- Энергетический метаболизм прокариот

Содержание
- 2. Метаболизм – совокупность ферментативных процессов, протекающих в клетке и обеспечивающих её энергетические и биосинтетические потребности. Энергетический
- 3. Классификация микроорганизмов По источнику углерода Автотрофы Гетеротрофы СО2 органические соединения В зависимости от донора электронов По
- 4. Факторы роста – вещества, которые прокариоты по каким-либо причинам не могут синтезировать самостоятельно из используемого источника
- 6. Бактерии, изучаемые медицинской микробиологией- хемоорганогетеротрофы Отличительной особенностью этой группы является то, что источник углерода у них
- 7. В общем виде процессы, способные служить источником энергии для прокариот, можно представить следующим образом Должен существовать
- 9. Способность использовать химическую энергию присуща всем без исключения организмам. Особенно многообразны возможности прокариот. Мономеры, образованные при
- 10. Основные катаболические системы клетки: гликолиз (путь Эмбдена-Мейергофа-Парнаса), окислительный пентозофосфатный путь, путь Энтнера-Дудорова цикл трикарбоновых кислот (ЦТК)
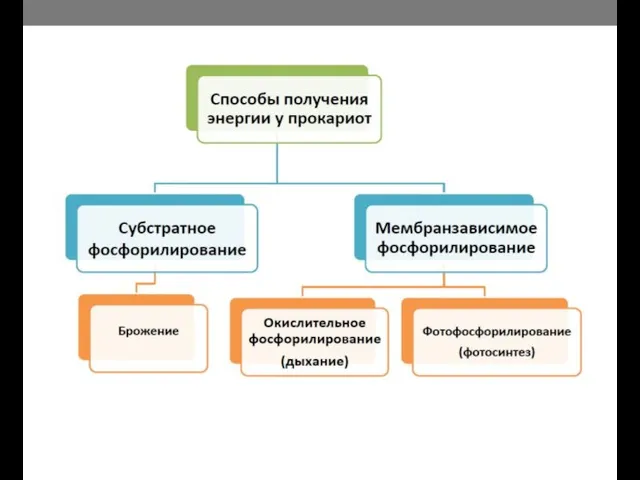
- 11. Схема энергетических процессов у прокариот.
- 13. Символ "~", введенный американским биохимиком Ф. Липманом (F. Lipmann), служит для обозначения макроэргической связи.
- 14. Брожение – наиболее примитивный способ получения энергии Донор и акцептор электронов – органические вещества, т.е. из
- 16. Продукты брожения глюкозы
- 17. Муравьинокислое брожение и семейство Enterobacteriaceae Некоторые микроорганизмы, образующие при брожении кислоты, объединяют в одну физиологическую группу
- 19. В процессе дыхания происходит окисление восстановленных веществ с относительно низким окислительно-восстановительным потенциалом, образующихся в реакциях метаболизма
- 20. Организация дыхательной цепи
- 21. Расположение переносчиков электронов в ЦПМ прокариот таково, что при работе любой электронотранспортной цепи (фотосинтетической или дыхательной)
- 23. Дыхание (продолжение) Освобождающаяся припреносе электронов энергия первоначально запасается в форме ΔμН+ Разрядка ΔμН+ происходит с участием
- 24. Особенности дыхательных цепей прокариот Дыхательная цепь бактерий: Расположена в цитоплазматической мембране Имеет непостоянный состав переносчиков (
- 25. Брожение Процесс протекает в цитоплазме Донор и акцептор электронов – органические метаболиты Анаэробный процесс Синтез АТФ
- 26. Типы анаэробного дыхания у эубактерий Различия дыхательных цепей аэробных и анаэробных бактерий – в конечном акцепторе
- 27. Классификация прокариот по отношению к молекулярному кислороду Облигатные (строгие) аэробы развиваются при наличии в атмосфере 20%
- 28. Облигатные анаэробы — бактерии, для которых наличие молекулярного кислорода является губительным (Bacteroides, Clostridium) Факультативные анаэробы могут
- 29. Молекулярный кислород. Токсичность Как фактор внешней среды О2 воздействует на организмы двояко: с одной стороны, он
- 30. Защитные механизмы клетки от кислорода и его токсических форм. Для нейтрализации токсических форм О2 существующие прокариоты
- 32. Скачать презентацию
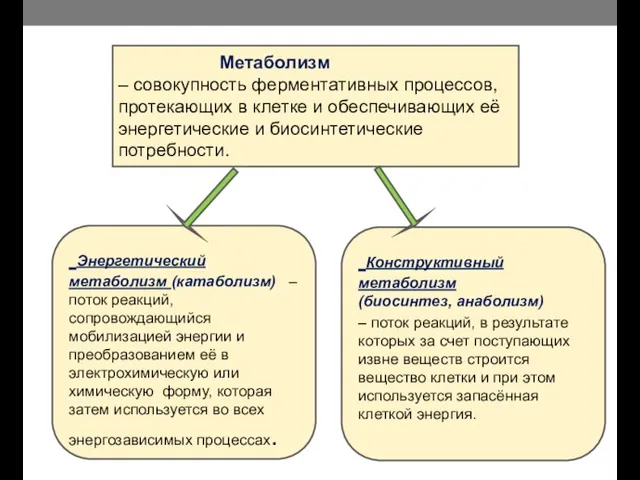
Метаболизм – совокупность ферментативных процессов, протекающих в клетке и
Метаболизм – совокупность ферментативных процессов, протекающих в клетке и

Классификация микроорганизмов
По источнику углерода
Автотрофы Гетеротрофы
СО2 органические соединения
В зависимости от донора электронов
По
Классификация микроорганизмов
По источнику углерода
Автотрофы Гетеротрофы
СО2 органические соединения
В зависимости от донора электронов
По

Факторы роста – вещества, которые прокариоты по каким-либо причинам не
Факторы роста – вещества, которые прокариоты по каким-либо причинам не


Бактерии, изучаемые медицинской микробиологией- хемоорганогетеротрофы
Отличительной особенностью этой группы является то, что
Бактерии, изучаемые медицинской микробиологией- хемоорганогетеротрофы
Отличительной особенностью этой группы является то, что
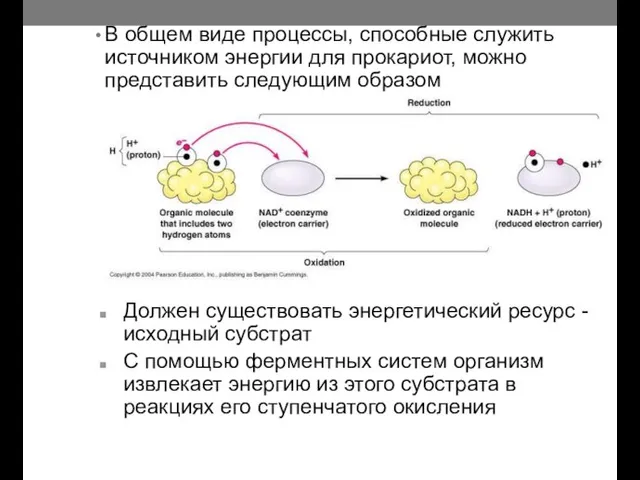
В общем виде процессы, способные служить источником энергии для прокариот, можно
В общем виде процессы, способные служить источником энергии для прокариот, можно

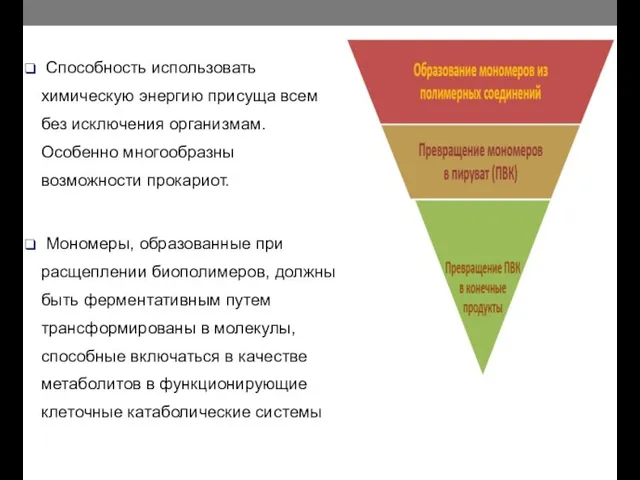
Способность использовать химическую энергию присуща всем без исключения организмам. Особенно
Способность использовать химическую энергию присуща всем без исключения организмам. Особенно

Основные катаболические системы клетки:
гликолиз (путь Эмбдена-Мейергофа-Парнаса),
окислительный пентозофосфатный путь,
Основные катаболические системы клетки:
гликолиз (путь Эмбдена-Мейергофа-Парнаса),
окислительный пентозофосфатный путь,
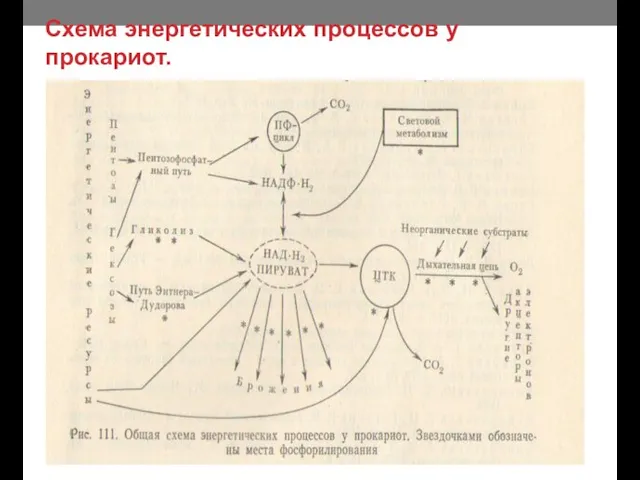
Схема энергетических процессов у прокариот.
Схема энергетических процессов у прокариот.
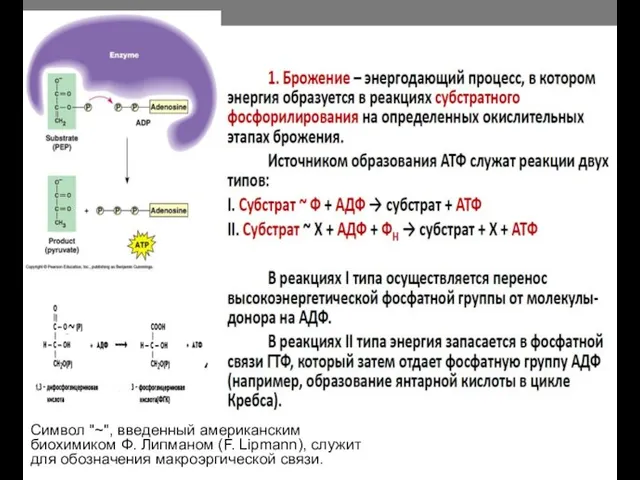
Символ "~", введенный американским биохимиком Ф. Липманом (F. Lipmann), служит для
Символ "~", введенный американским биохимиком Ф. Липманом (F. Lipmann), служит для

Брожение – наиболее примитивный способ получения энергии
Донор и акцептор электронов
Брожение – наиболее примитивный способ получения энергии
Донор и акцептор электронов
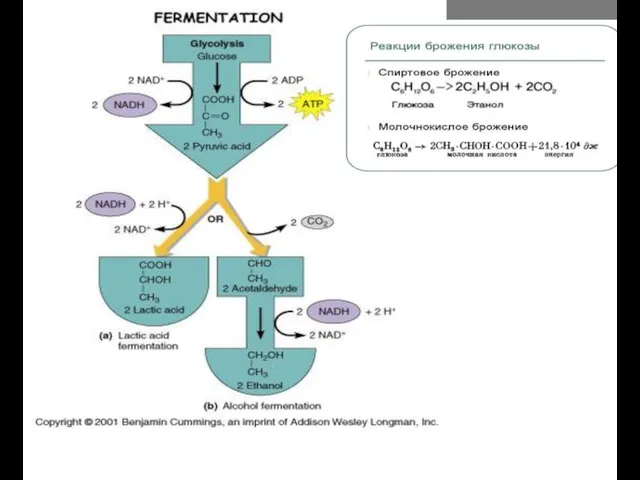
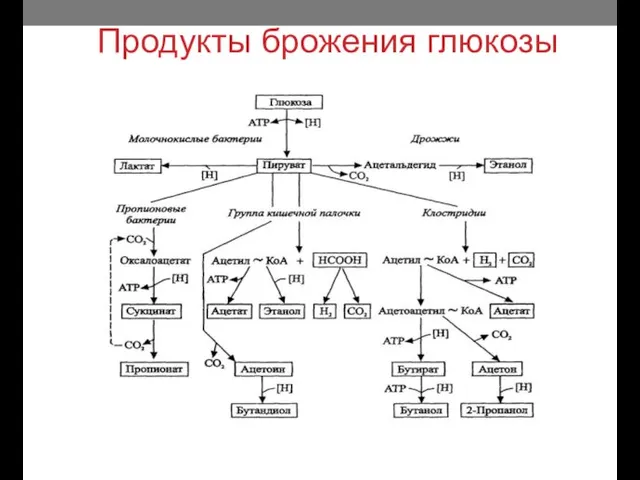
Продукты брожения глюкозы
Продукты брожения глюкозы

Муравьинокислое брожение и семейство Enterobacteriaceae
Некоторые микроорганизмы, образующие при брожении кислоты,
Муравьинокислое брожение и семейство Enterobacteriaceae
Некоторые микроорганизмы, образующие при брожении кислоты,


В процессе дыхания происходит окисление восстановленных веществ с относительно низким
В процессе дыхания происходит окисление восстановленных веществ с относительно низким
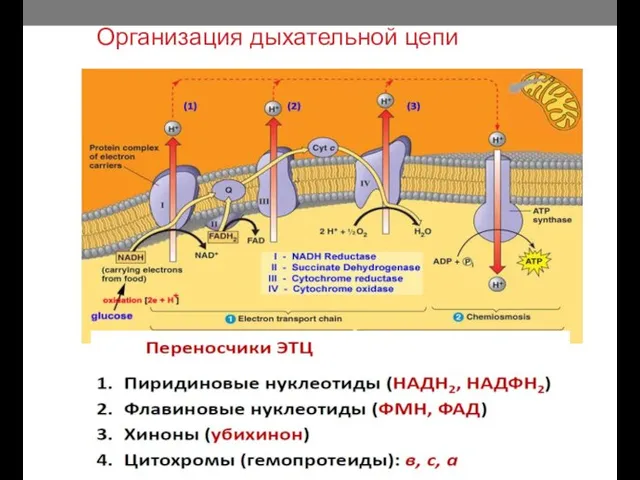
Организация дыхательной цепи
Организация дыхательной цепи
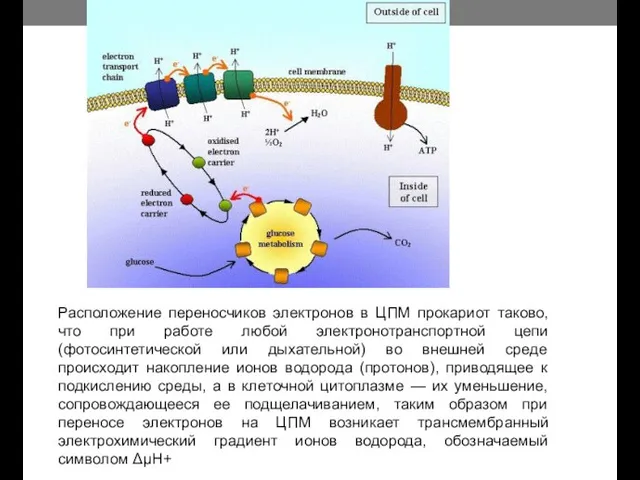
Расположение переносчиков электронов в ЦПМ прокариот таково, что при работе любой
Расположение переносчиков электронов в ЦПМ прокариот таково, что при работе любой


Дыхание (продолжение)
Освобождающаяся припреносе электронов энергия первоначально запасается в форме ΔμН+
Разрядка ΔμН+
Дыхание (продолжение)
Освобождающаяся припреносе электронов энергия первоначально запасается в форме ΔμН+
Разрядка ΔμН+

Особенности дыхательных цепей прокариот
Дыхательная цепь бактерий:
Расположена в цитоплазматической мембране
Имеет
Особенности дыхательных цепей прокариот
Дыхательная цепь бактерий:
Расположена в цитоплазматической мембране
Имеет

Брожение
Процесс протекает в цитоплазме
Донор и акцептор электронов – органические
Брожение
Процесс протекает в цитоплазме
Донор и акцептор электронов – органические

Типы анаэробного дыхания у эубактерий
Различия дыхательных цепей аэробных и анаэробных бактерий
Типы анаэробного дыхания у эубактерий
Различия дыхательных цепей аэробных и анаэробных бактерий
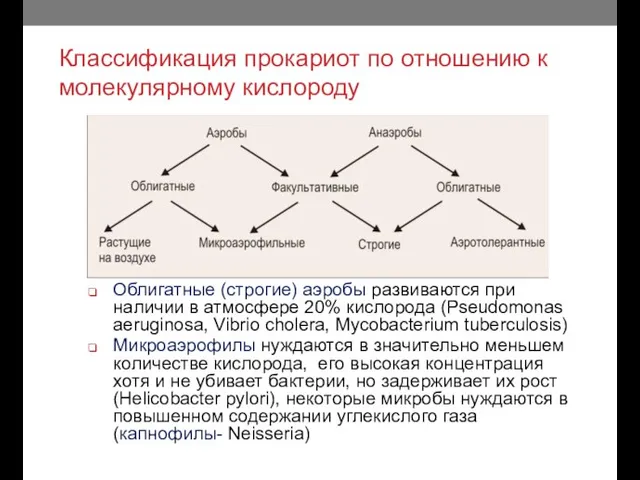
Классификация прокариот по отношению к молекулярному кислороду
Облигатные (строгие) аэробы развиваются при
Классификация прокариот по отношению к молекулярному кислороду
Облигатные (строгие) аэробы развиваются при

Облигатные анаэробы — бактерии, для которых наличие молекулярного кислорода является губительным
Облигатные анаэробы — бактерии, для которых наличие молекулярного кислорода является губительным
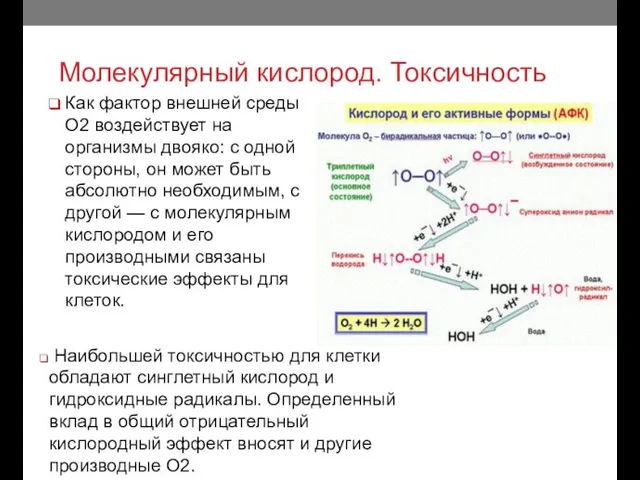
Молекулярный кислород. Токсичность
Как фактор внешней среды О2 воздействует на организмы двояко:
Молекулярный кислород. Токсичность
Как фактор внешней среды О2 воздействует на организмы двояко:

Защитные механизмы клетки от кислорода и его токсических форм.
Для нейтрализации токсических
Защитные механизмы клетки от кислорода и его токсических форм.
Для нейтрализации токсических
 Генетика человека
Генетика человека Танспортные системы организма
Танспортные системы организма Презентация на тему "Клетка" - скачать презентации по Биологии
Презентация на тему "Клетка" - скачать презентации по Биологии Тип Членистоногие. Класс Насекомые
Тип Членистоногие. Класс Насекомые Энергетический обмен
Энергетический обмен Тема «Общая характеристика грибов» Цель:определить место грибов в системе органического мира
Тема «Общая характеристика грибов» Цель:определить место грибов в системе органического мира Flora of the river Vodla
Flora of the river Vodla Назад в прошлое
Назад в прошлое Цікаві факти про «людину» та «їжу»
Цікаві факти про «людину» та «їжу»  Microbiologia si diagnosticul de laborator al infectilor stafilo-si streptococice
Microbiologia si diagnosticul de laborator al infectilor stafilo-si streptococice Период новорожденности
Период новорожденности Витамины
Витамины Правила кормления животных. Виды кормов. Витамины и минеральные добавки
Правила кормления животных. Виды кормов. Витамины и минеральные добавки Царство Грибы
Царство Грибы Биотехнология производства бактериальных удобрений
Биотехнология производства бактериальных удобрений Плоды - яблоко, костянка, сборная семянка, сборная костянка. Плоды - яблоко, костянка, сборная семянка, сборная костянка. В это семей
Плоды - яблоко, костянка, сборная семянка, сборная костянка. Плоды - яблоко, костянка, сборная семянка, сборная костянка. В это семей Презентация на тему "Проблемы селекции собак в свете некоторых положений современной генетики" - скачать презентации по Биол
Презентация на тему "Проблемы селекции собак в свете некоторых положений современной генетики" - скачать презентации по Биол Строение органов дыхания животных
Строение органов дыхания животных Великий круговорот жизни (окружающий мир, 3 класс)
Великий круговорот жизни (окружающий мир, 3 класс) Анализ микробных маркеров (по Осипову)
Анализ микробных маркеров (по Осипову) Болезни овощей
Болезни овощей Животные Южной Америки
Животные Южной Америки Endangered animals
Endangered animals Учебный проект по окружающему миру «Разнообразие животных в природе» 4 класс VIII вида
Учебный проект по окружающему миру «Разнообразие животных в природе» 4 класс VIII вида Презентация на тему "Этапы жизни" - скачать презентации по Биологии
Презентация на тему "Этапы жизни" - скачать презентации по Биологии Мыс Мартьян
Мыс Мартьян Многообразие водорослей их роль в природе практическое значение.
Многообразие водорослей их роль в природе практическое значение.  Энергетический обмен. (Лекция 4)
Энергетический обмен. (Лекция 4)