- Отдел Аскомицеты

Содержание
- 2. Отдел аскомицеты, или сумчатые грибы – Ascomycetes Число видов: около 55 000 Клеточные стенки содержат хитин
- 3. Бесполое размножение осуществляется посредством экзогенных спор – конидий, образующихся на специализированных видоизмененных частях мицелия – конидиеносцах.
- 4. Половой процесс и развитие сумок: I. Концы антеридия и трихогины плотно срастаются, разделяющая их перегородка лизируется
- 5. VII. Пара ядер разного происхождения остается в месте перегиба крючка. Появляющиеся две перегородки отделяют место перегиба
- 6. Половой процесс и развитие сумок у аскомицетов А – антеридий (1) и аскогон (2) с трихогиной
- 7. Различают сумки прототуникатные и эутуникатные. Прототуникатные сумки имеют тонкую однослойную оболочку, споры освобождаются пассивно после разрушения
- 8. Типы эутуникатных сумок А – унитуникатные; Б - битуникатные
- 9. В свою очередь сумки развиваются в плодовых телах трёх типов:
- 10. Клейстотеций – полностью замкнутое округлой формы плодовое тело. 1 – сумка.
- 11. Перитеций – полузамкнутое кувшиновидной формы плодовое тело с отверстием на верху; 1 – сумки ; 2
- 12. Апотеций – полностью открытое плодовое тело дисковидной или блюдцевидной формы. 1 – сумки ; 2 –
- 13. У видов класса Дотидеомицеты сумки образуются в полостях – локулах, возникающих в специальных сплетениях гиф –
- 14. Экология: широко распространены во всех географических областях. Большинство сапротрофы, обитающие в почве, на субстратах органического происхождения,
- 15. Систематика: деление отдела на подотделы основано на морфологических признаках и молекулярных данных. В составе отдела выделяют
- 17. Подотдел тафриномицеты – Taphrinomycotina, или низшие архиаскомицеты - Archiascomycotina Наиболее древняя исходная группа для остальных аскомицетов.
- 18. Taphrina padi
- 19. Подотдел сахаромицеты – Saccharomycotina, или гемиаскомицеты – Hemiascomycotina Вегетативное тело в виде мицелия либо почкующихся клеток.
- 20. Пивные дрожжи: А — почкующиеся клетки; Б — сумка; В — псевдомицелий.
- 21. Подотдел собственно аскомицеты – Ascomycotina, или пезизомицеты – Pezizomycotina . Самый обширный и наиболее важный подотдел.
- 22. В составе подотдела рассматриваются следующие классы: эвроциомицеты (Eurotiomycetes), сордариомицеты (Sordariomycetes), дотидеомицеты (Dothideomycetes), пезизомицеты (Pezizomycetes), леоциомицеты (Leotiomycetes),
- 23. Класс эвроциомицеты – Eurotiomycetes Морфологически наиболее примитивны. Плодовые тела клейстотеции с прототуникатными сумками расположенными беспорядочно. Освобождение
- 24. Конидиеносцы и конидии Penicillium
- 25. Строение конидиеносцев Penicillium
- 26. АЛЕКСАНДР ФЛЕМИНГ (6 августа 1881 г. - 11 марта 1955 г. )
- 28. Зинаида Виссарионовна Ермольева (27.10 1898 — 2.12.1974)
- 30. «Мраморный» сыр «Рокфор»
- 31. Сыр «Камамбер»
- 32. Aspergillus fumigatus
- 33. Aspergillus niger
- 34. Класс сордариомицеты – Sordariomycetes Плодовые тела перитеции с унитуникатными сумками. Основной рассматриваемый порядок: Hypocreales.
- 35. Перитеции Nectria cinnabarina
- 36. Склероций Claviceps purpurea
- 37. Жизненный цикл спорыньи пурпурной. А – колос ржи со склероциями; Б – склероций, проросший головчатыми стромами;
- 38. Больной «антониевым огнём» — деталь нижней части правой створки Изенгеймского алтаря (Маттиас Грюневальд, приблизительно 1506—1515 годы).
- 39. Стромы Cordyceps militaris
- 40. Класс дотидеомицеты – Dothideomycetes Сумки образуются в специальных полостях – локулах, возникающих в специальных сплетениях гиф
- 41. Venturia inaequales 1 – пораженные лист и плод яблони; 2 – конидиальное спороношение; 3 – псевдотеций.
- 42. Конидии Venturia на листьях яблони
- 43. Класс пезизомицеты – Pezizomycetes Сумки образуются в апотециях, реже во вторичных производных апотециев – вторично замкнутых
- 44. Morchella conica – сморчок конический
- 45. Verpa bohemica – сморчковая шапочка
- 46. Gyromitra gigas – строчок гигантский
- 47. Gyromitra infula – строчок осенний
- 48. Caloscypha fulgens – калосцифа блестящая
- 50. Geopyxis carbonaria – геопиксис угольный
- 51. Humaria hemisphaerica – гумария полушаровидная
- 52. Sarcoscypha austriaca – саркосцифа австрийская
- 53. Microstoma protractum – микростома вытянутая
- 54. Choiromyces meandriformis– трюфель белый, или троицкий
- 55. Белый трюфель: А – разрез плодового тела; Б – то же при большом увеличении: 1 –
- 56. Tuber melanosporum – черный французский, или перигорский трюфель
- 57. Tuber magnatum – трюфель итальянский, или настоящий белый
- 58. Класс леоциомицеты – Leotiomycetes Апотеции преимущественно мелкие, кожистые, развиваются поодиночке или на стромах. Сумки вскрываются трещиной
- 59. Monilia fructigena: А – пораженный плод яблони с конидиальным спороношением гриба; Б – конидиальная стадия; В
- 60. Конидии Monilinia fructigena
- 61. Стромы Rhytisma acerinum
- 62. Bisporella citrina – биспорелла лимонная
- 63. Класс леканоромицеты – Lecanoromycetes Почти все виды класса – лихенизированные грибы, т. е. грибы, входящие в
- 64. Таллом образован гетеротрофным грибом (микобионт) и автотрофной водорослью или цианобактерией (фотобионт). Микобионтами в подавляющем большинстве случаев
- 65. Графис письменный – пример накипного лишайника.
- 66. Ксантория постенная – пример листоватых лишайников.
- 67. Пельтигера собачья – пример листоватых лишайников.
- 68. Кладония звездчатая, включенная в Красную книгу Мордовии, – пример кустистого лишайника.
- 69. Анатомическое строение таллома сводится к двум типам. Гомеомерный таллом образован переплетением грибных гиф и клетками водоросли,
- 70. Анатомическое строение таллома лишайников. А – строение гомеомерного слоевища: 1 – нити цианобактерии; 2 – гифы
- 71. Экология: распространены лишайники широко. Для развития лишайников необходим лишь субстрат. Соответственно типу субстрата выделяют следующие экологические
- 72. Значение в природе: Лишайники являются пионерами, заселяющими вновь образовавшиеся участки суши. Разлагая горные породы, отмирающими участками
- 73. Значение лишайников в жизни человека ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ В ПИЩУ
- 74. Aspicilia esculenta
- 75. Cetraria islandica
- 76. ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ В… медицине
- 77. Уснея бородатая
- 78. Пельтигера собачья.
- 79. Лобария лёгочная.
- 80. Гипогимния вздутая.
- 81. Пармелия бороздчатая.
- 82. ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ В... парфюмерии
- 83. Evernia pruniastri
- 84. Эверния шелушащаяся.
- 86. ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ В... бальзамировании
- 88. Мумия…
- 89. Эверния шелушащаяся.
- 90. ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ В... изготовлении красок
- 91. Пармелия пупковидная.
- 92. Рочелла.
- 93. Охролехия.
- 94. ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ В... определении возраста памятников
- 95. Ризокарпон географический
- 96. Каменные изваяния на острове Пасха
- 99. Скачать презентацию

Отдел аскомицеты, или сумчатые грибы – Ascomycetes
Число видов: около 55 000
Отдел аскомицеты, или сумчатые грибы – Ascomycetes
Число видов: около 55 000

Бесполое размножение осуществляется посредством экзогенных спор – конидий, образующихся на специализированных
Бесполое размножение осуществляется посредством экзогенных спор – конидий, образующихся на специализированных

Половой процесс и развитие сумок:
I. Концы антеридия и трихогины плотно срастаются,
Половой процесс и развитие сумок:
I. Концы антеридия и трихогины плотно срастаются,

VII. Пара ядер разного происхождения остается в месте перегиба крючка. Появляющиеся
VII. Пара ядер разного происхождения остается в месте перегиба крючка. Появляющиеся
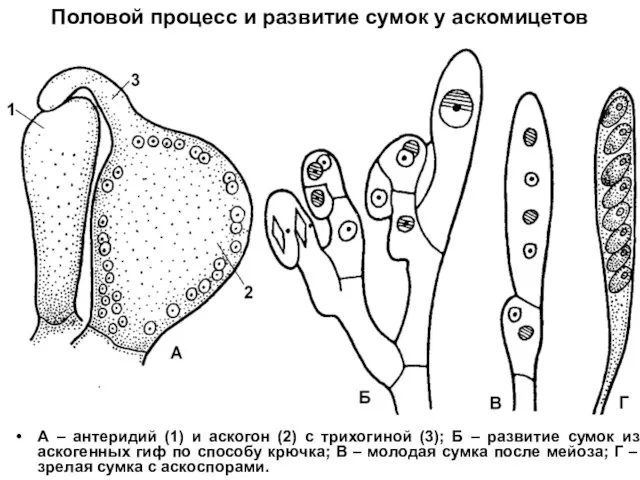
Половой процесс и развитие сумок у аскомицетов
А – антеридий (1) и
Половой процесс и развитие сумок у аскомицетов
А – антеридий (1) и

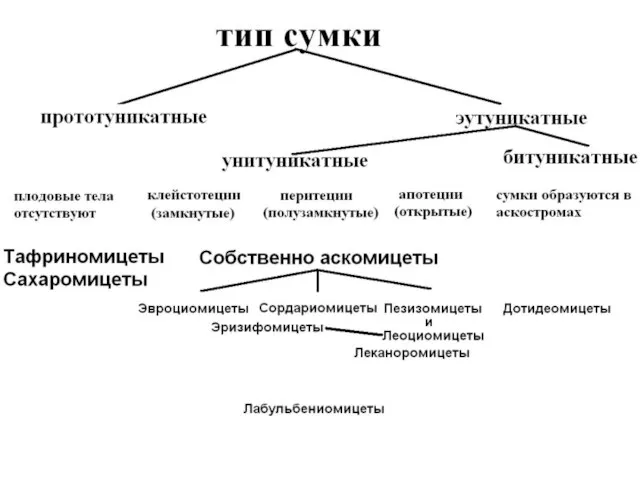
Различают сумки прототуникатные и эутуникатные.
Прототуникатные сумки имеют тонкую однослойную оболочку, споры
Различают сумки прототуникатные и эутуникатные.
Прототуникатные сумки имеют тонкую однослойную оболочку, споры
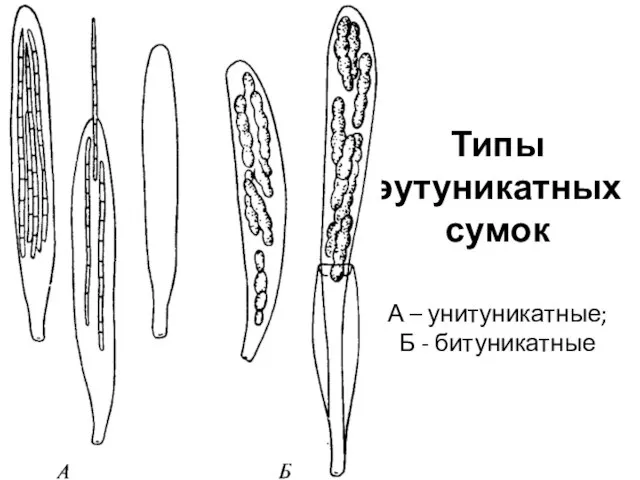
Типы эутуникатных сумок
А – унитуникатные;
Б - битуникатные
Типы эутуникатных сумок
А – унитуникатные;
Б - битуникатные

В свою очередь сумки развиваются в плодовых телах трёх типов:
В свою очередь сумки развиваются в плодовых телах трёх типов:
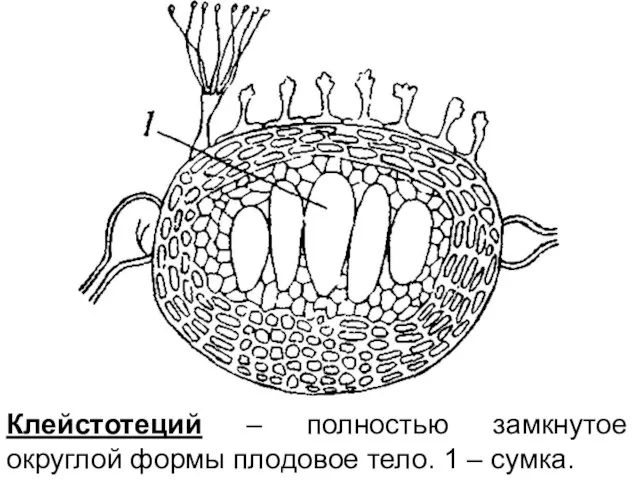
Клейстотеций – полностью замкнутое округлой формы плодовое тело. 1 – сумка.
Клейстотеций – полностью замкнутое округлой формы плодовое тело. 1 – сумка.
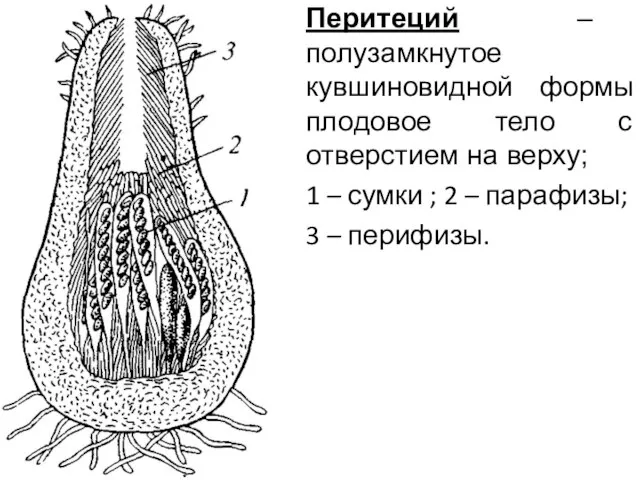
Перитеций – полузамкнутое кувшиновидной формы плодовое тело с отверстием на верху;
1
Перитеций – полузамкнутое кувшиновидной формы плодовое тело с отверстием на верху;
1
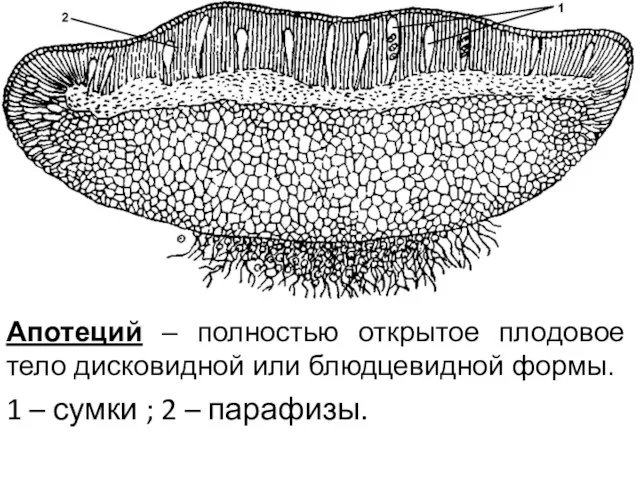
Апотеций – полностью открытое плодовое тело дисковидной или блюдцевидной формы.
1 –
Апотеций – полностью открытое плодовое тело дисковидной или блюдцевидной формы.
1 –
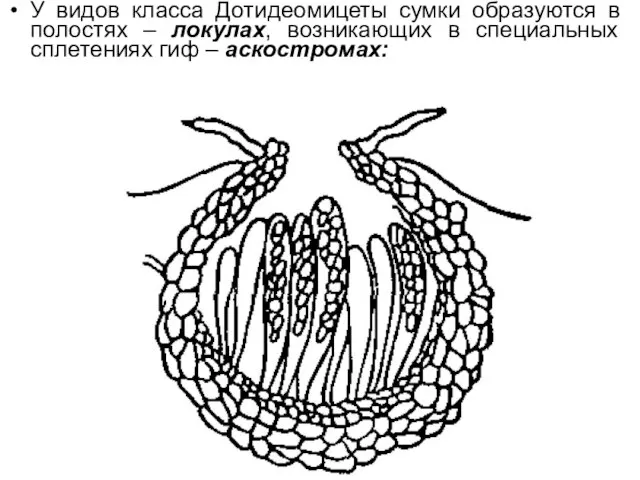
У видов класса Дотидеомицеты сумки образуются в полостях – локулах, возникающих
У видов класса Дотидеомицеты сумки образуются в полостях – локулах, возникающих

Экология: широко распространены во всех географических областях. Большинство сапротрофы, обитающие в
Экология: широко распространены во всех географических областях. Большинство сапротрофы, обитающие в

Систематика: деление отдела на подотделы основано на морфологических признаках и молекулярных
Систематика: деление отдела на подотделы основано на морфологических признаках и молекулярных

Подотдел тафриномицеты – Taphrinomycotina,
или низшие архиаскомицеты - Archiascomycotina
Наиболее древняя исходная
Подотдел тафриномицеты – Taphrinomycotina,
или низшие архиаскомицеты - Archiascomycotina
Наиболее древняя исходная

Taphrina padi
Taphrina padi

Подотдел сахаромицеты – Saccharomycotina,
или гемиаскомицеты – Hemiascomycotina
Вегетативное тело в виде
Подотдел сахаромицеты – Saccharomycotina,
или гемиаскомицеты – Hemiascomycotina
Вегетативное тело в виде
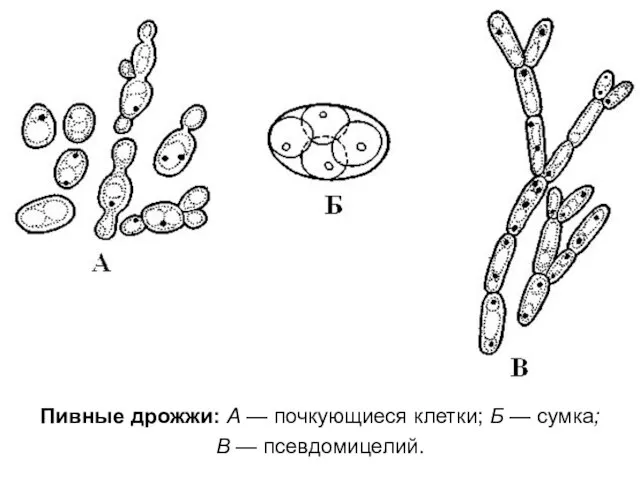
Пивные дрожжи: А — почкующиеся клетки; Б — сумка;
В —
Пивные дрожжи: А — почкующиеся клетки; Б — сумка;
В —

Подотдел собственно аскомицеты – Ascomycotina, или пезизомицеты – Pezizomycotina .
Самый обширный
Подотдел собственно аскомицеты – Ascomycotina, или пезизомицеты – Pezizomycotina .
Самый обширный

В составе подотдела рассматриваются следующие классы: эвроциомицеты (Eurotiomycetes), сордариомицеты (Sordariomycetes), дотидеомицеты
В составе подотдела рассматриваются следующие классы: эвроциомицеты (Eurotiomycetes), сордариомицеты (Sordariomycetes), дотидеомицеты

Класс эвроциомицеты – Eurotiomycetes
Морфологически наиболее примитивны. Плодовые тела клейстотеции с
Класс эвроциомицеты – Eurotiomycetes
Морфологически наиболее примитивны. Плодовые тела клейстотеции с

Конидиеносцы и конидии Penicillium
Конидиеносцы и конидии Penicillium
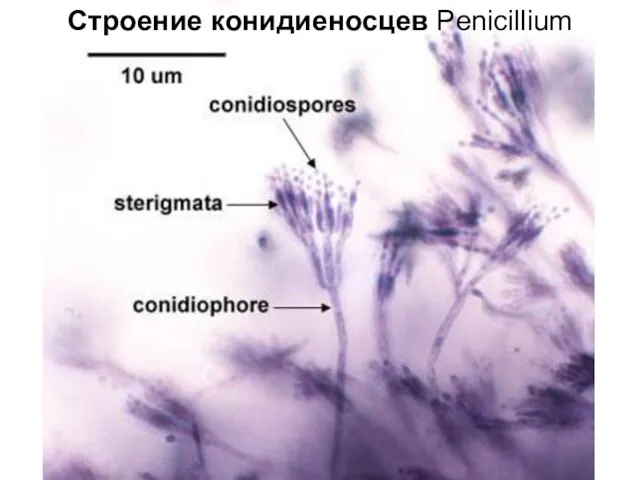
Строение конидиеносцев Penicillium
Строение конидиеносцев Penicillium

АЛЕКСАНДР ФЛЕМИНГ
(6 августа 1881 г. - 11 марта 1955 г. )
АЛЕКСАНДР ФЛЕМИНГ (6 августа 1881 г. - 11 марта 1955 г. )


Зинаида Виссарионовна Ермольева
(27.10 1898 — 2.12.1974)
Зинаида Виссарионовна Ермольева
(27.10 1898 — 2.12.1974)


«Мраморный» сыр «Рокфор»
«Мраморный» сыр «Рокфор»

Сыр «Камамбер»
Сыр «Камамбер»
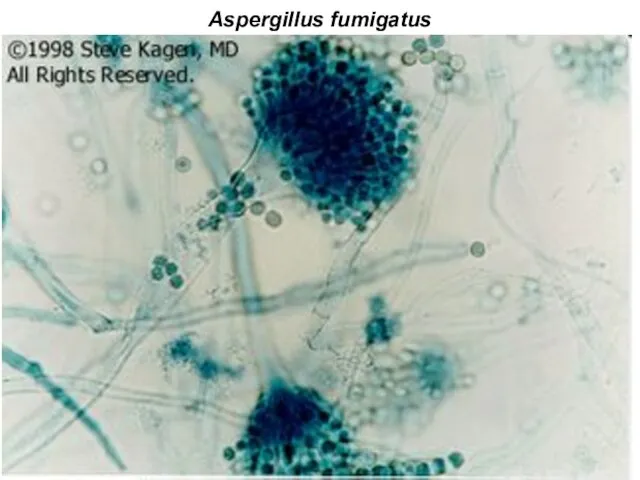
Aspergillus fumigatus
Aspergillus fumigatus
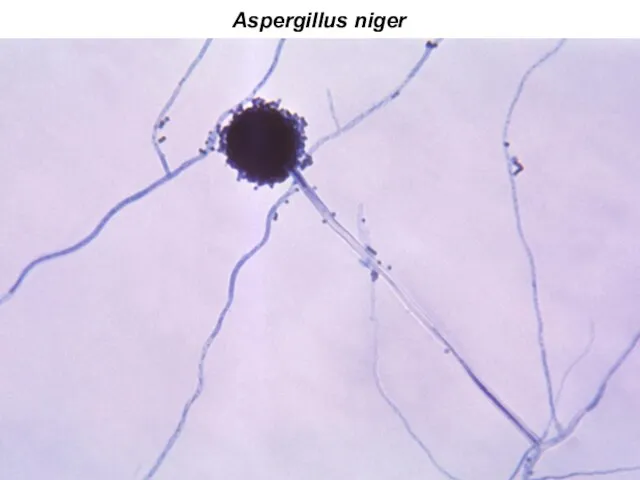
Aspergillus niger
Aspergillus niger

Класс сордариомицеты – Sordariomycetes
Плодовые тела перитеции с унитуникатными сумками.
Основной
Класс сордариомицеты – Sordariomycetes
Плодовые тела перитеции с унитуникатными сумками.
Основной

Перитеции Nectria cinnabarina
Перитеции Nectria cinnabarina

Склероций
Claviceps purpurea
Склероций
Claviceps purpurea
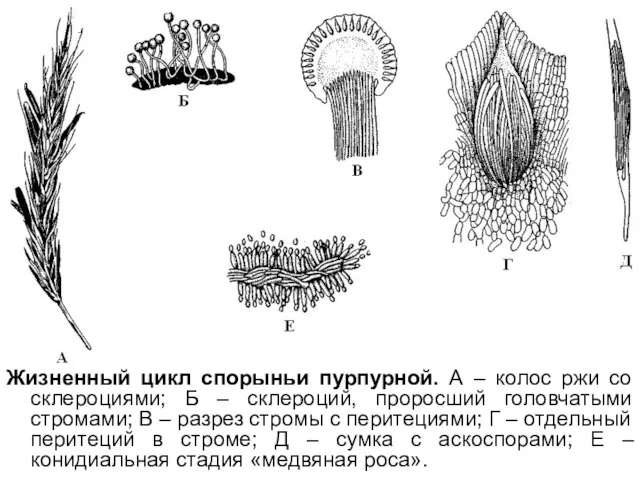
Жизненный цикл спорыньи пурпурной. А – колос ржи со склероциями; Б
Жизненный цикл спорыньи пурпурной. А – колос ржи со склероциями; Б
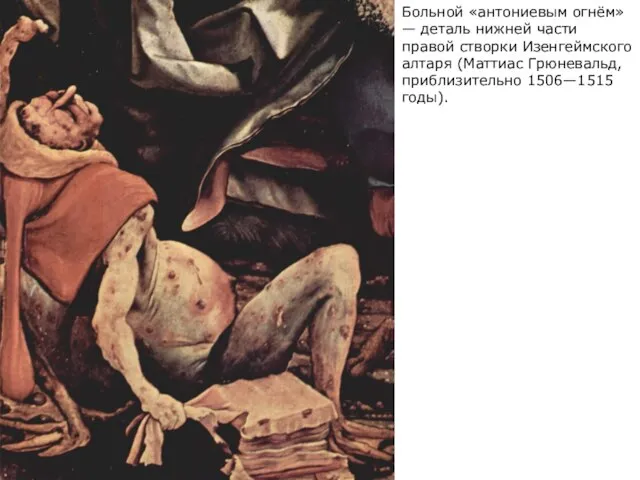
Больной «антониевым огнём» — деталь нижней части правой створки Изенгеймского алтаря
Больной «антониевым огнём» — деталь нижней части правой створки Изенгеймского алтаря

Стромы Cordyceps militaris
Стромы Cordyceps militaris

Класс дотидеомицеты – Dothideomycetes
Сумки образуются в специальных полостях – локулах,
Класс дотидеомицеты – Dothideomycetes
Сумки образуются в специальных полостях – локулах,
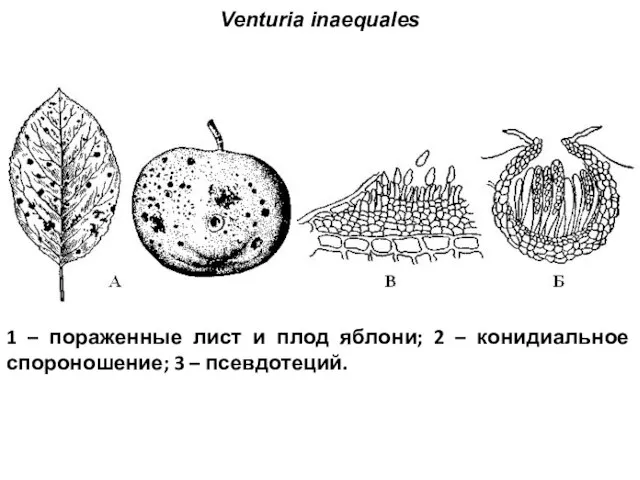
Venturia inaequales
1 – пораженные лист и плод яблони; 2 – конидиальное
Venturia inaequales
1 – пораженные лист и плод яблони; 2 – конидиальное
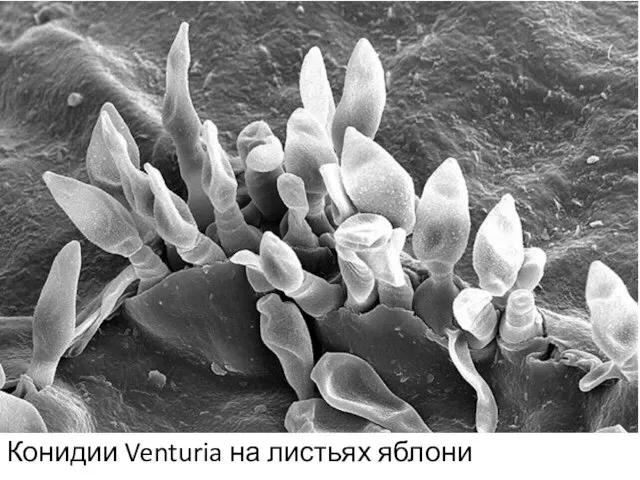
Конидии Venturia на листьях яблони
Конидии Venturia на листьях яблони

Класс пезизомицеты – Pezizomycetes
Сумки образуются в апотециях, реже во вторичных
Класс пезизомицеты – Pezizomycetes
Сумки образуются в апотециях, реже во вторичных

Morchella conica – сморчок конический
Morchella conica – сморчок конический

Verpa bohemica – сморчковая шапочка
Verpa bohemica – сморчковая шапочка

Gyromitra gigas – строчок гигантский
Gyromitra gigas – строчок гигантский

Gyromitra infula – строчок осенний
Gyromitra infula – строчок осенний

Caloscypha fulgens – калосцифа блестящая
Caloscypha fulgens – калосцифа блестящая


Geopyxis carbonaria – геопиксис угольный
Geopyxis carbonaria – геопиксис угольный

Humaria hemisphaerica – гумария полушаровидная
Humaria hemisphaerica – гумария полушаровидная

Sarcoscypha austriaca – саркосцифа австрийская
Sarcoscypha austriaca – саркосцифа австрийская

Microstoma protractum – микростома вытянутая
Microstoma protractum – микростома вытянутая

Choiromyces meandriformis– трюфель белый, или троицкий
Choiromyces meandriformis– трюфель белый, или троицкий
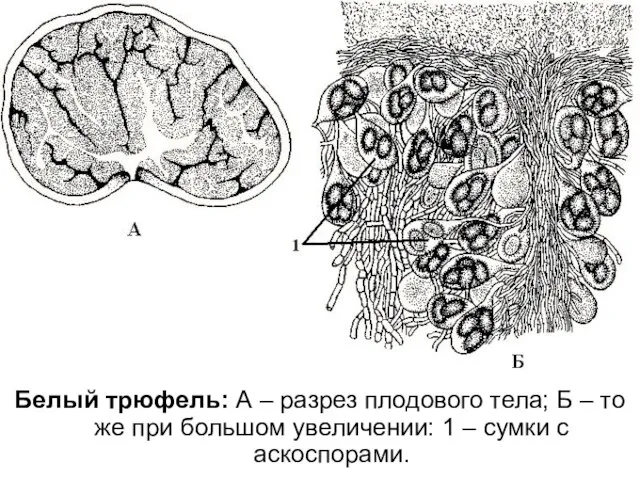
Белый трюфель: А – разрез плодового тела; Б – то же
Белый трюфель: А – разрез плодового тела; Б – то же

Tuber melanosporum – черный французский, или перигорский трюфель
Tuber melanosporum – черный французский, или перигорский трюфель

Tuber magnatum – трюфель итальянский, или настоящий белый
Tuber magnatum – трюфель итальянский, или настоящий белый

Класс леоциомицеты – Leotiomycetes
Апотеции преимущественно мелкие, кожистые, развиваются поодиночке или
Класс леоциомицеты – Leotiomycetes
Апотеции преимущественно мелкие, кожистые, развиваются поодиночке или
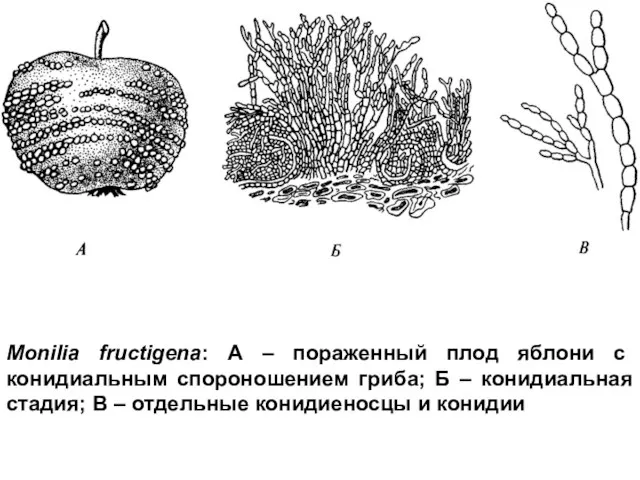
Monilia fructigena: А – пораженный плод яблони с конидиальным спороношением гриба;
Monilia fructigena: А – пораженный плод яблони с конидиальным спороношением гриба;

Конидии Monilinia fructigena
Конидии Monilinia fructigena

Стромы Rhytisma acerinum
Стромы Rhytisma acerinum

Bisporella citrina – биспорелла лимонная
Bisporella citrina – биспорелла лимонная

Класс леканоромицеты – Lecanoromycetes
Почти все виды класса – лихенизированные грибы,
Класс леканоромицеты – Lecanoromycetes
Почти все виды класса – лихенизированные грибы,

Таллом образован гетеротрофным грибом (микобионт) и автотрофной водорослью или цианобактерией (фотобионт).
Таллом образован гетеротрофным грибом (микобионт) и автотрофной водорослью или цианобактерией (фотобионт).

Графис письменный – пример накипного лишайника.
Графис письменный – пример накипного лишайника.

Ксантория постенная – пример листоватых лишайников.
Ксантория постенная – пример листоватых лишайников.

Пельтигера собачья – пример листоватых лишайников.
Пельтигера собачья – пример листоватых лишайников.

Кладония звездчатая, включенная в Красную книгу Мордовии, – пример кустистого лишайника.
Кладония звездчатая, включенная в Красную книгу Мордовии, – пример кустистого лишайника.

Анатомическое строение таллома сводится к двум типам. Гомеомерный таллом образован переплетением
Анатомическое строение таллома сводится к двум типам. Гомеомерный таллом образован переплетением

Анатомическое строение таллома лишайников.
А – строение гомеомерного слоевища: 1 – нити
Анатомическое строение таллома лишайников.
А – строение гомеомерного слоевища: 1 – нити

Экология: распространены лишайники широко. Для развития лишайников необходим лишь субстрат. Соответственно
Экология: распространены лишайники широко. Для развития лишайников необходим лишь субстрат. Соответственно

Значение в природе: Лишайники являются пионерами, заселяющими вновь образовавшиеся участки суши.

Значение лишайников в жизни человека
ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ В ПИЩУ
Значение лишайников в жизни человека
ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ В ПИЩУ

Aspicilia esculenta
Aspicilia esculenta

Cetraria islandica
Cetraria islandica

ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ В… медицине
ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ В… медицине

Уснея бородатая
Уснея бородатая

Пельтигера собачья.
Пельтигера собачья.

Лобария лёгочная.
Лобария лёгочная.

Гипогимния вздутая.
Гипогимния вздутая.

Пармелия бороздчатая.
Пармелия бороздчатая.

ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ В... парфюмерии
ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ В... парфюмерии

Evernia pruniastri
Evernia pruniastri

Эверния шелушащаяся.
Эверния шелушащаяся.


ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ В... бальзамировании
ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ В... бальзамировании


Мумия…
Мумия…

Эверния шелушащаяся.
Эверния шелушащаяся.

ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ В...
изготовлении красок
изготовлении красок

Пармелия пупковидная.
Пармелия пупковидная.
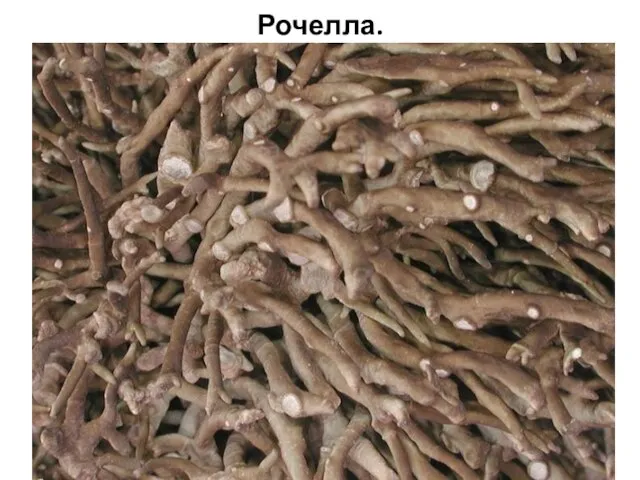
Рочелла.
Рочелла.

Охролехия.
Охролехия.

ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ В... определении возраста памятников
ИСПОЛЬЗОВАНИЕ ЛИШАЙНИКОВ В... определении возраста памятников

Ризокарпон географический
Ризокарпон географический

Каменные изваяния на острове Пасха
Каменные изваяния на острове Пасха

 Что такое фотодиссоциация или фотолиз воды
Что такое фотодиссоциация или фотолиз воды Аттестационная работа. Методическая разработка по выполнению исследовательской работы Плесневые грибы
Аттестационная работа. Методическая разработка по выполнению исследовательской работы Плесневые грибы Презентация на тему Плоские, круглые, кольчатые черви
Презентация на тему Плоские, круглые, кольчатые черви  Особенности происходящие в живом организме в физико-химической интерпретации
Особенности происходящие в живом организме в физико-химической интерпретации МОУ СОШ № 13 с углубленным изучением отдельных предметов Учитель биологии : О.П. Жмаева
МОУ СОШ № 13 с углубленным изучением отдельных предметов Учитель биологии : О.П. Жмаева Мульчирование
Мульчирование Презентация на тему: «Млекопитающие» Выполнила: ученица 3 класса Спасской основной общеобразовательной школы Титова Анастасия
Презентация на тему: «Млекопитающие» Выполнила: ученица 3 класса Спасской основной общеобразовательной школы Титова Анастасия Потребность пациента в сне и отдыхе
Потребность пациента в сне и отдыхе Плоды. Образование плода
Плоды. Образование плода Целлюлоза. Применение целлюлозы
Целлюлоза. Применение целлюлозы Иммунолюминесцентный анализ
Иммунолюминесцентный анализ Презентация на тему "Рост и развитие животных" - скачать презентации по Биологии
Презентация на тему "Рост и развитие животных" - скачать презентации по Биологии Весенний гомон птичьих стай
Весенний гомон птичьих стай Собака - друг человека
Собака - друг человека Полимеразная цепная реакция
Полимеразная цепная реакция Онтогенез
Онтогенез Строение, размножение, развитие млекопитающих
Строение, размножение, развитие млекопитающих Эволюция. Квест-игра
Эволюция. Квест-игра Биология почв
Биология почв Сучасні біологічні науки. Зв’язок біології з іншими науками
Сучасні біологічні науки. Зв’язок біології з іншими науками  Презентация на тему "Життя до народження" - скачать бесплатно презентации по Биологии
Презентация на тему "Життя до народження" - скачать бесплатно презентации по Биологии Отряд Приматы
Отряд Приматы  Декоративные достоинства лиственных деревьев
Декоративные достоинства лиственных деревьев Звери - обитатели живых уголков
Звери - обитатели живых уголков Малды сойысқа жығу
Малды сойысқа жығу Инкрустирующие вещества клеточной оболочки растений Презентация по биологии
Инкрустирующие вещества клеточной оболочки растений Презентация по биологии Морфология побега высших растений. Побеговые системы. (Лекция 7)
Морфология побега высших растений. Побеговые системы. (Лекция 7) Основы генетики
Основы генетики