- Na-K-АТФаза

Содержание
- 2. Ионные насосы (помпы) – интегральные белки, которые обеспечивают активный перенос ионов против градиента концентрации. Энергией для
- 3. Активный транспорт веществ через биологические мембраны имеет огромное значение. За счет активного транспорта в организме создаются
- 4. Суммарный поток ионов через мембрану должен был бы отсутствовать. Его наличие свидетельствовало бы о переносе ионов
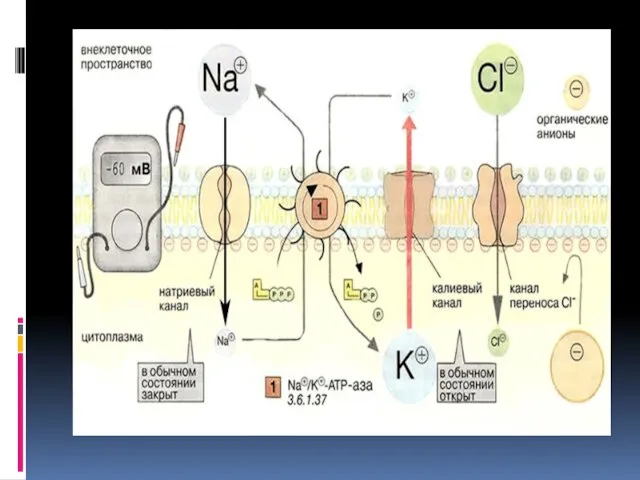
- 5. Na/K-АТФаза - фермент клеточной мембраны животных тканей, который избирательно выкачивает из клетки ионы натрия и аккумулирует
- 6. Na/K-АТФаза представляет собой сложный белок, встроенный в наружную мембрану клетки и имеющий центры связывания для ионов
- 8. Активный транспорт ионов против их концентрационного градиента зависит от наличия в клетке АТФ. В нейронах мозга,
- 9. Та часть бета-субъединицы, которая обращена во внеклеточную среду, несет на себе ковалентно присоединенные углеводные фрагменты. По
- 10. ПОЧЕМУ НАТРИЙ И КАЛИЙ БЫЛИ ВЫБРАНЫ ПРИРОДОЙ ДЛЯ ОТЛИЧИЯ ЖИВОГО ОТ НЕЖИВОГО Одно из неотъемлемых свойств
- 11. Поскольку гидратированный ион натрия близок по размерам к негидратированному иону калия, то ни по заряду, ни
- 13. КАК РАБОТАЕТ Na/K-ATФаза Специфическим механизмом распознавания ионов калия и натрия обладает и Na / K-АТФаза. Впервые
- 14. КАК РАБОТАЕТ Na/K-ATФаза Однако в покоящейся клетке соотношение концентраций ионов натрия и калия противоположно тому, которое
- 15. КАК РАБОТАЕТ Na/K-ATФаза Начинается гидролитический цикл с взаимодействия белка с ионами натрия. "Натриевая" конформация фермента (или
- 16. Позже было установлено, что при гидролизе АТФ действительно наблюдается образование фосфорилированного интермедиата. В настоящее время цикл
- 17. Потенциал покоя Мембранные транспортные структуры — натрий-калиевые насосы — создают предпосылки для возникновения потенциала покоя. Предпосылки
- 18. В норме, когда нервная клетка находится в физиологическом покое и готова к работе, у неё уже
- 19. Макроскопические физические тела, как правило, электрически нейтральны, т.е. в них в равных количествах содержатся как положительные,
- 20. Важно учитывать, что в нервной системе электрические заряды представлены не электронами, как в обычных металлических проводах,
- 21. Электрические силы связаны с химическими по уравнению Гольдмана. Его частным случаем является более простое уравнение Нернста,
- 24. Скачать презентацию

Ионные насосы (помпы) – интегральные белки, которые обеспечивают активный перенос ионов против
Ионные насосы (помпы) – интегральные белки, которые обеспечивают активный перенос ионов против

Активный транспорт веществ через биологические мембраны имеет огромное значение. За счет активного
Активный транспорт веществ через биологические мембраны имеет огромное значение. За счет активного

Суммарный поток ионов через мембрану должен был бы отсутствовать. Его наличие
Суммарный поток ионов через мембрану должен был бы отсутствовать. Его наличие

Na/K-АТФаза - фермент клеточной мембраны животных тканей, который избирательно выкачивает из
Na/K-АТФаза - фермент клеточной мембраны животных тканей, который избирательно выкачивает из

Na/K-АТФаза представляет собой сложный белок, встроенный в наружную мембрану клетки и
Na/K-АТФаза представляет собой сложный белок, встроенный в наружную мембрану клетки и


Активный транспорт ионов против их концентрационного градиента зависит от наличия в
Активный транспорт ионов против их концентрационного градиента зависит от наличия в

Та часть бета-субъединицы, которая обращена во внеклеточную среду, несет на себе
Та часть бета-субъединицы, которая обращена во внеклеточную среду, несет на себе

ПОЧЕМУ НАТРИЙ И КАЛИЙ БЫЛИ ВЫБРАНЫ ПРИРОДОЙ ДЛЯ ОТЛИЧИЯ ЖИВОГО ОТ
ПОЧЕМУ НАТРИЙ И КАЛИЙ БЫЛИ ВЫБРАНЫ ПРИРОДОЙ ДЛЯ ОТЛИЧИЯ ЖИВОГО ОТ

Поскольку гидратированный ион натрия близок по размерам к негидратированному иону калия,
Поскольку гидратированный ион натрия близок по размерам к негидратированному иону калия,


КАК РАБОТАЕТ Na/K-ATФаза
Специфическим механизмом распознавания ионов калия и натрия обладает и
КАК РАБОТАЕТ Na/K-ATФаза
Специфическим механизмом распознавания ионов калия и натрия обладает и

КАК РАБОТАЕТ Na/K-ATФаза
Однако в покоящейся клетке соотношение концентраций ионов натрия и
КАК РАБОТАЕТ Na/K-ATФаза
Однако в покоящейся клетке соотношение концентраций ионов натрия и

КАК РАБОТАЕТ Na/K-ATФаза
Начинается гидролитический цикл с взаимодействия белка с ионами натрия.
КАК РАБОТАЕТ Na/K-ATФаза
Начинается гидролитический цикл с взаимодействия белка с ионами натрия.

Позже было установлено, что при гидролизе АТФ действительно наблюдается образование фосфорилированного
Позже было установлено, что при гидролизе АТФ действительно наблюдается образование фосфорилированного

Потенциал покоя
Мембранные транспортные структуры — натрий-калиевые насосы — создают предпосылки для
Потенциал покоя
Мембранные транспортные структуры — натрий-калиевые насосы — создают предпосылки для

В норме, когда нервная клетка находится в физиологическом покое и готова
В норме, когда нервная клетка находится в физиологическом покое и готова

Макроскопические физические тела, как правило, электрически нейтральны, т.е. в них в
Макроскопические физические тела, как правило, электрически нейтральны, т.е. в них в

Важно учитывать, что в нервной системе электрические заряды представлены не электронами,
Важно учитывать, что в нервной системе электрические заряды представлены не электронами,

Электрические силы связаны с химическими по уравнению Гольдмана. Его частным случаем
Электрические силы связаны с химическими по уравнению Гольдмана. Его частным случаем
 Что такое нейронные сети?
Что такое нейронные сети? Ядро. Деление клетки
Ядро. Деление клетки Система кровообращения
Система кровообращения  «Влияние городской среды на здоровье человека» Дорохина А., Есаян Л., 10 «Б», Руководитель: Черёмухина И.А. ГОУ Гимназия 1526, Ю
«Влияние городской среды на здоровье человека» Дорохина А., Есаян Л., 10 «Б», Руководитель: Черёмухина И.А. ГОУ Гимназия 1526, Ю Силы в природе
Силы в природе Растительный организм и его особенности
Растительный организм и его особенности Посадка Пеперомии
Посадка Пеперомии Презентация на тему "Строение органоидов клетки" - скачать презентации по Биологии
Презентация на тему "Строение органоидов клетки" - скачать презентации по Биологии Гибриды среди животных
Гибриды среди животных Теломеры и теломераза. Структура, функции и пути регуляции активности
Теломеры и теломераза. Структура, функции и пути регуляции активности Популярные виды сухоцветов
Популярные виды сухоцветов Морфофункциональная функция характеристики маточной трубы
Морфофункциональная функция характеристики маточной трубы Интересные факты об экзотических фруктах и овощах
Интересные факты об экзотических фруктах и овощах Презентация на тему "Забавные эти животные" - скачать презентации по Биологии
Презентация на тему "Забавные эти животные" - скачать презентации по Биологии Хромосомная теория наследственности
Хромосомная теория наследственности Онтогенез. Сравнение зародышей позвоночных на разных стадиях эмбрионального развития
Онтогенез. Сравнение зародышей позвоночных на разных стадиях эмбрионального развития Экскурсия в лес. Создатель:Васильева Вита
Экскурсия в лес. Создатель:Васильева Вита Тварини минулого
Тварини минулого Презентация на тему Систематика
Презентация на тему Систематика  Генетика та основні етапи її розвитку. Тема 1
Генетика та основні етапи її розвитку. Тема 1 Явище змочування та капілярності в побуті,природі та техніці
Явище змочування та капілярності в побуті,природі та техніці Мозг необъятен как Вселенная
Мозг необъятен как Вселенная Цистолитиаз у лошади
Цистолитиаз у лошади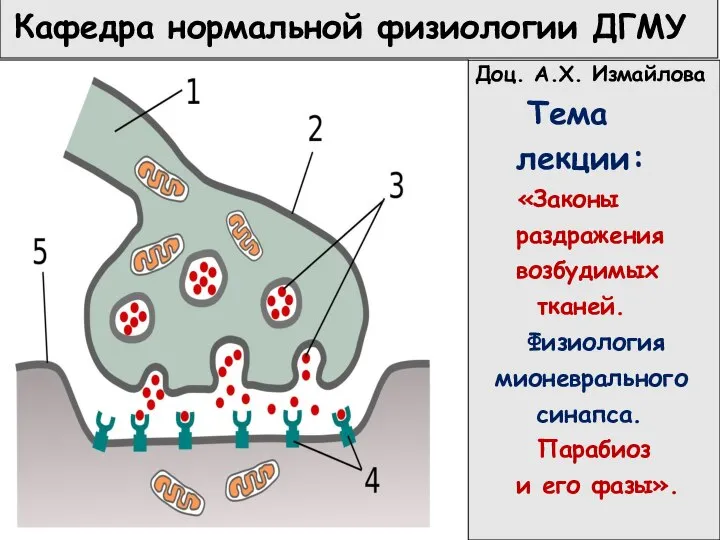 Законы раздражения возбудимых тканей. Физиология мионеврального синапса
Законы раздражения возбудимых тканей. Физиология мионеврального синапса Пищеварительная система человека. Тонкая кишка, печень. Поджелудочная железа. Селезенка
Пищеварительная система человека. Тонкая кишка, печень. Поджелудочная железа. Селезенка Царство Животные. Тип Губки. Тип Кишечнополостные
Царство Животные. Тип Губки. Тип Кишечнополостные Обмен веществ и энергии в клетке
Обмен веществ и энергии в клетке Волк - санитар леса
Волк - санитар леса