- Полуконсервативный характер репликации ДНК

Содержание
- 2. В одном цикле репликации каждая из двух цепей ДНК используется в качестве матрицы для образования комплементарной
- 3. Этап инициация репликационных вилок в точках начала: Origin репликации. Затем следует образование репликационного глазка. У E.coli
- 4. Матричный синтез ДНК-это механизм, используемый клеткой, чтобы копировать нуклеотидную последовательность одной цепи ДНК в комплементарную последовательность
- 5. Расплетание двойной спирали ДНК Для раскрытия двойной спирали необходимы особые белки. Они бывают двух типов: ДНК-хеликазы
- 6. Влияние белков на структуру одноцепочечной ДНК Каждая молекула белка предпочитает связывать ДНК, находясь в контакте с
- 7. Закручивание ДНК в ходе репликации В спирали ДНК образуется шарнир, формируемый белками –топоизомеразами. Это обратимая нуклеаза,
- 8. Реакция надрезания ДНК
- 9. Первый фермент, участвующий в реакции - ДНК-полимераза был открыт в 1957г. Свободные нуклеотиды, служащие субстратами для
- 10. Структура репликационной вилки Обе цепи дочерней ДНК полимеризуются в направлении 5’ к 3’, синтезируемая на отстающей
- 11. Коррекция репликации ДНК-полимераза выступает с экзонуклеазной активностью (удаляет ошибочно включенный нуклеотид). Экзонуклеазы-ферменты, которые отщепляют нуклеотиды одного
- 12. ДНК-полимеразы иногда все же ошибаются. Клетки имеют шанс исправить эти ошибки при помощи процесса Направляемая цепь
- 13. Синтез РНК-затравки РНК-праймаза – фермент, который синтезирует короткие РНК-затравки на отстающей цепи, используя ДНК в качестве
- 14. Синтез фрагмента ДНК на отстающей цепи В клетках эукариот РНК-затравки синтезируются на отстающей цепи с промежутками,
- 15. Реакция с ДНК-лигазой Фермент «сшивает» разрыв фосфодиэфирной связи. ДНК-лигаза использует молекулу АТР, чтобы активизировать 5’ конец
- 16. В репликационном составе эукариот белковых компонентов больше чем у бактерий. SSB белок у эукариот из 3
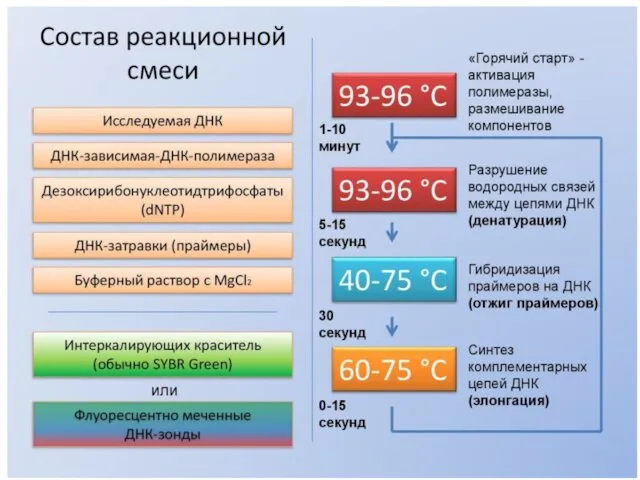
- 17. ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ (ПЦР) POLYMERASE CHAIN REACTION (PCR)
- 28. Скачать презентацию
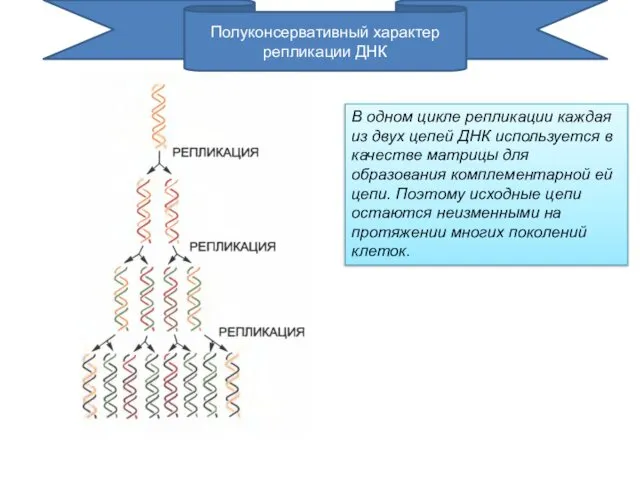
В одном цикле репликации каждая из двух цепей ДНК используется в
В одном цикле репликации каждая из двух цепей ДНК используется в
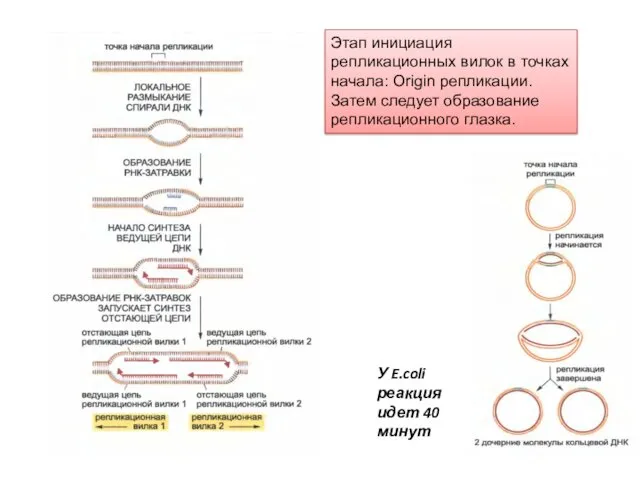
Этап инициация репликационных вилок в точках начала: Origin репликации. Затем следует
Этап инициация репликационных вилок в точках начала: Origin репликации. Затем следует
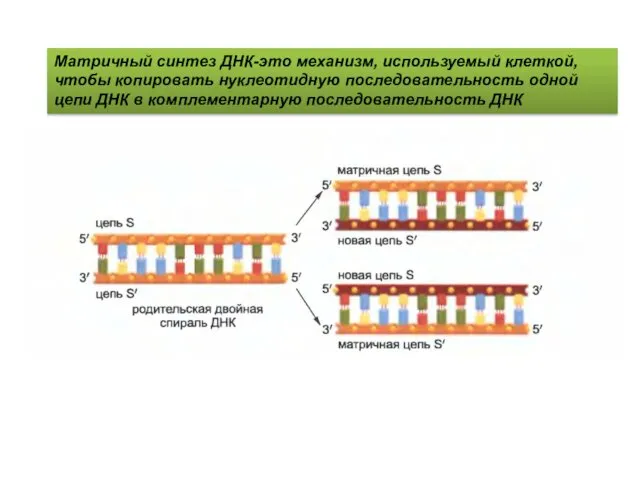
Матричный синтез ДНК-это механизм, используемый клеткой, чтобы копировать нуклеотидную последовательность одной
Матричный синтез ДНК-это механизм, используемый клеткой, чтобы копировать нуклеотидную последовательность одной
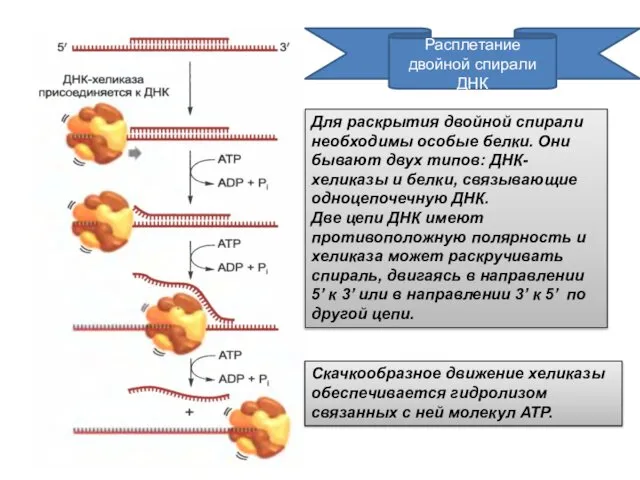
Расплетание двойной спирали ДНК
Для раскрытия двойной спирали необходимы особые белки. Они
Расплетание двойной спирали ДНК
Для раскрытия двойной спирали необходимы особые белки. Они
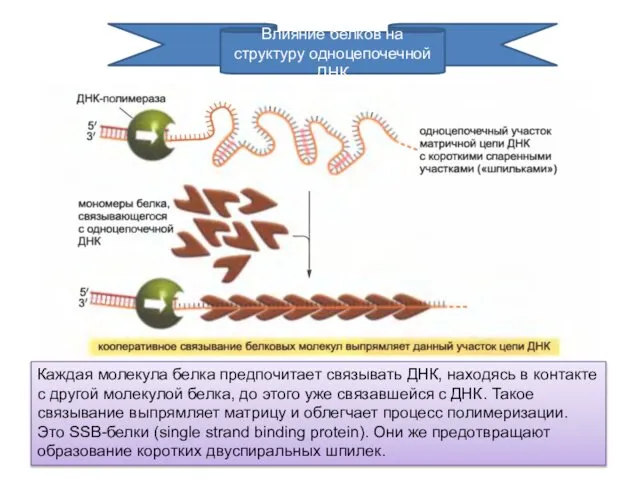
Влияние белков на структуру одноцепочечной ДНК
Каждая молекула белка предпочитает связывать ДНК,
Влияние белков на структуру одноцепочечной ДНК
Каждая молекула белка предпочитает связывать ДНК,
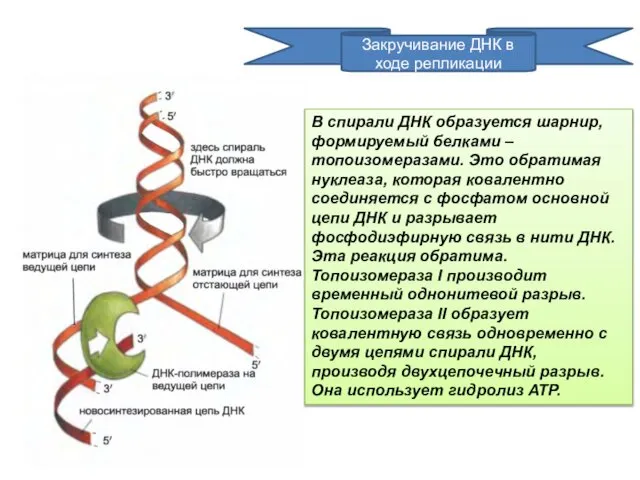
Закручивание ДНК в ходе репликации
В спирали ДНК образуется шарнир, формируемый белками
Закручивание ДНК в ходе репликации
В спирали ДНК образуется шарнир, формируемый белками
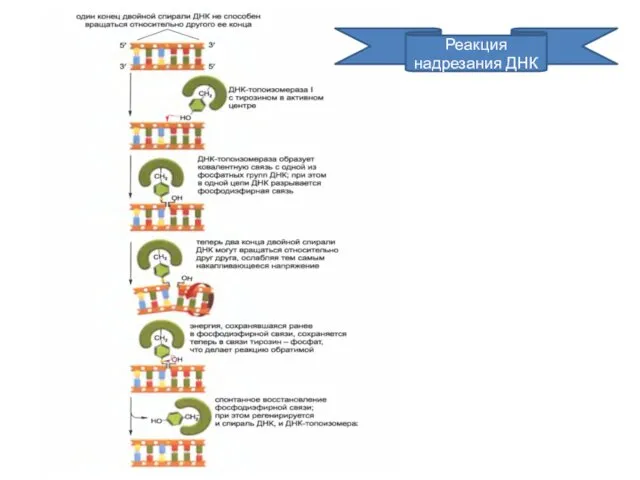
Реакция надрезания ДНК
Реакция надрезания ДНК
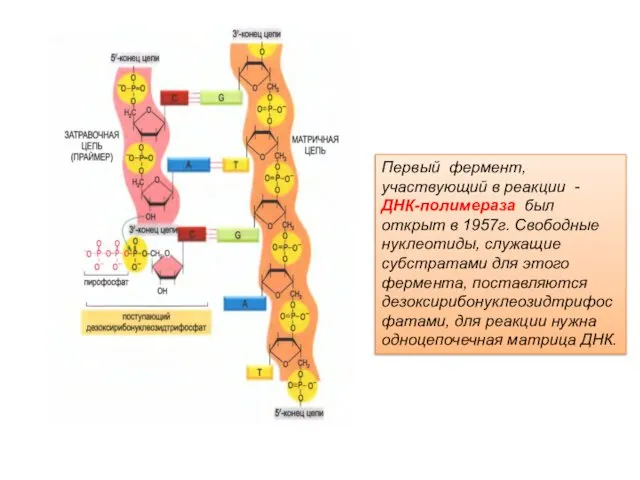
Первый фермент, участвующий в реакции - ДНК-полимераза был открыт в 1957г.
Первый фермент, участвующий в реакции - ДНК-полимераза был открыт в 1957г.
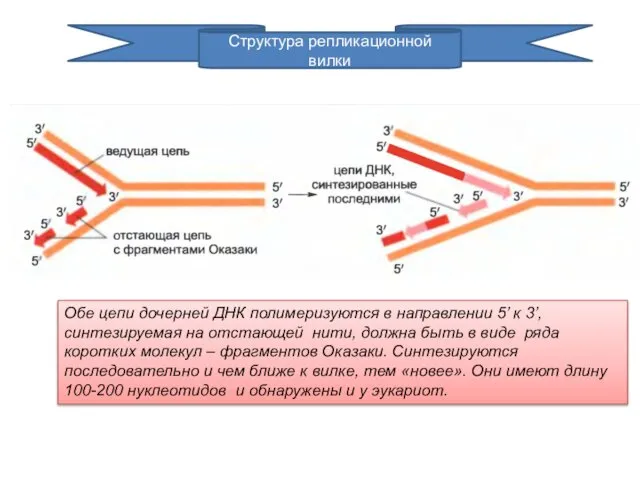
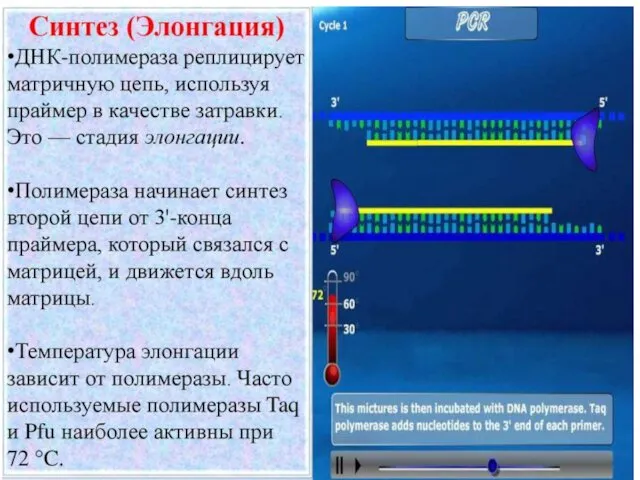
Структура репликационной вилки
Обе цепи дочерней ДНК полимеризуются в направлении 5’ к
Структура репликационной вилки
Обе цепи дочерней ДНК полимеризуются в направлении 5’ к
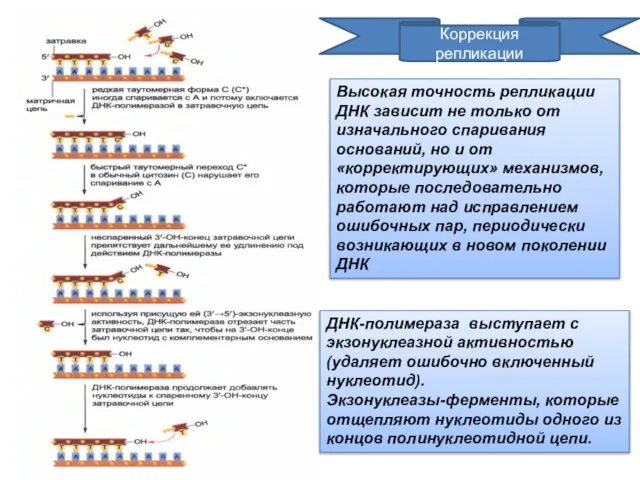
Коррекция
репликации
ДНК-полимераза выступает с экзонуклеазной активностью (удаляет ошибочно включенный нуклеотид).
Экзонуклеазы-ферменты, которые
Коррекция
репликации
ДНК-полимераза выступает с экзонуклеазной активностью (удаляет ошибочно включенный нуклеотид).
Экзонуклеазы-ферменты, которые
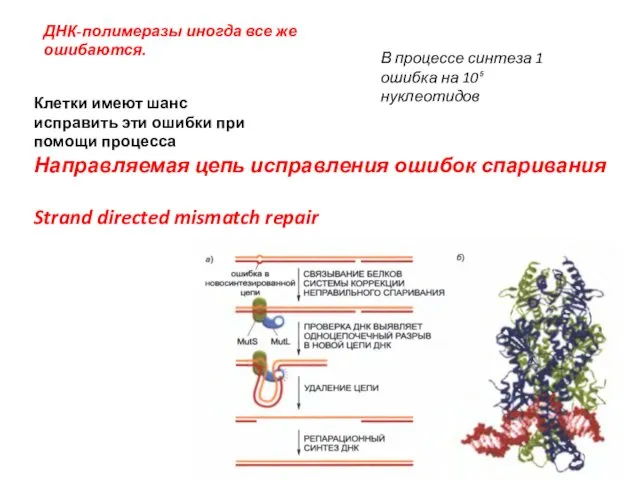
ДНК-полимеразы иногда все же ошибаются.
Клетки имеют шанс исправить эти ошибки при
ДНК-полимеразы иногда все же ошибаются.
Клетки имеют шанс исправить эти ошибки при
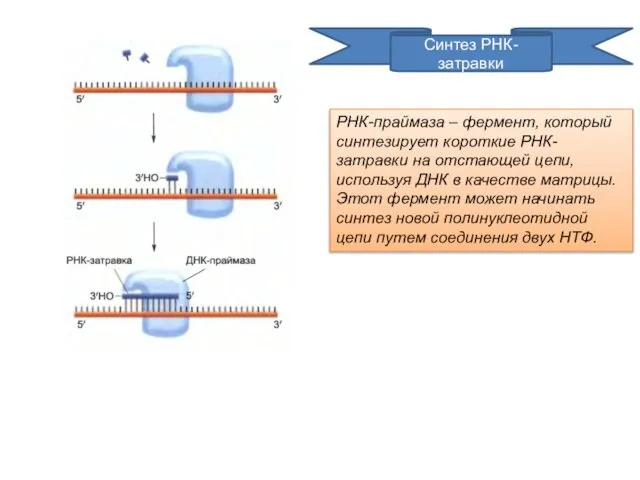
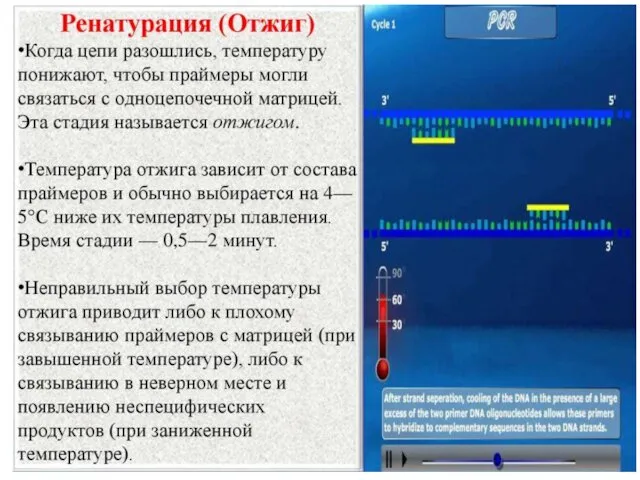
Синтез РНК-затравки
РНК-праймаза – фермент, который синтезирует короткие РНК-затравки на отстающей цепи,
Синтез РНК-затравки
РНК-праймаза – фермент, который синтезирует короткие РНК-затравки на отстающей цепи,
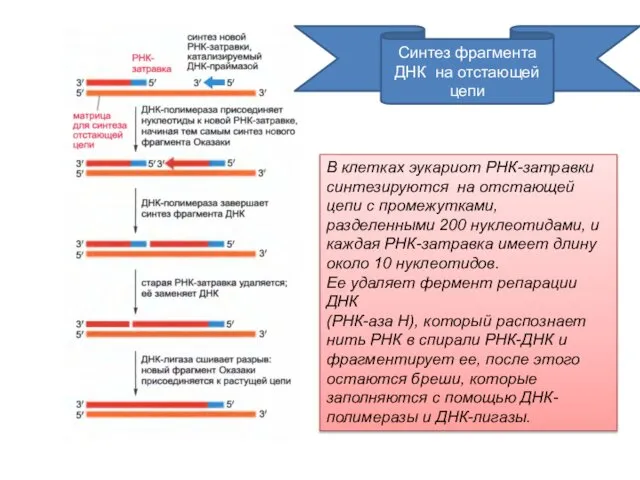
Синтез фрагмента ДНК на отстающей цепи
В клетках эукариот РНК-затравки синтезируются
Синтез фрагмента ДНК на отстающей цепи
В клетках эукариот РНК-затравки синтезируются
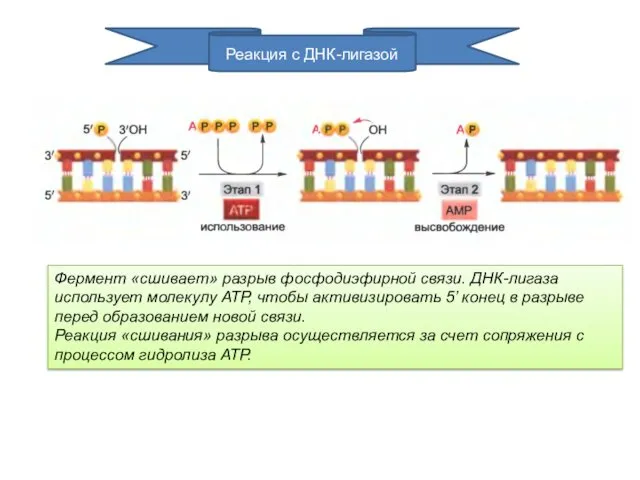
Реакция с ДНК-лигазой
Фермент «сшивает» разрыв фосфодиэфирной связи. ДНК-лигаза использует молекулу АТР,
Реакция с ДНК-лигазой
Фермент «сшивает» разрыв фосфодиэфирной связи. ДНК-лигаза использует молекулу АТР,

В репликационном составе эукариот белковых компонентов больше чем у бактерий.
SSB белок
В репликационном составе эукариот белковых компонентов больше чем у бактерий.
SSB белок

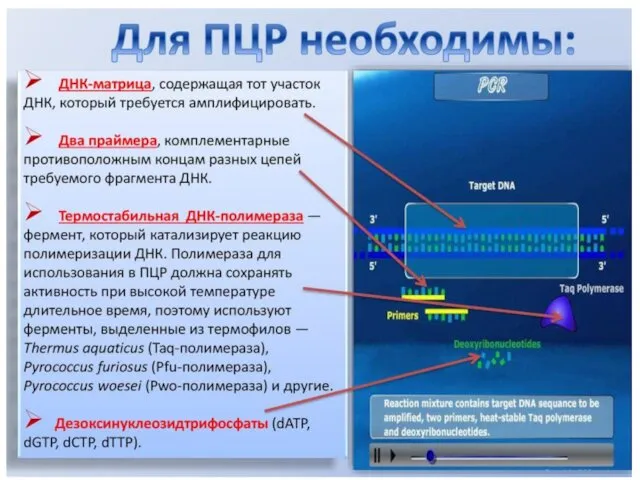
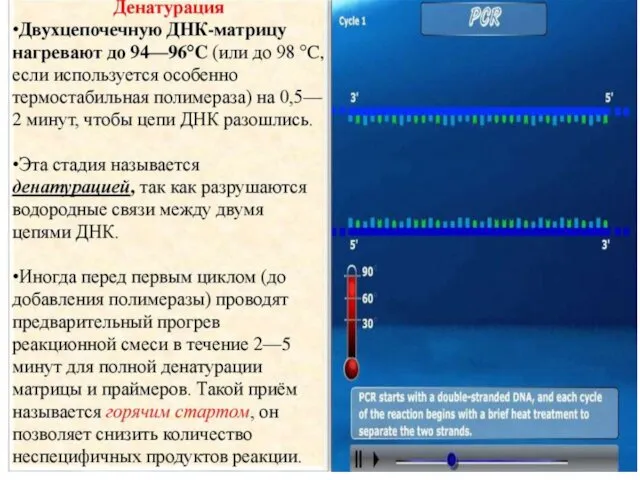
ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ (ПЦР)
POLYMERASE CHAIN REACTION (PCR)
ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ (ПЦР)
POLYMERASE CHAIN REACTION (PCR)




 Комбинативная изменчивость
Комбинативная изменчивость Строение и функции уха
Строение и функции уха Царство грибов
Царство грибов Физиология дыхания
Физиология дыхания Мыши - семейство млекопитающих отряда грызунов
Мыши - семейство млекопитающих отряда грызунов Тема урока: Роль кожи в терморегуляции организма.
Тема урока: Роль кожи в терморегуляции организма. Отряды Насекомоядные и Рукокрылые
Отряды Насекомоядные и Рукокрылые «Щупальцы мозга»
«Щупальцы мозга» Презентация на тему "Жуки" - скачать бесплатно презентации по Биологии
Презентация на тему "Жуки" - скачать бесплатно презентации по Биологии Пищеварительная трубка
Пищеварительная трубка Жизнедеятельность клетки, её деление и рост
Жизнедеятельность клетки, её деление и рост Органические вещества клетки. Углеводы
Органические вещества клетки. Углеводы Функция и строение плечевого сустава
Функция и строение плечевого сустава Биологические ритмы. Фотопериодизм.
Биологические ритмы. Фотопериодизм.  Повреждение органелл
Повреждение органелл Презентация на тему "КЛЕТКА" - скачать бесплатно презентации по Биологии_
Презентация на тему "КЛЕТКА" - скачать бесплатно презентации по Биологии_ Нейросифилис: атипичная цереброваскулярная и паретическая формы Самсон Xачатрян Кафедра неврологии Ереванского Государственного Медицинского Университета им. М.Гераци, Медицинский Центр “Эребуни”
Нейросифилис: атипичная цереброваскулярная и паретическая формы Самсон Xачатрян Кафедра неврологии Ереванского Государственного Медицинского Университета им. М.Гераци, Медицинский Центр “Эребуни”  Загальна характеристика класу Павукоподібні
Загальна характеристика класу Павукоподібні ШИРОКОЛИСТВЕННЫЙ ЛЕС КАК ПРИРОДНОЕ СООБЩЕСТВО Подготовила Учитель МКОУ Тальменская СОШ №1 Акимова О.А.
ШИРОКОЛИСТВЕННЫЙ ЛЕС КАК ПРИРОДНОЕ СООБЩЕСТВО Подготовила Учитель МКОУ Тальменская СОШ №1 Акимова О.А.  Водно-солевой обмен у животных
Водно-солевой обмен у животных Генетика клеточного цикла. Протеолиз циклина под контролем АРС. (Глава 4)
Генетика клеточного цикла. Протеолиз циклина под контролем АРС. (Глава 4) Красный волк
Красный волк Анатомия сердца
Анатомия сердца Учение Ч. Дарвина об искусственном отборе
Учение Ч. Дарвина об искусственном отборе Умови, необхідні для забезпечення життєдіяльності рослин
Умови, необхідні для забезпечення життєдіяльності рослин  Энергетический метаболизм прокариот
Энергетический метаболизм прокариот Презентация на тему Закономерности развития живой природы. Эволюционное учение
Презентация на тему Закономерности развития живой природы. Эволюционное учение Физиология ретикулярной формации
Физиология ретикулярной формации