- Нервные волокна

Содержание
- 2. Миелинизированное волокно Для понимания механизмов проведения по нервным волокнам возбуждения необходимо учитывать как электрические, так и
- 3. Рис. 1-83. Схема строения миелинизированного нервного волокна (А), шванновской клетки (Б) и структура перехвата Ранвье (В)
- 4. Шванновская клетка Миелин представляет собой упаковку листков специфических плазматических мембран, продуцируемых глиальными клетками, которые обертываются вокруг
- 5. Рис. 1-84. Рост мембраны шванновской клетки и формирование миелиновой оболочки аксона. А - рост мембраны шванновской
- 6. Распространение по аксону пассивного электротонического потенциала Пассивный электротонический потенциал распространяется по нервному волокну на небольшие расстояния,
- 7. Рис. 1-85. Распространение пассивного электротонического потенциала на примере немиелинизированного волокна. А - аксон в состоянии покоя.
- 8. Распространение потенциала действия Распространение возбуждения в форме потенциала действия принципиально отличается от распространения пассивного электротонического потенциала.
- 9. Рис. 1-86. Распространение потенциала действия на примере немиелинизированного волокна. А - аксон в состоянии покоя. Потенциалуправляемые
- 10. Механизм распространения потенциала действия по немиелинизированному волокну Рис. 1-87. Механизм проведения потенциала действия в немиелинизированном волокне.
- 11. В зонах (2), находящихся в состоянии покоя, локальные круговые токи электротонически сдвигают потенциал покоя в сторону
- 12. Механизм распространения потенциала действия по миелинизированному волокну Рис. 1-88. Механизм проведения потенциала действия в миелинизированном волокне.
- 13. Раздражение перехвата Ранвье (0) нервного волокна приводит к открытию потенциалуправляемых Na+-ионных каналов, что ведет к возникновению
- 14. Проведение возбуждения по двум типам волокон Распространение возбуждения в немиелинизированном и миелинизированном нервном волокне суммарно и
- 15. Рис. 1-89. Сравнение общих принципов проведения возбуждения по немиелинизированному и миелинизированному нервному волокну
- 16. Внеклеточные потенциалы В качестве предварительного усилителя обычно применяют усилитель постоянного тока с дифференциальным входом (рис. 1-90
- 17. Рис. 1-90. Потенциалы нервного волокна и их регистрация. А - дифференциальный усилитель для внеклеточной регистрации потенциалов
- 18. Законы проведения возбуждения Существует несколько необходимых условий проведения возбуждения по нерву, названных «законами» проведения возбуждения по
- 20. Скачать презентацию

Миелинизированное волокно
Для понимания механизмов проведения по нервным волокнам возбуждения необходимо учитывать
Миелинизированное волокно
Для понимания механизмов проведения по нервным волокнам возбуждения необходимо учитывать

Рис. 1-83. Схема строения миелинизированного нервного волокна (А), шванновской клетки (Б)
Рис. 1-83. Схема строения миелинизированного нервного волокна (А), шванновской клетки (Б)

Шванновская клетка
Миелин представляет собой упаковку листков специфических плазматических мембран, продуцируемых глиальными
Шванновская клетка
Миелин представляет собой упаковку листков специфических плазматических мембран, продуцируемых глиальными
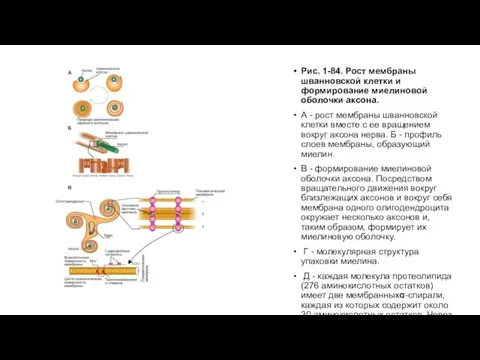
Рис. 1-84. Рост мембраны шванновской клетки и формирование миелиновой оболочки аксона.
А
Рис. 1-84. Рост мембраны шванновской клетки и формирование миелиновой оболочки аксона.
А

Распространение по аксону пассивного электротонического потенциала
Пассивный электротонический потенциал распространяется по нервному
Распространение по аксону пассивного электротонического потенциала
Пассивный электротонический потенциал распространяется по нервному

Рис. 1-85. Распространение пассивного электротонического потенциала на примере немиелинизированного волокна.
А -
Рис. 1-85. Распространение пассивного электротонического потенциала на примере немиелинизированного волокна.
А -

Распространение потенциала действия
Распространение возбуждения в форме потенциала действия принципиально отличается от
Распространение потенциала действия
Распространение возбуждения в форме потенциала действия принципиально отличается от

Рис. 1-86. Распространение потенциала действия на примере немиелинизированного волокна.
А - аксон
Рис. 1-86. Распространение потенциала действия на примере немиелинизированного волокна.
А - аксон

Механизм распространения потенциала действия по немиелинизированному волокну
Рис. 1-87. Механизм проведения потенциала
Механизм распространения потенциала действия по немиелинизированному волокну
Рис. 1-87. Механизм проведения потенциала
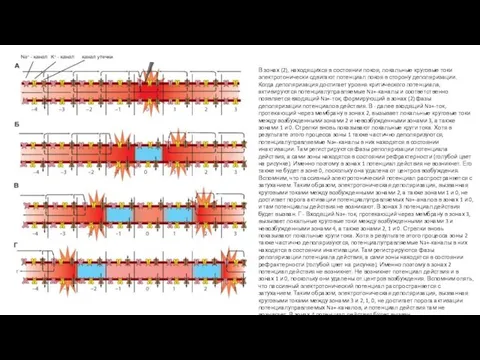
В зонах (2), находящихся в состоянии покоя, локальные круговые токи электротонически
В зонах (2), находящихся в состоянии покоя, локальные круговые токи электротонически

Механизм распространения потенциала действия по миелинизированному волокну
Рис. 1-88. Механизм проведения потенциала
Механизм распространения потенциала действия по миелинизированному волокну
Рис. 1-88. Механизм проведения потенциала

Раздражение перехвата Ранвье (0) нервного волокна приводит к открытию потенциалуправляемых Na+-ионных
Раздражение перехвата Ранвье (0) нервного волокна приводит к открытию потенциалуправляемых Na+-ионных

Проведение возбуждения по двум типам волокон
Распространение возбуждения в немиелинизированном и миелинизированном
Проведение возбуждения по двум типам волокон
Распространение возбуждения в немиелинизированном и миелинизированном

Рис. 1-89. Сравнение общих принципов проведения возбуждения по немиелинизированному и миелинизированному
Рис. 1-89. Сравнение общих принципов проведения возбуждения по немиелинизированному и миелинизированному

Внеклеточные потенциалы
В качестве предварительного усилителя обычно применяют усилитель постоянного тока с
Внеклеточные потенциалы
В качестве предварительного усилителя обычно применяют усилитель постоянного тока с
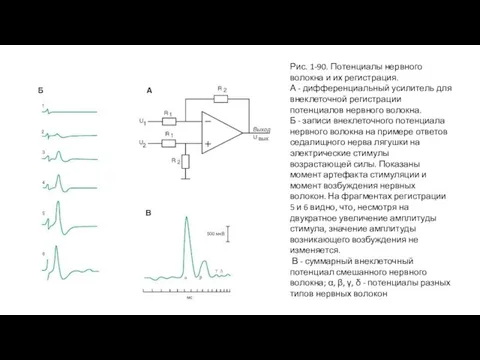
Рис. 1-90. Потенциалы нервного волокна и их регистрация.
А - дифференциальный усилитель
Рис. 1-90. Потенциалы нервного волокна и их регистрация.
А - дифференциальный усилитель

Законы проведения возбуждения
Существует несколько необходимых условий проведения возбуждения по нерву, названных
Законы проведения возбуждения
Существует несколько необходимых условий проведения возбуждения по нерву, названных
 Опорно-двигательная система. Значение опорно-двигательной системы. Строение костей
Опорно-двигательная система. Значение опорно-двигательной системы. Строение костей Влияние распорядка учебного дня на работоспособность и концентрацию и оптимизация временных затрат на учебный процесс
Влияние распорядка учебного дня на работоспособность и концентрацию и оптимизация временных затрат на учебный процесс Мухи: польза и вред. Исследовательский проект
Мухи: польза и вред. Исследовательский проект Презентация на тему Организм побеждает болезни
Презентация на тему Организм побеждает болезни Органическое вещество почвы
Органическое вещество почвы Птицы из красной книги Башкортостана. Региональный компонент на уроках математики в начальной школе
Птицы из красной книги Башкортостана. Региональный компонент на уроках математики в начальной школе Биосинтез заменимых аминокислот. Деградация нуклеиновых кислот. (Лекция 4)
Биосинтез заменимых аминокислот. Деградация нуклеиновых кислот. (Лекция 4) Наследственность, изменчивость
Наследственность, изменчивость Импринтинг. Нарушения импринтинга как причина наследственной патологии. Семинар 7
Импринтинг. Нарушения импринтинга как причина наследственной патологии. Семинар 7 Интеллектуалды ойын
Интеллектуалды ойын Где живут микробы
Где живут микробы Кольчатые черви
Кольчатые черви ВІЛ та СНІД
ВІЛ та СНІД  Как помочь птицам
Как помочь птицам Регуляция дыхания. Транспорт О2 кровью
Регуляция дыхания. Транспорт О2 кровью Заттардың тасымалдануы. Гуморальді және жасушалық иммунитетті салыстыру
Заттардың тасымалдануы. Гуморальді және жасушалық иммунитетті салыстыру Растение подорожник
Растение подорожник Фотосинтез Презентация для 6 класса Проценко Л.В._
Фотосинтез Презентация для 6 класса Проценко Л.В._ Клетка как биологическая система
Клетка как биологическая система Строение головного мозга. Продолговатый мозг. Задний мозг. Лекция № 23
Строение головного мозга. Продолговатый мозг. Задний мозг. Лекция № 23 Гордость и чудо Астраханского края - лотос
Гордость и чудо Астраханского края - лотос Прокариотическая клетка
Прокариотическая клетка Физиология кровообращения
Физиология кровообращения Мускулатура рыб. Физиологические основы движения. Лекция 4
Мускулатура рыб. Физиологические основы движения. Лекция 4 Основные эволюционные преобразования в кровеносной системе хордовых
Основные эволюционные преобразования в кровеносной системе хордовых Презентация на тему Значение, строение и функционирование нервной системы
Презентация на тему Значение, строение и функционирование нервной системы Живая природа - прототип человеческих изобретений
Живая природа - прототип человеческих изобретений Птица удод
Птица удод