Основы молекулярной генетики. Генетические механизмы. Биосинтез белков и нуклеиновых кислот. (Лекция 3)
- Основы молекулярной генетики. Генетические механизмы. Биосинтез белков и нуклеиновых кислот. (Лекция 3)

Содержание
- 2. Основной постулат молекулярной биологии и генетики: ДНК ------> РНК-------> белок Основными генетическими механизмами являются процессы: репликация
- 3. 1. Репликация (от ДНК к ДНК) 2. Транскрипция (от ДНК к РНК) 3. Трансляция (от РНК
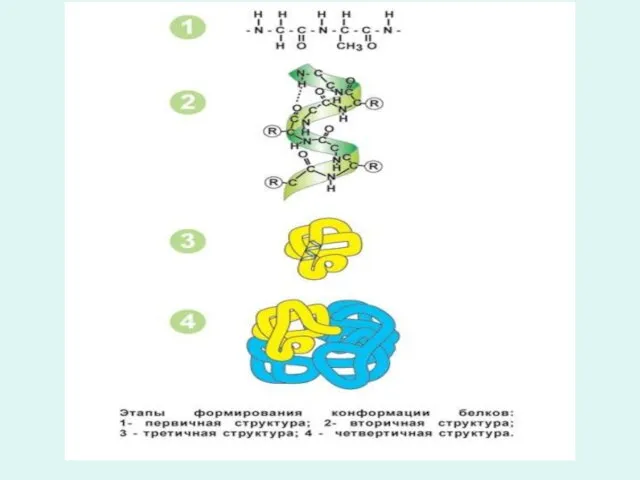
- 4. Синтез белка - это многоступенчатый энергозависимый процесс, который связан с функцией хромосом ядра клеток и функцией
- 5. Молекула ДНК за счет остатков фосфорной кислоты заряжается отрицательно и присоединяет к своей поверхности по всей
- 6. Хромосомы
- 7. Структурная организация хромосом В хромосоме спираль ДНК соединяется группами из восьми молекул белка гистона и образует
- 8. Нуклеосома
- 11. Перед клеточным делением все элементы клетки удваиваются, в том числе и хромосомы (процесс репликации). Из полюсов
- 12. Микротрубочки, примерно так, как строится кирпичная труба, строятся из белка тубулина Молекулы тубулина складываются один на
- 14. Микротрубочки начинают тянуть хромосомы к полюсам веретена. Это сигнал к действию фермента, который аккуратно разрезает пару
- 15. В 1932 году Нобелевский лауреат Герман Мёллер обратил внимание на особое поведение концевых участков хромосом, которые
- 16. Постепенное укорочение ДНК хромосом во время репликации является одной из теорий "старения" клеток (А.М. Оловников,1971) Л.
- 17. Причины укорочения теломеров хромосом Для деления клетки необходимо, чтобы перед этим произошло удвоение хромосом. Ферменты ведущие
- 18. Теломеры Прогрессивное укорочение теломер является счетно-ограничительным механизмом митотических циклов и играет роль часов, отсчитывающих число делений
- 19. Теломераза и бессмертие клетки В организме здорового человека есть клетки, которые могут делиться бесконечное количество раз
- 20. Особенности структуры нуклеиновых кислот Нуклеотиды ДНК и РНК
- 21. Нуклеиновые кислоты любого типа (ДНК, РНК) состоят из мономеров, называемые нуклеотидами. Полинуклеотидные цепи, приобретая соответствующую пространственную
- 22. МОНОНУКЛЕОТИДЫ РНК
- 23. Строение молекулы ДНК В молекуле ДНК полинуклеотидные цепи построены из нуклеотидов :d-АМФ, d-ГМФ, d-ЦМФ, d-ТМФ, т.е.
- 24. Молекула ДНК - две комплиментарные полинуклеотидные цепи
- 26. Правила комплиментарности Чаргафа В молекуле ДНК: 1. Количество аденина равно тимину, гуанина равно цитозину ( А=Т,
- 27. Структура генетического кода Кодовым элементом в полинуклеотидной цепи ДНК, определяющим включение соответствующей аминокислоты в полипептидную цепь,
- 28. Триплетный код
- 29. ЧТО ТАКОЕ ГЕН? Под термином ген (цистрон) следует понимать участок ДНК, в котором в триплетной последовательности
- 30. Строение РНК РНК в отличие от ДНК бывает по большей части одноцепочечной. Формы РНК - транспортная
- 31. Строение мРНК Матричная (мРНК) или информационная РНК (иРНК) составляет 3-5% всей содержащейся в клетке РНК. Это
- 32. Рибосомная РНК Рибосомная РНК, составляющая более 80% всей РНК клетки кодируется особыми генами находящимися в нескольких
- 33. Транспортная РНК (тРНК) На долю тРНК приходится примерно 15% всей клеточной РНК. У тРНК самая короткая
- 35. Структурные участки тРНК В тРНК имеются следующие структурные участки: 1.Акцепторный участок - представлен триплетом ЦЦА. Гидроксил
- 36. МАТРИЧНЫЙ СИНТЕЗ ДНК Процесс самовоспроизведения ДНК называется репликацией (редупликация). Удвоение ДНК происходит вследствие того, что цепи
- 37. Молекулярные основы репликации и репарации ДНК Для репликации ДНК необходимо: 1 - наличие структурного материала для
- 38. 4 - Ферменты: - Расплетающие белки и ДНК-хеликаза - «Затравочная» ДНК-зависимая РНК-полимераза ( ДНК-праймаза), - ДНК-полимеразы
- 39. Известно, что молекула ДНК состоит из двух комплиментарных и антипараллельно направленых полинуклеотидных цепей Каждая полинуклеотидная цепь
- 40. Иногда ДНК-полимеразы могут давать "задний ход", то есть двигаться в направлении 5' 3'. В том случае,
- 41. Синтез ДНК (репликация) Суммарно процесс синтеза ДНК можно представить следующей схемой: m(dАТФ+dТТФ)+ n(dГТФ+dЦТФ)------>ДНК + (m+n)Н4Р2О7 Важнейшими
- 42. Дезокси-АТФ
- 45. Для того чтобы раскрутить двойную спираль ДНК необходимы белки-ферменты ДНК-хеликазы. ДНК-хеликазы быстро движутся вдоль цепей ДНК,
- 47. ДНК-полимеразы не могут начинать синтеза ДНК на матрице без затравочного полинуклеотидного фрагмента, а способны только добавлять
- 48. Удаление крайних РНК-праймеров, комплементарных 3'-концам обеих цепей линейной материнской молекулы ДНК, приводит к тому, что дочерние
- 49. Синтез ДНК Под воздействием расплетающих белков (ДНК-хеликаза и дестабилизирующие белки) происходит разрыв водородных связей между комплиментарными
- 50. Далее к праймеру с помощью ДНК-полимеразы 3 присоединяются соответствующие дезоксирибонуклеозиды в направлении 5` 3` В результате
- 51. Синтез дочерней полинуклеотидной цепи вдоль другой полинуклеотидной цепи матричной ДНК, ввиду ее антинаправленности (3` 5`) и
- 52. Эта реакция идет в две стадии: 1.ДНК-лигазы реагирует с НАД, которая служит донатором АМФ. При этом
- 55. Репарация ошибок репликации В ходе репликации самопроизвольно или под воздействием различных внешних факторов (радиация, ультрафиолетовое излучение,
- 56. Ошибки репликации Ошибки репликации, возникающие во время синтеза полинуклеотидных цепей дочерних молекул ДНК, могут исправляться ДНК-полимеразой
- 57. Транскрипция . Функциональная организация оперона. Согласно современным представлениям (Жакоб, Моно, 1965) гены молекулы ДНК, принимающие участие
- 58. ОПЕРОН Информативные участки оперона (экзоны) представлены структурными генами или цистронами, в триплетной последовательности которых закодирована информация
- 59. Функциональная организация о п е р о н а - транскриптона ( Жакоб, Моно, 1965).
- 60. Функции генов оперона Ген промотор - с него начинается транскципция. К нему присоединяются белки, запускающие транскрипцию
- 61. Механизм транскрипции (биосинтез РНК). Транскрипция идет в три фазы: фаза инициации, фаза элонгации и фаза терминации.
- 62. В фазу инициации первой, исходной реакцией синтеза РНК является реакция присоединения 5`-3` фосфоэфирной связью к АТФ
- 63. Фаза элонгации Затем наступает фаза элонгации - наращивание полинуклеотидной цепи РНК. В результате перемещения РНК-полимеразы вдоль
- 64. Терминация Терминация (завершение) транскрипции происходит после достижения РНК-полимеразой терминирующих кодонов, являющихся стоп-сигналами. Одновременно, специальный белок -
- 65. Первичный транскрипт -пре-РНК Первичные продукты транскрипции являются полными копиями структурных генов ДНК. При этом в нем
- 66. Процессинг В ходе процессинга удаляются неинформативные участки в пре-РНК и образуются функционально зрелые молекулы РНК. Процессинг
- 68. Молекулярные основы трансляции В процессе трансляции можно выделить два этапа, которые имеют разную локализацию в клетке:
- 69. Механизм рекогниции Эти ферменты (аминоацил -тРНК- синтетазы ) катализируют реакции активации аминокислот с образованием аминоацил-аденилата (а)
- 70. Биосинтез белка и факторы трансляции Далее тРНК путем простой диффузии переносят присоединенную к ним аминокислоту к
- 71. Рибосомы Рибосомы являются субклеточными образованиями, состоящими из двух субъединиц - большой и малой, отличающиеся молекулярной массой
- 72. Сборка рибосом и фазы трансляции Появление в клетке факторов инициации синтеза белка, мРНК, met-тРНК приводит к
- 73. Фаза инициации: Биосинтез полипептидной цепи белковой молекулы начинается с появлением в цитозоле мРНК, которая в присутствии
- 74. Формирование Р-(пептидильного) и А-(аминоацильного) центров рибосом После образования комплекса met-тРНК -мРНК -малая субъдиница рибосом -ГТФ, фактор
- 75. Фаза элонгации: В присутствии фактора элонгации ЕF1 и за счет энергии гидролиза ГТФ в свободный А-центр
- 76. Фаза терминации: Синтез полипептидной цепи продолжается до тех пор, пока на пути рибосом не встретится один
- 77. Синтез белка
- 78. Посттрансляционные изменения В результате трансляции не всегда сразу образуется функционально активный белок, хотя с формированием третичной
- 80. Ингибиторы матричных биосинтезов Токсины выделяемые патогенными бактериами, например, дифтерийный токсин, блокирует транслокацию рибосом, прекращая трансляцию, что
- 81. Антибиотики как ингибиторы матричных синтезов Антибиотики, взаимодействующие с ДНК и нарушающие ее матричную функцию, подавляющие репликацию
- 83. Синтез белка
- 84. Регуляция биосинтеза белка Регуляция синтеза белка в системе оперона идет в двух направлениях: 1.Положительный регуляторный контроль
- 85. ОПЕРОН Информативные участки оперона (экзоны) представлены структурными генами или цистронами, в триплетной последовательности которых закодирована информация
- 86. Функциональная организация о п е р о н а - транскриптона ( Жакоб, Моно, 1965).
- 87. Функции генов оперона Ген промотор - с него начинается транскципция. К нему присоединяются белки, запускающие транскрипцию
- 88. Регуляторные гены Кроме собственно регуляторных генов в системе оперона обнаружены целый ряд других генов, принимающих участие
- 89. Протоонкогены Протоонкогены имеются во всех нормальных клетках. В связи с очень большой схожестью со структурой вирусных
- 90. Мутации протоонкогенов Протоонкогены находятся под тщательным и жестким контролем других генов. Мутации протоонкогенов выводят их из-под
- 91. Факторы роста Факторы роста известны как белки, индуцирующие синтез ДНКФакторы роста известны как белки, индуцирующие синтез
- 92. Большинство полипептидных факторов роста действует одновременно по паракринномуБольшинство полипептидных факторов роста действует одновременно по паракринному и
- 93. В регуляторных белках, обладающих интракринным действием, имеются сигнальные последовательности, обеспечивающие внутриклеточную локализацию. До сих пор очень
- 94. Последние исследования показали, женщины страдающие раком молочной железы, у которых выявлена опухоль, экспрессирующая рецепторы эпидермального фактора
- 95. Установлено, вырабатываемый в организме человека фактор роста нейрегулин-1 способен эффективно защитить нервные клетки от разрушительных последствий
- 96. Современные препараты, применяемые при инсульте, эффективны только том случае, если больной начал принимать их в течение
- 97. Опухолевые вирусы и онкогены Некоторые опухолевые вирусы не содержат онкоген, но, встраиваясь в хромосому рядом с
- 98. Превращение клеточных протоонкогенов Превращение клеточных протоонкогенов в онкогены может происходить в результате и в результате повышения
- 99. Онкогенные вирусы Установлено, что гены и даже целые участки хромосом высших организмов могут иногда перемещаться с
- 100. Ретровирусы Как показали исследования, во многих зрелых онкогенных РНК-содержащих вирусах (ретровирусы) и в том числе в
- 101. Онкогены По фенотипическим проявлениям различают две группы онкогенов. Одна группа - ядерные (иммортелизующие) онкогены, приводящие к
- 102. Канцерогенез Некоторые опухолевые вирусы не содержат онкоген, но, встраиваясь в хромосому рядом с протоонкогеном, активируют его,
- 103. Протеин р53 В последние годы найдено еще одно, по-видимому, наиболее общее звено канцерогенеза - гены-супрессоры опухолей,
- 105. Скачать презентацию

Основной постулат молекулярной биологии и генетики:
ДНК ------> РНК-------> белок
Основными
Основной постулат молекулярной биологии и генетики:
ДНК ------> РНК-------> белок
Основными
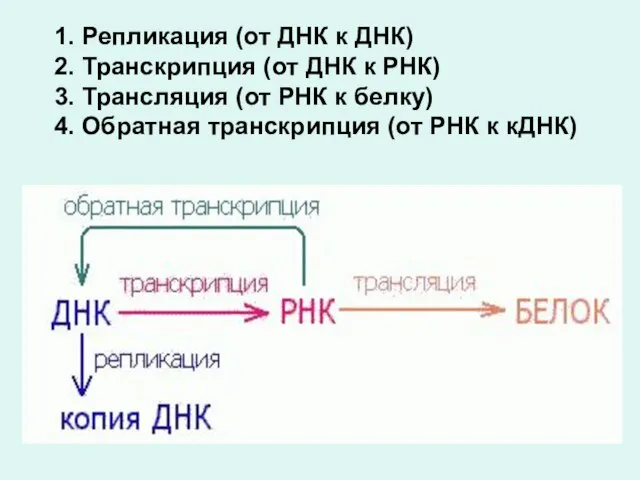
1. Репликация (от ДНК к ДНК)
2. Транскрипция (от ДНК к
1. Репликация (от ДНК к ДНК)
2. Транскрипция (от ДНК к

Синтез белка - это многоступенчатый энергозависимый процесс, который связан с функцией
Синтез белка - это многоступенчатый энергозависимый процесс, который связан с функцией

Молекула ДНК за счет остатков фосфорной кислоты заряжается отрицательно и присоединяет
Молекула ДНК за счет остатков фосфорной кислоты заряжается отрицательно и присоединяет

Хромосомы
Хромосомы

Структурная организация хромосом
В хромосоме спираль ДНК соединяется группами из восьми
Структурная организация хромосом
В хромосоме спираль ДНК соединяется группами из восьми
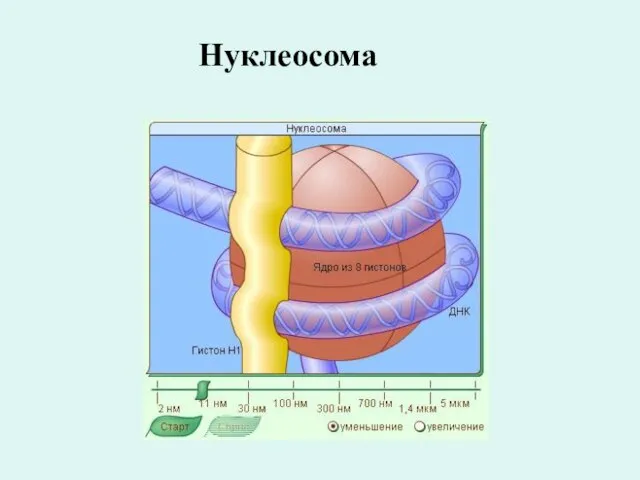
Нуклеосома
Нуклеосома
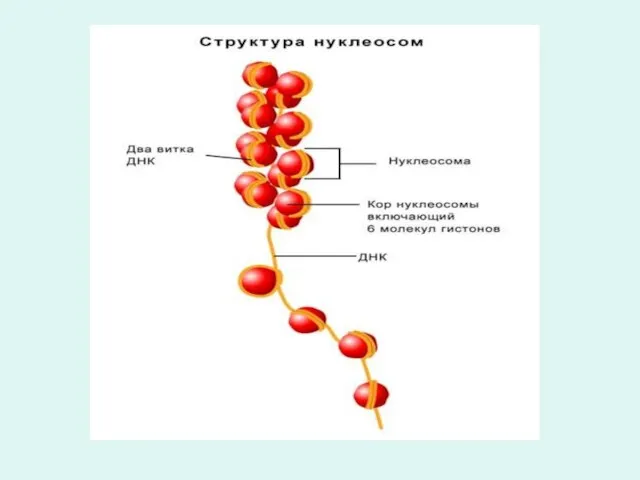

Перед клеточным делением все элементы клетки удваиваются, в том числе и
Перед клеточным делением все элементы клетки удваиваются, в том числе и

Микротрубочки, примерно так, как строится кирпичная труба, строятся из белка тубулина
Молекулы
Микротрубочки, примерно так, как строится кирпичная труба, строятся из белка тубулина
Молекулы
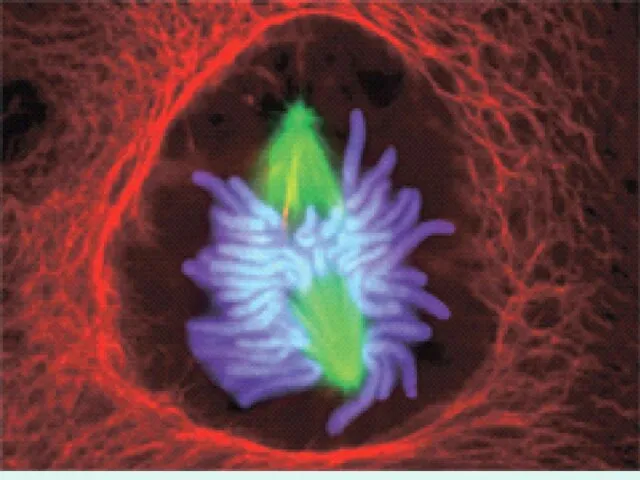

Микротрубочки начинают тянуть хромосомы к полюсам веретена.
Это сигнал к действию фермента, который
Это сигнал к действию фермента, который

В 1932 году Нобелевский лауреат Герман Мёллер обратил внимание на особое
В 1932 году Нобелевский лауреат Герман Мёллер обратил внимание на особое

Постепенное укорочение ДНК хромосом во время репликации является одной из теорий
Постепенное укорочение ДНК хромосом во время репликации является одной из теорий

Причины укорочения теломеров хромосом
Для деления клетки необходимо, чтобы перед этим произошло
Причины укорочения теломеров хромосом
Для деления клетки необходимо, чтобы перед этим произошло

Теломеры
Прогрессивное укорочение теломер является счетно-ограничительным механизмом митотических циклов и играет роль
Теломеры
Прогрессивное укорочение теломер является счетно-ограничительным механизмом митотических циклов и играет роль

Теломераза и бессмертие клетки
В организме здорового человека есть клетки, которые могут
Теломераза и бессмертие клетки
В организме здорового человека есть клетки, которые могут

Особенности структуры нуклеиновых кислот
Нуклеотиды ДНК и РНК
Особенности структуры нуклеиновых кислот
Нуклеотиды ДНК и РНК

Нуклеиновые кислоты любого типа (ДНК, РНК) состоят из мономеров, называемые нуклеотидами.
Нуклеиновые кислоты любого типа (ДНК, РНК) состоят из мономеров, называемые нуклеотидами.
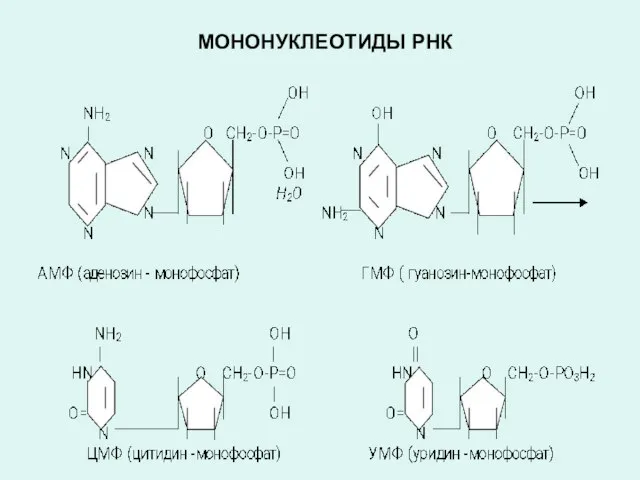
МОНОНУКЛЕОТИДЫ РНК
МОНОНУКЛЕОТИДЫ РНК

Строение молекулы ДНК
В молекуле ДНК полинуклеотидные цепи построены из нуклеотидов :d-АМФ,
Строение молекулы ДНК
В молекуле ДНК полинуклеотидные цепи построены из нуклеотидов :d-АМФ,
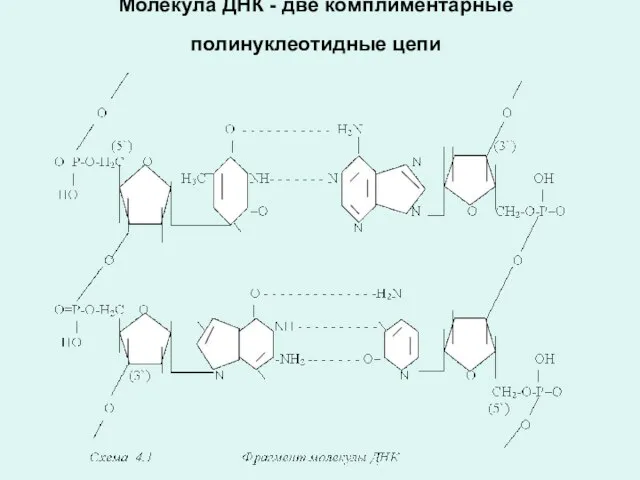
Молекула ДНК - две комплиментарные полинуклеотидные цепи
Молекула ДНК - две комплиментарные полинуклеотидные цепи
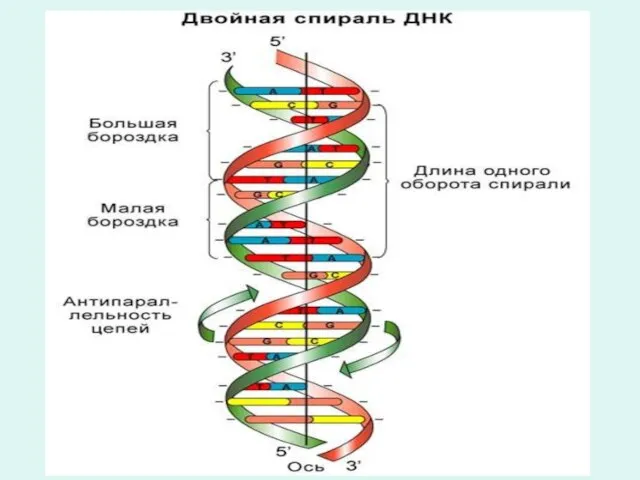

Правила комплиментарности Чаргафа
В молекуле ДНК:
1. Количество аденина равно тимину, гуанина равно
Правила комплиментарности Чаргафа
В молекуле ДНК:
1. Количество аденина равно тимину, гуанина равно

Структура генетического кода
Кодовым элементом в полинуклеотидной цепи ДНК, определяющим включение соответствующей
Структура генетического кода
Кодовым элементом в полинуклеотидной цепи ДНК, определяющим включение соответствующей

Триплетный код
Триплетный код

ЧТО ТАКОЕ ГЕН?
Под термином ген (цистрон) следует понимать участок ДНК, в
ЧТО ТАКОЕ ГЕН?
Под термином ген (цистрон) следует понимать участок ДНК, в

Строение РНК
РНК в отличие от ДНК бывает по большей части одноцепочечной.
Строение РНК
РНК в отличие от ДНК бывает по большей части одноцепочечной.

Строение мРНК
Матричная (мРНК) или информационная РНК (иРНК) составляет 3-5% всей содержащейся
Строение мРНК
Матричная (мРНК) или информационная РНК (иРНК) составляет 3-5% всей содержащейся

Рибосомная РНК
Рибосомная РНК, составляющая более 80% всей РНК клетки кодируется особыми
Рибосомная РНК
Рибосомная РНК, составляющая более 80% всей РНК клетки кодируется особыми

Транспортная РНК (тРНК)
На долю тРНК приходится примерно 15% всей клеточной РНК.
Транспортная РНК (тРНК)
На долю тРНК приходится примерно 15% всей клеточной РНК.


Структурные участки тРНК
В тРНК имеются следующие структурные участки:
1.Акцепторный участок - представлен
Структурные участки тРНК
В тРНК имеются следующие структурные участки:
1.Акцепторный участок - представлен

МАТРИЧНЫЙ СИНТЕЗ ДНК
Процесс самовоспроизведения ДНК называется репликацией (редупликация).
Удвоение ДНК происходит
МАТРИЧНЫЙ СИНТЕЗ ДНК
Процесс самовоспроизведения ДНК называется репликацией (редупликация).
Удвоение ДНК происходит

Молекулярные основы репликации и репарации ДНК
Для репликации ДНК необходимо:
1 - наличие
Молекулярные основы репликации и репарации ДНК
Для репликации ДНК необходимо:
1 - наличие

4 - Ферменты:
- Расплетающие белки и ДНК-хеликаза
- «Затравочная» ДНК-зависимая РНК-полимераза (
4 - Ферменты:
- Расплетающие белки и ДНК-хеликаза
- «Затравочная» ДНК-зависимая РНК-полимераза (

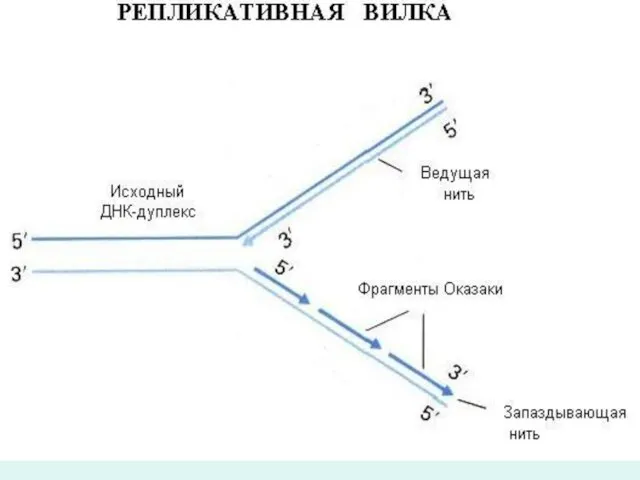
Известно, что молекула ДНК состоит из двух комплиментарных и антипараллельно направленых
Известно, что молекула ДНК состоит из двух комплиментарных и антипараллельно направленых

Иногда ДНК-полимеразы могут давать "задний ход", то есть двигаться в направлении
Иногда ДНК-полимеразы могут давать "задний ход", то есть двигаться в направлении

Синтез ДНК (репликация)
Суммарно процесс синтеза ДНК можно представить следующей схемой:
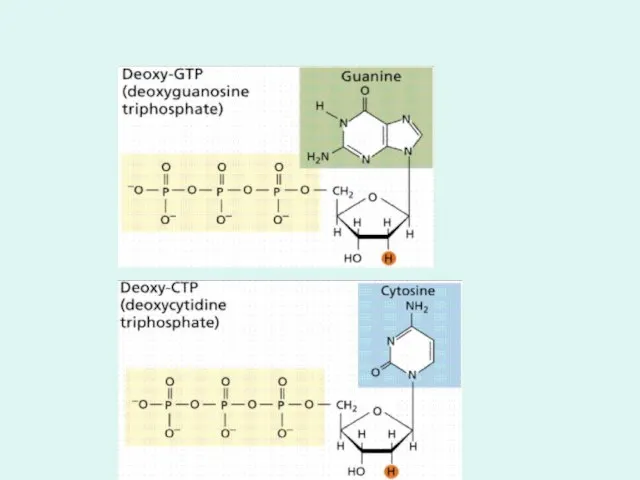
m(dАТФ+dТТФ)+
Синтез ДНК (репликация)
Суммарно процесс синтеза ДНК можно представить следующей схемой:
m(dАТФ+dТТФ)+
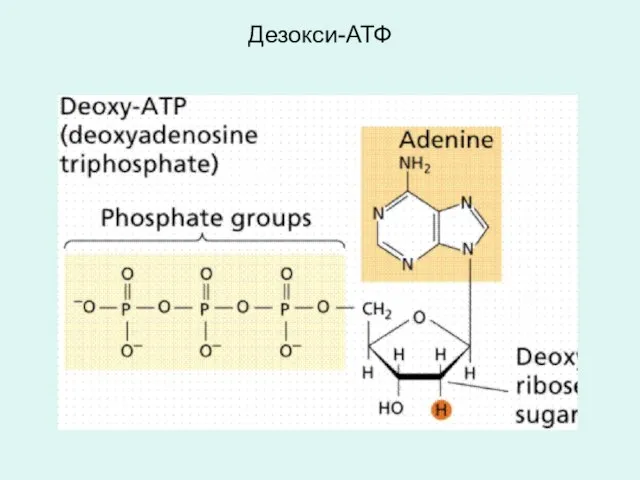
Дезокси-АТФ
Дезокси-АТФ

Для того чтобы раскрутить двойную спираль ДНК необходимы белки-ферменты ДНК-хеликазы.
ДНК-хеликазы
Для того чтобы раскрутить двойную спираль ДНК необходимы белки-ферменты ДНК-хеликазы.
ДНК-хеликазы

ДНК-полимеразы не могут начинать синтеза ДНК на матрице без затравочного полинуклеотидного
ДНК-полимеразы не могут начинать синтеза ДНК на матрице без затравочного полинуклеотидного

Удаление крайних РНК-праймеров, комплементарных 3'-концам обеих цепей линейной материнской молекулы ДНК,
Удаление крайних РНК-праймеров, комплементарных 3'-концам обеих цепей линейной материнской молекулы ДНК,

Синтез ДНК
Под воздействием расплетающих белков (ДНК-хеликаза и дестабилизирующие белки) происходит разрыв
Синтез ДНК
Под воздействием расплетающих белков (ДНК-хеликаза и дестабилизирующие белки) происходит разрыв

Далее к праймеру с помощью ДНК-полимеразы 3 присоединяются соответствующие дезоксирибонуклеозиды в
Далее к праймеру с помощью ДНК-полимеразы 3 присоединяются соответствующие дезоксирибонуклеозиды в

Синтез дочерней полинуклеотидной цепи вдоль другой полинуклеотидной цепи матричной ДНК, ввиду
Синтез дочерней полинуклеотидной цепи вдоль другой полинуклеотидной цепи матричной ДНК, ввиду

Эта реакция идет в две стадии:
1.ДНК-лигазы реагирует с НАД, которая служит
Эта реакция идет в две стадии:
1.ДНК-лигазы реагирует с НАД, которая служит

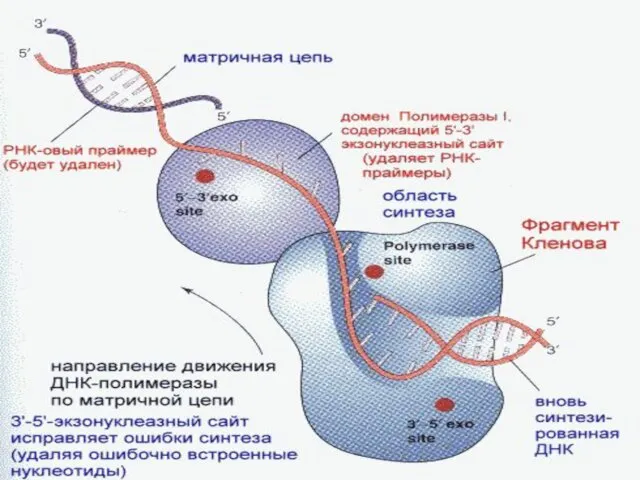

Репарация ошибок репликации
В ходе репликации самопроизвольно или под воздействием различных внешних
Репарация ошибок репликации
В ходе репликации самопроизвольно или под воздействием различных внешних

Ошибки репликации
Ошибки репликации, возникающие во время синтеза полинуклеотидных цепей дочерних молекул
Ошибки репликации
Ошибки репликации, возникающие во время синтеза полинуклеотидных цепей дочерних молекул

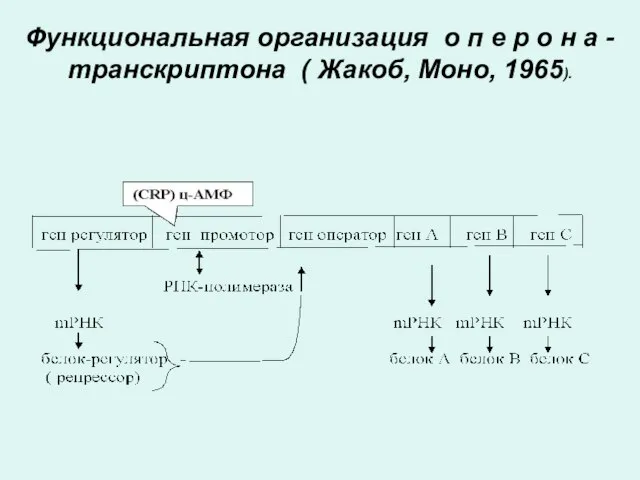
Транскрипция . Функциональная организация оперона.
Согласно современным представлениям (Жакоб, Моно, 1965) гены
Транскрипция . Функциональная организация оперона.
Согласно современным представлениям (Жакоб, Моно, 1965) гены

ОПЕРОН
Информативные участки оперона (экзоны) представлены структурными генами или цистронами, в триплетной
ОПЕРОН
Информативные участки оперона (экзоны) представлены структурными генами или цистронами, в триплетной
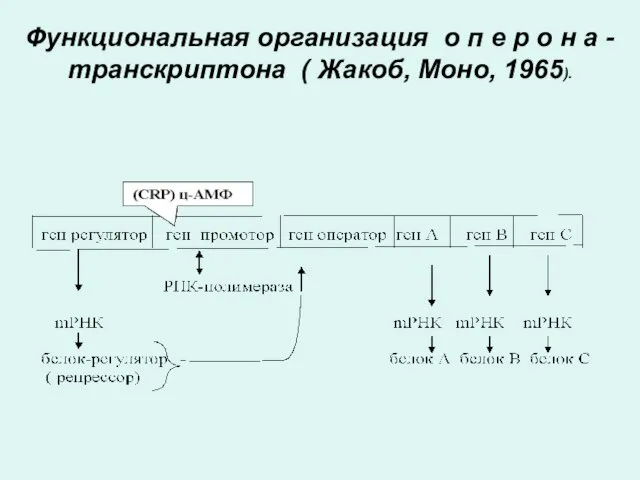
Функциональная организация о п е р о н а - транскриптона
Функциональная организация о п е р о н а - транскриптона

Функции генов оперона
Ген промотор - с него начинается транскципция.
К нему
Функции генов оперона
Ген промотор - с него начинается транскципция.
К нему

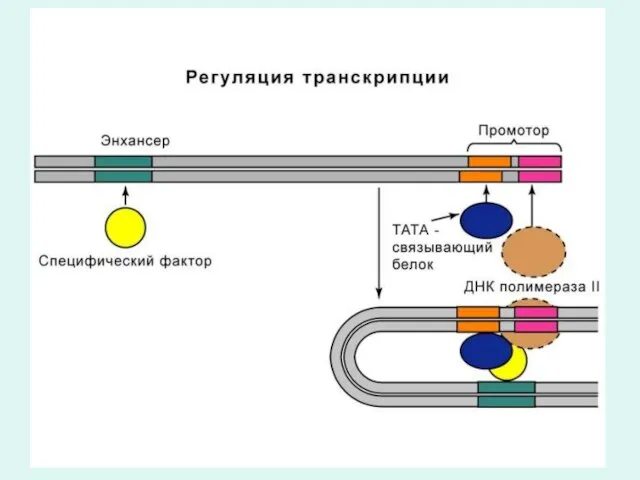
Механизм транскрипции (биосинтез РНК).
Транскрипция идет в три фазы: фаза инициации, фаза
Механизм транскрипции (биосинтез РНК).
Транскрипция идет в три фазы: фаза инициации, фаза

В фазу инициации первой, исходной реакцией синтеза РНК является реакция присоединения
В фазу инициации первой, исходной реакцией синтеза РНК является реакция присоединения

Фаза элонгации
Затем наступает фаза элонгации - наращивание полинуклеотидной цепи РНК.
В
Фаза элонгации
Затем наступает фаза элонгации - наращивание полинуклеотидной цепи РНК.
В

Терминация
Терминация (завершение) транскрипции происходит после достижения РНК-полимеразой терминирующих кодонов, являющихся стоп-сигналами.
Терминация
Терминация (завершение) транскрипции происходит после достижения РНК-полимеразой терминирующих кодонов, являющихся стоп-сигналами.

Первичный транскрипт -пре-РНК
Первичные продукты транскрипции являются полными копиями структурных генов ДНК.
Первичный транскрипт -пре-РНК
Первичные продукты транскрипции являются полными копиями структурных генов ДНК.

Процессинг
В ходе процессинга удаляются неинформативные участки в пре-РНК и образуются функционально
Процессинг
В ходе процессинга удаляются неинформативные участки в пре-РНК и образуются функционально

Молекулярные основы трансляции
В процессе трансляции можно выделить два этапа, которые имеют
Молекулярные основы трансляции
В процессе трансляции можно выделить два этапа, которые имеют
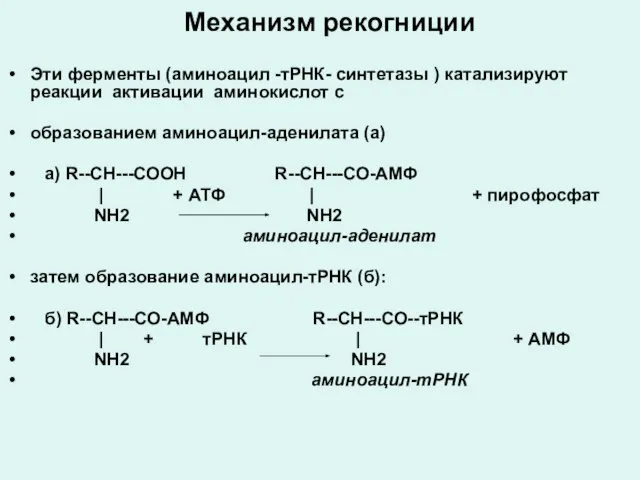
Механизм рекогниции
Эти ферменты (аминоацил -тРНК- синтетазы ) катализируют реакции активации аминокислот
Механизм рекогниции
Эти ферменты (аминоацил -тРНК- синтетазы ) катализируют реакции активации аминокислот

Биосинтез белка и факторы трансляции
Далее тРНК путем простой диффузии переносят
Биосинтез белка и факторы трансляции
Далее тРНК путем простой диффузии переносят

Рибосомы
Рибосомы являются субклеточными образованиями, состоящими из двух субъединиц - большой и
Рибосомы
Рибосомы являются субклеточными образованиями, состоящими из двух субъединиц - большой и

Сборка рибосом и фазы трансляции
Появление в клетке факторов инициации синтеза белка,
Сборка рибосом и фазы трансляции
Появление в клетке факторов инициации синтеза белка,

Фаза инициации:
Биосинтез полипептидной цепи белковой молекулы начинается с появлением в цитозоле
Фаза инициации:
Биосинтез полипептидной цепи белковой молекулы начинается с появлением в цитозоле

Формирование Р-(пептидильного) и А-(аминоацильного) центров рибосом
После образования комплекса met-тРНК -мРНК -малая
Формирование Р-(пептидильного) и А-(аминоацильного) центров рибосом
После образования комплекса met-тРНК -мРНК -малая

Фаза элонгации:
В присутствии фактора элонгации ЕF1 и за счет энергии
Фаза элонгации:
В присутствии фактора элонгации ЕF1 и за счет энергии

Фаза терминации:
Синтез полипептидной цепи продолжается до тех пор, пока на пути
Фаза терминации:
Синтез полипептидной цепи продолжается до тех пор, пока на пути
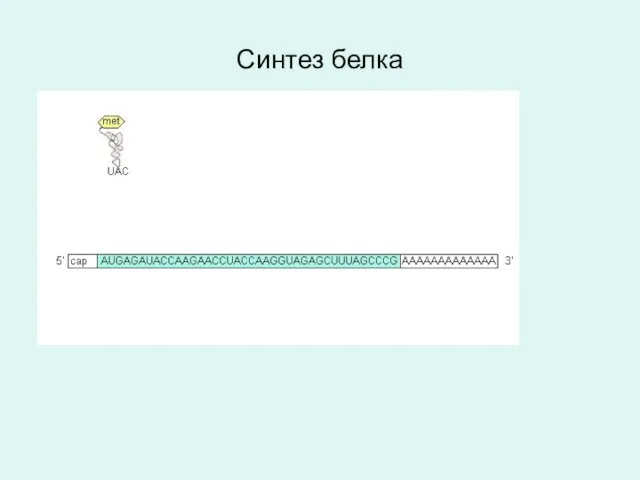
Синтез белка
Синтез белка

Посттрансляционные изменения
В результате трансляции не всегда сразу образуется функционально активный белок,
Посттрансляционные изменения
В результате трансляции не всегда сразу образуется функционально активный белок,

Ингибиторы матричных биосинтезов
Токсины выделяемые патогенными бактериами, например, дифтерийный токсин, блокирует транслокацию
Ингибиторы матричных биосинтезов
Токсины выделяемые патогенными бактериами, например, дифтерийный токсин, блокирует транслокацию

Антибиотики как ингибиторы матричных синтезов
Антибиотики, взаимодействующие с ДНК и нарушающие ее
Антибиотики как ингибиторы матричных синтезов
Антибиотики, взаимодействующие с ДНК и нарушающие ее
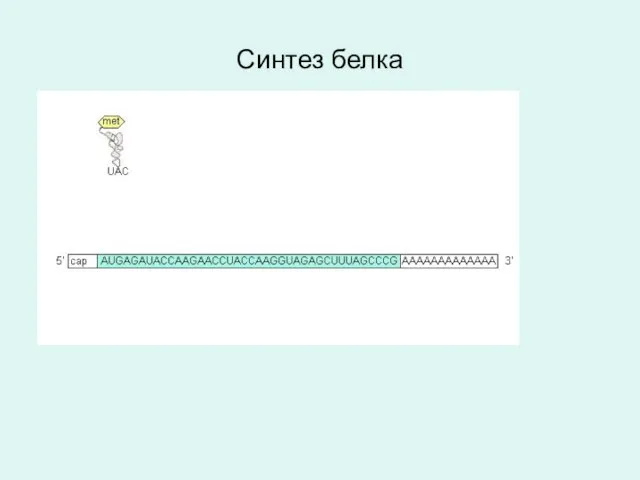
Синтез белка
Синтез белка

Регуляция биосинтеза белка
Регуляция синтеза белка в системе оперона идет в
Регуляция биосинтеза белка
Регуляция синтеза белка в системе оперона идет в

ОПЕРОН
Информативные участки оперона (экзоны) представлены структурными генами или цистронами, в триплетной
ОПЕРОН
Информативные участки оперона (экзоны) представлены структурными генами или цистронами, в триплетной
Функциональная организация о п е р о н а - транскриптона
Функциональная организация о п е р о н а - транскриптона

Функции генов оперона
Ген промотор - с него начинается транскципция.
К нему
Функции генов оперона
Ген промотор - с него начинается транскципция.
К нему

Регуляторные гены
Кроме собственно регуляторных генов в системе оперона обнаружены целый ряд
Регуляторные гены
Кроме собственно регуляторных генов в системе оперона обнаружены целый ряд

Протоонкогены
Протоонкогены имеются во всех нормальных клетках. В связи с очень большой
Протоонкогены
Протоонкогены имеются во всех нормальных клетках. В связи с очень большой

Мутации протоонкогенов
Протоонкогены находятся под тщательным и жестким контролем других генов.
Мутации
Мутации протоонкогенов
Протоонкогены находятся под тщательным и жестким контролем других генов.
Мутации

Факторы роста
Факторы роста известны как белки, индуцирующие синтез ДНКФакторы роста известны
Факторы роста
Факторы роста известны как белки, индуцирующие синтез ДНКФакторы роста известны

Большинство полипептидных факторов роста действует одновременно по паракринномуБольшинство полипептидных факторов роста
Большинство полипептидных факторов роста действует одновременно по паракринномуБольшинство полипептидных факторов роста

В регуляторных белках, обладающих интракринным действием, имеются сигнальные последовательности, обеспечивающие внутриклеточную
В регуляторных белках, обладающих интракринным действием, имеются сигнальные последовательности, обеспечивающие внутриклеточную

Последние исследования показали, женщины страдающие раком молочной железы, у которых выявлена опухоль,
Последние исследования показали, женщины страдающие раком молочной железы, у которых выявлена опухоль,

Установлено, вырабатываемый в организме человека фактор роста нейрегулин-1 способен эффективно защитить
Установлено, вырабатываемый в организме человека фактор роста нейрегулин-1 способен эффективно защитить

Современные препараты, применяемые при инсульте, эффективны только том случае, если больной
Современные препараты, применяемые при инсульте, эффективны только том случае, если больной

Опухолевые вирусы и онкогены
Некоторые опухолевые вирусы не содержат онкоген, но, встраиваясь
Опухолевые вирусы и онкогены
Некоторые опухолевые вирусы не содержат онкоген, но, встраиваясь

Превращение клеточных протоонкогенов
Превращение клеточных протоонкогенов в онкогены может происходить в результате
Превращение клеточных протоонкогенов
Превращение клеточных протоонкогенов в онкогены может происходить в результате

Онкогенные вирусы
Установлено, что гены и даже целые участки хромосом высших организмов
Онкогенные вирусы
Установлено, что гены и даже целые участки хромосом высших организмов

Ретровирусы
Как показали исследования, во многих зрелых онкогенных РНК-содержащих вирусах (ретровирусы) и
Ретровирусы
Как показали исследования, во многих зрелых онкогенных РНК-содержащих вирусах (ретровирусы) и

Онкогены
По фенотипическим проявлениям различают две группы онкогенов.
Одна группа - ядерные
Онкогены
По фенотипическим проявлениям различают две группы онкогенов.
Одна группа - ядерные

Канцерогенез
Некоторые опухолевые вирусы не содержат онкоген, но, встраиваясь в хромосому рядом
Канцерогенез
Некоторые опухолевые вирусы не содержат онкоген, но, встраиваясь в хромосому рядом

Протеин р53
В последние годы найдено еще одно, по-видимому, наиболее общее
Протеин р53
В последние годы найдено еще одно, по-видимому, наиболее общее
 Где зимуют птицы?
Где зимуют птицы?  Жизненные процессы, протекающие в корнях растений
Жизненные процессы, протекающие в корнях растений Рефлекс. Рефлекторная дуга
Рефлекс. Рефлекторная дуга Влияние различных растворов на сроки выгонки и цветения веток вишни, каштана и тополя
Влияние различных растворов на сроки выгонки и цветения веток вишни, каштана и тополя Строение древесного стебля голосеменного растения. Строение древесного стебля двудольного растения
Строение древесного стебля голосеменного растения. Строение древесного стебля двудольного растения Комбинативная изменчивость
Комбинативная изменчивость Витамины. Авитаминоз
Витамины. Авитаминоз Прості та складні суцвіття
Прості та складні суцвіття Паукообразные. Общая характеристика насекомых
Паукообразные. Общая характеристика насекомых Особенности строения растительной клетки
Особенности строения растительной клетки Взаимосвязи животных в природе.(7 класс)
Взаимосвязи животных в природе.(7 класс) ПОТРЕБНОСТЬ ОРГАНИЗМА В КИСЛОРОДЕ СТРОЕНИЕ ДЫХАТЕЛЬНОЙ СИСТЕМЫ ЧЕЛОВЕКА
ПОТРЕБНОСТЬ ОРГАНИЗМА В КИСЛОРОДЕ СТРОЕНИЕ ДЫХАТЕЛЬНОЙ СИСТЕМЫ ЧЕЛОВЕКА Зоны корня. Корень – орган минерального питания растений
Зоны корня. Корень – орган минерального питания растений  Тип Моллюски. Общая харка типа
Тип Моллюски. Общая харка типа Bill Gates is one of the most influential people in the world. He is cofounder of the most recognized brands in the computer
Bill Gates is one of the most influential people in the world. He is cofounder of the most recognized brands in the computer Ткани. Типы тканей и их свойства
Ткани. Типы тканей и их свойства Многообразие цветковых растений
Многообразие цветковых растений Лотос Комарова
Лотос Комарова Методы биологических исследований. Приготовление микропрепарата и работа с ним
Методы биологических исследований. Приготовление микропрепарата и работа с ним Оценка степени пылевого загрязнения снега и его влияние на прорастание семян
Оценка степени пылевого загрязнения снега и его влияние на прорастание семян Презентация на тему "Почвенная среда обитания" - скачать презентации по Биологии
Презентация на тему "Почвенная среда обитания" - скачать презентации по Биологии antropogenez-stadii-antropogeneza (1)
antropogenez-stadii-antropogeneza (1) Внутреннее строение рыб
Внутреннее строение рыб Пастеризация молока
Пастеризация молока Презентация на тему Учение Дарвина об искусственном отборе
Презентация на тему Учение Дарвина об искусственном отборе Презентация на тему "Самые опасные животные" - скачать бесплатно презентации по Биологии
Презентация на тему "Самые опасные животные" - скачать бесплатно презентации по Биологии Как размножаются живые организмы Урок биологии в 6 классе
Как размножаются живые организмы Урок биологии в 6 классе  Презентация на тему "Птицы Мурманской области" - скачать бесплатно презентации по Биологии
Презентация на тему "Птицы Мурманской области" - скачать бесплатно презентации по Биологии