системе. Стереоцилии
- 12. Волосковые рецепторы возбуждаются при изгибе волосков от меньшего к большему. Почему? Длительное время считали, что здесь
- 13. Волосковые рецепторы возбуждаются при изгибе волосков от меньшего к большему. Почему? Длительное время считали, что здесь
- 14. Все оказалось несколько сложнее и интереснее: электронная микроскопия позволила разглядеть белковые нити, соединяющие кончики волосков. Было
- 15. Все оказалось несколько сложнее и интереснее: электронная микроскопия позволила разглядеть белковые нити, соединяющие кончики волосков. Было
- 16. Реакция на линейное ускорение: смещение желеобразной массы за счет инерции при появлении ускорения (либо изменения положения
- 17. Два правых рецептора – активация, два левых – торможение. В целом гребень волосков каждого рецептора настроен
- 18. Сигнал от каждого рецептора топически («поточечно») передается в вестибулярные ядра, нейроны которых врожденно связаны с восприятием
- 19. Нерв от внутреннего уха идет в ромбовидную ямку (продолговатый мозг и мост). Вестибулярные ядра, как эволюционно
- 20. Дальнейшая быстрая передача вестибу-лярной информа-ции (прежде всего, для коррекции движений): в спинной мозг (вестибулярные рефлексы) в
- 21. Таламус: фильтр инф-ии (функция внимания). В коре больших полушарий вестиб. сигналы используются для построения и коррекции
- 22. Укачивание – иррадиация (распространение) возбуждения по центрам ромбовидной ямки.
- 24. Филипп Пети (1949); 1974 – на высоте более 400 м между Близнецами 45 мин, ОСКАР 2009
- 25. Вестибулярная система: «количество стимула» = величина ускорения, «качество стимула» = направление ускорения. Слуховая система: «количество стимула»
- 26. Наружное ухо: «рупор» для сбора колебаний воздуха. Среднее ухо: энергия колебаний воздуха улавливается барабанной перепонкой и
- 27. Membrana tympanica: барабанная перепонка Соотношение S барабанной перепонки к S овального окна = 20 : 1
- 28. 4. Мышца, напрягающая барабанную перепонку (m. tensor tympani) и стременная мышца (m. stapedius) – адаптация к
- 29. Распространение колебаний по каналам улитки: из верхнего канала в нижний + колебания базилярной мембраны. Улитковый проток
- 30. Поперечный разрез улитки. Лестница преддверия (верхний канал), барабанная лестница (нижний канал) и улитковый проток (средний канал);
- 31. Улитка (растянутое изображение): чем дальше от овального окна, тем шире базилярная мембрана. Георг Бекеши: чем ниже
- 32. Улитка: разные части базилярной мембраны отвечают на разные частоты (20-20000 Гц) Высокие частоты Низкие частоты У
- 33. Кортиев орган (organum spirale). Внутренние ВК – 1 ряд (собственно сенсорные). Наружные ВК – 3 ряда
- 34. 1. Ядро. 2. Стереоцилии. 3. Базилярная мембрана. 4, 6. Афферентные (чувствительные) окончания. 5, 7. Эфферентные (двигательные)
- 35. Звук вызывает смещение базилярной мембраны относительно текториальной, изгиб волосков и возбуждение рецепторов. Сигнал от волосковых клеток
- 36. Принцип работы кортиева органа Наружные ВК начина-ют сокращаться и «тянут на себя» текто-риальную мембрану, усиливая колебания
- 37. Улитка – частотно-амплитудный анализатор («на выходе» - спектр звука, тонотопия). Поскольку латентный период развития рецепторного потенциала
- 38. Улитка – частотно-амплитудный анализатор («на выходе» возникает спектр звука). В ЦНС – тонотопические карты. Активность волосковых
- 39. Диапазон слышимости от 20-30 Гц до 10-20 кГц. «Ля» 1-й октавы – 440 Гц Средн. частота
- 40. Слуховые центры головного мозга. 1. Улитковые ядра; ядра верхней оливы. Сравнение сигна-лов от правой и левой
- 41. На основе работы улитковых ядер – способность дельфинов и летучих мышей к эхолокации (анализ отражения звука-щелчка
- 42. Слуховые центры головного мозга. 2. Нижние холмики четверохолмия: новизна (изменение частоты и громкос-ти, движение источника звука).
- 43. Слуховые центры головного мозга. 2. Нижние холмики четверохолмия: новизна (изменение частоты и громкос-ти, движение источника звука).
- 44. Слуховая кора: Первичная – височная доля, по границе боковой борозды. Завершение частотно-амплитудного анализа, наиболее точная тонотопическая
- 45. Ниже расположена вторичная слуховая кора – опознавание звуковых образов как совокупности частот (шумы, «звуки природы» и
- 46. Ниже расположена вторичная слуховая кора – опознавание звуковых образов как совокупности частот (шумы, «звуки природы» и
- 47. Задняя часть височной доли – третичная слуховая кора: узнавание наиболее сложных слуховых образов (музыки, речи). Узнавание
- 48. Задняя часть височной доли – третичная слуховая кора: узнавание наиболее сложных слуховых образов (музыки, речи). Узнавание
- 49. Индивидуальные особенности организации мозга и слуховой системы во многом определяют восприятие музыки («изоляция», «порядок»). Диапазон слышимости
- 50. Грустная музыка Минорные аккорды Веселая музыка мажорные аккорды Mitterschiffthaler et al., 2007
- 51. Слева: представление движения; справа – вращение под музыку. Связь слуховой и двигательной систем.
- 52. Связь слуховой и двигательной систем. ЛОКОМОЦИЯ → ритм похода → ритм барабана → музыка и танец
- 53. Положит. эмоции, связанные с выделением дофамина: удовольствие от удовольствие от восприятия движений, танца нового, от творчества
- 54. http://mglinets.narod.ru/mypubl/mgl8.htm http://mglinets.narod.ru/mypubl/mgl6.htm Molecular evolution of the vertebrate mechanosensory cell and ear BERND FRITZSCH, KIRK W. BEISEL,
- 55. Импланты улитки используют при нарушениях работы волосковых клеток (как правило, генетически обусловленных). Пример: нарушение состава эндолимфы,
- 56. Дали, Кандинский, Шагал, Матисс
- 57. Спасибо за внимание! Через неделю (30-го июня): «Вкус, обоняние и немного осязания», последняя лекция цикла.
- 58. Спектрограммы песни зебровой амадины (около 10 сек) и русской народной песни (около 3 сек).
- 59. Спектрограммы голоса человека. а - гласный звук "а-а" здорового содержит только основную частоту и кратные ей
- 60. На спектрограмме горлового пения хорошо видно, что в спектре звука присутствует всего одна частота с ее
- 62. Скачать презентацию
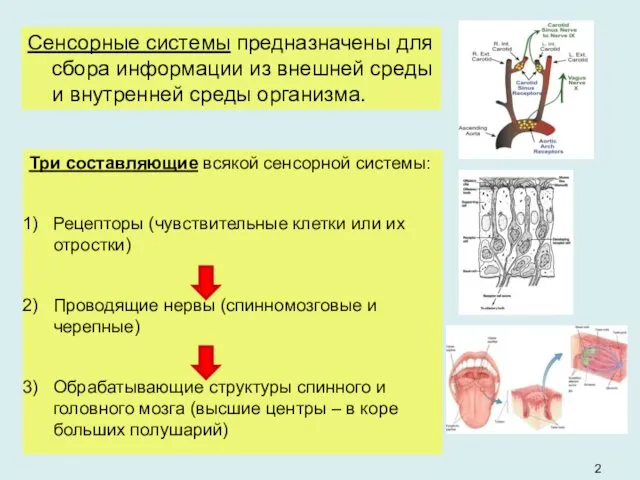
Три составляющие всякой сенсорной системы:
Рецепторы (чувствительные клетки или их отростки)
Проводящие нервы
Три составляющие всякой сенсорной системы:
Рецепторы (чувствительные клетки или их отростки)
Проводящие нервы
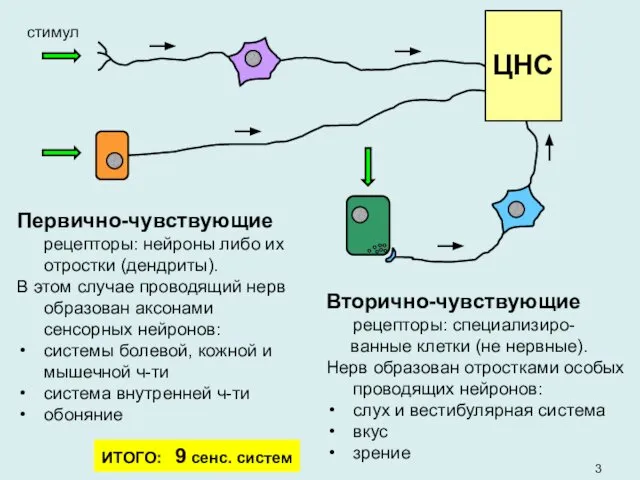
стимул
Вторично-чувствующие рецепторы: специализиро-
ванные клетки (не нервные).
Нерв образован отростками особых
стимул
Вторично-чувствующие рецепторы: специализиро-
ванные клетки (не нервные).
Нерв образован отростками особых
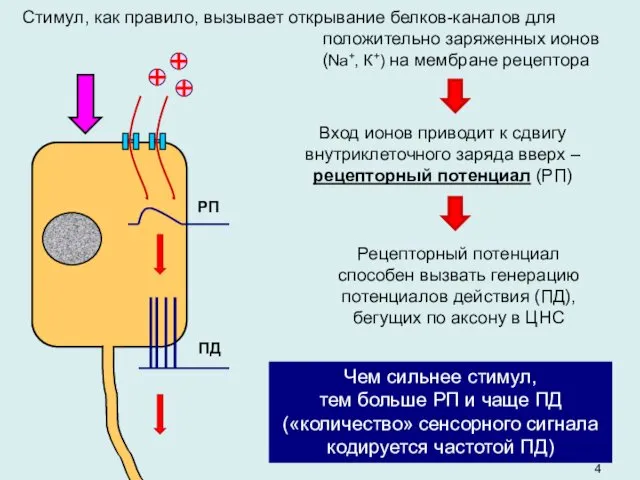
Стимул, как правило, вызывает открывание белков-каналов для положительно заряженных ионов (Na+,
Стимул, как правило, вызывает открывание белков-каналов для положительно заряженных ионов (Na+,
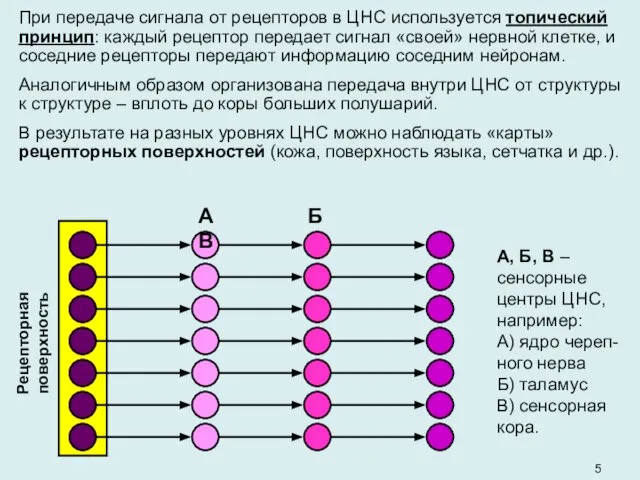
При передаче сигнала от рецепторов в ЦНС используется топический принцип: каждый
При передаче сигнала от рецепторов в ЦНС используется топический принцип: каждый
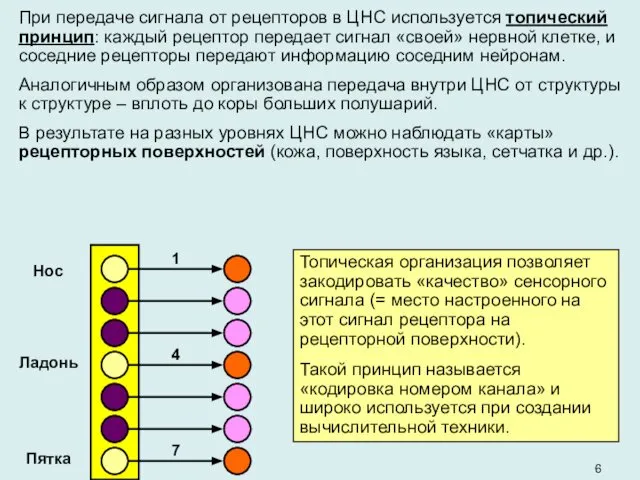
Топическая организация позволяет закодировать «качество» сенсорного сигнала (= место настроенного на
Топическая организация позволяет закодировать «качество» сенсорного сигнала (= место настроенного на
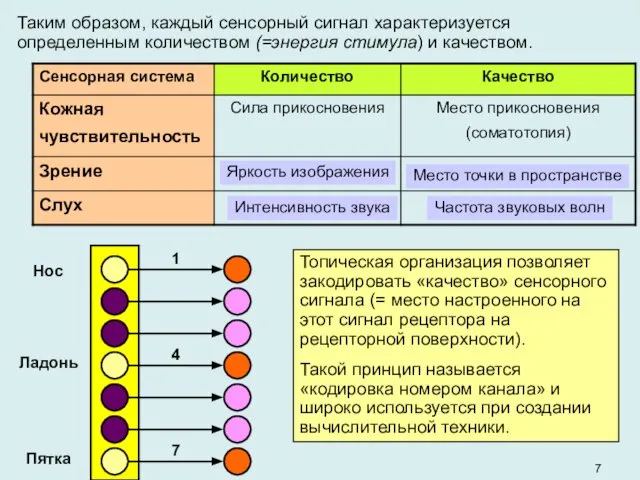
Таким образом, каждый сенсорный сигнал характеризуется определенным количеством (=энергия стимула) и
Таким образом, каждый сенсорный сигнал характеризуется определенным количеством (=энергия стимула) и

Общий «орган чувства» – внутреннее ухо. Состоит из улитки (слуховая часть),
Общий «орган чувства» – внутреннее ухо. Состоит из улитки (слуховая часть),
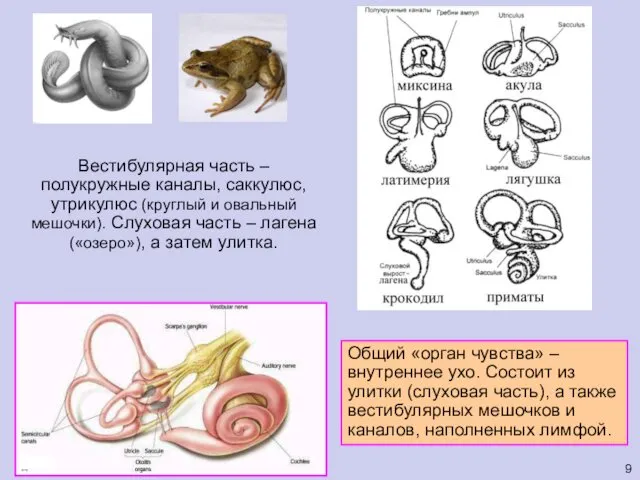
Вестибулярная часть – полукружные каналы, саккулюс, утрикулюс (круглый и овальный мешочки).
Вестибулярная часть – полукружные каналы, саккулюс, утрикулюс (круглый и овальный мешочки).
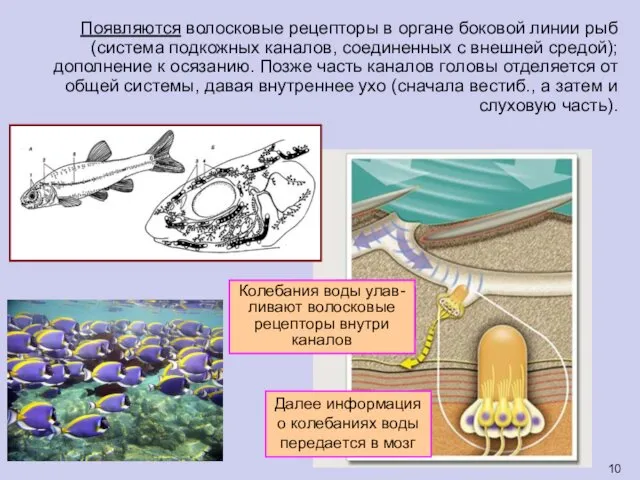
Появляются волосковые рецепторы в органе боковой линии рыб (система подкожных каналов,
Появляются волосковые рецепторы в органе боковой линии рыб (система подкожных каналов,
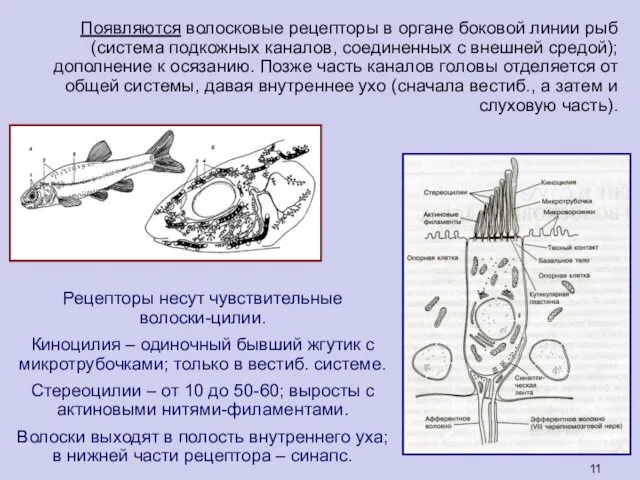
Рецепторы несут чувствительные
волоски-цилии.
Киноцилия – одиночный бывший жгутик с микротрубочками; только
Рецепторы несут чувствительные
волоски-цилии.
Киноцилия – одиночный бывший жгутик с микротрубочками; только
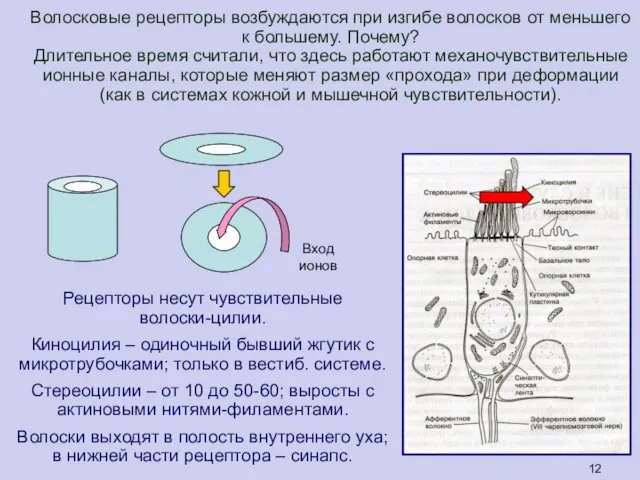
Волосковые рецепторы возбуждаются при изгибе волосков от меньшего к большему. Почему?
Длительное
Волосковые рецепторы возбуждаются при изгибе волосков от меньшего к большему. Почему?
Длительное
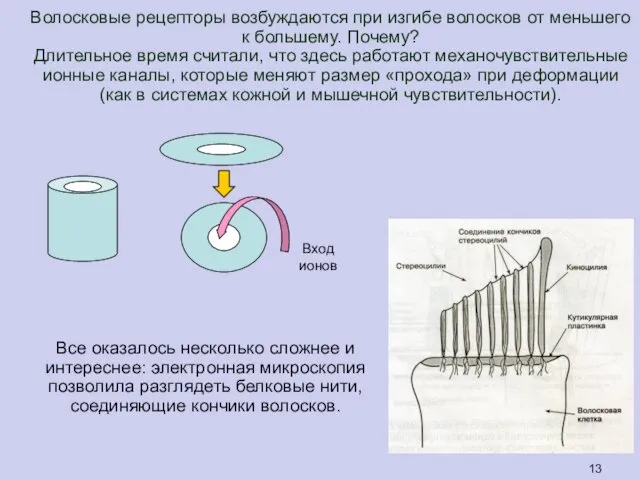
Волосковые рецепторы возбуждаются при изгибе волосков от меньшего к большему. Почему?
Длительное
Волосковые рецепторы возбуждаются при изгибе волосков от меньшего к большему. Почему?
Длительное
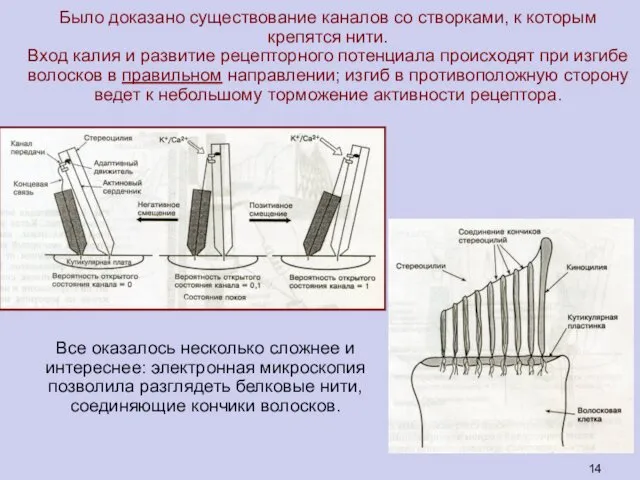
Все оказалось несколько сложнее и
интереснее: электронная микроскопия позволила разглядеть белковые
Все оказалось несколько сложнее и
интереснее: электронная микроскопия позволила разглядеть белковые
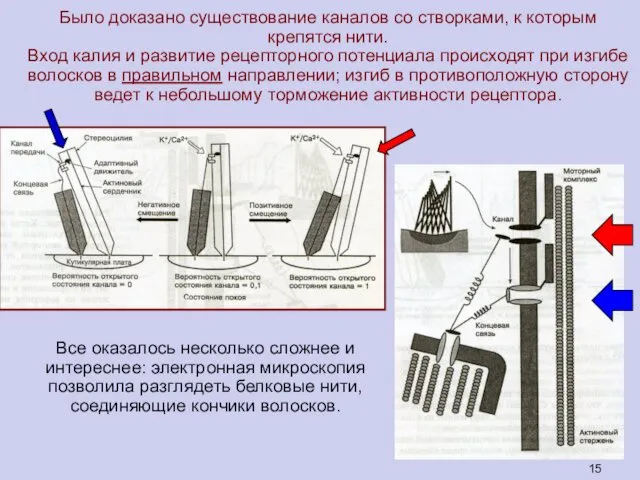
Все оказалось несколько сложнее и
интереснее: электронная микроскопия позволила разглядеть белковые
Все оказалось несколько сложнее и
интереснее: электронная микроскопия позволила разглядеть белковые
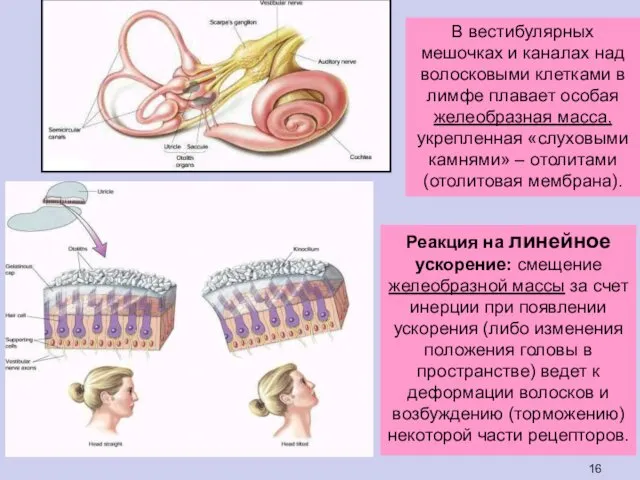
Реакция на линейное ускорение: смещение желеобразной массы за счет инерции при
Реакция на линейное ускорение: смещение желеобразной массы за счет инерции при
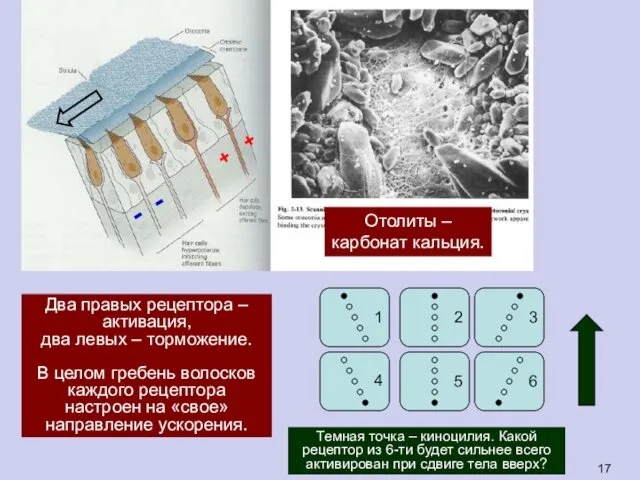
Два правых рецептора –
активация,
два левых – торможение.
В целом гребень волосков
Два правых рецептора –
активация,
два левых – торможение.
В целом гребень волосков

Сигнал от каждого рецептора топически («поточечно») передается в вестибулярные ядра, нейроны
Сигнал от каждого рецептора топически («поточечно») передается в вестибулярные ядра, нейроны
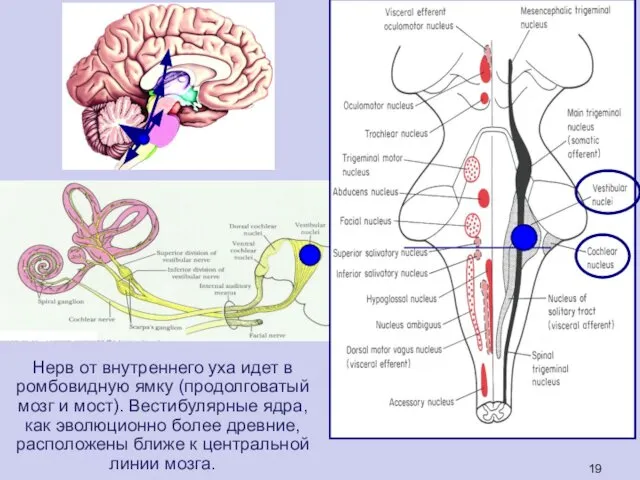
Нерв от внутреннего уха идет в ромбовидную ямку (продолговатый мозг и
Нерв от внутреннего уха идет в ромбовидную ямку (продолговатый мозг и
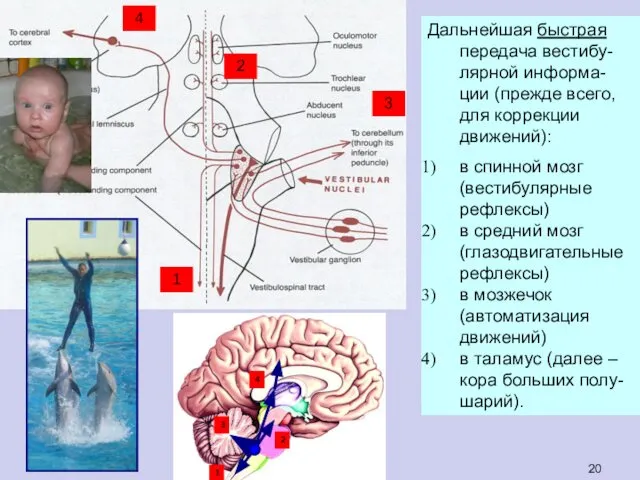
Дальнейшая быстрая передача вестибу-лярной информа-ции (прежде всего, для коррекции движений):
в спинной
Дальнейшая быстрая передача вестибу-лярной информа-ции (прежде всего, для коррекции движений):
в спинной
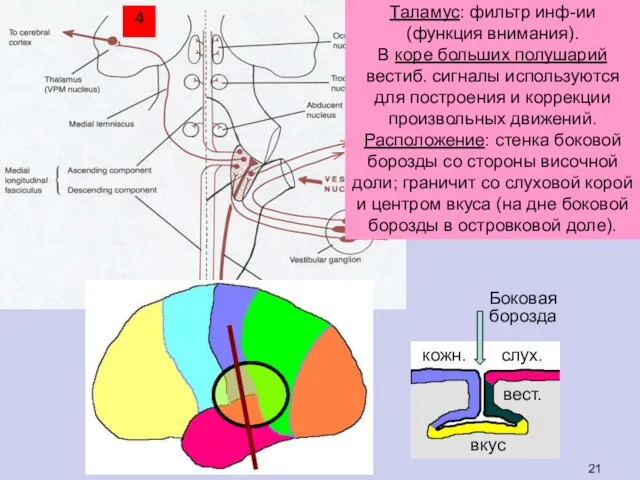
Таламус: фильтр инф-ии (функция внимания).
В коре больших полушарий вестиб. сигналы используются
Таламус: фильтр инф-ии (функция внимания).
В коре больших полушарий вестиб. сигналы используются
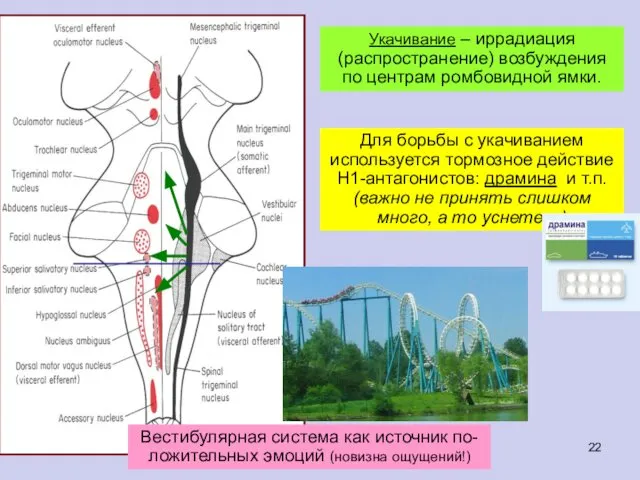
Укачивание – иррадиация (распространение) возбуждения по центрам ромбовидной ямки.
Укачивание – иррадиация (распространение) возбуждения по центрам ромбовидной ямки.


Филипп Пети (1949); 1974 – на высоте более 400 м между
Филипп Пети (1949); 1974 – на высоте более 400 м между
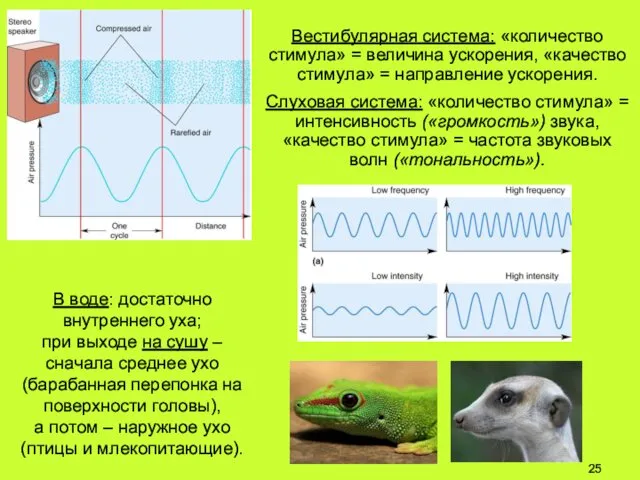
Вестибулярная система: «количество стимула» = величина ускорения, «качество стимула» = направление
Вестибулярная система: «количество стимула» = величина ускорения, «качество стимула» = направление
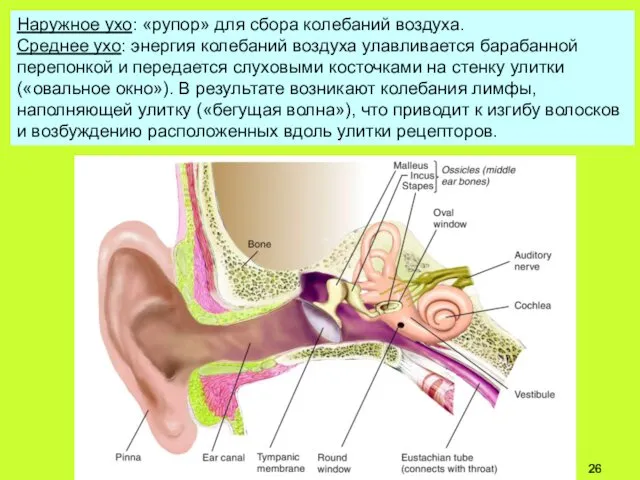
Наружное ухо: «рупор» для сбора колебаний воздуха.
Среднее ухо: энергия колебаний воздуха
Наружное ухо: «рупор» для сбора колебаний воздуха.
Среднее ухо: энергия колебаний воздуха

Membrana tympanica: барабанная перепонка
Соотношение S барабанной перепонки к
S овального окна
Membrana tympanica: барабанная перепонка
Соотношение S барабанной перепонки к
S овального окна
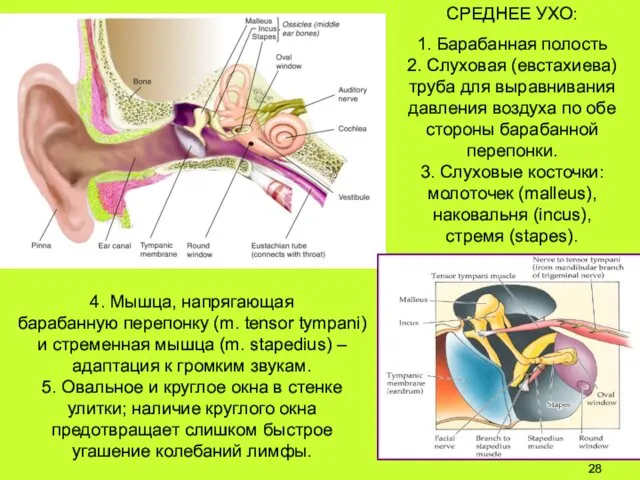
4. Мышца, напрягающая
барабанную перепонку (m. tensor tympani)
и стременная мышца
4. Мышца, напрягающая
барабанную перепонку (m. tensor tympani)
и стременная мышца
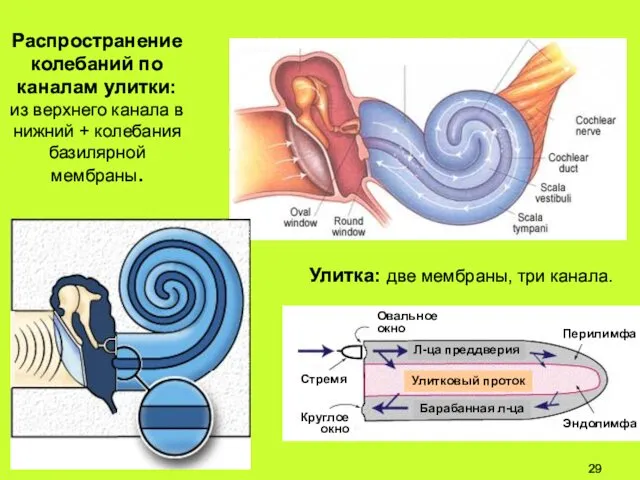
Распространение
колебаний по
каналам улитки:
из верхнего канала в нижний + колебания
Распространение
колебаний по
каналам улитки:
из верхнего канала в нижний + колебания

Поперечный разрез улитки.
Лестница преддверия (верхний канал), барабанная лестница (нижний канал) и
Поперечный разрез улитки.
Лестница преддверия (верхний канал), барабанная лестница (нижний канал) и
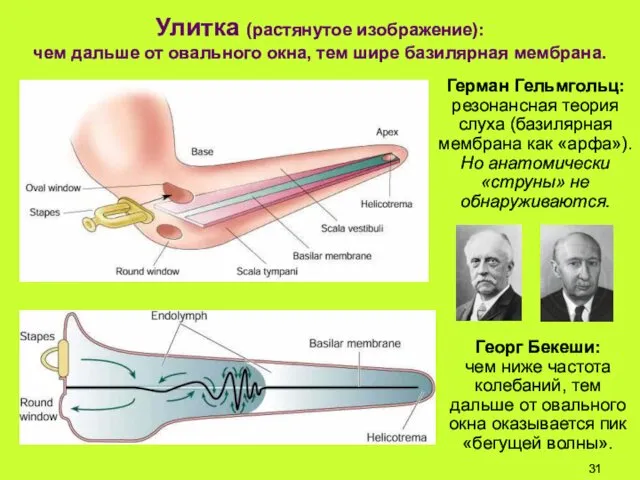
Улитка (растянутое изображение):
чем дальше от овального окна, тем шире базилярная мембрана.
Георг
Улитка (растянутое изображение):
чем дальше от овального окна, тем шире базилярная мембрана.
Георг
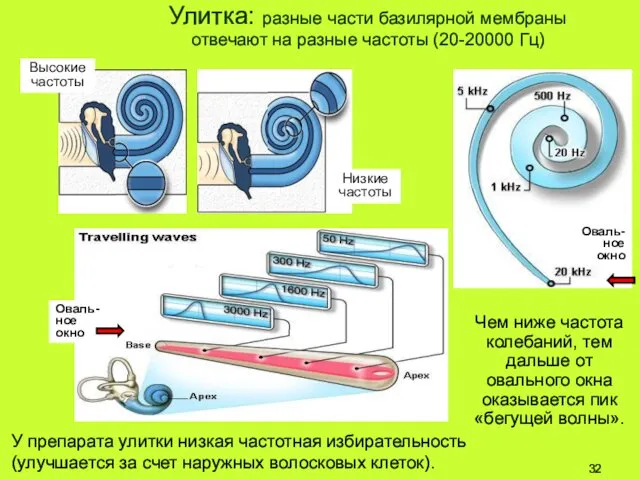
Улитка: разные части базилярной мембраны
отвечают на разные частоты (20-20000 Гц)
Высокие
Улитка: разные части базилярной мембраны
отвечают на разные частоты (20-20000 Гц)
Высокие
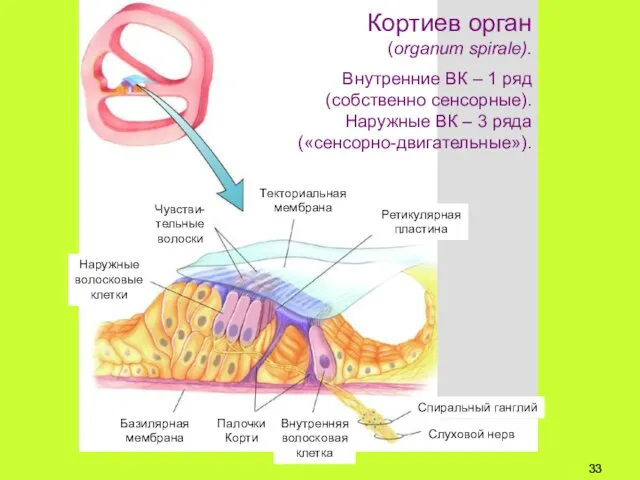
Кортиев орган
(organum spirale).
Внутренние ВК – 1 ряд
(собственно сенсорные).
Наружные ВК –
Кортиев орган
(organum spirale).
Внутренние ВК – 1 ряд
(собственно сенсорные).
Наружные ВК –
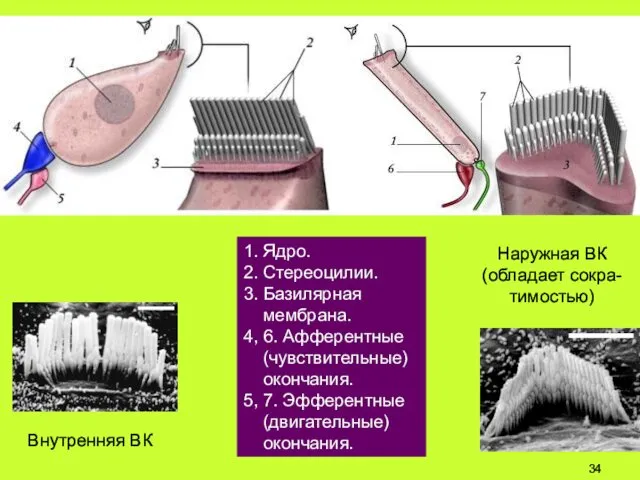
1. Ядро.
2. Стереоцилии.
3. Базилярная
мембрана.
4, 6. Афферентные
(чувствительные)
1. Ядро.
2. Стереоцилии.
3. Базилярная
мембрана.
4, 6. Афферентные
(чувствительные)
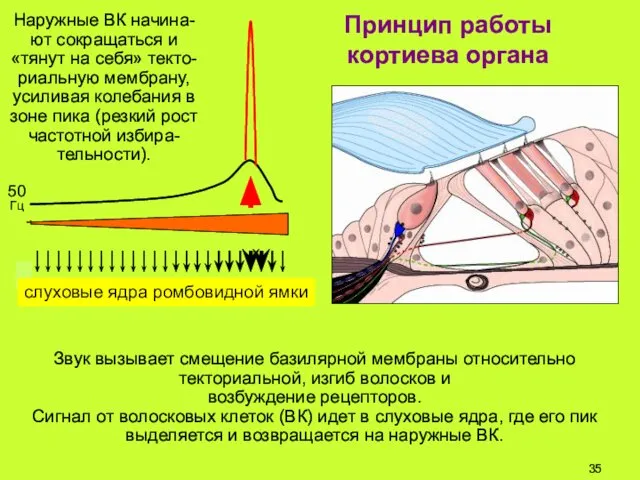
Звук вызывает смещение базилярной мембраны относительно текториальной, изгиб волосков и
возбуждение
Звук вызывает смещение базилярной мембраны относительно текториальной, изгиб волосков и
возбуждение
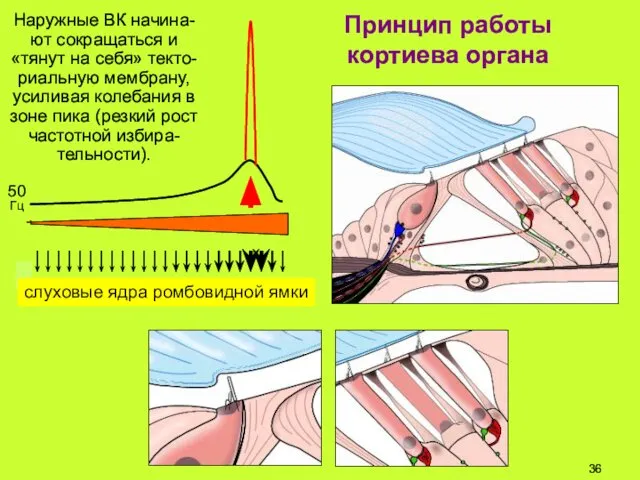
Принцип работы
кортиева органа
Наружные ВК начина-ют сокращаться и «тянут на себя»
Принцип работы
кортиева органа
Наружные ВК начина-ют сокращаться и «тянут на себя»
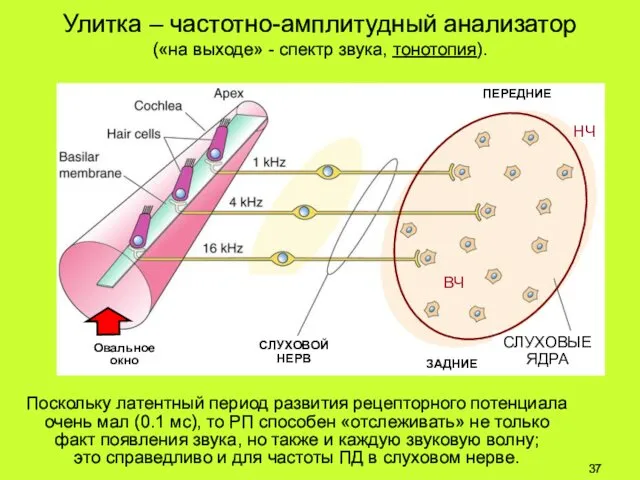
Улитка – частотно-амплитудный анализатор («на выходе» - спектр звука, тонотопия).
Поскольку латентный
Улитка – частотно-амплитудный анализатор («на выходе» - спектр звука, тонотопия).
Поскольку латентный
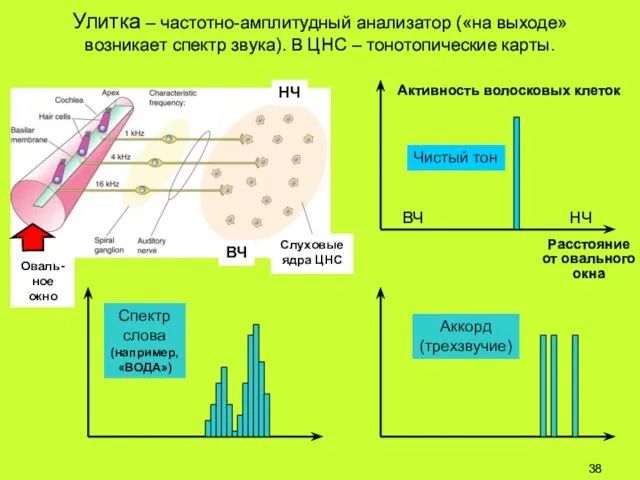
Улитка – частотно-амплитудный анализатор («на выходе» возникает спектр звука). В ЦНС
Улитка – частотно-амплитудный анализатор («на выходе» возникает спектр звука). В ЦНС
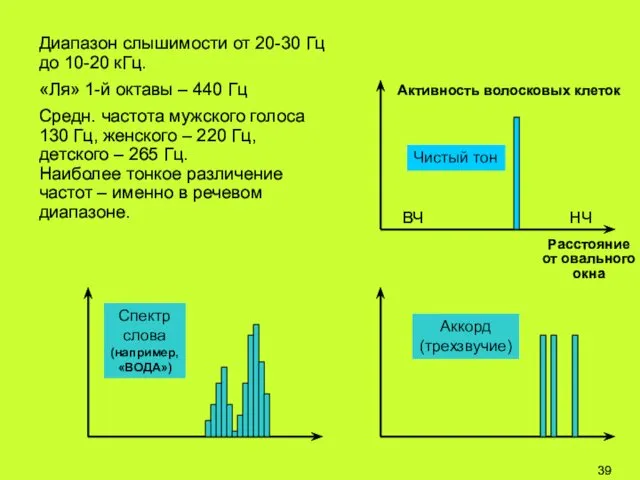
Диапазон слышимости от 20-30 Гц до 10-20 кГц.
«Ля» 1-й октавы –
Диапазон слышимости от 20-30 Гц до 10-20 кГц.
«Ля» 1-й октавы –
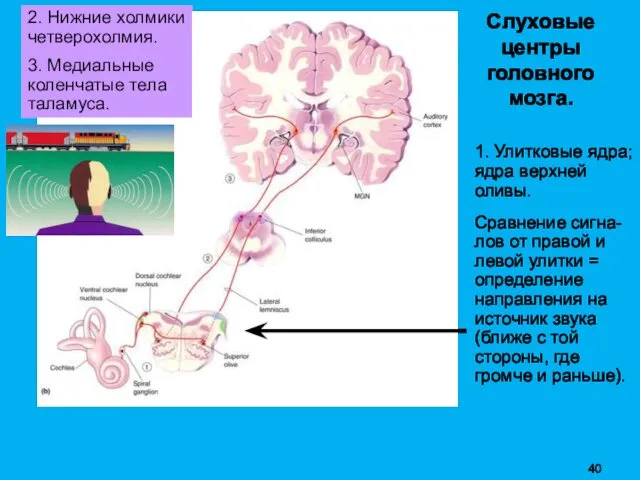
Слуховые
центры
головного
мозга.
1. Улитковые ядра; ядра верхней оливы.
Сравнение сигна-лов от правой и
Слуховые
центры
головного
мозга.
1. Улитковые ядра; ядра верхней оливы.
Сравнение сигна-лов от правой и
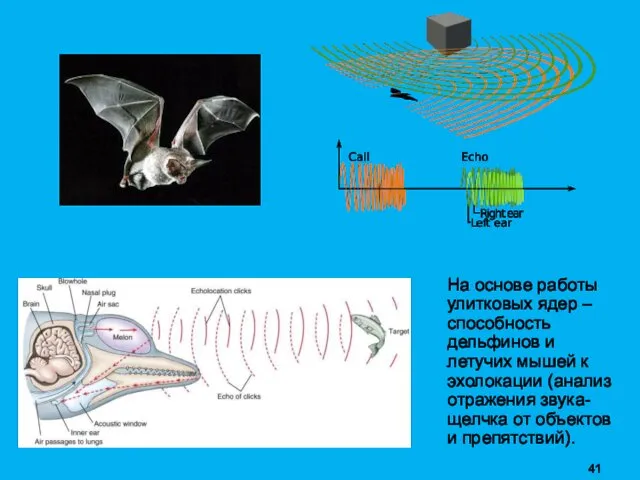
На основе работы улитковых ядер – способность дельфинов и летучих мышей
На основе работы улитковых ядер – способность дельфинов и летучих мышей
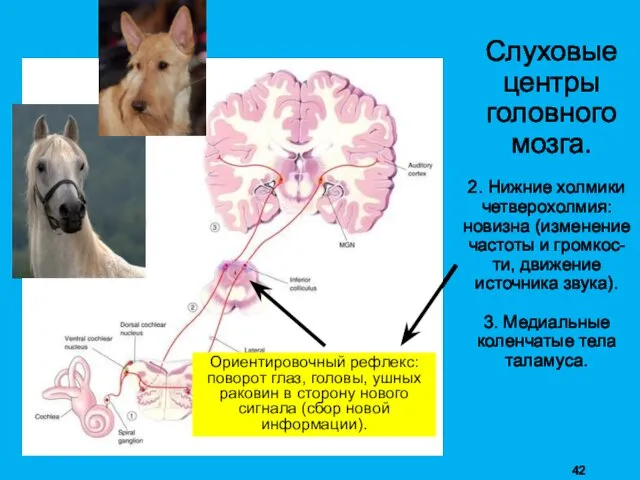
Слуховые
центры
головного
мозга.
2. Нижние холмики четверохолмия: новизна (изменение частоты и громкос-ти, движение
Слуховые
центры
головного
мозга.
2. Нижние холмики четверохолмия: новизна (изменение частоты и громкос-ти, движение
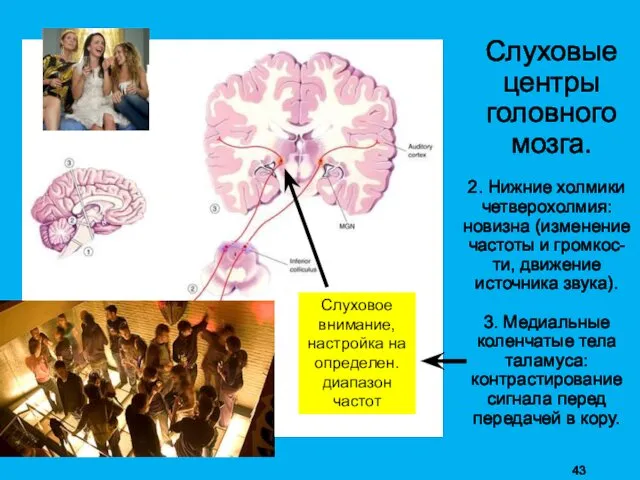
Слуховые
центры
головного
мозга.
2. Нижние холмики четверохолмия: новизна (изменение частоты и громкос-ти, движение
Слуховые
центры
головного
мозга.
2. Нижние холмики четверохолмия: новизна (изменение частоты и громкос-ти, движение
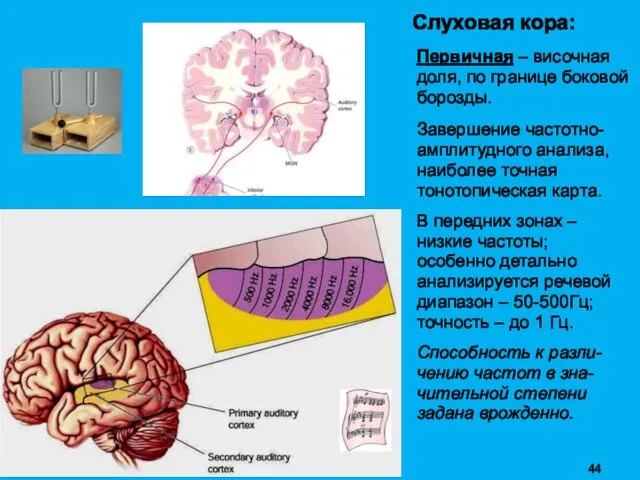
Слуховая кора:
Первичная – височная доля, по границе боковой борозды.
Завершение частотно-амплитудного анализа,
Слуховая кора:
Первичная – височная доля, по границе боковой борозды.
Завершение частотно-амплитудного анализа,
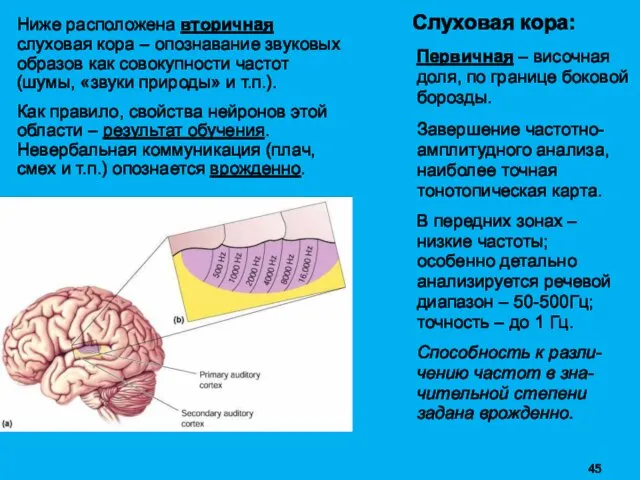
Ниже расположена вторичная слуховая кора – опознавание звуковых образов как совокупности
Ниже расположена вторичная слуховая кора – опознавание звуковых образов как совокупности
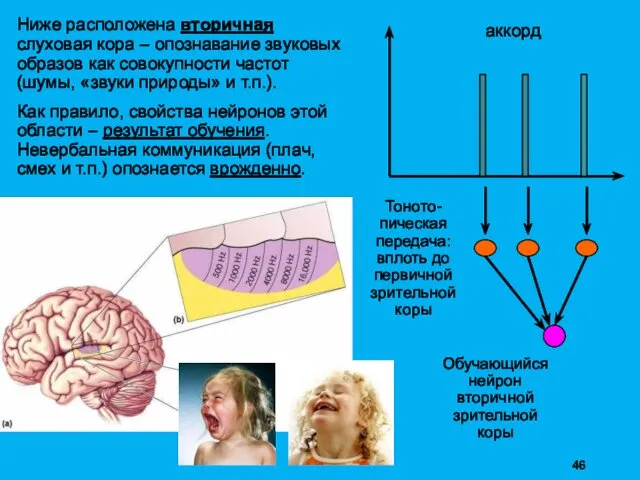
Ниже расположена вторичная слуховая кора – опознавание звуковых образов как совокупности
Ниже расположена вторичная слуховая кора – опознавание звуковых образов как совокупности
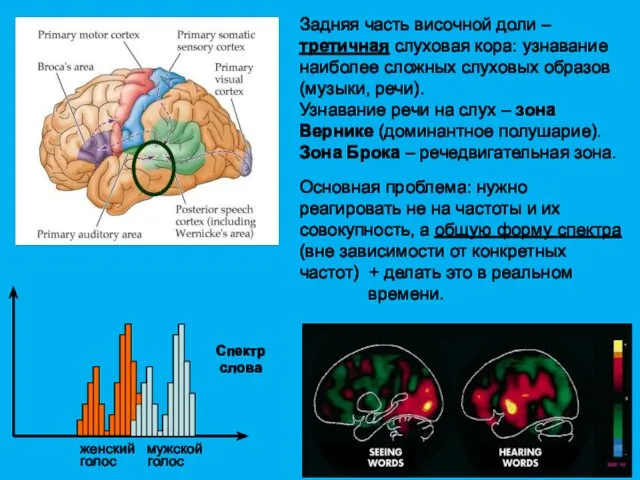
Задняя часть височной доли – третичная слуховая кора: узнавание наиболее сложных
Задняя часть височной доли – третичная слуховая кора: узнавание наиболее сложных
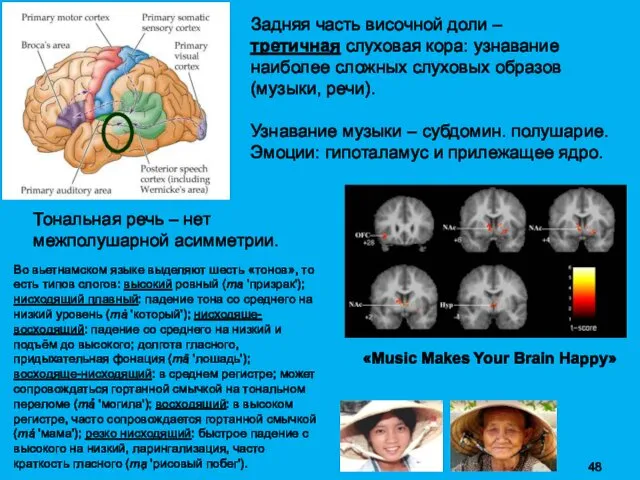
Задняя часть височной доли –
третичная слуховая кора: узнавание наиболее сложных
Задняя часть височной доли –
третичная слуховая кора: узнавание наиболее сложных
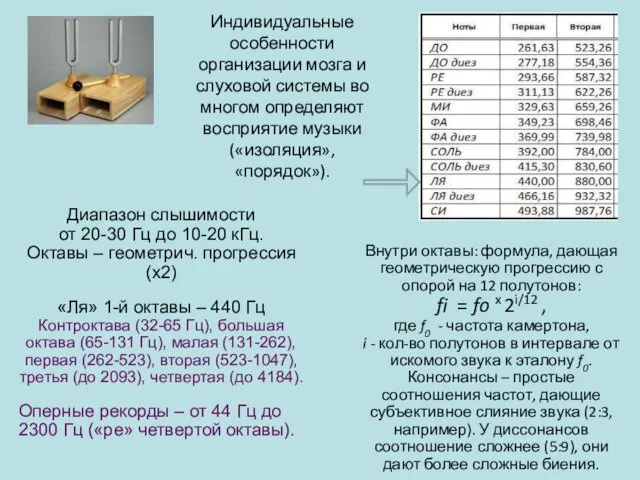
Индивидуальные особенности организации мозга и слуховой системы во многом определяют восприятие
Индивидуальные особенности организации мозга и слуховой системы во многом определяют восприятие
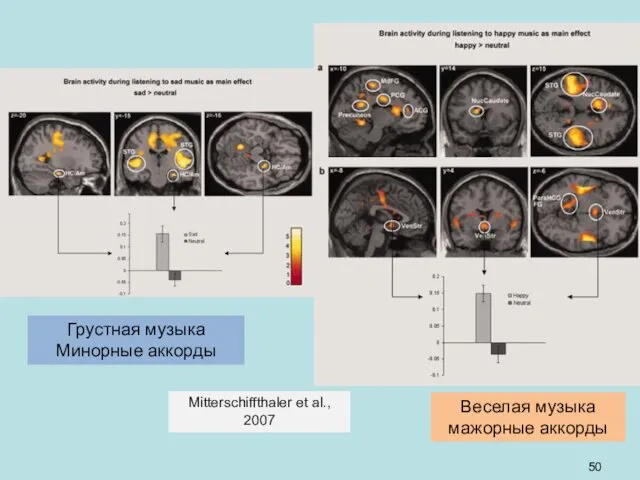
Грустная музыка
Минорные аккорды
Веселая музыка
мажорные аккорды
Mitterschiffthaler et al.,
2007
Грустная музыка
Минорные аккорды
Веселая музыка
мажорные аккорды
Mitterschiffthaler et al.,
2007
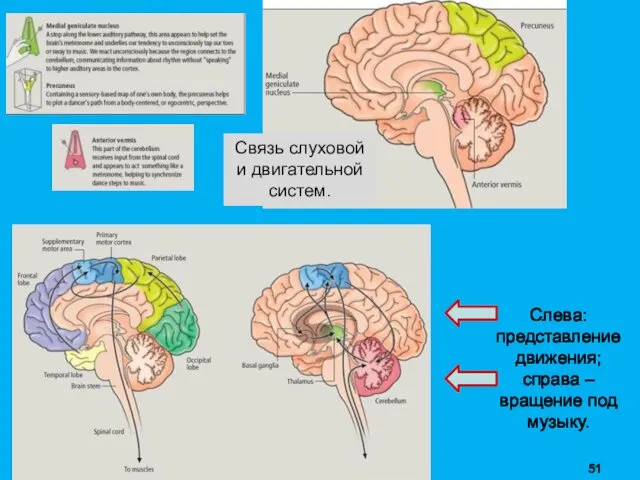
Слева: представление движения;
справа – вращение под музыку.
Связь слуховой и двигательной систем.
Слева: представление движения;
справа – вращение под музыку.
Связь слуховой и двигательной систем.
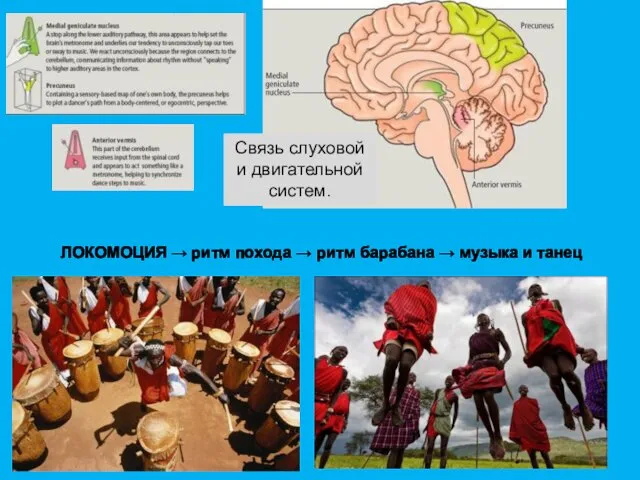
Связь слуховой и двигательной систем.
ЛОКОМОЦИЯ → ритм похода → ритм барабана
Связь слуховой и двигательной систем.
ЛОКОМОЦИЯ → ритм похода → ритм барабана
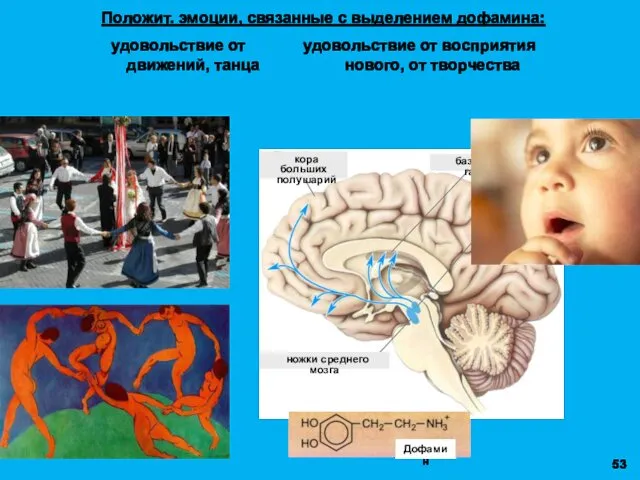
Положит. эмоции, связанные с выделением дофамина:
удовольствие от удовольствие от восприятия
движений,
Положит. эмоции, связанные с выделением дофамина:
удовольствие от удовольствие от восприятия
движений,

http://mglinets.narod.ru/mypubl/mgl8.htm
http://mglinets.narod.ru/mypubl/mgl6.htm
Molecular evolution of the vertebrate mechanosensory cell and ear
BERND FRITZSCH, KIRK
http://mglinets.narod.ru/mypubl/mgl8.htm
http://mglinets.narod.ru/mypubl/mgl6.htm
Molecular evolution of the vertebrate mechanosensory cell and ear
BERND FRITZSCH, KIRK
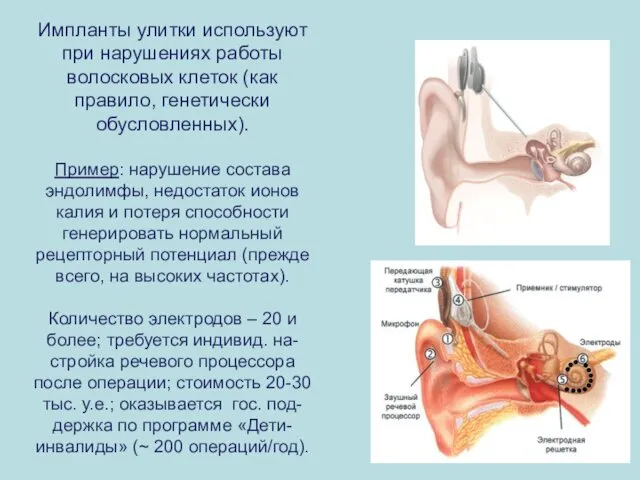
Импланты улитки используют при нарушениях работы волосковых клеток (как правило, генетически
Импланты улитки используют при нарушениях работы волосковых клеток (как правило, генетически

Дали, Кандинский, Шагал, Матисс
Дали, Кандинский, Шагал, Матисс
Спасибо за внимание!
Через неделю (30-го июня):
«Вкус, обоняние и немного
Спасибо за внимание!
Через неделю (30-го июня):
«Вкус, обоняние и немного
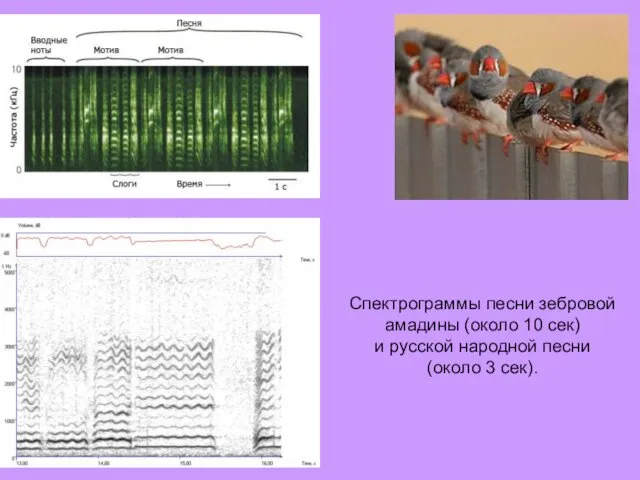
Спектрограммы песни зебровой амадины (около 10 сек)
и русской народной песни
Спектрограммы песни зебровой амадины (около 10 сек)
и русской народной песни
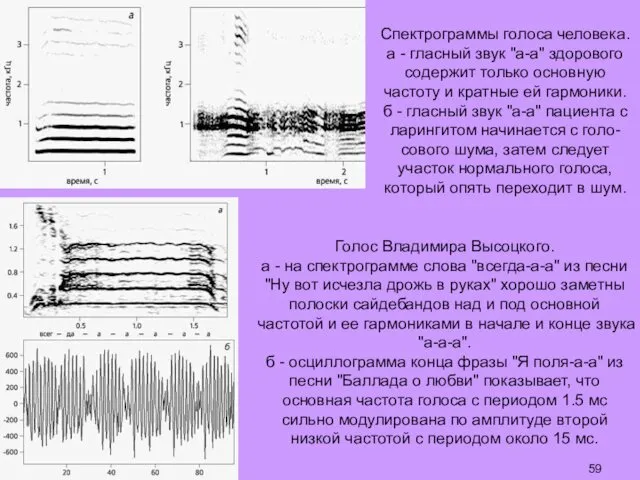
Спектрограммы голоса человека.
а - гласный звук "а-а" здорового содержит только
Спектрограммы голоса человека.
а - гласный звук "а-а" здорового содержит только
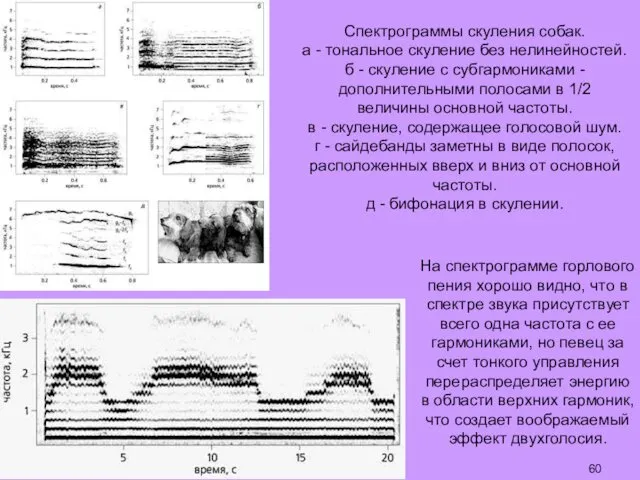
На спектрограмме горлового пения хорошо видно, что в спектре звука присутствует
На спектрограмме горлового пения хорошо видно, что в спектре звука присутствует
 Сердечно-сосудистая система
Сердечно-сосудистая система  Деревья (trees) «…великое Дерево Жизни заполняет земную кору своими мертвыми и сломанными ветвями и покрывает поверхность вечно ветвящимися и прекрасными побегами» Ч. Дарвин
Деревья (trees) «…великое Дерево Жизни заполняет земную кору своими мертвыми и сломанными ветвями и покрывает поверхность вечно ветвящимися и прекрасными побегами» Ч. Дарвин Тип кишечнополостные, общая характеристика
Тип кишечнополостные, общая характеристика Презентация на тему "Речной рак" - скачать презентации по Биологии
Презентация на тему "Речной рак" - скачать презентации по Биологии Генетические методы типирования HLA системы метод секвенирования
Генетические методы типирования HLA системы метод секвенирования Презентация на тему Царство Грибов
Презентация на тему Царство Грибов  Витаминдер биотехнологиясы. Витаминдердің жеке биотехнологиясы. Нормофлора препараттарының жалпы сипаттамасы
Витаминдер биотехнологиясы. Витаминдердің жеке биотехнологиясы. Нормофлора препараттарының жалпы сипаттамасы Продуктивність насаджень дуба червоного в умовах ДП Старосамбірське лісомисливське господарство Сусідовицького лісництва
Продуктивність насаджень дуба червоного в умовах ДП Старосамбірське лісомисливське господарство Сусідовицького лісництва Исследовательская работа: «количественное определение витамина С в продуктах питания йодометрическим методом» Выполнили: учени
Исследовательская работа: «количественное определение витамина С в продуктах питания йодометрическим методом» Выполнили: учени Презентация на тему Весеннее пробуждение растений
Презентация на тему Весеннее пробуждение растений  Ткани растений
Ткани растений Презентация по экологии Оценка пищевого статуса при развитии адаптационной резистентности организма.
Презентация по экологии Оценка пищевого статуса при развитии адаптационной резистентности организма.  Презентация на тему "Мир вокруг нас. Игровые моменты" - скачать презентации по Биологии
Презентация на тему "Мир вокруг нас. Игровые моменты" - скачать презентации по Биологии Строение клеток прокариот Подготовила ученица 10-А класса ЭМЛ Огурцова Валерия
Строение клеток прокариот Подготовила ученица 10-А класса ЭМЛ Огурцова Валерия Заповідник Асканія-Нова
Заповідник Асканія-Нова Фотосинтез
Фотосинтез Презентация на тему Эволюция пищеварительной системы животных
Презентация на тему Эволюция пищеварительной системы животных Внутреннее строение рыб
Внутреннее строение рыб Клас Птахи
Клас Птахи Семечковые и косточковые плоды. Субтропические и тропические плоды
Семечковые и косточковые плоды. Субтропические и тропические плоды Царство животные
Царство животные Память. Типологии и свойства памяти. Процессы памяти. Виды памяти. Индивидуальные особенности памяти
Память. Типологии и свойства памяти. Процессы памяти. Виды памяти. Индивидуальные особенности памяти Презентация на тему "Чарльз Роберт Дарвин. Жизнь, Биография, Научные труды" - скачать бесплатно презентации по Биологии
Презентация на тему "Чарльз Роберт Дарвин. Жизнь, Биография, Научные труды" - скачать бесплатно презентации по Биологии Тип Плоские черви (Plathelminthes)
Тип Плоские черви (Plathelminthes)  Зчеплене успадкування
Зчеплене успадкування Взаимосвязь между структурой, свойствами и функциями клеточной мембраны
Взаимосвязь между структурой, свойствами и функциями клеточной мембраны Животные Африки
Животные Африки Многообразие растений
Многообразие растений