Продолжительность жизни животных в единицах физического и биологического времени А.Ф. Алимов, Т.И. Казанцева Зоологический инсти
- Продолжительность жизни животных в единицах физического и биологического времени А.Ф. Алимов, Т.И. Казанцева Зоологический инсти

Содержание
- 2. При рассмотрении понятия времени применительно к живым организмам возникают три главных вопроса: Какова максимально возможная продолжительность
- 3. Мы попытались рассмотреть эти вопросы для животных, непрерывно растущих в течение всей жизни, и животных, прекращающих
- 4. Физическое и биологическое время Физическим временем будем называть время, в котором мы измеряем длительности различных событий,
- 5. Биологические системы существуют благодаря специфическим процессам, протекающим в них с определенными скоростями. Каждый из этих процессов
- 6. Проблема биологического времени разрабатывается в разных аспектах со второй половины XIX века. Рассматриваются различные биологические системы
- 7. Нас интересует то «собственное» время, в котором существует организм как целое. Поэтому за основу мы взяли
- 8. Общепризнано, что морфологические и физиологические свойства животного тесно связаны с массой тела M. Описано множество аллометрических
- 9. Известна единственная попытка определения внутреннего времени через процесс роста массы организма (Backman G. Wachstum und organische
- 10. Бакман выделяет три цикла роста многоклеточного организма: овулярный, эмбриональный и период основного роста. «Органическое» время определяется
- 11. Метаболизм Общепризнанным является способ определения внутреннего (физиологического) времени организма через удельную скорость метаболизма. Наиболее строгое определение
- 12. Из этого определения следует: Физиологическим временем названо количество энергии, использованной единицей массы организма за некоторый интервал
- 13. Высказано и активно обсуждается предположение, что суммарное за время жизни количество энергии, использованное единицей массы, определяет
- 14. Определение единицы внутреннего времени через скорость роста массы организма Обозначим: – функция изменения массы организма с
- 15. Величина единицы внутреннего времени не постоянна. Она растет с увеличением возраста организма, измеренного в единицах физического
- 16. За одну единицу физического времени единица массы прирастает на cm(t) единиц массы. Поэтому одна единица физического
- 17. Продолжительность жизни рыб в единицах τ(t), рассчитанная на основе процесса роста массы Уравнение Берталанфи (Bertalanffy, 1942;
- 18. Оценка tmax для рыб tmax оценивается как вторая точка перегиба кривой скорости роста массы (m’(t)) (Алимов
- 19. Точность оценки параметров кривой Берталанфи по эмпирической выборке низка, однако величина Θm оценивается значительно точнее. единиц
- 20. Результаты расчетов величины Θm [ед. внутр. времени] для некоторых популяций рыб из разных водоемов (m0, M∞,
- 21. Общий аллометрический закон: Продолжительность жизни в единицах физического времени tmax эквивалентна М1/4. Для рассмотренных видов рыб
- 22. Определение единицы внутреннего времени для животных, прекращающих рост, на основе процесса метаболизма Обозначим: M – масса
- 23. Количество энергии, усвоенной за интервал времени [t1, t2], численно равно длине этого интервала в единицах внутреннего
- 24. Определение q(t) и tmax для птиц Основание: R.E. Ricklefs “Intrinsic aging-related mortality in birds” // J.
- 25. Гипотеза: Внутренней причиной роста смертности с возрастом является уменьшение удельной скорости метаболизма. Нами предложена формула зависимости
- 26. Аппроксимация зависимостей ω(M) и (qcrit/q0)(M) Все оценки получены на основе массива данных из работы Рикклефса, который
- 27. Для оценки качества расчетов tmax по полученной формуле мы применили ее к 33 видам воробьиных и
- 28. Пример результатов расчета tmax (годы) по нашей формуле (*) и формулам других авторов (номера формул указаны
- 29. Расчет максимальной продолжительности жизни в единицах внутреннего времени (θq) для птиц q0 tM q0 tM В
- 30. Удельная скорость обмена выражена в [ккал/г/год]; Результаты расчетов Значения продолжительности жизни в единицах внутреннего времени для
- 31. Предположения, лежащие в основе всего сказанного выше, имеют общий характер. Поэтому данный подход можно применить к
- 32. Выводы Предложены способы определения максимальной продолжительности жизни животных ( tmax ), учитывающие особенности их роста и
- 34. Скачать презентацию

При рассмотрении понятия времени применительно к живым организмам возникают три главных
При рассмотрении понятия времени применительно к живым организмам возникают три главных

Мы попытались рассмотреть эти вопросы для животных, непрерывно растущих в течение
Мы попытались рассмотреть эти вопросы для животных, непрерывно растущих в течение

Физическое и биологическое время
Физическим временем будем называть время, в котором мы
Физическое и биологическое время
Физическим временем будем называть время, в котором мы

Биологические системы существуют благодаря специфическим процессам, протекающим в них с определенными
Биологические системы существуют благодаря специфическим процессам, протекающим в них с определенными

Проблема биологического времени разрабатывается в разных аспектах со второй половины XIX
Проблема биологического времени разрабатывается в разных аспектах со второй половины XIX

Нас интересует то «собственное» время, в котором существует организм как целое.
Нас интересует то «собственное» время, в котором существует организм как целое.

Общепризнано, что морфологические и физиологические свойства животного тесно связаны с массой
Общепризнано, что морфологические и физиологические свойства животного тесно связаны с массой

Известна единственная попытка определения внутреннего времени через процесс роста массы организма
Известна единственная попытка определения внутреннего времени через процесс роста массы организма

Бакман выделяет три цикла роста многоклеточного организма: овулярный, эмбриональный и период
Бакман выделяет три цикла роста многоклеточного организма: овулярный, эмбриональный и период

Метаболизм
Общепризнанным является способ определения внутреннего (физиологического) времени организма через удельную скорость
Метаболизм
Общепризнанным является способ определения внутреннего (физиологического) времени организма через удельную скорость

Из этого определения следует:
Физиологическим временем названо количество энергии, использованной единицей массы
Из этого определения следует:
Физиологическим временем названо количество энергии, использованной единицей массы

Высказано и активно обсуждается предположение, что суммарное за время жизни
Высказано и активно обсуждается предположение, что суммарное за время жизни

Определение единицы внутреннего времени через скорость роста массы организма
Обозначим:
– функция изменения
Определение единицы внутреннего времени через скорость роста массы организма
Обозначим:
– функция изменения

Величина единицы внутреннего времени не постоянна. Она растет с увеличением возраста
Величина единицы внутреннего времени не постоянна. Она растет с увеличением возраста

За одну единицу физического времени единица массы прирастает на cm(t) единиц
За одну единицу физического времени единица массы прирастает на cm(t) единиц
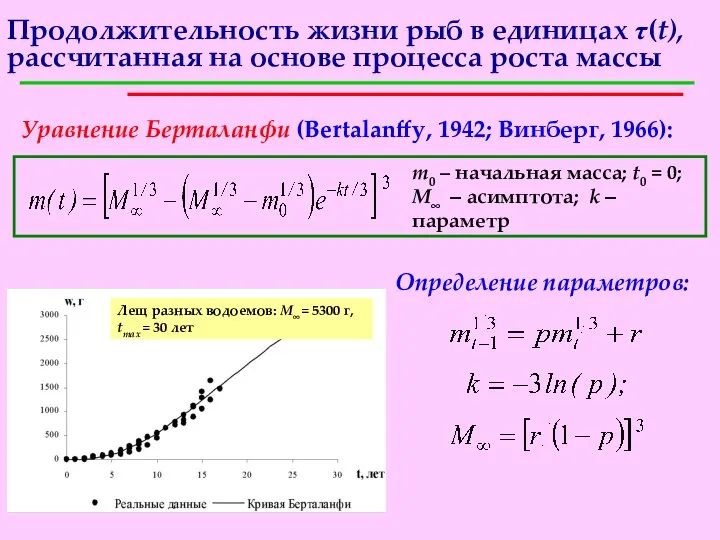
Продолжительность жизни рыб в единицах τ(t), рассчитанная на основе процесса роста
Продолжительность жизни рыб в единицах τ(t), рассчитанная на основе процесса роста
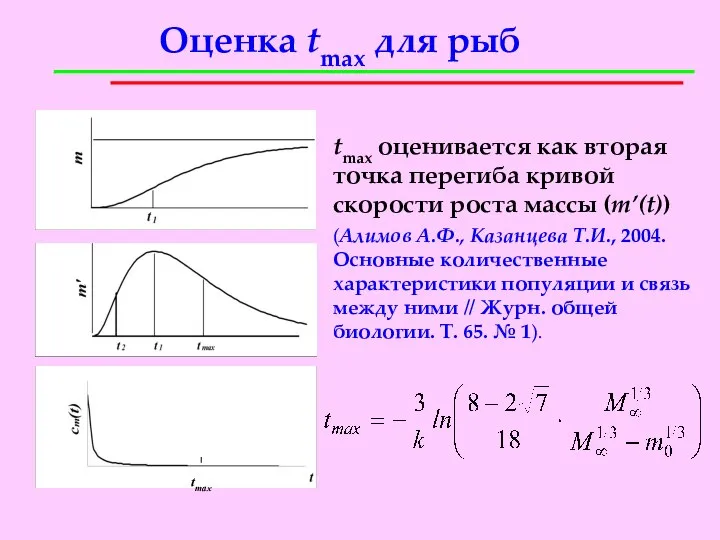
Оценка tmax для рыб
tmax оценивается как вторая точка перегиба кривой скорости
Оценка tmax для рыб
tmax оценивается как вторая точка перегиба кривой скорости
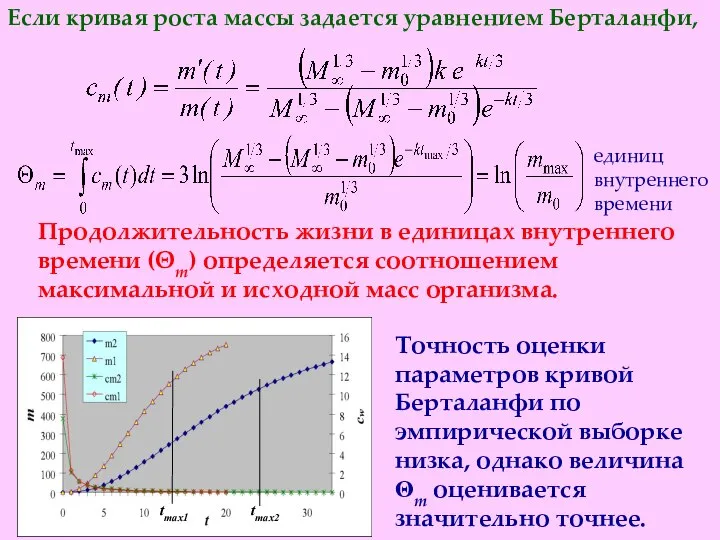
Точность оценки параметров кривой Берталанфи по эмпирической выборке низка, однако величина
Точность оценки параметров кривой Берталанфи по эмпирической выборке низка, однако величина
![Результаты расчетов величины Θm [ед. внутр. времени] для некоторых популяций рыб](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1187567/slide-19.jpg)
Результаты расчетов величины Θm [ед. внутр. времени] для некоторых популяций рыб
Результаты расчетов величины Θm [ед. внутр. времени] для некоторых популяций рыб
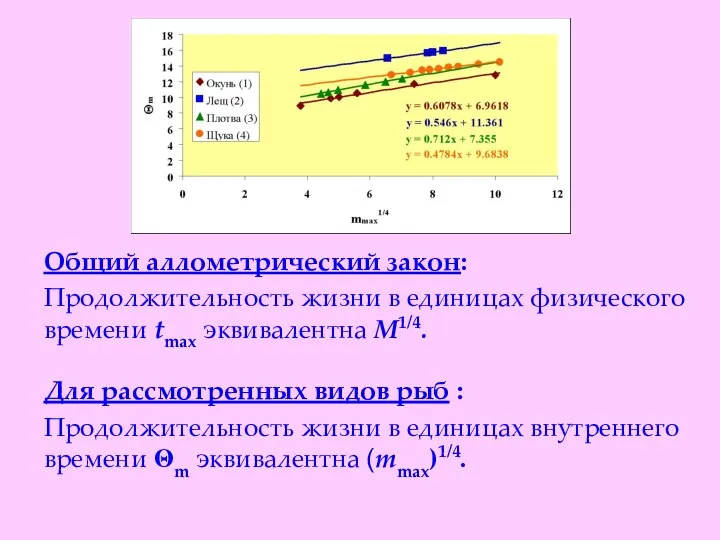
Общий аллометрический закон:
Продолжительность жизни в единицах физического времени tmax эквивалентна
Общий аллометрический закон:
Продолжительность жизни в единицах физического времени tmax эквивалентна

Определение единицы внутреннего времени для животных, прекращающих рост, на основе процесса
Определение единицы внутреннего времени для животных, прекращающих рост, на основе процесса
![Количество энергии, усвоенной за интервал времени [t1, t2], численно равно длине](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1187567/slide-22.jpg)
Количество энергии, усвоенной за интервал времени [t1, t2], численно равно длине
Количество энергии, усвоенной за интервал времени [t1, t2], численно равно длине

Определение q(t) и tmax для птиц
Основание: R.E. Ricklefs “Intrinsic aging-related mortality
Определение q(t) и tmax для птиц
Основание: R.E. Ricklefs “Intrinsic aging-related mortality

Гипотеза: Внутренней причиной роста смертности с возрастом является уменьшение удельной скорости
Гипотеза: Внутренней причиной роста смертности с возрастом является уменьшение удельной скорости
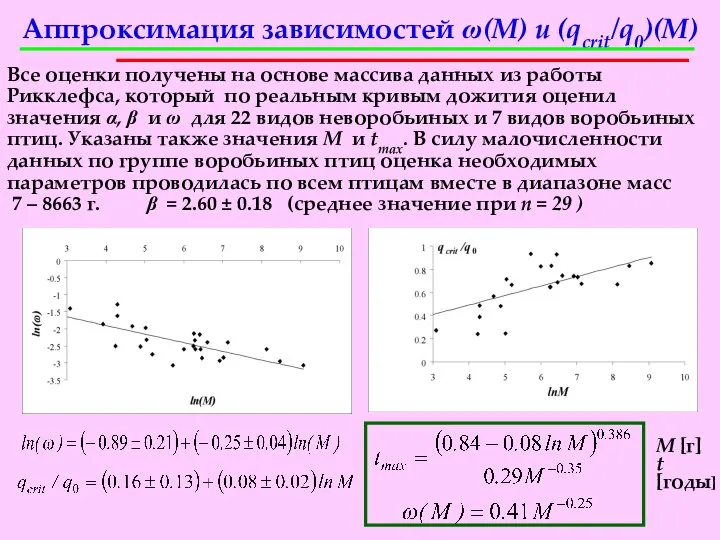
Аппроксимация зависимостей ω(M) и (qcrit/q0)(M)
Все оценки получены на основе массива данных
Аппроксимация зависимостей ω(M) и (qcrit/q0)(M)
Все оценки получены на основе массива данных

Для оценки качества расчетов tmax по полученной формуле мы применили ее
Для оценки качества расчетов tmax по полученной формуле мы применили ее

Пример результатов расчета tmax (годы) по нашей формуле (*) и формулам
Пример результатов расчета tmax (годы) по нашей формуле (*) и формулам

Расчет максимальной продолжительности жизни в единицах внутреннего времени (θq) для птиц
q0
tM
q0
tM
В
Расчет максимальной продолжительности жизни в единицах внутреннего времени (θq) для птиц
q0
tM
q0
tM
В
![Удельная скорость обмена выражена в [ккал/г/год]; Результаты расчетов Значения продолжительности жизни](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1187567/slide-29.jpg)
Удельная скорость обмена выражена в [ккал/г/год];
Результаты расчетов
Значения продолжительности жизни в
Удельная скорость обмена выражена в [ккал/г/год];
Результаты расчетов
Значения продолжительности жизни в

Предположения, лежащие в основе всего сказанного выше, имеют общий характер. Поэтому
Предположения, лежащие в основе всего сказанного выше, имеют общий характер. Поэтому

Выводы
Предложены способы определения максимальной продолжительности жизни животных ( tmax ), учитывающие
Выводы
Предложены способы определения максимальной продолжительности жизни животных ( tmax ), учитывающие
 Хвойные растения
Хвойные растения  Хоровод цветов (игра по мотивам телевизионной передачи Своя игра)
Хоровод цветов (игра по мотивам телевизионной передачи Своя игра) Жасмонаты. Жасмоновая кислота
Жасмонаты. Жасмоновая кислота Кайнозойская эра
Кайнозойская эра Глотка, пищевод и желудок
Глотка, пищевод и желудок Викторина «МЛЕКОПИТАЮЩИЕ»
Викторина «МЛЕКОПИТАЮЩИЕ»  Хрящевые рыбы
Хрящевые рыбы Презентация на тему "Захворювання органів дихання" - скачать бесплатно презентации по Биологии
Презентация на тему "Захворювання органів дихання" - скачать бесплатно презентации по Биологии Суставы, типы суставов у животных. Виды соединения суставов конечностей
Суставы, типы суставов у животных. Виды соединения суставов конечностей Физиология зрительного анализатора
Физиология зрительного анализатора Вебинар. Строение клетки
Вебинар. Строение клетки Это орган растения, который ему жизненно необходим. Это орган растения, который ему жизненно необходим. Этот орган появляется
Это орган растения, который ему жизненно необходим. Это орган растения, который ему жизненно необходим. Этот орган появляется  Отделы растений. Покрытосеменные. 5 класс
Отделы растений. Покрытосеменные. 5 класс Удивительные природные материалы
Удивительные природные материалы Презентация на тему ДНК и РНК
Презентация на тему ДНК и РНК Презентация на тему Внутренняя среда организма
Презентация на тему Внутренняя среда организма Растения
Растения Препараты по гистологии
Препараты по гистологии Презентация на тему "Голосеменные растения 7 класс" - скачать презентации по Биологии
Презентация на тему "Голосеменные растения 7 класс" - скачать презентации по Биологии Презентация открытого урока по теме «Рельеф Мордовии» Автор: Честнова Ольга Валерьевна учитель географии средней школы №39
Презентация открытого урока по теме «Рельеф Мордовии» Автор: Честнова Ольга Валерьевна учитель географии средней школы №39 ТЕМА: TROCHOZOA (схема развития трохофоры) и олигомерные трохофорные
ТЕМА: TROCHOZOA (схема развития трохофоры) и олигомерные трохофорные  Значение бактерий
Значение бактерий Презентация на тему Зоология – наука о животных 7 класс.
Презентация на тему Зоология – наука о животных 7 класс. КОЛОКОЛА ТРЕВОГИ
КОЛОКОЛА ТРЕВОГИ Тип плоские черви Многообразие плоских червей Выполнила учитель биологии Пискунова О.А. МОУ СОШ №3
Тип плоские черви Многообразие плоских червей Выполнила учитель биологии Пискунова О.А. МОУ СОШ №3  Презентация по экологии Оценка пищевого статуса при развитии адаптационной резистентности организма.
Презентация по экологии Оценка пищевого статуса при развитии адаптационной резистентности организма.  Законы России об охране животного мира. Система мониторинга
Законы России об охране животного мира. Система мониторинга Взаимоотношения между организмами
Взаимоотношения между организмами