- Обмен белков и аминокислот

Содержание
- 2. План лекции Азотистый баланс, биологическая ценность белков пищи Переваривание белков в ЖКТ Всасывание аминокислот в кишечнике
- 3. Суточная потребность в белке За сутки в организме взрослого человека распадается и вновь синтезируется до 400
- 4. Обмен белков и аминокислот
- 5. Азотистый баланс Это разница между количеством азота, поступающего с пищей, и количеством выделяемого азота. Азотистый баланс
- 6. Биологическая ценность белков определяется Аминокислотным составом Соотношением заменимых и незаменимых аминокислот: 6:1 Степенью усвоения т.е. как
- 7. СО2 К+ НСО3- Н+ Сl- Сl- Плазма рН 7,2 Просвет Желудка рН 1,0 – 2,0 Переваривание:
- 8. Переваривание белков в ЖКТ В желудочном соке – пепсиноген→пепсин: 1-е молекулы медленно под действием HCl остальные
- 9. Переваривание белков в ЖКТ Специфичность действия протеаз
- 10. Транспорт аминокислот Активный, Na-зависимый (Симпорт) Транспортеры (специфические переносчики): Нейтральных, с короткими радикалами – АЛА, СЕР, ТРЕ
- 11. Нарушение транспорта аминокислот Болезнь Хартнупа – дефект переносчика нейтральных аминокислот Иминоглицинурия – ПРО, оксипролин, ГЛИ Цистинурия
- 12. Толстый кишечник Брожение, гниение – результат действия кишечных бактерий: образуются газы СН4, СО2, Н2S, уксусная, молочная,
- 13. Внутриклеточное переваривание Лизосомы – 40 гидролаз Эндоцитоз, фагоцитоз Катепсины – тканевые ферменты
- 14. Обмен аминокислот Пути поступления аминокислот: а) транспорт из внеклеточной жидкости (при всасывании пищевых аминокислот); б) синтез
- 15. Трансаминирование –перенос аминогруппы с аминокислоты на α-кетокислоту. При этом количество аминокислот не меняется, т.к. образуются новая
- 16. Трансаминирование Аминокислота NH3 NH3 α-кетокислота Орнитиновый цикл Пируват ацетил-КоА мочевина Глюкоза Липиды, Кетоновые тела экскреция С
- 17. Аланинаминотрансфераза (АЛТ)
- 18. Коферментами аминотрансфераз являются производные витамина В6 (пиридоксина)
- 19. Механизм трансаминирования – образование шиффовых оснований (1стадия) Субстрат 1 - аминокислота Продукт 1 - кетокислота Н2О
- 20. Дезаминирование - отщепление аминогруппы с образованием аммиака Существует четыре типа реакций: 1. Восстановительное дезаминирование ( +2H+)
- 21. Реакции дезаминирования подвергаются все аминокислоты кроме ЛИЗ Окислительное (прямое дезаминирование) – фермент ГЛУТАМАТДЕГИДРОГЕНАЗА, ММ 312 кD,
- 22. Непрямое (трансдезаминирование) – основной путь катаболизма аминокислот I этап – трансаминирование с α-кетоглутаратом с образованием ГЛУ
- 23. Основные источники NH3 Аминокислоты, биогенные амины, нуклеотиды NH3 транспортируется из тканей в печень в виде 3
- 24. Симптомы гипераммониемии: головокружение, тошнота, рвота, судороги, потеря сознания, отек мозга Основной диагностический признак – повышение концентрации
- 25. Механизм токсического действия аммиака NH3 легко проникает через мембраны в клетки В митохондриях – гипоэнергетическое состояние,
- 26. Лечение больных с гипераммониемией направлено на снижение концентрации NH3 Малобелковая диета Введение в организм с пищей
- 27. В обезвреживании NH3 – центральная роль принадлежит ГЛУ Основной реакцией обезвреживания NH3 в мышцах, мозге и
- 28. Глюкозо-аланиновый цикл – это образование АЛА в мышцах, его перенос в печень и перенос глюкозы, синтезированной
- 29. Основной путь обезвреживания NH3 в ЦНС – синтез глутамина под действием глутаминсинтазы Источники NH3 : АМФ,
- 30. В кишечнике глутамат подвергается трансаминированию с пируватом Глутамин Глутамат Пируват Фекалии α-кетогглутарат Аланин Печень Мочевина
- 31. В почках происходит гидролиз глутамина под действием глутаминазы, активность которой возрастает при ацидозе Н+ NH4А Глутамин
- 32. Орнитиновый цикл (только в печени) Обезвреживание NH3 Синтез АРГИНИНА Молекула мочевины содержит 2 атома N: 1
- 33. Орнитиновый цикл Реакции орнитинового цикла протекают в цитоплазме. 1-я реакция протекает в матриксе митохондрий: синтез карбамоилфосфата
- 34. Орнитиновый цикл Аммиак пересекает внутреннюю митохондриальную мембрану в составе аминокислоты цитруллин. Реакция катализируется ферментом орнитин-транскарбамоил-синтетазой (ОТ-к):
- 35. Орнитиновый цикл
- 36. Орнитиновый цикл
- 37. Декарбоксилирование аминокислот – отщепление α -карбоксильной группы аминокислот с образованием аминов Реакция катализируется декарбоксилазами, коферментом которых
- 38. Биогенные амины Серотонин – вазоконстриктор, сокращает гладкую мускулатуру, антидепрессант Мелатонин регулирует суточные и сезонные изменения метаболизма
- 39. Биогенные амины
- 40. Биогенные амины Ацетилхолин – нейромедиатор вегетативной нервной системы донор метильных групп – SAM серин→этаноамин→холин
- 41. Основные метаболические превращения фенилаланина и тирозина Блокирование реакций при фенилкетонурии (1-фенилаланингидроксилаза), тирозинозе (2-п-гидрокси-фенилпируватдиоксигеназа), альбинизме (3) и
- 42. Фенилкетонурия Наследственное заболевание – мутация в гене фенилаланингидроксилазы – в крови накапливаются фенилаланин, фенилпируват, фениллактат, фенилацетат,
- 43. Синтез катехоламинов в надпочечниках а – Тирозингидроксилаза (О2, Н4БП, НАДФН) б – ДОФА-декарбоксилаза (ПФ) в- Дофамингидроксилаза
- 44. В щитовидной железе синтезируются йодтиронины
- 45. Обмен серосодержащих аминокислот
- 46. Cинтез цистина
- 47. Cинтез S-аденозилметионина (SAM)
- 48. Перенос одноуглеродных групп Реакция, в которой переносится метильная группа (-СН3), называется реакцией метилирования. Донором метильной группы
- 50. Скачать презентацию

План лекции
Азотистый баланс, биологическая ценность белков пищи
Переваривание белков в ЖКТ
Всасывание аминокислот
План лекции
Азотистый баланс, биологическая ценность белков пищи
Переваривание белков в ЖКТ
Всасывание аминокислот

Суточная потребность в белке
За сутки в организме взрослого человека распадается
Суточная потребность в белке
За сутки в организме взрослого человека распадается
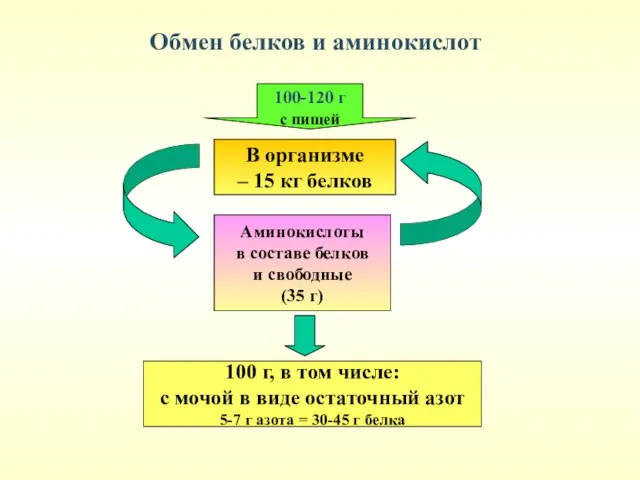
Обмен белков и аминокислот
Обмен белков и аминокислот

Азотистый баланс
Это разница между количеством азота, поступающего с пищей, и количеством
Азотистый баланс
Это разница между количеством азота, поступающего с пищей, и количеством

Биологическая ценность белков определяется
Аминокислотным составом
Соотношением заменимых и незаменимых аминокислот: 6:1
Степенью
Биологическая ценность белков определяется
Аминокислотным составом
Соотношением заменимых и незаменимых аминокислот: 6:1
Степенью
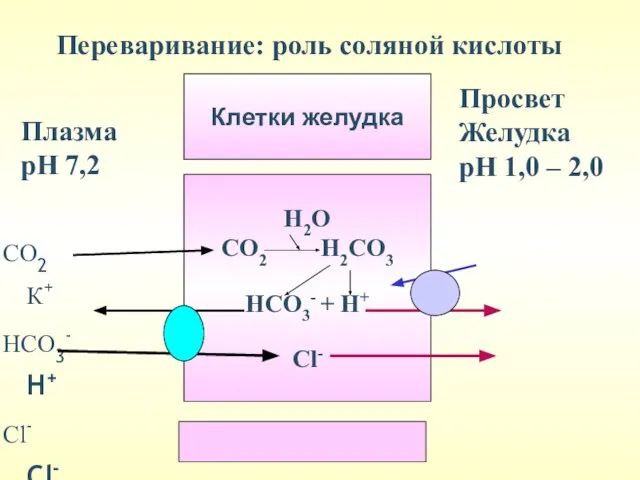
СО2 К+
НСО3- Н+
Сl- Сl-
Плазма
рН 7,2
Просвет
Желудка
рН 1,0 – 2,0
Переваривание: роль соляной
НСО3- Н+
Сl- Сl-
Плазма
рН 7,2
Просвет
Желудка
рН 1,0 – 2,0
Переваривание: роль соляной

Переваривание белков в ЖКТ
В желудочном соке – пепсиноген→пепсин: 1-е молекулы медленно
Переваривание белков в ЖКТ
В желудочном соке – пепсиноген→пепсин: 1-е молекулы медленно
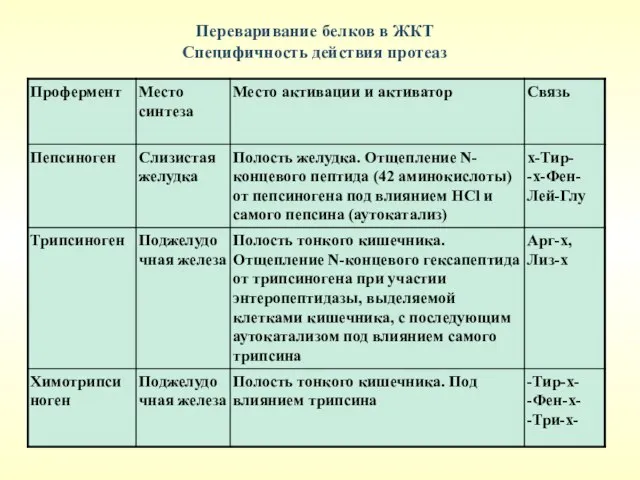
Переваривание белков в ЖКТ
Специфичность действия протеаз
Переваривание белков в ЖКТ
Специфичность действия протеаз

Транспорт аминокислот
Активный, Na-зависимый (Симпорт)
Транспортеры (специфические переносчики):
Нейтральных, с короткими радикалами –
Транспорт аминокислот
Активный, Na-зависимый (Симпорт)
Транспортеры (специфические переносчики):
Нейтральных, с короткими радикалами –

Нарушение транспорта аминокислот
Болезнь Хартнупа – дефект переносчика нейтральных аминокислот
Иминоглицинурия – ПРО,
Нарушение транспорта аминокислот
Болезнь Хартнупа – дефект переносчика нейтральных аминокислот
Иминоглицинурия – ПРО,

Толстый кишечник
Брожение, гниение – результат действия кишечных бактерий: образуются газы СН4,
Толстый кишечник
Брожение, гниение – результат действия кишечных бактерий: образуются газы СН4,
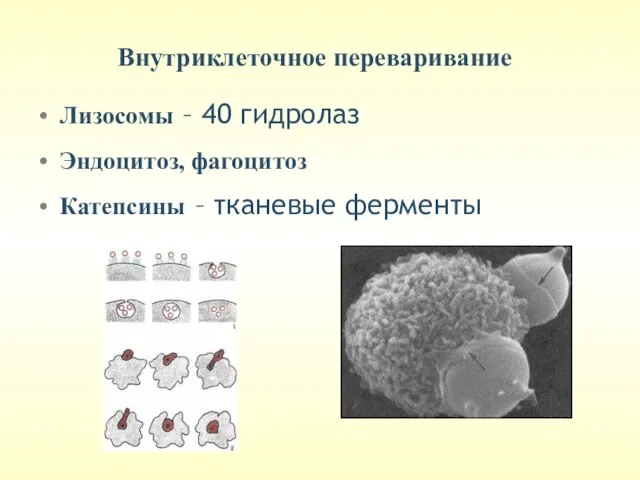
Внутриклеточное переваривание
Лизосомы – 40 гидролаз
Эндоцитоз, фагоцитоз
Катепсины – тканевые ферменты
Внутриклеточное переваривание
Лизосомы – 40 гидролаз
Эндоцитоз, фагоцитоз
Катепсины – тканевые ферменты

Обмен аминокислот
Пути поступления аминокислот:
а) транспорт из внеклеточной жидкости (при всасывании пищевых
Обмен аминокислот
Пути поступления аминокислот: а) транспорт из внеклеточной жидкости (при всасывании пищевых

Трансаминирование –перенос аминогруппы с аминокислоты на α-кетокислоту. При этом количество аминокислот
Трансаминирование –перенос аминогруппы с аминокислоты на α-кетокислоту. При этом количество аминокислот
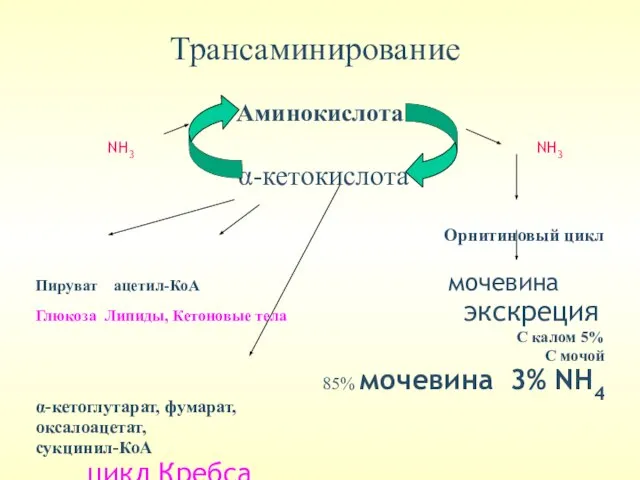
Трансаминирование
Аминокислота
NH3 NH3
α-кетокислота
Орнитиновый цикл
Пируват ацетил-КоА мочевина
Глюкоза Липиды, Кетоновые
Трансаминирование
Аминокислота
NH3 NH3
α-кетокислота
Орнитиновый цикл
Пируват ацетил-КоА мочевина
Глюкоза Липиды, Кетоновые
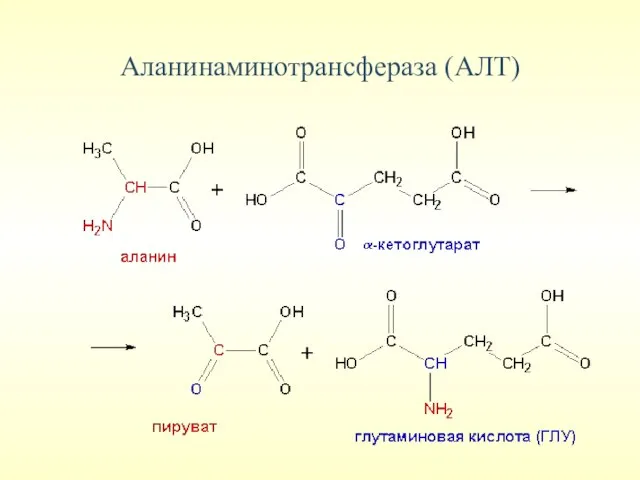
Аланинаминотрансфераза (АЛТ)
Аланинаминотрансфераза (АЛТ)
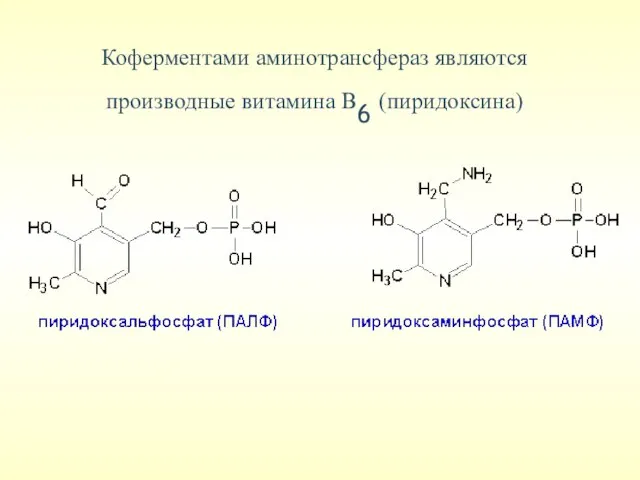
Коферментами аминотрансфераз являются производные витамина В6 (пиридоксина)
Коферментами аминотрансфераз являются производные витамина В6 (пиридоксина)
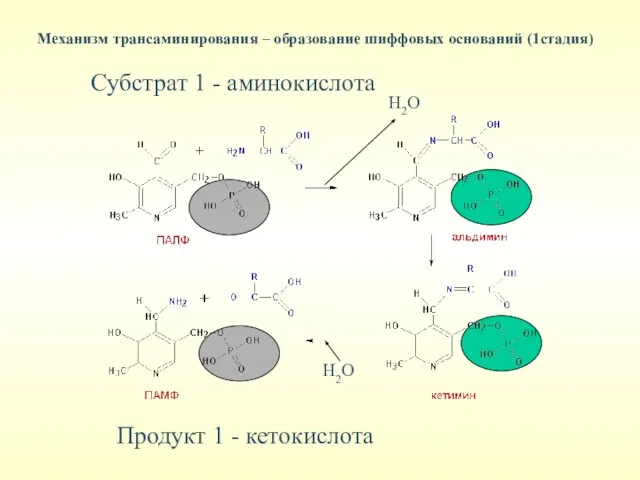
Механизм трансаминирования – образование шиффовых оснований (1стадия)
Субстрат 1 - аминокислота
Продукт 1
Механизм трансаминирования – образование шиффовых оснований (1стадия)
Субстрат 1 - аминокислота
Продукт 1

Дезаминирование - отщепление аминогруппы с образованием аммиака
Существует четыре типа реакций:
1. Восстановительное
Дезаминирование - отщепление аминогруппы с образованием аммиака
Существует четыре типа реакций:
1. Восстановительное

Реакции дезаминирования
подвергаются все аминокислоты кроме ЛИЗ
Окислительное (прямое дезаминирование) – фермент
Реакции дезаминирования
подвергаются все аминокислоты кроме ЛИЗ
Окислительное (прямое дезаминирование) – фермент

Непрямое (трансдезаминирование) – основной путь катаболизма аминокислот
I этап – трансаминирование
с
Непрямое (трансдезаминирование) – основной путь катаболизма аминокислот
I этап – трансаминирование
с

Основные источники NH3
Аминокислоты, биогенные амины, нуклеотиды
NH3 транспортируется из тканей в
Основные источники NH3
Аминокислоты, биогенные амины, нуклеотиды
NH3 транспортируется из тканей в

Симптомы гипераммониемии:
головокружение, тошнота, рвота, судороги, потеря сознания, отек мозга
Основной диагностический признак
Симптомы гипераммониемии:
головокружение, тошнота, рвота, судороги, потеря сознания, отек мозга
Основной диагностический признак

Механизм токсического действия аммиака
NH3 легко проникает через мембраны в клетки
В митохондриях
Механизм токсического действия аммиака
NH3 легко проникает через мембраны в клетки
В митохондриях

Лечение больных с гипераммониемией направлено на снижение концентрации NH3
Малобелковая диета
Введение
Лечение больных с гипераммониемией направлено на снижение концентрации NH3
Малобелковая диета
Введение

В обезвреживании NH3 – центральная роль принадлежит ГЛУ
Основной реакцией обезвреживания NH3
В обезвреживании NH3 – центральная роль принадлежит ГЛУ
Основной реакцией обезвреживания NH3
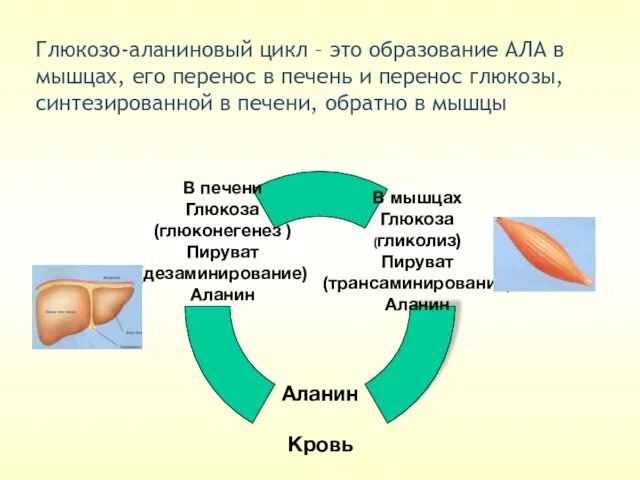
Глюкозо-аланиновый цикл – это образование АЛА в мышцах, его перенос в
Глюкозо-аланиновый цикл – это образование АЛА в мышцах, его перенос в

Основной путь обезвреживания NH3 в ЦНС – синтез глутамина под действием
Основной путь обезвреживания NH3 в ЦНС – синтез глутамина под действием
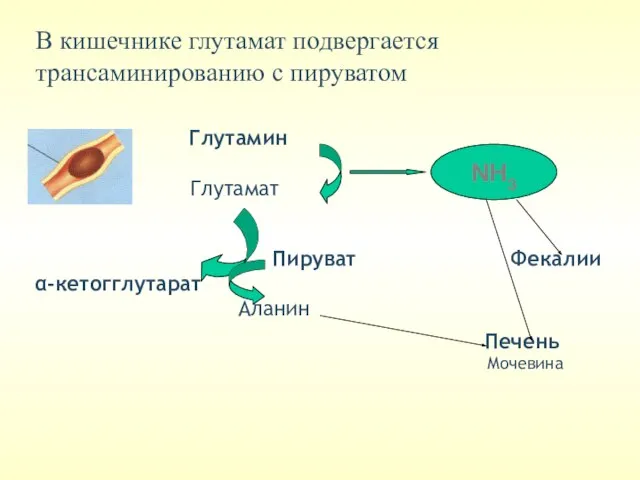
В кишечнике глутамат подвергается трансаминированию с пируватом
Глутамин
Глутамат
Пируват Фекалии
α-кетогглутарат
Аланин
Печень
В кишечнике глутамат подвергается трансаминированию с пируватом
Глутамин
Глутамат
Пируват Фекалии
α-кетогглутарат
Аланин
Печень
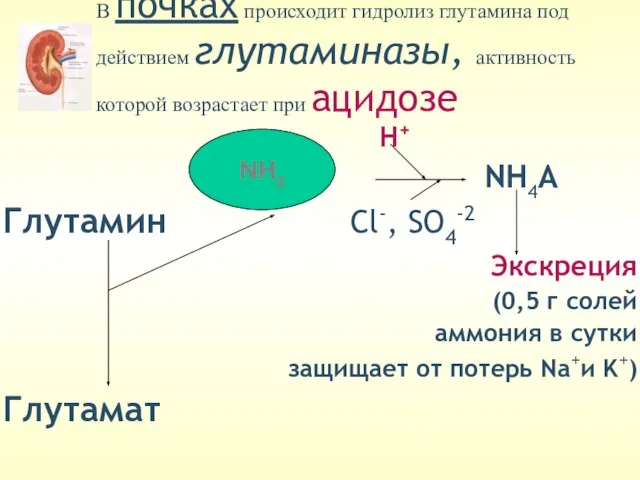
В почках происходит гидролиз глутамина под действием глутаминазы, активность которой возрастает
В почках происходит гидролиз глутамина под действием глутаминазы, активность которой возрастает

Орнитиновый цикл
(только в печени)
Обезвреживание NH3
Синтез АРГИНИНА
Молекула мочевины содержит 2 атома
Орнитиновый цикл
(только в печени)
Обезвреживание NH3
Синтез АРГИНИНА
Молекула мочевины содержит 2 атома

Орнитиновый цикл
Реакции орнитинового цикла протекают в цитоплазме.
1-я реакция протекает в
Орнитиновый цикл
Реакции орнитинового цикла протекают в цитоплазме.
1-я реакция протекает в
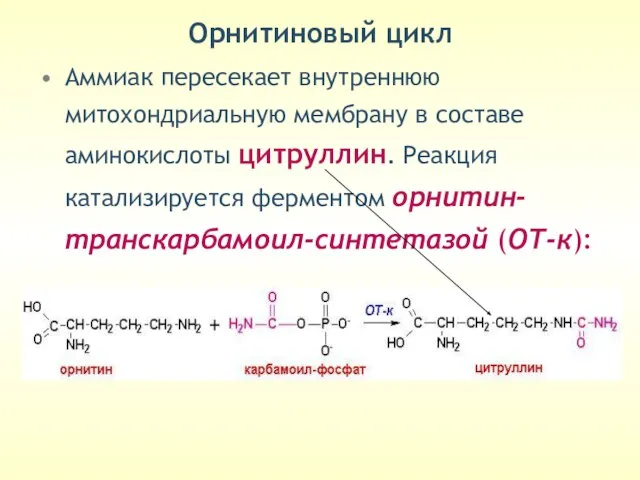
Орнитиновый цикл
Аммиак пересекает внутреннюю митохондриальную мембрану в составе аминокислоты цитруллин. Реакция
Орнитиновый цикл
Аммиак пересекает внутреннюю митохондриальную мембрану в составе аминокислоты цитруллин. Реакция
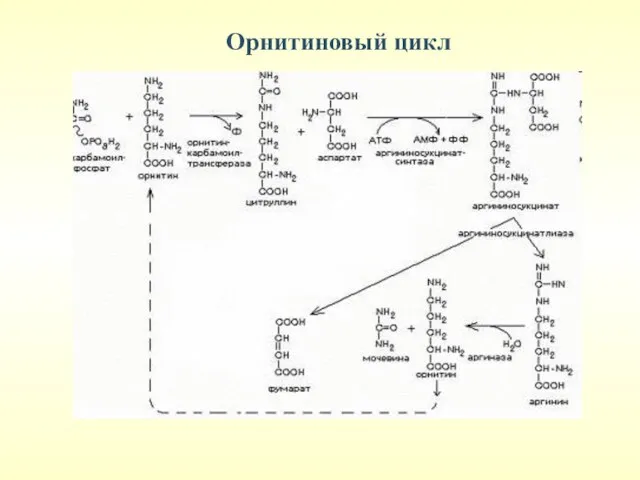
Орнитиновый цикл
Орнитиновый цикл
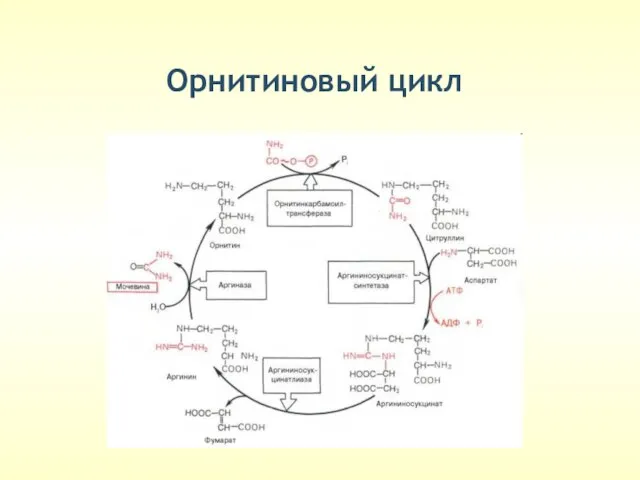
Орнитиновый цикл
Орнитиновый цикл

Декарбоксилирование аминокислот – отщепление α -карбоксильной группы аминокислот с образованием аминов
Реакция
Декарбоксилирование аминокислот – отщепление α -карбоксильной группы аминокислот с образованием аминов
Реакция
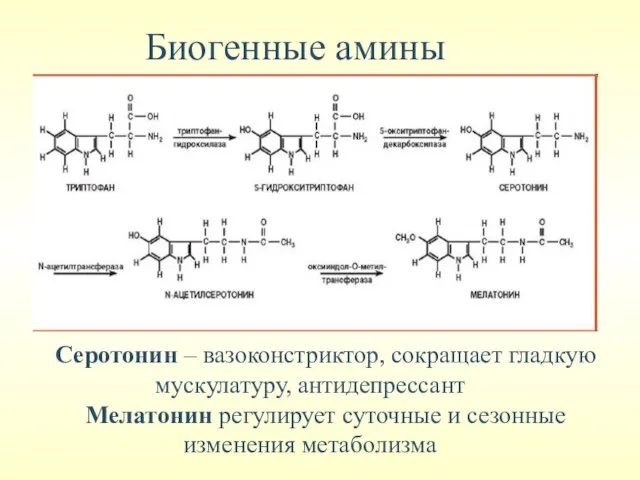
Биогенные амины
Серотонин – вазоконстриктор, сокращает гладкую мускулатуру, антидепрессант
Мелатонин регулирует суточные и
Биогенные амины
Серотонин – вазоконстриктор, сокращает гладкую мускулатуру, антидепрессант
Мелатонин регулирует суточные и
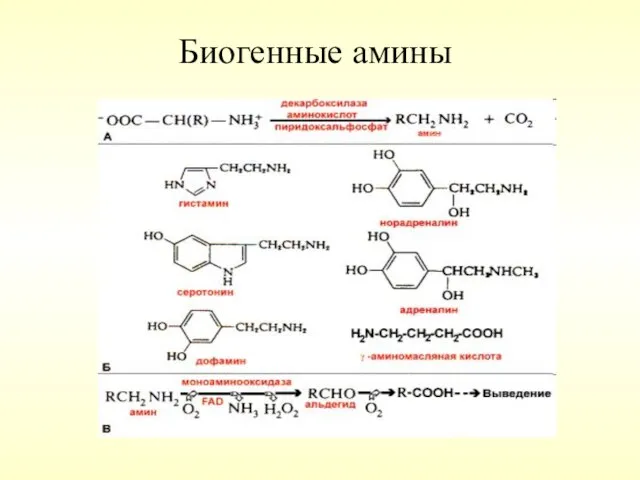
Биогенные амины
Биогенные амины
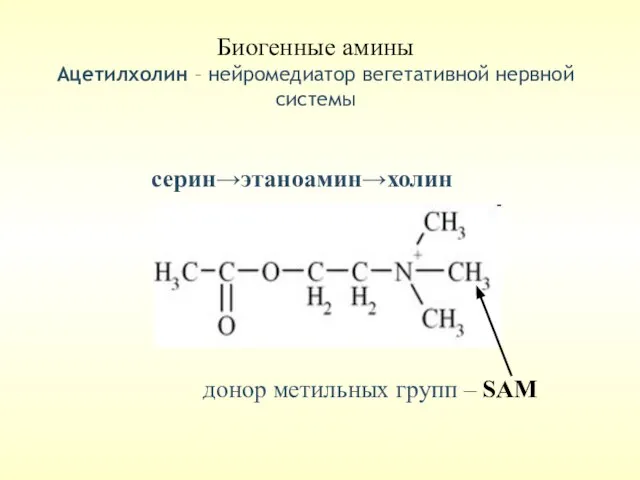
Биогенные амины
Ацетилхолин – нейромедиатор вегетативной нервной системы
донор метильных групп –
Биогенные амины
Ацетилхолин – нейромедиатор вегетативной нервной системы
донор метильных групп –
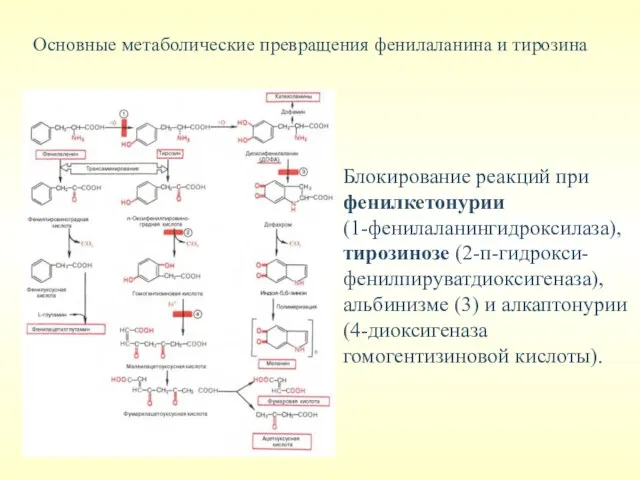
Основные метаболические превращения фенилаланина и тирозина
Блокирование реакций при фенилкетонурии
(1-фенилаланингидроксилаза),
Основные метаболические превращения фенилаланина и тирозина
Блокирование реакций при фенилкетонурии
(1-фенилаланингидроксилаза),

Фенилкетонурия
Наследственное заболевание – мутация в гене фенилаланингидроксилазы – в крови накапливаются
Фенилкетонурия
Наследственное заболевание – мутация в гене фенилаланингидроксилазы – в крови накапливаются
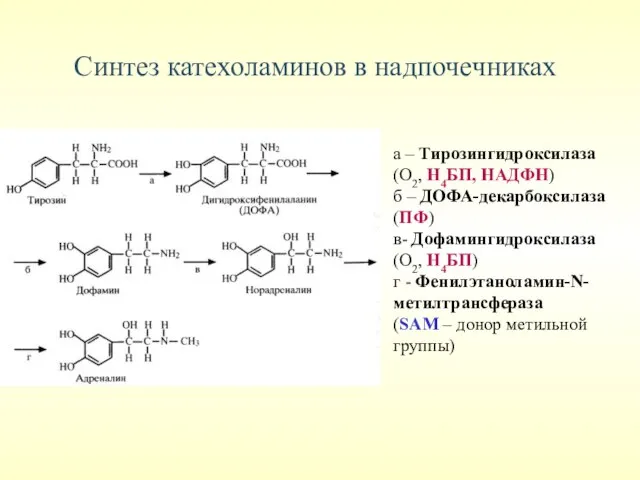
Синтез катехоламинов в надпочечниках
а – Тирозингидроксилаза
(О2, Н4БП, НАДФН)
б – ДОФА-декарбоксилаза
(ПФ)
в- Дофамингидроксилаза
(О2,
Синтез катехоламинов в надпочечниках
а – Тирозингидроксилаза
(О2, Н4БП, НАДФН)
б – ДОФА-декарбоксилаза
(ПФ)
в- Дофамингидроксилаза
(О2,
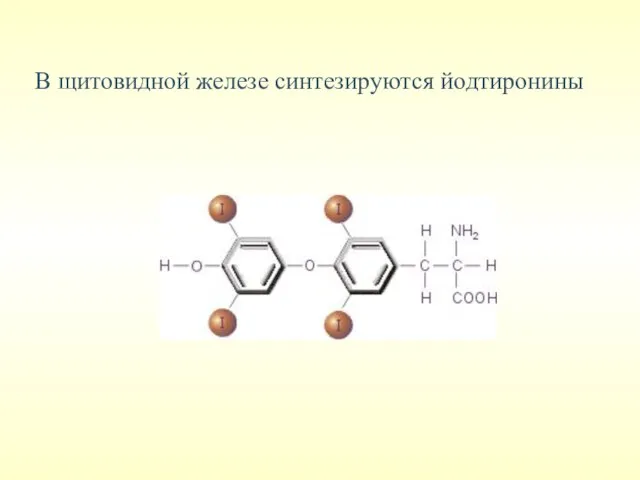
В щитовидной железе синтезируются йодтиронины
В щитовидной железе синтезируются йодтиронины
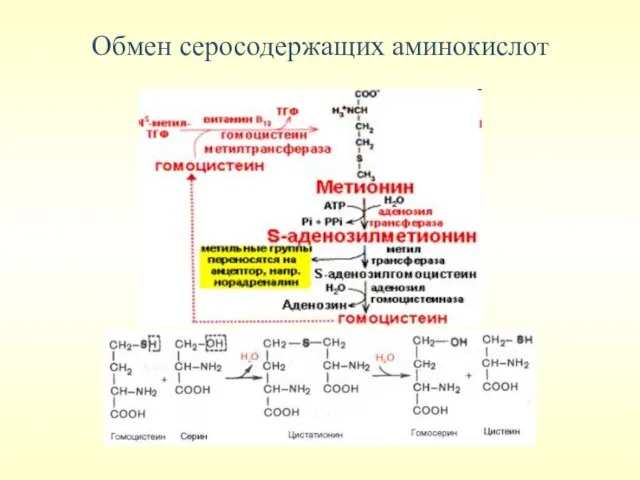
Обмен серосодержащих аминокислот
Обмен серосодержащих аминокислот
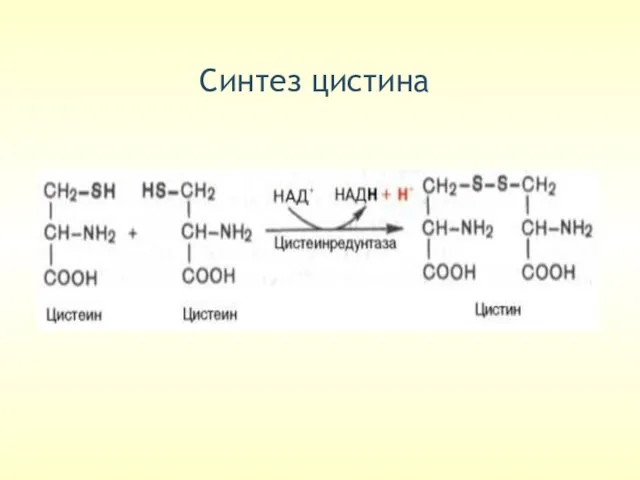
Cинтез цистина
Cинтез цистина
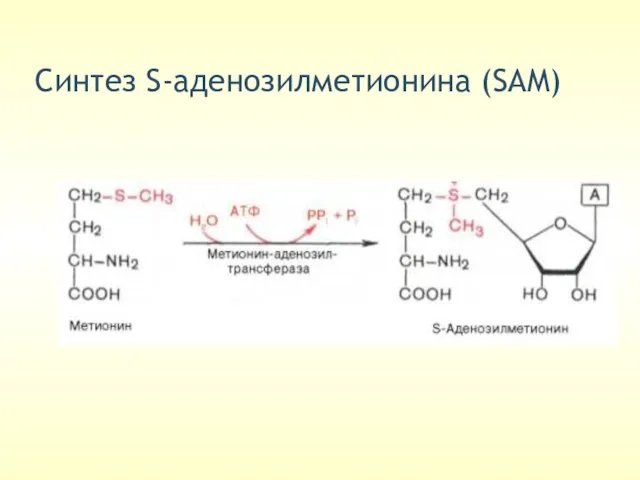
Cинтез S-аденозилметионина (SAM)
Cинтез S-аденозилметионина (SAM)
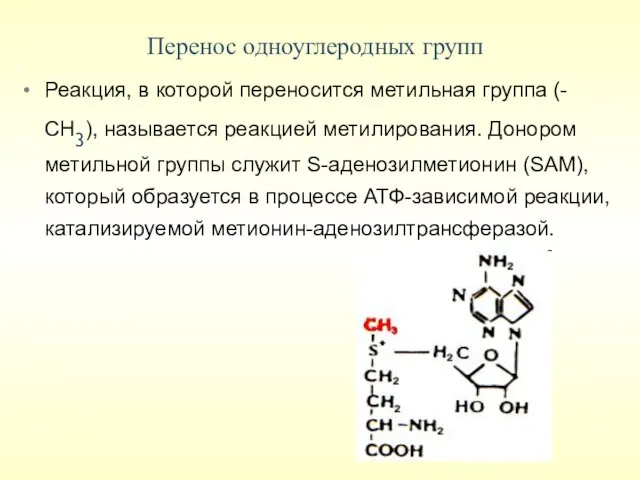
Перенос одноуглеродных групп
Реакция, в которой переносится метильная группа (-СН3), называется реакцией
Перенос одноуглеродных групп
Реакция, в которой переносится метильная группа (-СН3), называется реакцией
 Изменения в жизни животных осенью
Изменения в жизни животных осенью Плесневые грибы и дрожжи
Плесневые грибы и дрожжи Презентация на тему ГЕНЕТИЧЕСКИЕ ОСНОВЫ СЕЛЕКЦИИ ОРГАНИЗМОВ
Презентация на тему ГЕНЕТИЧЕСКИЕ ОСНОВЫ СЕЛЕКЦИИ ОРГАНИЗМОВ  Подцарство Одноклеточные или Простейшие Вводная характеристика простейших. Тип саркожгутиковых.
Подцарство Одноклеточные или Простейшие Вводная характеристика простейших. Тип саркожгутиковых.  Интересные факты о генетике
Интересные факты о генетике Среды жизни планета Земля
Среды жизни планета Земля Пищеварительная система
Пищеварительная система Презентация на тему "Мифы о курении и реальность" - презентации по Биологии
Презентация на тему "Мифы о курении и реальность" - презентации по Биологии Свойства живого. Живые системы
Свойства живого. Живые системы Промысловые рыбы
Промысловые рыбы Интегрированный урок «Процесс дыхания. Газообмен в лёгких»
Интегрированный урок «Процесс дыхания. Газообмен в лёгких»  Полость живота. Полость брюшины. Отношение органов брюшине. Забрюшинное пространство. Производные брюшины (занятие 4)
Полость живота. Полость брюшины. Отношение органов брюшине. Забрюшинное пространство. Производные брюшины (занятие 4) Класс Костные рыбы
Класс Костные рыбы Factors of evolution
Factors of evolution Органы цветковых растений
Органы цветковых растений Колострум (colostrum). Уникальные компоненты колострума
Колострум (colostrum). Уникальные компоненты колострума Презентация Дарвинский заповедник
Презентация Дарвинский заповедник Анатомия и физиология, как науки
Анатомия и физиология, как науки Сосна сибирская кедровая. Исследование условий роста на территории Вологодской области
Сосна сибирская кедровая. Исследование условий роста на территории Вологодской области Общие вопросы анатомии и физиологии сенсорных систем
Общие вопросы анатомии и физиологии сенсорных систем Осень в природе. Дары осени
Осень в природе. Дары осени ТКАНИ И ОРГАНЫ ЧЕЛОВЕКА Урок по биологии 8 класс Учитель Жданова О. В., МБОУ СОШ № 25 г.Липецк
ТКАНИ И ОРГАНЫ ЧЕЛОВЕКА Урок по биологии 8 класс Учитель Жданова О. В., МБОУ СОШ № 25 г.Липецк  Реакции глюконеогенеза
Реакции глюконеогенеза Класс Пресмыкающиеся
Класс Пресмыкающиеся Зона мішаних лісів. Українське Полісся
Зона мішаних лісів. Українське Полісся Самые лучшие сторожевые и охранные собаки Сторожевые собаки – лучшие охранники дома и территории. Сторожевыми называют собак,
Самые лучшие сторожевые и охранные собаки Сторожевые собаки – лучшие охранники дома и территории. Сторожевыми называют собак, Гипотеза стационарного состояния жизни
Гипотеза стационарного состояния жизни Эволюционная теория Дарвина
Эволюционная теория Дарвина