- Мышечные ткани

Содержание
- 2. Структурно-функциональные признаки мышечных тканей 1.Возбудимость – способность отвечать на раздражитель изменением мембранного потенциала. 2.Проводимость – способность
- 3. По строению различают мышечные ткани : 1.Гладкие – состоят из клеток, не обладающих исчерченностью. 2.Поперечнополосатые –
- 4. Гистогенетическая классификация (учитывает источник развития) 1.Из мезенхимы – висцеральная гладкая мышечная ткань (стенки полых органов, трубчатые
- 5. Скелетная поперечнополосатая мышечная ткань 1.Называют произвольной, т.к. её сокращение контролируется нашей волей (исключение – мышцы, поддерживающие
- 6. Гистогенез скелетной мышечной ткани. 1.Из клеток миотомов образуются миобласты, которые образуют две линии дифференцировки. 2.Миобласты одной
- 7. Гистогенез скелетной мышечной ткани 3.Из мышечных трубочек образуются зрелые мышечные волокна, в которых миофибриллы располагаются в
- 8. Миосателлитоцит 1.Камбиальные клетки скелетной мышечной ткани в течение всей жизни человека. 2.Прилежат к поверхности симпласта, располагаясь
- 9. Продолжение 4.Активируются: - при регенерации мышечных волокон после их повреждения; -при усиленной нагрузке сливаются с миосимпластом,
- 10. Поперечнополосатое мышечное волокно 1.Структурно- функциональная единица скелетной мышечной ткани. 2.Цилиндрическое образование. 3.Диаметр–10-100 мкм, длина – 10-30
- 11. Базальная мембрана 1.Толстая, располагается снаружи от сарколеммы миосимпласта и миосателлитоцитов. 2.В мембрану вплетаются коллагеновые волокна РСТ,
- 12. Миосимпласт 1.Включает ядра, саркоплазму, сарколемму. 2.Ядра:-светлые, - длинная ось вдоль волокна, -располагаются на периферии под сарколеммой,
- 13. Функциональные аппараты мышечного волокна 1.Сократительный. 2.Передачи возбуждения с сарколеммы на сократительный аппарат. 3.Опорный. 4.Энергетический. 5.Синтетический. 6.Лизосомальный.
- 14. Сократительный аппарат Представлен миофибриллами. Миофибриллы: 1.Располагаются продольно в центральной части саркоплазмы. 2.Отделяются друг от друга митохондриями
- 15. Сократительный аппарат 3.Имеют вид нитей. 4.Диаметр -1-2 мкм. 5.Длина сопоставима с длиной мышечного волокна. 6.Количество от
- 16. Саркомер 1.Структурнофункциональная единица миофибриллы. 2.Это участок миофибриллы, который расположен между двумя соседними телофрагмами (Z) и включает
- 17. Саркомер (продолжение) 3.Это упорядоченная система толстых и тонких нитей (филаментов). 4. Z-линия (тонкая тёмная) ---телофрагма, рассекает
- 18. Толстые филаменты саркомера 1.Сосредоточены в А-диске. 2.Связаны с мезофрагмой. 3.Образованы молекулами фибриллярного сократительного белка миозина. 4.Белок
- 19. Тонкие филаменты саркомера 1.Образуют I-диск. 2.Частично проникают в А-диск между толстыми филаментами. 3.Одним концом прикреплены к
- 20. Теория скользящих нитей (механизм мышечного сокращения) 1.Тонкие нити вдвигаются в промежутки между толстыми нитями без изменения
- 21. Аппарат передачи возбуждения 1.Образован ретикулярным элементом (L-система) и Т-системой. 2.L-система (longus – продольный) – система уплощённых
- 22. Аппарат передачи возбуждения (продолжение) 3.Т-трубочки (transversus поперечный) – это впячивания сарколеммы, которые отходят от неё под
- 23. Опорный аппарат мышечного волокна 1.Телофрагма (Z- линия) – белок альфа-актинин – это область прикрепления тонких филаментов.
- 24. Энергетический аппарат мышечного волокна: 1.Митохондрии. 2.Включения гликогена. 3.Липидные включения. 4.Миоглобин – железосодержащий кислород- связывающий пигмент. Синтетический
- 25. Типы мышечных волокон 1.Красные волокна (первый тип): -малой толщины; -обилие миоглобина; - обилие цитохромов (дыхательных ферментов)
- 26. Типы мышечных волокон (продолжение) 2.Белые волокна (второй тип): -толще, чем красные; -содержат меньше миоглобина; -митохондрий меньше;
- 27. Типы мышечных волокон (продолжение) 3.Промежуточные волокна (третий тип): -сокращаются быстро; -устойчивы к утомлению. Большинство мышц человека
- 28. Регенерация скелетной мышечной ткани 1. Физиологическая регенерация волокон непрерывно осуществляется в нормальных условиях и состоит в
- 29. продолжение -миграция фагоцитов в область повреждения мышечных волокон; -фагоцитоз разрушенных фрагментов ; -деление камбиальных клеток миосателлитоцитов
- 30. Мышца как орган 1.Состоит из пучков мышечных волокон, связанных воедино системой оболочек: -эпимизием; -перимизием; -эндомизием. Эпимизий
- 31. продолжение Перимизий –рыхлая волокнистая соединительная ткань, отходящая от внутренней поверхности эпимизия вглубь мышцы и окружающая пучки
- 32. продолжение Эндомизий – тонкие прослойки рыхлой волокнистой соединительной ткани, окружающие каждое мышечное волокно. Коллагеновые волокна эндомизия
- 33. Кровоснабжение скелетной мышцы Осуществляется артериями, которые проникают в мышцу вместе с нервами через эпимизий, идут в
- 34. Двигательная иннервация мышечных волокон Обеспечивается аксонами двигательных нейронов, которые образуют на мышечных волокнах нервно-мышечные синапсы (моторные
- 35. Чувствительная иннервация мышечных волокон Обеспечивается нервно-мышечными веретёнами – это рецепторы растяжения мышечных волокон, которые располагаются в
- 36. Гладкая мышечная ткань мезенхимного типа 1.Источник развития мезенхима, которая выселяется: а) из спланхнотома и образует унитарную
- 37. Гладкий миоцит 1.Форма веретеновидная. 2.Длина –50-500(1000) мкм. 3.Самые крупные миоциты матки при беременности, самые мелкие- в
- 38. Сократительный аппарат миоцита 1.Представлен тонкими актиновыми и толстыми миозиновыми филаментами. 2.Филаменты не формируют миофибрилл и саркомеров.
- 39. продолжение 5.Сокращение индуцируется притоком ионов кальция в цитоплазму из агр.-ЭПС и кавеол. 6.Это влечёт за собой
- 40. продолжение 8.Часть миозиновых «мостиков» не отсоединяется от актина, а остаётся с ним связанной. Поэтому гладкие миоциты
- 41. Тонкие актиновые филаменты 1. По количеству преобладают над толстыми филаментами. 2.Располагаются в цитоплазме пучками параллельно вдоль
- 42. Толстые миозиновые филаменты 1.Обладают различной длиной. 2.Покрыты миозиновыми головками по всей длине, поэтому не имеют центральной
- 43. Опорный аппарат 1.Система элементов цитоскелета. 2.Плотные тельца. 3.Базальная мембрана окружает плазмолемму каждого миоцита. На отдельных участках
- 44. Аппарат передачи возбуждения 1.агр.-ЭПС (L-система) рудиментарна и состоит из мелких цистерн и пузырьков. 2.Т-трубочки отсутствуют. 3.Имеются
- 45. Энергетический аппарат: митохондрии, включения гликогена, липидов. Располагаются в основном у полюсов ядра. Синтетический аппарат: гр.-ЭПС, свободные
- 46. Регуляция сократительной активности гладких миоцитов 1.Нервная регуляция: двигательные нервные окончания обнаруживаются на отдельных клетках и имеют
- 47. продолжение 2.Гуморальная регуляция: гладкие миоциты в отличие от скелетных мышечных волокон имеют на плазмолемме рецепторы к
- 48. Регенерация гладкой мышечной ткани 1. Физиологическая –путём обновления клеточных органелл. 2.Гипертрофия (увеличение количества и размеров всех
- 49. Мышечная ткань эпидермального происхождения 1.По строению –гладкие. 2.С железистыми клетками связаны десмосомами. 3.Снаружи покрыты базальной мембраной.
- 50. Гладкая мышечная ткань нейрального происхождения (мультиунитарная) 1.Сфинктер и дилататор радужки. 2.Форма веретеновидная. 3.На каждом миоците имеется
- 51. Сердечная поперечнополосатая мышечная ткань (миокард сердца).
- 52. Гистогенез сердечной мышечной ткани. 1.Источник развития – миоэпикардиальная пластинка (мезодерма висцерального листка спланхнотома). 2.Образующиеся кардиомиоциты выстраиваются
- 53. Вставочный диск Включает: 1.Десмосомы –механическое соединение кардиомиоцитов. В десмосомы вплетаются миофибриллы. 2.Нексусы (щелевые контакты) -обеспечивают быстрое
- 54. Типы кардиомиоцитов 1.Сократительные (типичные, рабочие). 2.Секретрные. 3.Проводящие (атипичные).
- 55. Сократительные кардиомиоциты 1.Форма призматическая или отростчатая. 2.Одно или два ядра в центре клетки. 3.Миофибриллы располагаются по
- 56. Секреторные кардиомиоциты 1.Располагаются в предсердиях. 2.Форма отростчатачя. 3.Слабое развитие сократительного аппарата. 4.Хорошо развит синтетический аппарат. 5.
- 57. Энергетический. синтетический , лизосомальный аппараты. См.выше - скелетная мышечная ткань
- 58. Проводящие кардиомиоциты 1.Крупнее, чем сократительные. 2.Богаче цитоплазмой. 3.Мало миофибрилл, поэтому при окраске гематоксилином и эозином выглядят
- 59. Типы проводящих кардиомиоцитов 1.Р-клетки (клетки пейсмекеры): -располагаются в центре синусного узла и немного в предсердно-желудочковом узле;
- 60. продолжение 3.Клетки Пуркинье: -самые крупные не только в проводящей системе, но и во всём миокарде; -входят
- 61. Регенерация сердечной мышечной ткани 1.На субмикроскопическом уровне – обновление всех структур клеток. 2.В случае гибели кардиомиоцитов
- 62. Кровоснабжение сердечной мышечной ткани 1.По уровню кровоснабжения превосходит головной мозг, уступая лишь почке. 2.Ветви коронарных артерий
- 63. Иннервация сердечной мышечной ткани 1.Осущестляется нервными волокнами симпатического и парасимпатического отделов вегетативной нервной системы. 2.Вегетативная нервная
- 65. Скачать презентацию

Структурно-функциональные признаки мышечных тканей
1.Возбудимость – способность отвечать на раздражитель изменением мембранного
Структурно-функциональные признаки мышечных тканей
1.Возбудимость – способность отвечать на раздражитель изменением мембранного

По строению различают мышечные ткани :
1.Гладкие – состоят из клеток, не
По строению различают мышечные ткани :
1.Гладкие – состоят из клеток, не

Гистогенетическая классификация (учитывает источник развития)
1.Из мезенхимы – висцеральная гладкая мышечная ткань
Гистогенетическая классификация (учитывает источник развития)
1.Из мезенхимы – висцеральная гладкая мышечная ткань
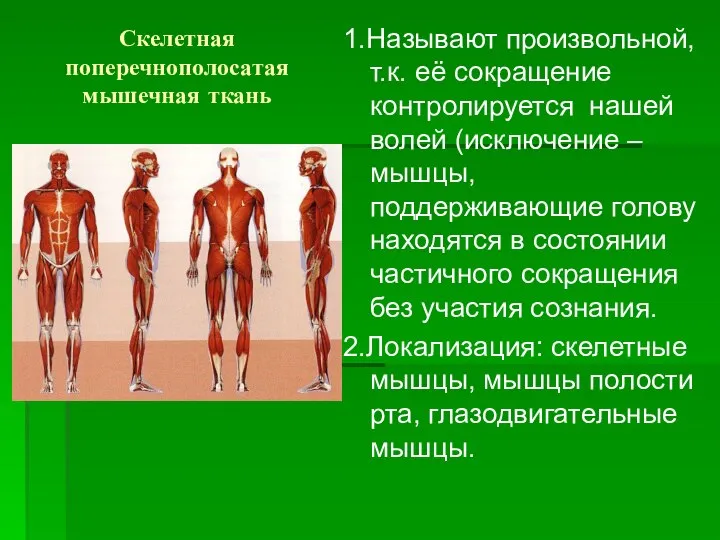
Скелетная поперечнополосатая мышечная ткань
1.Называют произвольной, т.к. её сокращение контролируется нашей волей
Скелетная поперечнополосатая мышечная ткань
1.Называют произвольной, т.к. её сокращение контролируется нашей волей

Гистогенез скелетной мышечной ткани.
1.Из клеток миотомов
образуются миобласты,
которые образуют две
линии
Гистогенез скелетной мышечной ткани.
1.Из клеток миотомов
образуются миобласты,
которые образуют две
линии

Гистогенез скелетной мышечной ткани
3.Из мышечных трубочек
образуются зрелые
мышечные волокна, в
которых миофибриллы
располагаются в
Гистогенез скелетной мышечной ткани
3.Из мышечных трубочек
образуются зрелые
мышечные волокна, в
которых миофибриллы
располагаются в

Миосателлитоцит
1.Камбиальные клетки скелетной мышечной ткани в течение всей жизни человека.
2.Прилежат к
Миосателлитоцит
1.Камбиальные клетки скелетной мышечной ткани в течение всей жизни человека.
2.Прилежат к

Продолжение
4.Активируются:
- при регенерации мышечных волокон после их повреждения;
-при усиленной нагрузке сливаются
Продолжение
4.Активируются:
- при регенерации мышечных волокон после их повреждения;
-при усиленной нагрузке сливаются
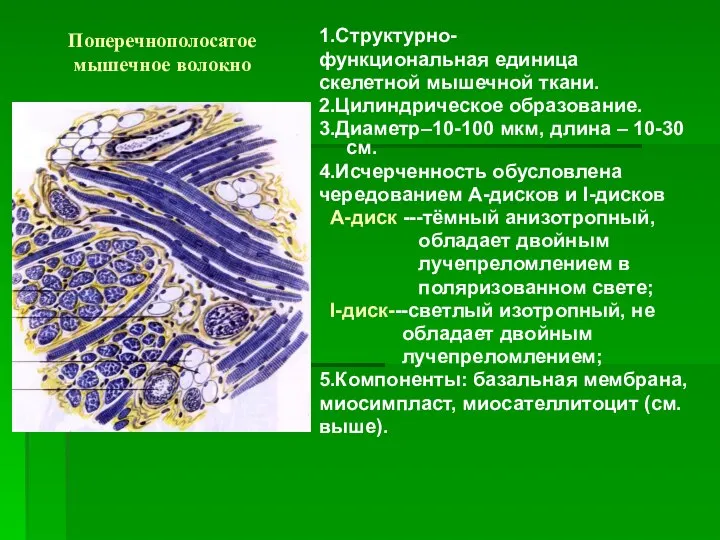
Поперечнополосатое мышечное волокно
1.Структурно-
функциональная единица
скелетной мышечной ткани.
2.Цилиндрическое образование.
3.Диаметр–10-100 мкм, длина – 10-30
Поперечнополосатое мышечное волокно
1.Структурно-
функциональная единица
скелетной мышечной ткани.
2.Цилиндрическое образование.
3.Диаметр–10-100 мкм, длина – 10-30

Базальная мембрана
1.Толстая, располагается снаружи от сарколеммы миосимпласта и миосателлитоцитов.
2.В мембрану вплетаются
Базальная мембрана
1.Толстая, располагается снаружи от сарколеммы миосимпласта и миосателлитоцитов.
2.В мембрану вплетаются

Миосимпласт
1.Включает ядра, саркоплазму, сарколемму.
2.Ядра:-светлые,
- длинная ось вдоль волокна,
-располагаются на
Миосимпласт
1.Включает ядра, саркоплазму, сарколемму.
2.Ядра:-светлые,
- длинная ось вдоль волокна,
-располагаются на

Функциональные аппараты мышечного волокна
1.Сократительный.
2.Передачи возбуждения с сарколеммы на сократительный аппарат.
3.Опорный.
4.Энергетический.
5.Синтетический.
6.Лизосомальный.
Функциональные аппараты мышечного волокна
1.Сократительный.
2.Передачи возбуждения с сарколеммы на сократительный аппарат.
3.Опорный.
4.Энергетический.
5.Синтетический.
6.Лизосомальный.

Сократительный аппарат
Представлен
миофибриллами.
Миофибриллы:
1.Располагаются
продольно
в центральной части
саркоплазмы.
2.Отделяются друг от
друга митохондриями
и цистернами агр-ЭПС
(саркоплазматический ретикулум).
Сократительный аппарат
Представлен
миофибриллами.
Миофибриллы:
1.Располагаются
продольно
в центральной части
саркоплазмы.
2.Отделяются друг от
друга митохондриями
и цистернами агр-ЭПС
(саркоплазматический ретикулум).

Сократительный аппарат
3.Имеют вид нитей.
4.Диаметр -1-2 мкм.
5.Длина сопоставима с длиной мышечного волокна.
6.Количество
Сократительный аппарат
3.Имеют вид нитей.
4.Диаметр -1-2 мкм.
5.Длина сопоставима с длиной мышечного волокна.
6.Количество

Саркомер
1.Структурнофункциональная единица миофибриллы.
2.Это участок
миофибриллы, который
расположен между двумя
соседними телофрагмами
(Z) и включает А-диск
Саркомер
1.Структурнофункциональная единица миофибриллы.
2.Это участок
миофибриллы, который
расположен между двумя
соседними телофрагмами
(Z) и включает А-диск
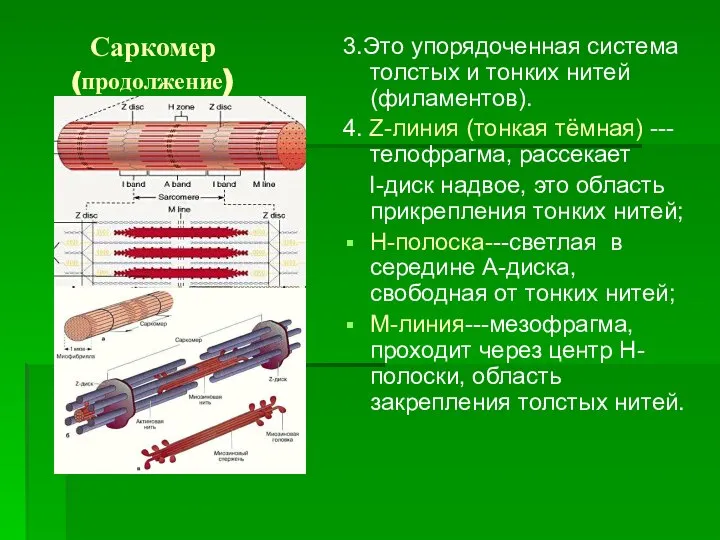
Саркомер (продолжение)
3.Это упорядоченная система толстых и тонких нитей (филаментов).
4. Z-линия (тонкая
Саркомер (продолжение)
3.Это упорядоченная система толстых и тонких нитей (филаментов).
4. Z-линия (тонкая

Толстые филаменты саркомера
1.Сосредоточены в А-диске.
2.Связаны с мезофрагмой.
3.Образованы молекулами фибриллярного сократительного белка
Толстые филаменты саркомера
1.Сосредоточены в А-диске.
2.Связаны с мезофрагмой.
3.Образованы молекулами фибриллярного сократительного белка

Тонкие филаменты саркомера
1.Образуют I-диск.
2.Частично проникают в А-диск между толстыми филаментами.
3.Одним концом
Тонкие филаменты саркомера
1.Образуют I-диск.
2.Частично проникают в А-диск между толстыми филаментами.
3.Одним концом

Теория скользящих нитей (механизм мышечного сокращения)
1.Тонкие нити вдвигаются в
промежутки между толстыми
нитями
Теория скользящих нитей (механизм мышечного сокращения)
1.Тонкие нити вдвигаются в
промежутки между толстыми
нитями

Аппарат передачи возбуждения
1.Образован ретикулярным
элементом (L-система) и
Т-системой.
2.L-система (longus –
продольный) – система
уплощённых
Аппарат передачи возбуждения
1.Образован ретикулярным
элементом (L-система) и
Т-системой.
2.L-система (longus –
продольный) – система
уплощённых

Аппарат передачи возбуждения
(продолжение)
3.Т-трубочки (transversus поперечный) – это впячивания сарколеммы, которые отходят
Аппарат передачи возбуждения
(продолжение)
3.Т-трубочки (transversus поперечный) – это впячивания сарколеммы, которые отходят

Опорный аппарат мышечного волокна
1.Телофрагма (Z- линия) – белок альфа-актинин
– это область
Опорный аппарат мышечного волокна
1.Телофрагма (Z- линия) – белок альфа-актинин
– это область

Энергетический аппарат мышечного волокна:
1.Митохондрии.
2.Включения гликогена.
3.Липидные включения.
4.Миоглобин – железосодержащий кислород- связывающий пигмент.
Синтетический
Энергетический аппарат мышечного волокна:
1.Митохондрии.
2.Включения гликогена.
3.Липидные включения.
4.Миоглобин – железосодержащий кислород- связывающий пигмент.
Синтетический

Типы мышечных волокон
1.Красные волокна (первый тип):
-малой толщины;
-обилие миоглобина;
- обилие цитохромов (дыхательных
Типы мышечных волокон
1.Красные волокна (первый тип):
-малой толщины;
-обилие миоглобина;
- обилие цитохромов (дыхательных

Типы мышечных волокон
(продолжение)
2.Белые волокна (второй тип):
-толще, чем красные;
-содержат меньше миоглобина;
-митохондрий меньше;
-АТФ-фаза
Типы мышечных волокон
(продолжение)
2.Белые волокна (второй тип):
-толще, чем красные;
-содержат меньше миоглобина;
-митохондрий меньше;
-АТФ-фаза

Типы мышечных волокон
(продолжение)
3.Промежуточные волокна (третий тип):
-сокращаются быстро;
-устойчивы к утомлению.
Большинство мышц человека
Типы мышечных волокон
(продолжение)
3.Промежуточные волокна (третий тип):
-сокращаются быстро;
-устойчивы к утомлению.
Большинство мышц человека

Регенерация скелетной мышечной ткани
1. Физиологическая регенерация волокон непрерывно осуществляется в нормальных
Регенерация скелетной мышечной ткани
1. Физиологическая регенерация волокон непрерывно осуществляется в нормальных

продолжение
-миграция фагоцитов в область повреждения мышечных волокон;
-фагоцитоз разрушенных фрагментов ;
-деление камбиальных
продолжение
-миграция фагоцитов в область повреждения мышечных волокон;
-фагоцитоз разрушенных фрагментов ;
-деление камбиальных
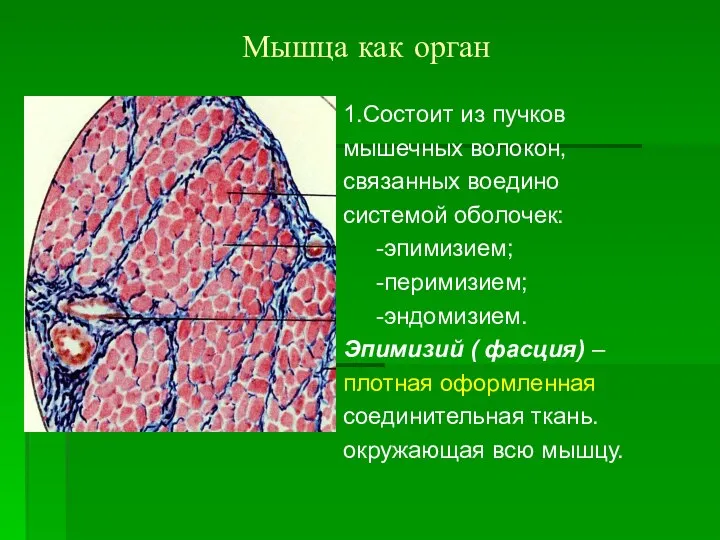
Мышца как орган
1.Состоит из пучков
мышечных волокон,
связанных воедино
системой оболочек:
-эпимизием;
-перимизием;
-эндомизием.
Эпимизий
Мышца как орган
1.Состоит из пучков
мышечных волокон,
связанных воедино
системой оболочек:
-эпимизием;
-перимизием;
-эндомизием.
Эпимизий

продолжение
Перимизий –рыхлая волокнистая соединительная ткань, отходящая от внутренней поверхности эпимизия вглубь
продолжение
Перимизий –рыхлая волокнистая соединительная ткань, отходящая от внутренней поверхности эпимизия вглубь

продолжение
Эндомизий – тонкие прослойки рыхлой волокнистой соединительной ткани, окружающие каждое мышечное
продолжение
Эндомизий – тонкие прослойки рыхлой волокнистой соединительной ткани, окружающие каждое мышечное
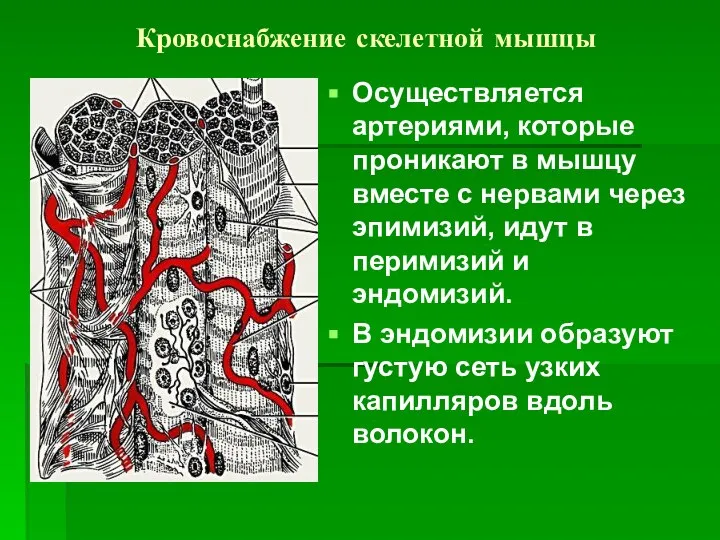
Кровоснабжение скелетной мышцы
Осуществляется артериями, которые проникают в мышцу вместе с нервами
Кровоснабжение скелетной мышцы
Осуществляется артериями, которые проникают в мышцу вместе с нервами
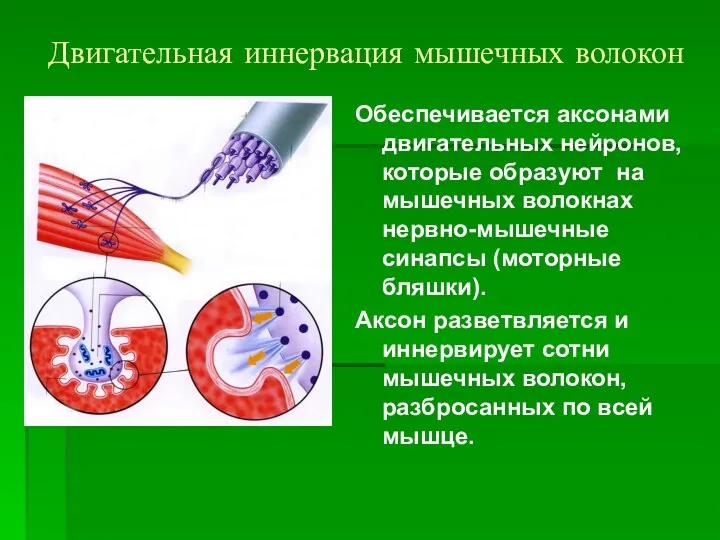
Двигательная иннервация мышечных волокон
Обеспечивается аксонами двигательных нейронов, которые образуют на мышечных
Двигательная иннервация мышечных волокон
Обеспечивается аксонами двигательных нейронов, которые образуют на мышечных

Чувствительная иннервация мышечных волокон
Обеспечивается нервно-мышечными веретёнами – это рецепторы растяжения мышечных
Чувствительная иннервация мышечных волокон
Обеспечивается нервно-мышечными веретёнами – это рецепторы растяжения мышечных
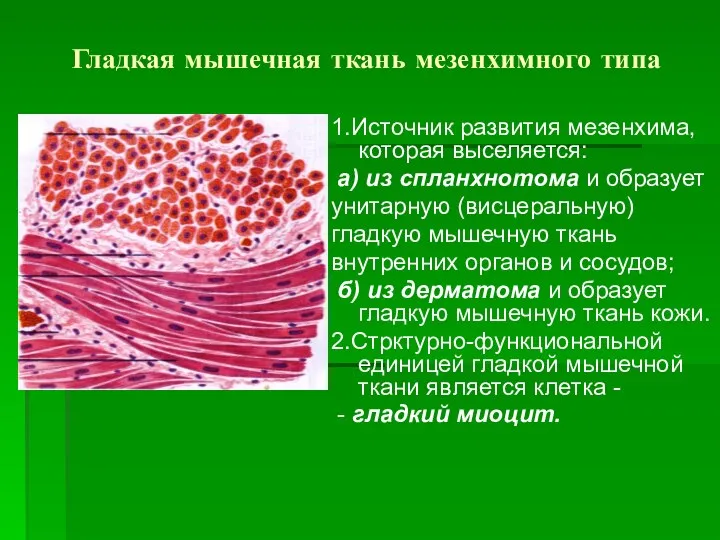
Гладкая мышечная ткань мезенхимного типа
1.Источник развития мезенхима, которая выселяется:
а) из
Гладкая мышечная ткань мезенхимного типа
1.Источник развития мезенхима, которая выселяется:
а) из
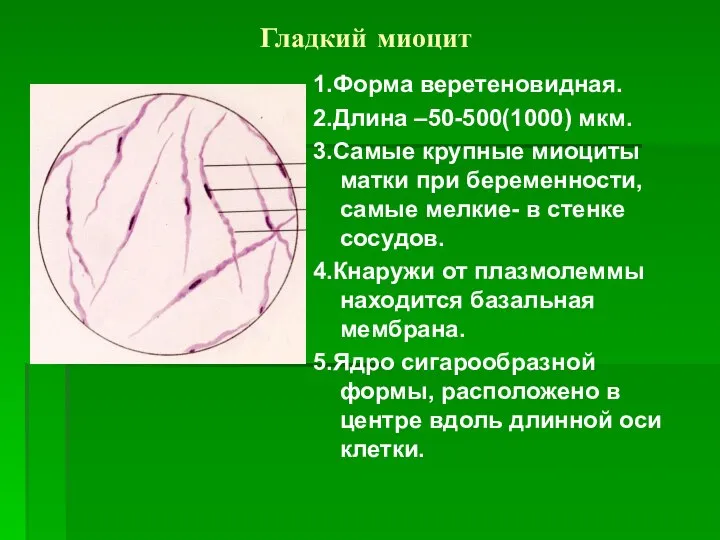
Гладкий миоцит
1.Форма веретеновидная.
2.Длина –50-500(1000) мкм.
3.Самые крупные миоциты матки при беременности, самые
Гладкий миоцит
1.Форма веретеновидная.
2.Длина –50-500(1000) мкм.
3.Самые крупные миоциты матки при беременности, самые

Сократительный аппарат миоцита
1.Представлен тонкими актиновыми и толстыми миозиновыми филаментами.
2.Филаменты не формируют
Сократительный аппарат миоцита
1.Представлен тонкими актиновыми и толстыми миозиновыми филаментами.
2.Филаменты не формируют

продолжение
5.Сокращение индуцируется притоком ионов кальция в цитоплазму из агр.-ЭПС и кавеол.
6.Это
продолжение
5.Сокращение индуцируется притоком ионов кальция в цитоплазму из агр.-ЭПС и кавеол.
6.Это

продолжение
8.Часть миозиновых «мостиков» не отсоединяется от актина, а остаётся с ним
продолжение
8.Часть миозиновых «мостиков» не отсоединяется от актина, а остаётся с ним
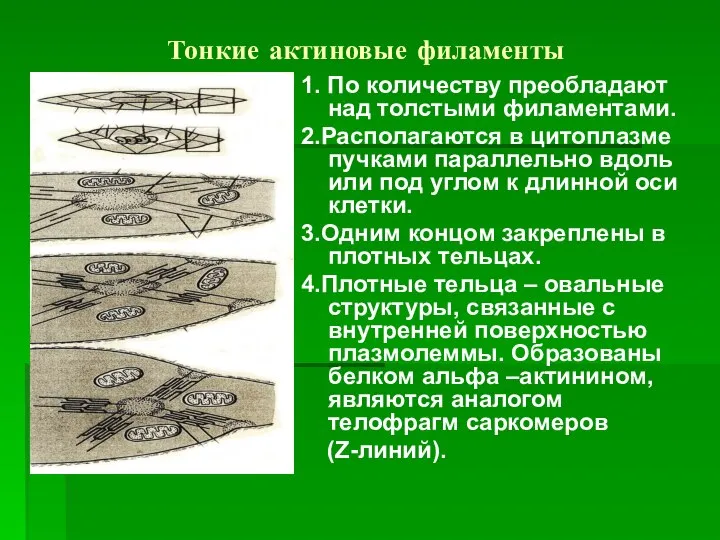
Тонкие актиновые филаменты
1. По количеству преобладают над толстыми филаментами.
2.Располагаются в цитоплазме
Тонкие актиновые филаменты
1. По количеству преобладают над толстыми филаментами.
2.Располагаются в цитоплазме

Толстые миозиновые филаменты
1.Обладают различной длиной.
2.Покрыты миозиновыми головками по всей длине, поэтому
Толстые миозиновые филаменты
1.Обладают различной длиной.
2.Покрыты миозиновыми головками по всей длине, поэтому

Опорный аппарат
1.Система элементов цитоскелета.
2.Плотные тельца.
3.Базальная мембрана окружает плазмолемму каждого миоцита. На
Опорный аппарат
1.Система элементов цитоскелета.
2.Плотные тельца.
3.Базальная мембрана окружает плазмолемму каждого миоцита. На

Аппарат передачи возбуждения
1.агр.-ЭПС (L-система) рудиментарна и состоит из мелких цистерн и
Аппарат передачи возбуждения
1.агр.-ЭПС (L-система) рудиментарна и состоит из мелких цистерн и

Энергетический аппарат: митохондрии, включения гликогена, липидов. Располагаются в основном у полюсов
Энергетический аппарат: митохондрии, включения гликогена, липидов. Располагаются в основном у полюсов

Регуляция сократительной активности гладких миоцитов
1.Нервная регуляция: двигательные нервные окончания обнаруживаются на
Регуляция сократительной активности гладких миоцитов
1.Нервная регуляция: двигательные нервные окончания обнаруживаются на

продолжение
2.Гуморальная регуляция: гладкие миоциты в отличие от скелетных мышечных волокон имеют
продолжение
2.Гуморальная регуляция: гладкие миоциты в отличие от скелетных мышечных волокон имеют

Регенерация гладкой мышечной ткани
1. Физиологическая –путём обновления клеточных органелл.
2.Гипертрофия (увеличение количества
Регенерация гладкой мышечной ткани
1. Физиологическая –путём обновления клеточных органелл.
2.Гипертрофия (увеличение количества

Мышечная ткань эпидермального происхождения
1.По строению –гладкие.
2.С железистыми клетками связаны десмосомами.
3.Снаружи покрыты
Мышечная ткань эпидермального происхождения
1.По строению –гладкие.
2.С железистыми клетками связаны десмосомами.
3.Снаружи покрыты

Гладкая мышечная ткань нейрального происхождения (мультиунитарная)
1.Сфинктер и дилататор радужки.
2.Форма веретеновидная.
3.На каждом
Гладкая мышечная ткань нейрального происхождения (мультиунитарная)
1.Сфинктер и дилататор радужки.
2.Форма веретеновидная.
3.На каждом
Сердечная поперечнополосатая мышечная ткань (миокард сердца).
Сердечная поперечнополосатая мышечная ткань (миокард сердца).

Гистогенез сердечной мышечной ткани.
1.Источник развития – миоэпикардиальная пластинка (мезодерма висцерального листка
Гистогенез сердечной мышечной ткани.
1.Источник развития – миоэпикардиальная пластинка (мезодерма висцерального листка
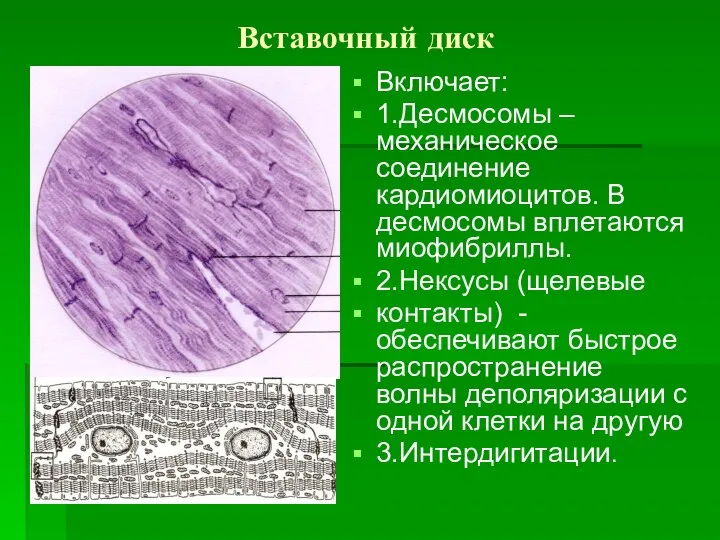
Вставочный диск
Включает:
1.Десмосомы –механическое соединение кардиомиоцитов. В десмосомы вплетаются миофибриллы.
2.Нексусы (щелевые
контакты) -обеспечивают
Вставочный диск
Включает:
1.Десмосомы –механическое соединение кардиомиоцитов. В десмосомы вплетаются миофибриллы.
2.Нексусы (щелевые
контакты) -обеспечивают

Типы кардиомиоцитов
1.Сократительные (типичные, рабочие).
2.Секретрные.
3.Проводящие (атипичные).
Типы кардиомиоцитов
1.Сократительные (типичные, рабочие).
2.Секретрные.
3.Проводящие (атипичные).

Сократительные кардиомиоциты
1.Форма призматическая или отростчатая.
2.Одно или два ядра в центре клетки.
3.Миофибриллы
Сократительные кардиомиоциты
1.Форма призматическая или отростчатая.
2.Одно или два ядра в центре клетки.
3.Миофибриллы

Секреторные кардиомиоциты
1.Располагаются в предсердиях.
2.Форма отростчатачя.
3.Слабое развитие сократительного аппарата.
4.Хорошо развит синтетический аппарат.
5.
Секреторные кардиомиоциты
1.Располагаются в предсердиях.
2.Форма отростчатачя.
3.Слабое развитие сократительного аппарата.
4.Хорошо развит синтетический аппарат.
5.

Энергетический. синтетический , лизосомальный аппараты.
См.выше - скелетная мышечная ткань
Энергетический. синтетический , лизосомальный аппараты.
См.выше - скелетная мышечная ткань

Проводящие кардиомиоциты
1.Крупнее, чем сократительные.
2.Богаче цитоплазмой.
3.Мало миофибрилл, поэтому при окраске гематоксилином и
Проводящие кардиомиоциты
1.Крупнее, чем сократительные.
2.Богаче цитоплазмой.
3.Мало миофибрилл, поэтому при окраске гематоксилином и

Типы проводящих кардиомиоцитов
1.Р-клетки (клетки пейсмекеры):
-располагаются в центре синусного узла и немного
Типы проводящих кардиомиоцитов
1.Р-клетки (клетки пейсмекеры):
-располагаются в центре синусного узла и немного

продолжение
3.Клетки Пуркинье:
-самые крупные не только в проводящей системе, но и во
продолжение
3.Клетки Пуркинье:
-самые крупные не только в проводящей системе, но и во

Регенерация сердечной мышечной ткани
1.На субмикроскопическом уровне – обновление всех структур клеток.
2.В
Регенерация сердечной мышечной ткани
1.На субмикроскопическом уровне – обновление всех структур клеток.
2.В

Кровоснабжение сердечной мышечной ткани
1.По уровню кровоснабжения превосходит головной мозг, уступая лишь
Кровоснабжение сердечной мышечной ткани
1.По уровню кровоснабжения превосходит головной мозг, уступая лишь

Иннервация сердечной мышечной ткани
1.Осущестляется нервными волокнами симпатического и парасимпатического отделов вегетативной
Иннервация сердечной мышечной ткани
1.Осущестляется нервными волокнами симпатического и парасимпатического отделов вегетативной
 Физиология нервной ткани
Физиология нервной ткани  Снегири на ветке
Снегири на ветке Аминокислоты и белки растений
Аминокислоты и белки растений Проект Мои домашние животные
Проект Мои домашние животные Общие пути катаболизма
Общие пути катаболизма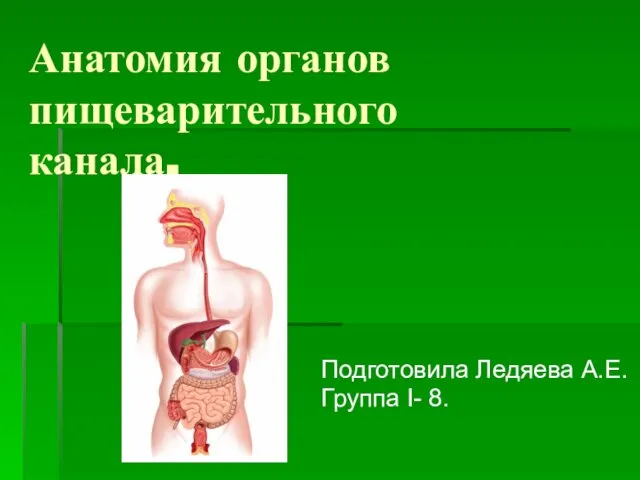 Анатомия органов пищеварительного канала
Анатомия органов пищеварительного канала Отделы: Хвощевидные и Плауновидные. Вымершие папоротникообразные
Отделы: Хвощевидные и Плауновидные. Вымершие папоротникообразные Чарльз Дарвин. Жизнь и биография
Чарльз Дарвин. Жизнь и биография Творческое название проекта: Полезен или вреден шоколад?
Творческое название проекта: Полезен или вреден шоколад? Презентация на тему "Устройство увеличительных приборов" - скачать презентации по Биологии
Презентация на тему "Устройство увеличительных приборов" - скачать презентации по Биологии Российский Государственный Университет им. А.И. Герцена. В.Т.Кудрявцев. Воображение ребенка: природа и развитие // Психологический
Российский Государственный Университет им. А.И. Герцена. В.Т.Кудрявцев. Воображение ребенка: природа и развитие // Психологический  Презентация на тему "Формула здоровья" - скачать презентации по Биологии
Презентация на тему "Формула здоровья" - скачать презентации по Биологии Презентация на тему Царство Грибов
Презентация на тему Царство Грибов  Презентация на тему "Жан Батист Ламарк" - скачать бесплатно презентации по Биологии
Презентация на тему "Жан Батист Ламарк" - скачать бесплатно презентации по Биологии Лекарственные растения
Лекарственные растения Фізіологія вегетативної нервової системи (ВНС)
Фізіологія вегетативної нервової системи (ВНС) Задачи: Изучить особенности строения, функции и гигиену кожи
Задачи: Изучить особенности строения, функции и гигиену кожи Животные Австралии
Животные Австралии Серцевий цикл. Робота серця Презентацію підготувала вчитель біології Черкаської спеціалізованої школи №33 ім. В.Симоненка Гр
Серцевий цикл. Робота серця Презентацію підготувала вчитель біології Черкаської спеціалізованої школи №33 ім. В.Симоненка Гр МИКРОЭЛЕМЕНТЫ И ИХ РОЛЬ В ОРГАНИЗМЕ ЧЕЛОВЕКА
МИКРОЭЛЕМЕНТЫ И ИХ РОЛЬ В ОРГАНИЗМЕ ЧЕЛОВЕКА Гормоны гипофиза. (Лекция 12)
Гормоны гипофиза. (Лекция 12) Презентация на тему "Где растут экзотические фрукты" - скачать бесплатно презентации по Биологии
Презентация на тему "Где растут экзотические фрукты" - скачать бесплатно презентации по Биологии Ферменты. Классификация ферментов. Особенности строения и свойств ферментов
Ферменты. Классификация ферментов. Особенности строения и свойств ферментов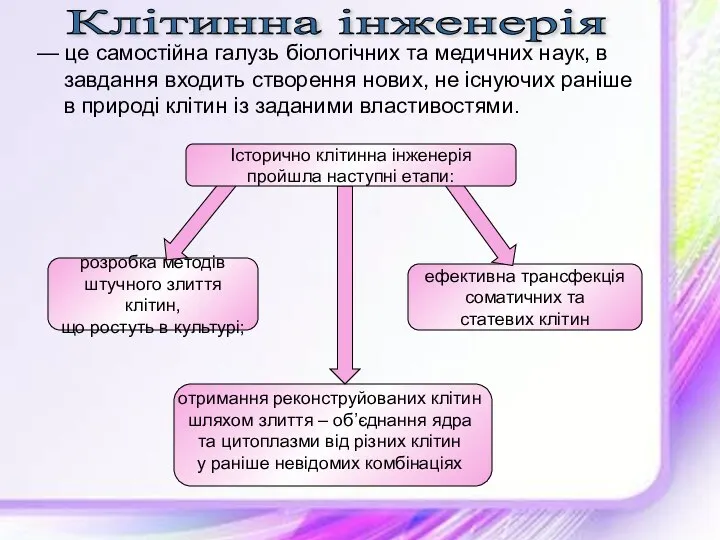 — це самостійна галузь біологічних та медичних наук, в завдання входить створення нових, не існуючих раніше в природі клітин із
— це самостійна галузь біологічних та медичних наук, в завдання входить створення нових, не існуючих раніше в природі клітин із Как помочь птицам
Как помочь птицам Ядовитая эволюция Составила учитель биологии Логунцова Ю.В. МОУ « Средняя школа №14» г.Кимры Тверской области.
Ядовитая эволюция Составила учитель биологии Логунцова Ю.В. МОУ « Средняя школа №14» г.Кимры Тверской области. Как питаются паразиты?
Как питаются паразиты? Подкласс Однодольные
Подкласс Однодольные