- Репликация и экспрессии геномов ДНК-вирусов бактерий и эукариот

Содержание
- 4. Роджер и Артур Корнберги
- 5. ДНК-вирусы растений. Каулимовирусы.
- 10. Каулимовирусы (Caulimoviruses)
- 14. ДНК-вирусы растений. Геминивирусы.
- 15. Bean golden mosaic virus genus
- 19. Banana bunchy top virus
- 20. Faba bean necrotic yellows virus
- 23. Репликация и экспрессии геномов ДНК-вирусов бактерий и эукариот
- 25. Роджер и Артур Корнберги
- 27. Бактериофаги
- 28. РНК-содержащие бактериофаги
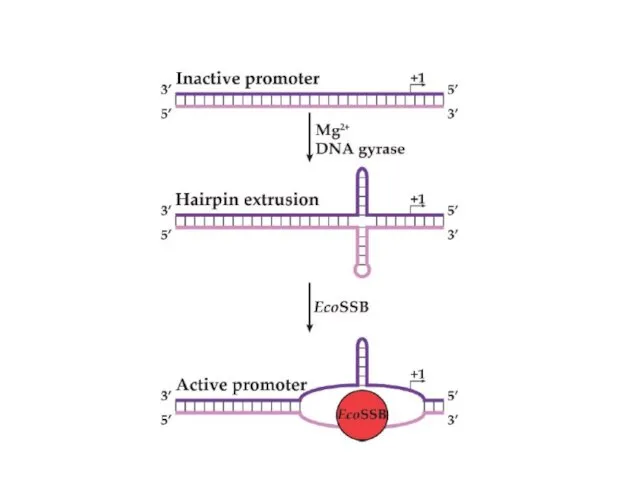
- 29. Механизмы синтеза РНК довольно консервативны и однообразны среди вирусов. Однако, большое разнообразие наблюдается в механизмах регуляции
- 30. Принципы регуляции транскрипции: 1. Системность. Один механизм регуляции транскрипции может контролировать экспрессию группы генов. 2. Каскадность.
- 31. Как правило, регулируемыми стадиями транскрипции являются инициация и терминация. В принципе, в некоторых системах может регулироваться
- 32. Можно различить несколько уровней сложности систем транскрипции фаговых геномов в зависимости от степени модификации транскрипционного аппарата
- 33. Бактериальная ДНК-зависимая РНК-полимераза включает “коровую“ часть, включающую две α субъединицы, β- и β’-субъединицы, а также ω-субъединицу.
- 34. Частота инициации зависит от силы промотора. Чем сильнее промотор, тем выше сродство к нему полимеразы, а,
- 35. В комплексе с другими субъединицами σ-фактор, включающий 613 аминокислотных остатков имеет не глобулярную, а вытянутую форму.
- 36. Домены, расположенные на дистальных концах σ-субъединицы, узнают два элемента промотора. Стандартный промотор РНК-полимеразы включает в себя
- 37. Сродство промотора и σ-субъединицы зависит прежде всего от оптимального расстояния между консенсусными элементами и их первичной
- 38. РНК-полимераза связывает промотор поэтапно. Сначала формируется достаточно слабый так называемый «закрытый» комплекс, который спонтанно изомеризуется с
- 39. Терминация транскрипции. У прокариот имеется два механизма регуляции терминации транскрипции. Это ρ (Rho)-зависимая терминация и ρ-независимая
- 41. Итак, комбинацией промоторов и терминаторов различной силы можно регулировать количество синтезируемой мРНК, а, значит, и количество
- 42. Бактериофаги
- 44. В геноме фагов есть две транскрипционные единицы. Одна из транскрипционных единиц содержит белки, нужные в достаточно
- 45. Такой принцип количественной транскрипционной регуляции называется каскадным (не путать с темпоральной каскадной регуляцией). Возможно, еще более
- 47. Итак, фаг φХ174 дает нам прекрасный пример позитивной каскадной количественной регуляции, когда экспрессия гена возрастает, если
- 48. Регуляция экспрессии генов фага λ Фаг λ - Умеренный бактериофаг 48,502 bp Система регуляции транскрипции фага
- 49. Жизненный цикл литических фагов
- 50. Явление лизогении
- 51. Индукция профага
- 52. Устройство генома лямбдоидных фагов
- 53. БАКТЕРИОФАГ ЛЯМБДА
- 54. На ранней стадии происходит активация транскрипции с помощью клеточных белков. Белок DnaA, участвующий в инициации репликации
- 55. С промотора PL происходит транскрипция гена N. Белок N – это полипептид массой 12 кДа, выполняющий
- 56. Мотивы nut состоят из последовательности box A длиной 8 н.т., спейсера длиной 9-10 н.т. и boxB,
- 58. Важнейшая роль на стадии выбора пути инфекции принадлежит белку CII. Основные способы регуляции процесса связаны с
- 59. В клетке активность системы Hfl регулируется системой катаболитной репрессии, включающей сAMP и CRP (сAMP-responsive protein). Эта
- 60. Белок CII длиной 97 аминокислот действует как транскрипционный фактор, открывающий новые промоторы. CII узнает последовательности ДНК
- 62. Белок CII длиной 97 аминокислот действует как транскрипционный фактор, открывающий новые промоторы. CII узнает последовательности ДНК
- 63. БАКТЕРИОФАГ ЛЯМБДА
- 64. Нужны внешние воздействия для выхода фага из лизогенного состояния. Таким событием, например, может быть повреждение ДНК
- 65. БАКТЕРИОФАГ ЛЯМБДА
- 66. Белок cro длиной 66 остатков, содержит только один домен и узнает те же операторные участки, что
- 67. Белок Q, как и N, является антитерминатором транскрипции и абсолютно необходим для литического пути развития. Он
- 68. БАКТЕРИОФАГ ЛЯМБДА
- 69. Сборка головки фага
- 70. Сборка хвостового отростка
- 71. БАКТЕРИОФАГ Т4
- 72. Гены Т4 делятся на истинно ранние, ранние, средние и поздние. Около 40 промоторов ранних генов узнаются
- 73. Среди продуктов истинно ранних генов фага обнаруживаются еще две АДФ-рибозил-трансферазы. Соответствующие гены называются ModA (23 кДа)
- 74. Для транскрипции средних генов нужны вирусные белки Asi A (antisigma) и Mot (Modification of transcription). Asi
- 76. Промоторы поздних генов содержат элементы, существенно отличающие их от клеточных промоторов. Для их транскрипции нужны следующие
- 77. Известно, что gp55 является фактором, обеспечивающим весьма слабое сродство к ДНК по сравнению с σ70-субъединицей и
- 78. Alt (ModA, ModB) АДФ-рибозилтрансферазы для α-субъединиц РНК-полимеразы Asi A (antisigma), Mot Mod - АДФ-рибозилтрансфераза
- 79. БАКТЕРИОФАГ Т7
- 80. Все гены фага Т7 транскрибируются с одной и той же цепи ДНК слева направо. Ген 0.3
- 81. Первоначально в клетку бактерии попадает левый конец генома размером около 850 пар оснований. Этот район включает
- 82. Ранние транскрипты у фага Т7 подвергаются процессингу РНКазой III. Точек разрезания в ранней области генома имеется
- 83. Важнейшую роль в инфекционном цикле фага играет ген 0.3, который экспрессируется в ходе инфекции первым. Его
- 84. Пять фагоспецифических промоторов для генов класса III являются гораздо более сильными, чем промоторы класса II. На
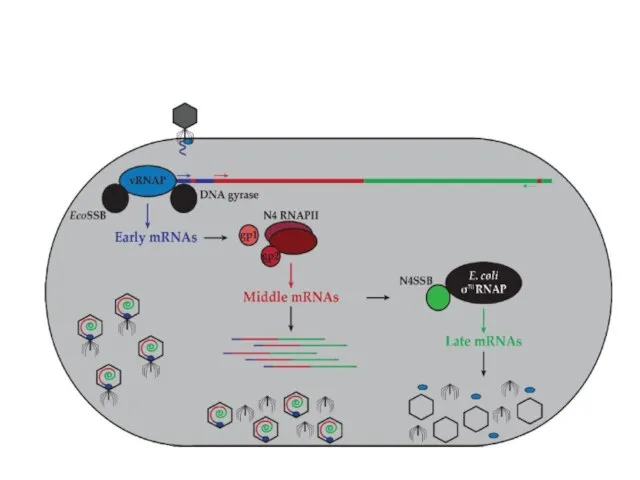
- 85. Регуляция транскрипции у фага N4 Геном этого фага представлен линейной двуцепочечной ДНК длиной 72 000 п.н.
- 88. Средние гены транскрибируются N4 РНК-полимеразой II, являющейся продуктом ранних генов и состоящей из двух белков gр15
- 90. Поздние гены транскрибируются клеточной σ70-содержащей РНК-полимеразой. Однако, поздние промоторы N4 проявляют весьма слабую гомологию консенсусам –10
- 93. ДНК полимераза III E.coli Фермент состоит из нескольких субъединиц и представляет собой гетероолигомер. Состоит из трех
- 95. Enterobacteria phage P2 33.5 Kbp
- 96. Репликация ДНК бактериофага λ Инициация репликации. Участок инициации репликации Ori первоначально узнается вирусным белком – О-белком
- 97. Если ассоциация белка О с комплексом DnaB-Р-белок не произойдет (в случае недостатка DnaB в клетке или
- 99. В ходе двунаправленной репликации по схеме Кернса в клетках, зараженных фагом λ образуются около 50 молекул
- 100. Созревание ДНК-конкатемеров В итоге поздней репликации образуются конкатемерные молекулы, которые надо разрезать на мономеры так, чтобы
- 101. Bacillus subtilis bacteriophage Φ29 21.1 Kbp
- 104. T7 phage
- 108. T7 phage
- 113. T4 bacteriophage
- 115. Скачать презентацию


Роджер и Артур Корнберги
Роджер и Артур Корнберги
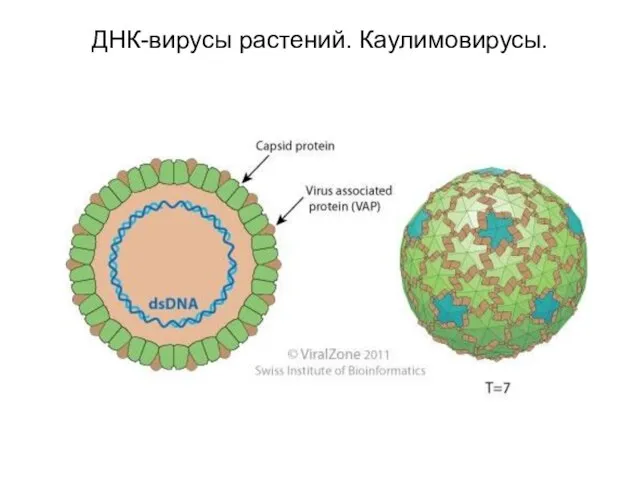
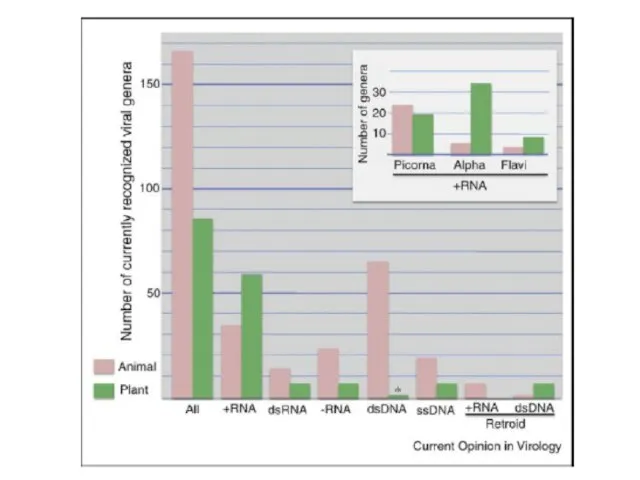
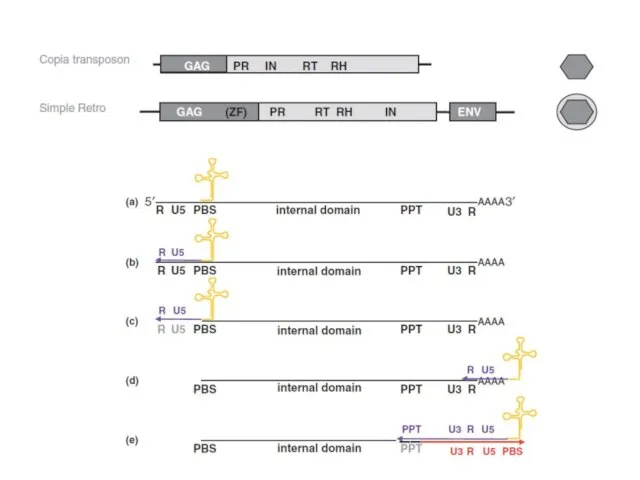
ДНК-вирусы растений. Каулимовирусы.
ДНК-вирусы растений. Каулимовирусы.
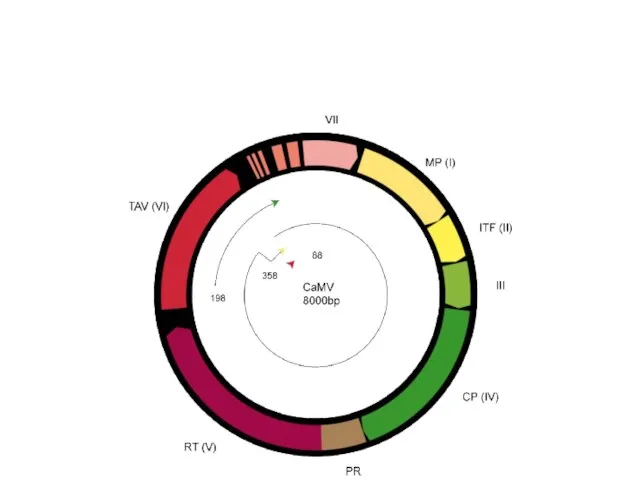
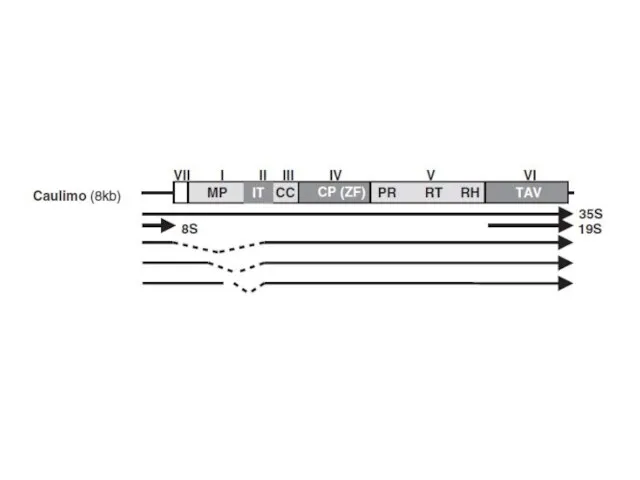


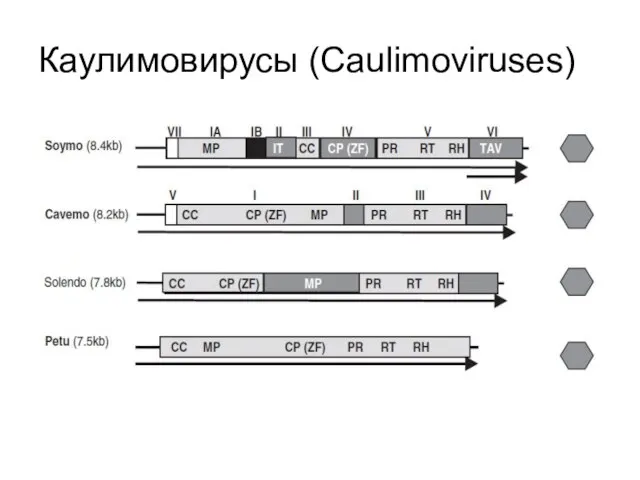
Каулимовирусы (Caulimoviruses)
Каулимовирусы (Caulimoviruses)


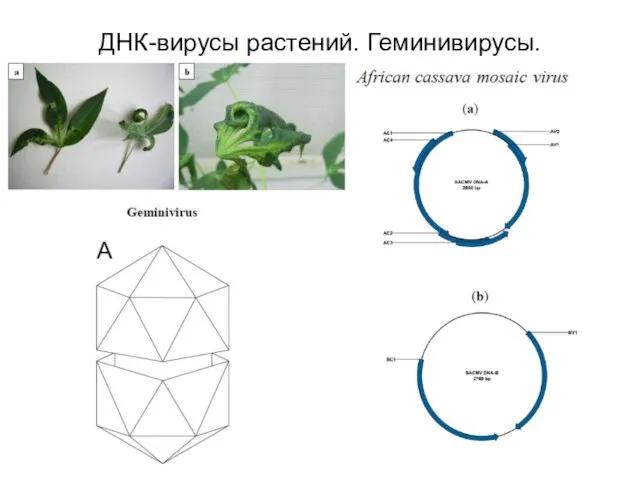
ДНК-вирусы растений. Геминивирусы.
ДНК-вирусы растений. Геминивирусы.
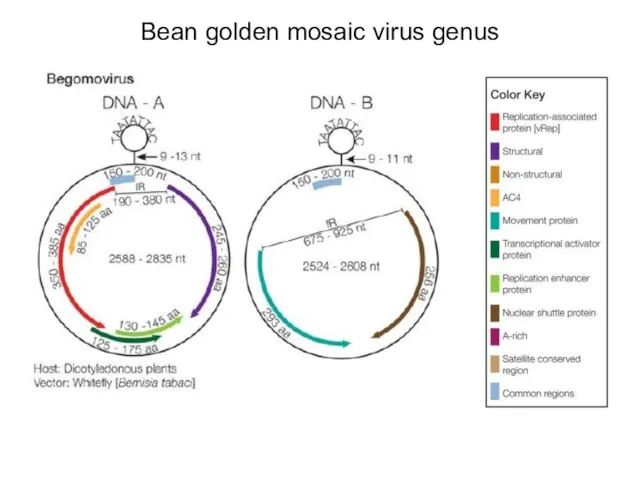
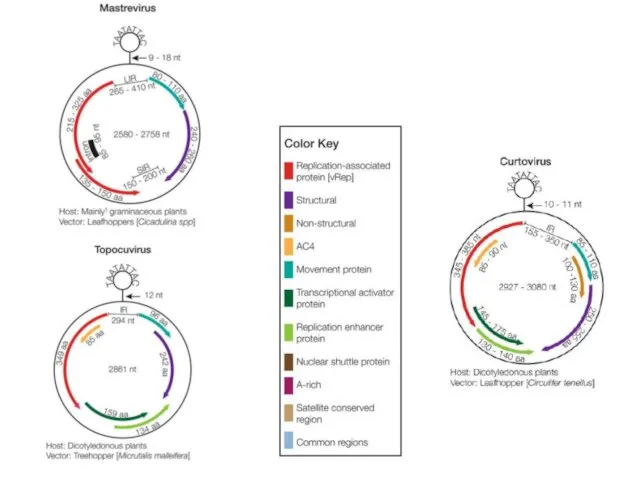
Bean golden mosaic virus genus
Bean golden mosaic virus genus


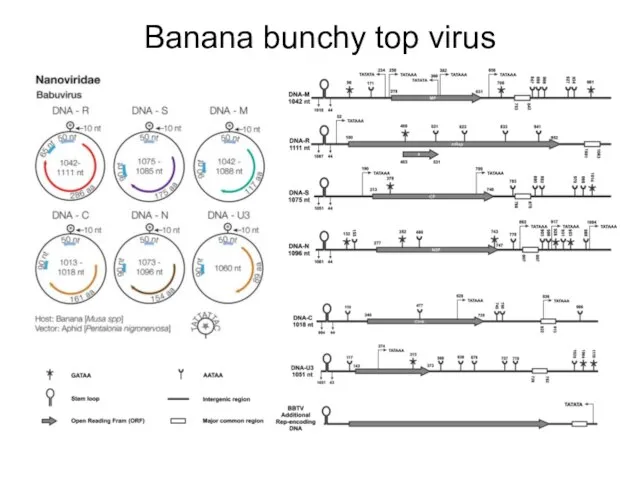
Banana bunchy top virus
Banana bunchy top virus

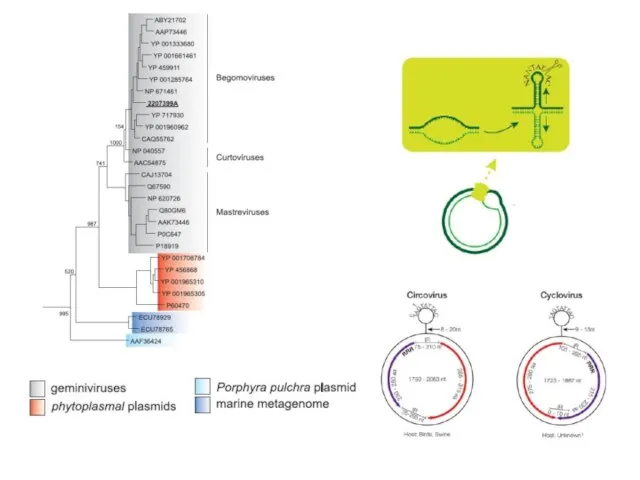
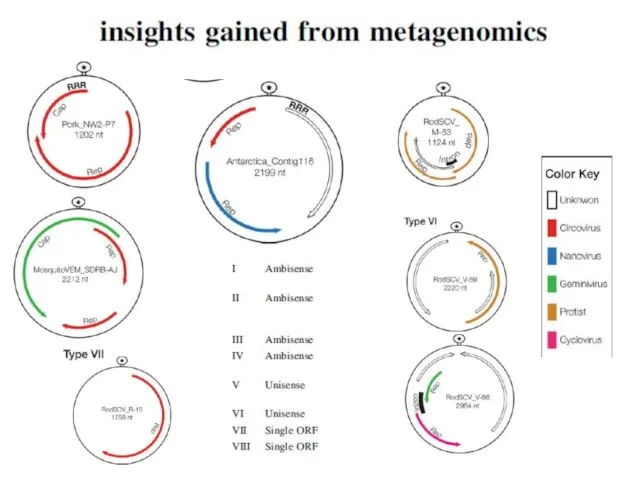
Faba bean necrotic yellows virus
Faba bean necrotic yellows virus

Репликация и экспрессии геномов ДНК-вирусов бактерий и эукариот
Репликация и экспрессии геномов ДНК-вирусов бактерий и эукариот


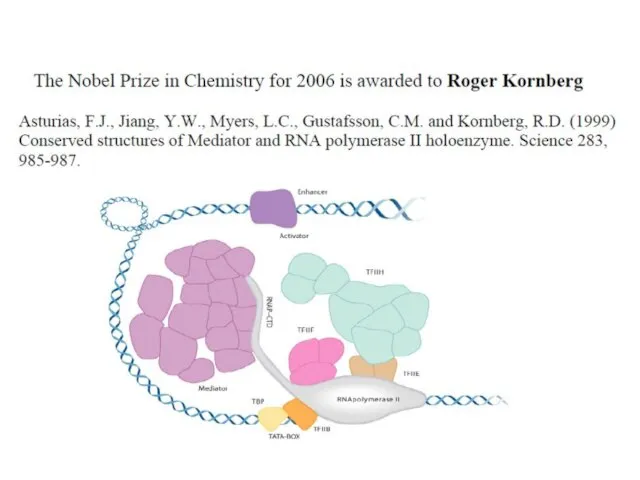
Роджер и Артур Корнберги
Роджер и Артур Корнберги
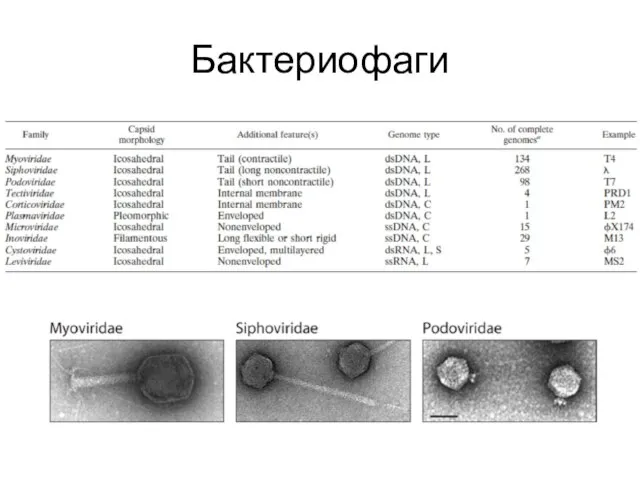
Бактериофаги
Бактериофаги

РНК-содержащие бактериофаги
РНК-содержащие бактериофаги

Механизмы синтеза РНК довольно консервативны и однообразны среди вирусов. Однако, большое
Механизмы синтеза РНК довольно консервативны и однообразны среди вирусов. Однако, большое

Принципы регуляции транскрипции:
1. Системность. Один механизм регуляции транскрипции может контролировать экспрессию
Принципы регуляции транскрипции:
1. Системность. Один механизм регуляции транскрипции может контролировать экспрессию

Как правило, регулируемыми стадиями транскрипции являются инициация и терминация. В
Как правило, регулируемыми стадиями транскрипции являются инициация и терминация. В

Можно различить несколько уровней сложности систем транскрипции фаговых геномов в зависимости
Можно различить несколько уровней сложности систем транскрипции фаговых геномов в зависимости
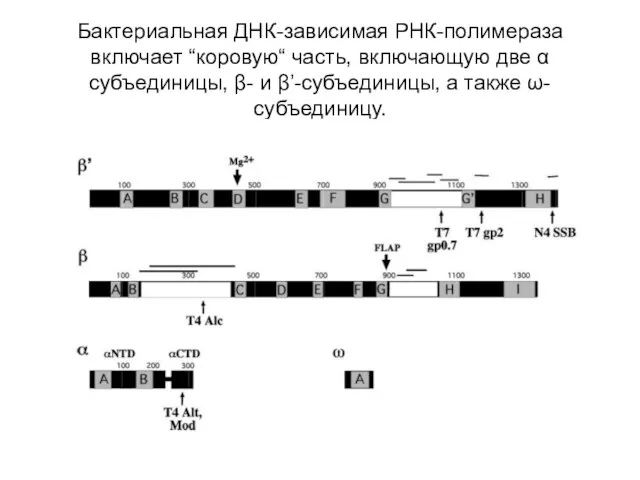
Бактериальная ДНК-зависимая РНК-полимераза
включает “коровую“ часть, включающую две α субъединицы, β- и
Бактериальная ДНК-зависимая РНК-полимераза включает “коровую“ часть, включающую две α субъединицы, β- и

Частота инициации зависит от силы промотора. Чем сильнее промотор, тем выше
Частота инициации зависит от силы промотора. Чем сильнее промотор, тем выше

В комплексе с другими субъединицами σ-фактор, включающий 613 аминокислотных остатков имеет
В комплексе с другими субъединицами σ-фактор, включающий 613 аминокислотных остатков имеет
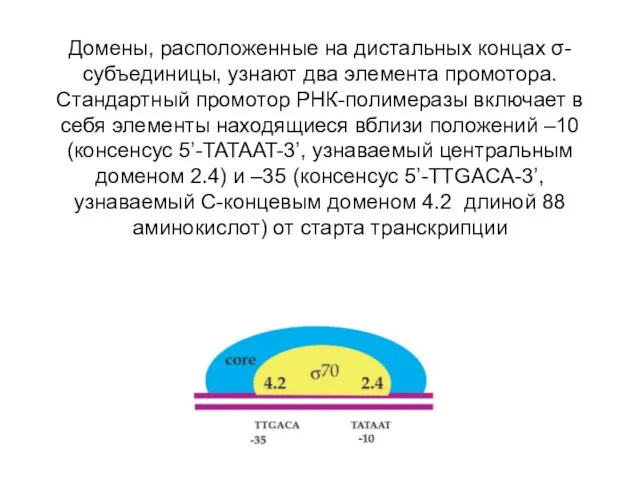
Домены, расположенные на дистальных концах σ-субъединицы, узнают два элемента промотора. Стандартный
Домены, расположенные на дистальных концах σ-субъединицы, узнают два элемента промотора. Стандартный

Сродство промотора и σ-субъединицы зависит прежде всего от оптимального расстояния между
Сродство промотора и σ-субъединицы зависит прежде всего от оптимального расстояния между

РНК-полимераза связывает промотор поэтапно. Сначала формируется достаточно слабый так называемый
РНК-полимераза связывает промотор поэтапно. Сначала формируется достаточно слабый так называемый

Терминация транскрипции.
У прокариот имеется два механизма регуляции терминации транскрипции. Это ρ
Терминация транскрипции.
У прокариот имеется два механизма регуляции терминации транскрипции. Это ρ
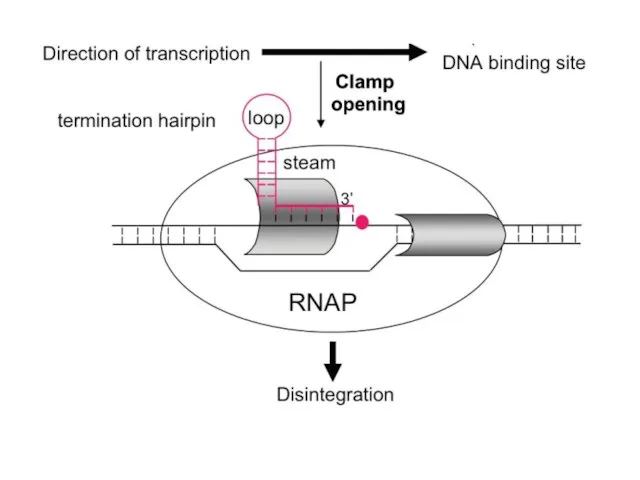

Итак, комбинацией промоторов и терминаторов различной силы можно регулировать количество синтезируемой
Итак, комбинацией промоторов и терминаторов различной силы можно регулировать количество синтезируемой
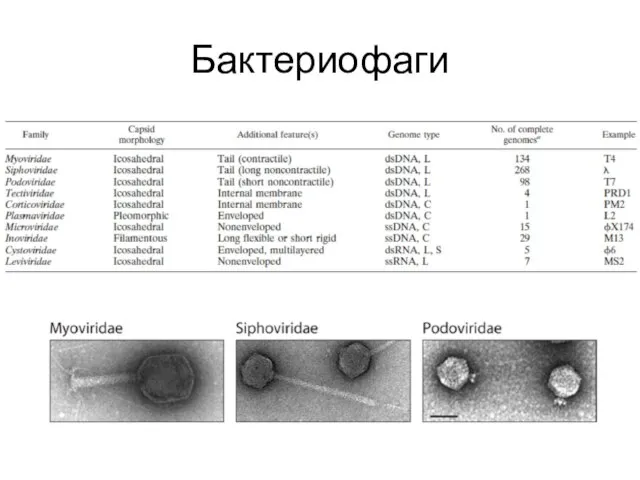
Бактериофаги
Бактериофаги

В геноме фагов есть две транскрипционные единицы. Одна из транскрипционных единиц
В геноме фагов есть две транскрипционные единицы. Одна из транскрипционных единиц

Такой принцип количественной транскрипционной регуляции называется каскадным (не путать с темпоральной
Такой принцип количественной транскрипционной регуляции называется каскадным (не путать с темпоральной
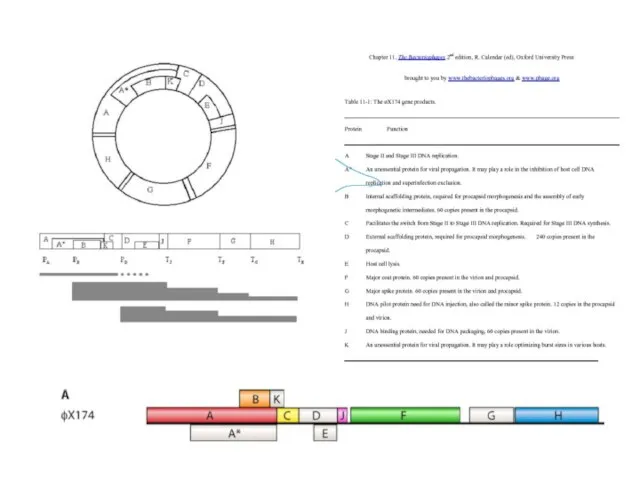

Итак, фаг φХ174 дает нам прекрасный пример позитивной каскадной количественной регуляции,
Итак, фаг φХ174 дает нам прекрасный пример позитивной каскадной количественной регуляции,
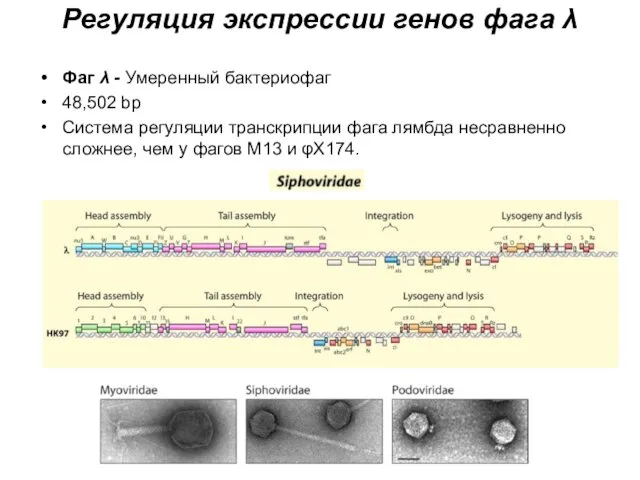
Регуляция экспрессии генов фага λ
Фаг λ - Умеренный бактериофаг
48,502 bp
Регуляция экспрессии генов фага λ
Фаг λ - Умеренный бактериофаг
48,502 bp
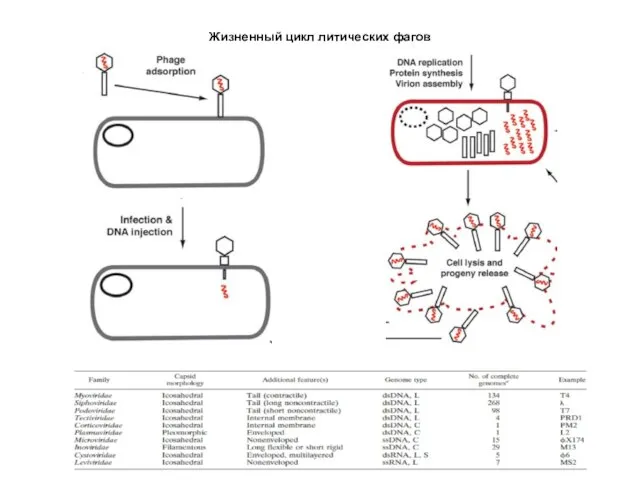
Жизненный цикл литических фагов
Жизненный цикл литических фагов
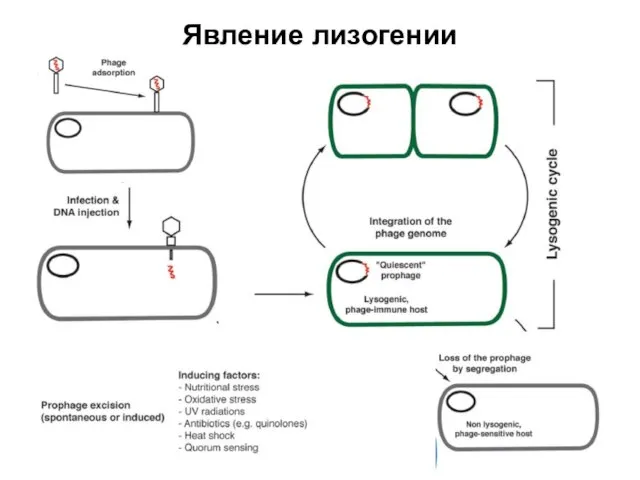
Явление лизогении
Явление лизогении
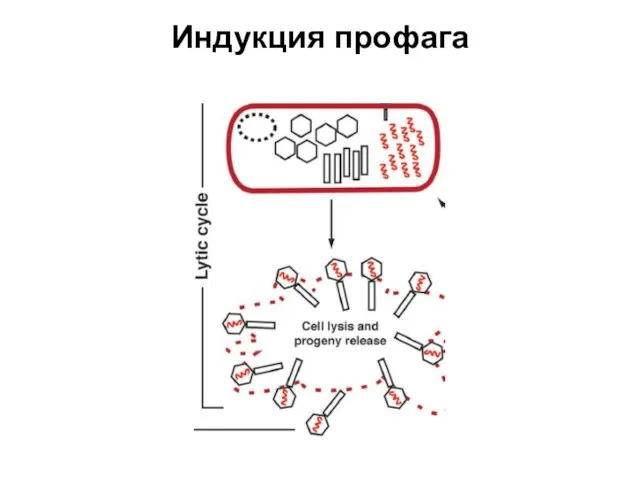
Индукция профага
Индукция профага
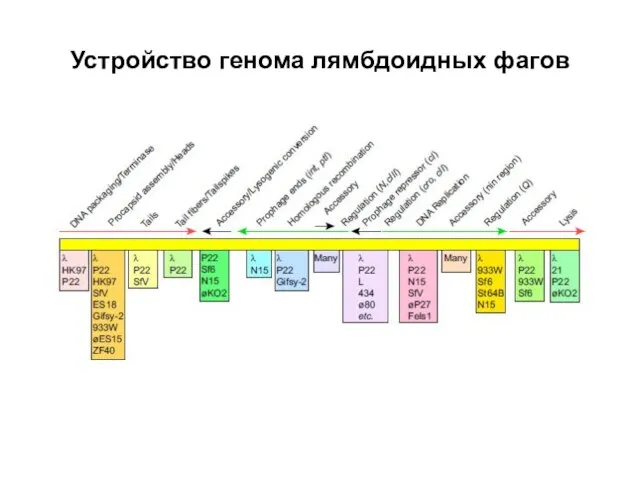
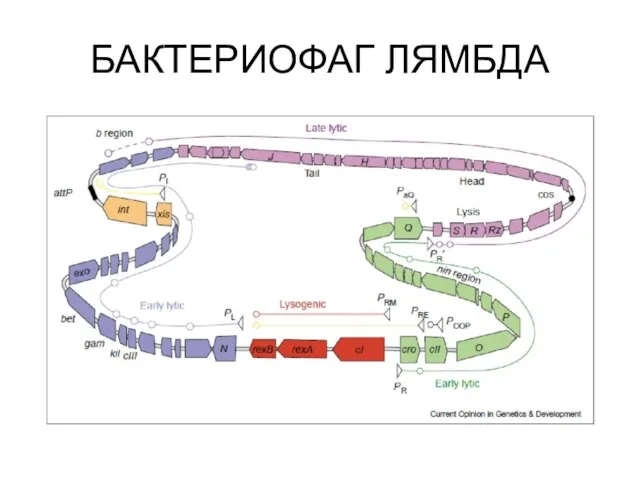
Устройство генома лямбдоидных фагов
Устройство генома лямбдоидных фагов
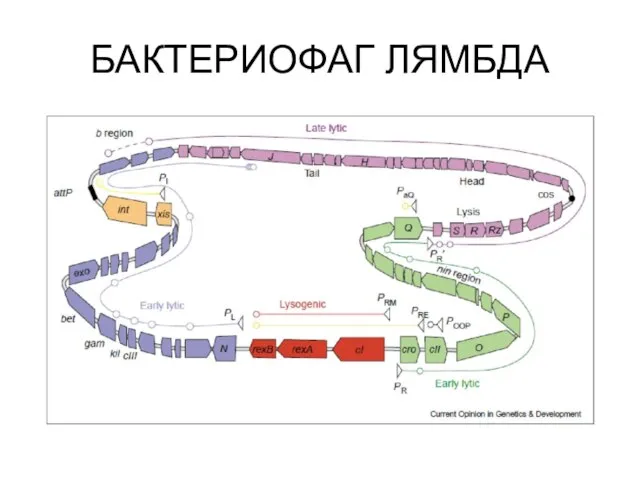
БАКТЕРИОФАГ ЛЯМБДА
БАКТЕРИОФАГ ЛЯМБДА

На ранней стадии происходит активация транскрипции с помощью клеточных белков.
Белок
На ранней стадии происходит активация транскрипции с помощью клеточных белков. Белок
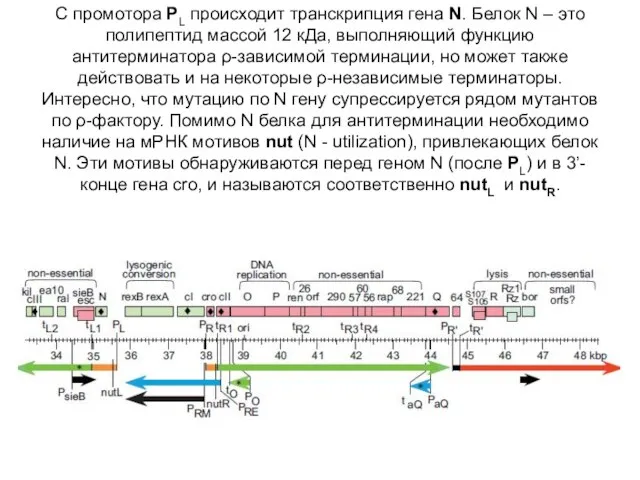
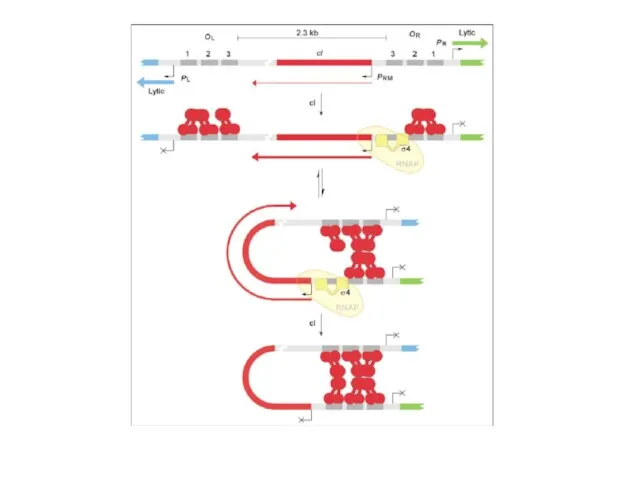
С промотора PL происходит транскрипция гена N. Белок N – это
С промотора PL происходит транскрипция гена N. Белок N – это
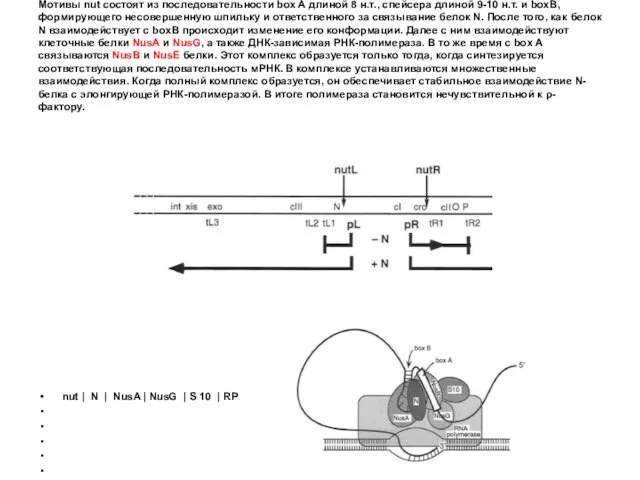
Мотивы nut состоят из последовательности box A длиной 8 н.т., спейсера
Мотивы nut состоят из последовательности box A длиной 8 н.т., спейсера

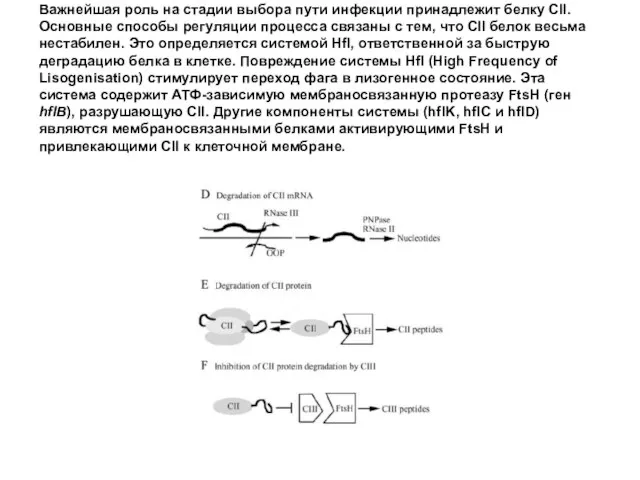
Важнейшая роль на стадии выбора пути инфекции принадлежит белку CII. Основные
Важнейшая роль на стадии выбора пути инфекции принадлежит белку CII. Основные
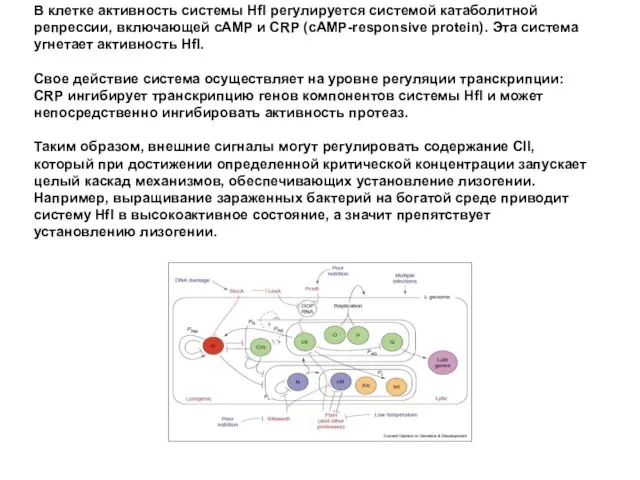
В клетке активность системы Hfl регулируется системой катаболитной репрессии, включающей сAMP
В клетке активность системы Hfl регулируется системой катаболитной репрессии, включающей сAMP
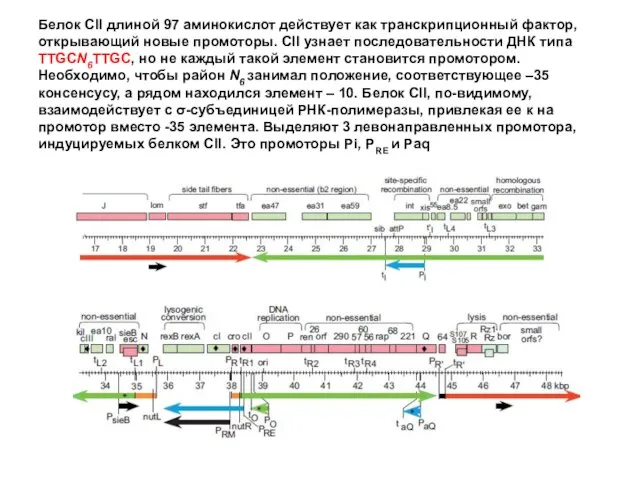
Белок CII длиной 97 аминокислот действует как транскрипционный фактор, открывающий новые
Белок CII длиной 97 аминокислот действует как транскрипционный фактор, открывающий новые
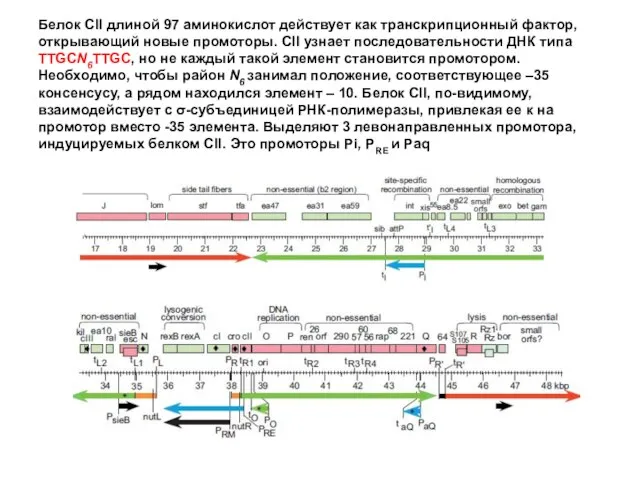
Белок CII длиной 97 аминокислот действует как транскрипционный фактор, открывающий новые
Белок CII длиной 97 аминокислот действует как транскрипционный фактор, открывающий новые
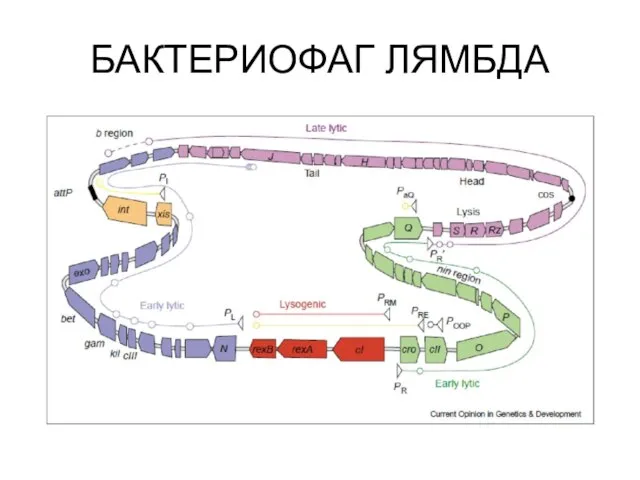
БАКТЕРИОФАГ ЛЯМБДА
БАКТЕРИОФАГ ЛЯМБДА

Нужны внешние воздействия для выхода фага из лизогенного состояния. Таким событием,
Нужны внешние воздействия для выхода фага из лизогенного состояния. Таким событием,
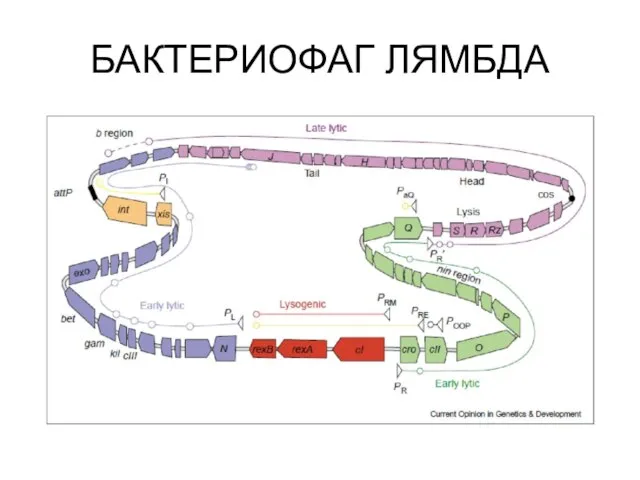
БАКТЕРИОФАГ ЛЯМБДА
БАКТЕРИОФАГ ЛЯМБДА
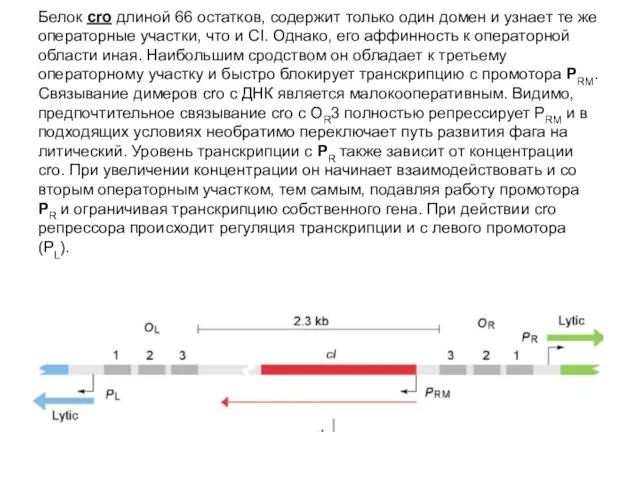
Белок cro длиной 66 остатков, содержит только один домен и узнает
Белок cro длиной 66 остатков, содержит только один домен и узнает
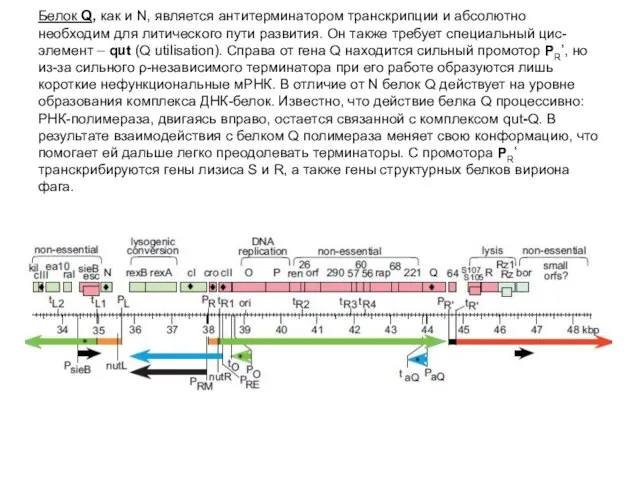
Белок Q, как и N, является антитерминатором транскрипции и абсолютно необходим
Белок Q, как и N, является антитерминатором транскрипции и абсолютно необходим
БАКТЕРИОФАГ ЛЯМБДА
БАКТЕРИОФАГ ЛЯМБДА
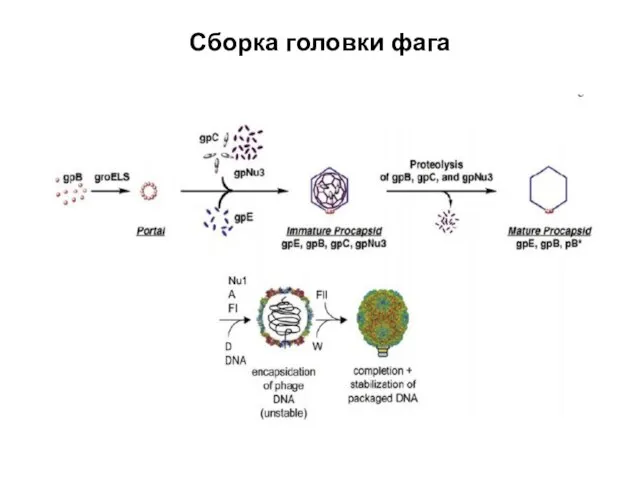
Сборка головки фага
Сборка головки фага
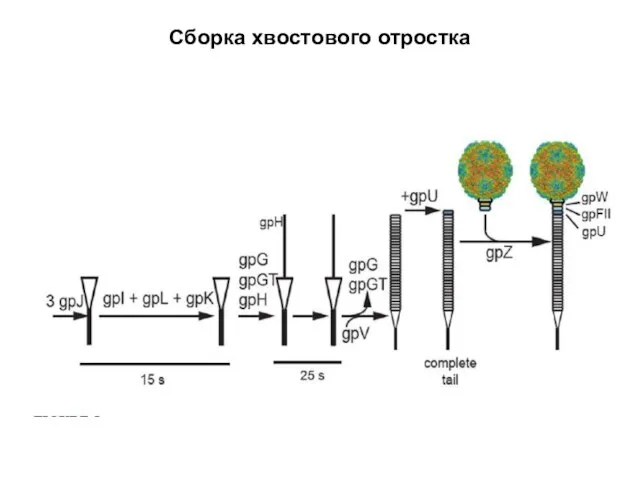
Сборка хвостового отростка
Сборка хвостового отростка
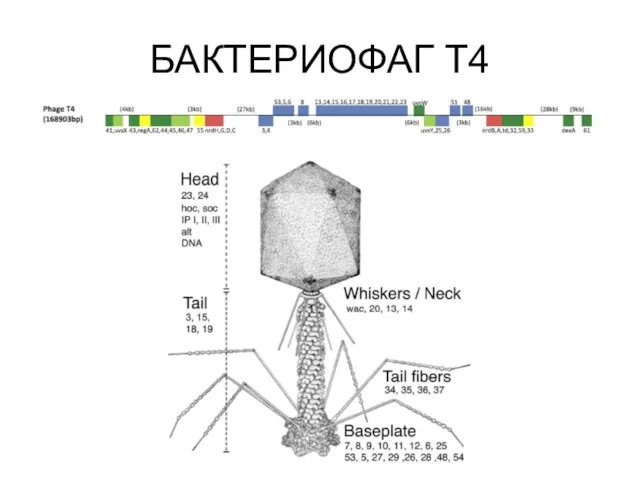
БАКТЕРИОФАГ Т4
БАКТЕРИОФАГ Т4

Гены Т4 делятся на истинно ранние, ранние, средние и поздние. Около
Гены Т4 делятся на истинно ранние, ранние, средние и поздние. Около
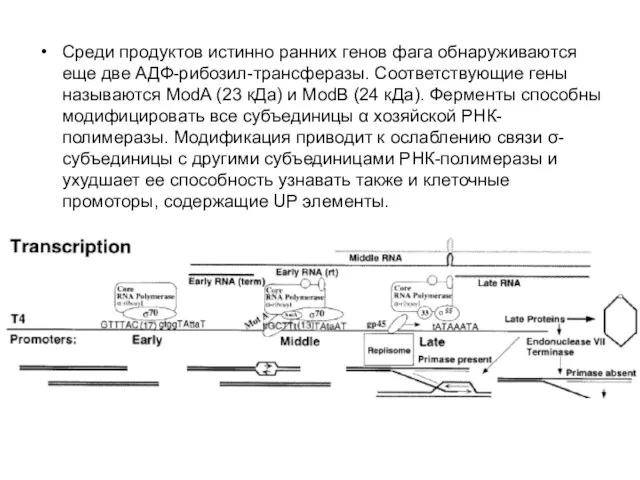
Среди продуктов истинно ранних генов фага обнаруживаются еще две АДФ-рибозил-трансферазы. Соответствующие
Среди продуктов истинно ранних генов фага обнаруживаются еще две АДФ-рибозил-трансферазы. Соответствующие

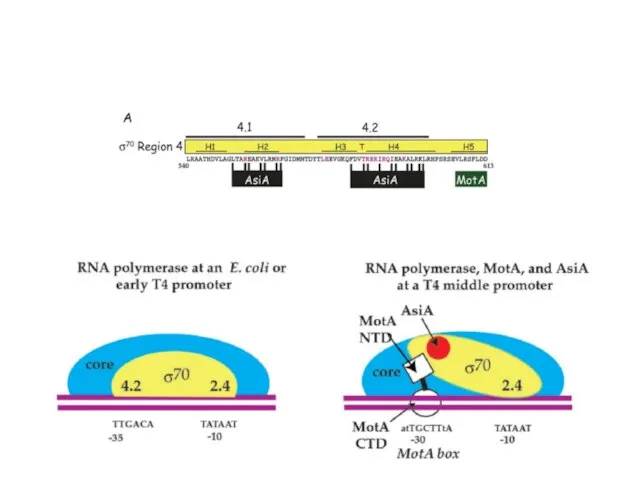
Для транскрипции средних генов нужны вирусные белки Asi A (antisigma) и
Для транскрипции средних генов нужны вирусные белки Asi A (antisigma) и
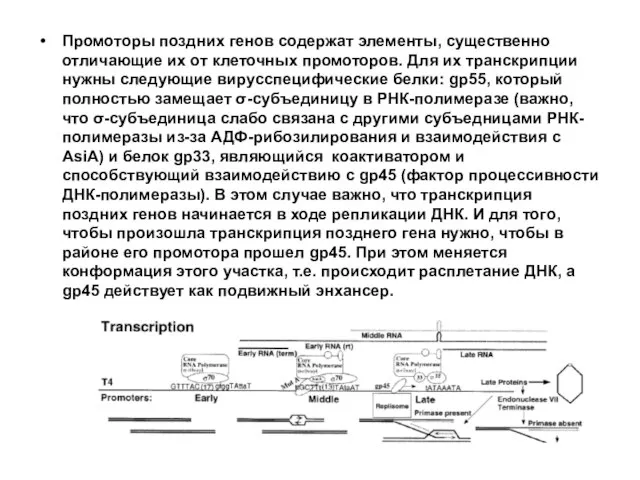
Промоторы поздних генов содержат элементы, существенно отличающие их от клеточных промоторов.
Промоторы поздних генов содержат элементы, существенно отличающие их от клеточных промоторов.

Известно, что gp55 является фактором, обеспечивающим весьма слабое сродство к ДНК
Известно, что gp55 является фактором, обеспечивающим весьма слабое сродство к ДНК
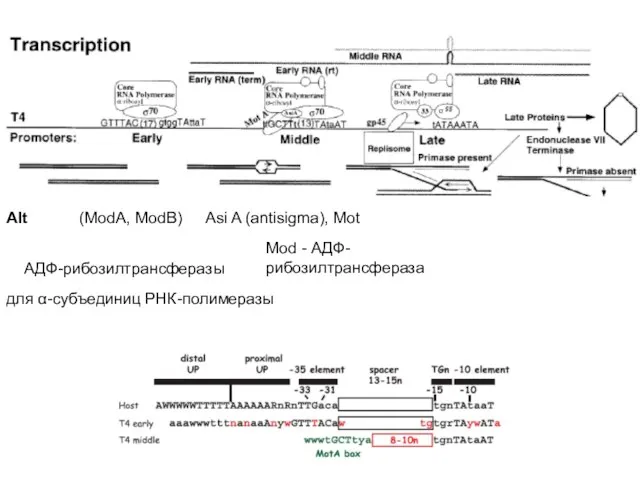
Alt
(ModA, ModB)
АДФ-рибозилтрансферазы
для α-субъединиц РНК-полимеразы
Asi A (antisigma), Mot
Mod
Alt
(ModA, ModB)
АДФ-рибозилтрансферазы
для α-субъединиц РНК-полимеразы
Asi A (antisigma), Mot
Mod
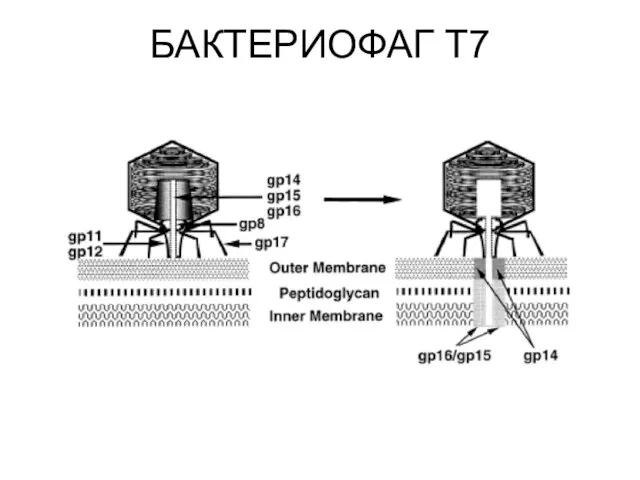
БАКТЕРИОФАГ Т7
БАКТЕРИОФАГ Т7

Все гены фага Т7 транскрибируются с одной и той же цепи
Все гены фага Т7 транскрибируются с одной и той же цепи

Первоначально в клетку бактерии попадает левый конец генома размером около 850
Первоначально в клетку бактерии попадает левый конец генома размером около 850

Ранние транскрипты у фага Т7 подвергаются процессингу РНКазой III. Точек разрезания
Ранние транскрипты у фага Т7 подвергаются процессингу РНКазой III. Точек разрезания
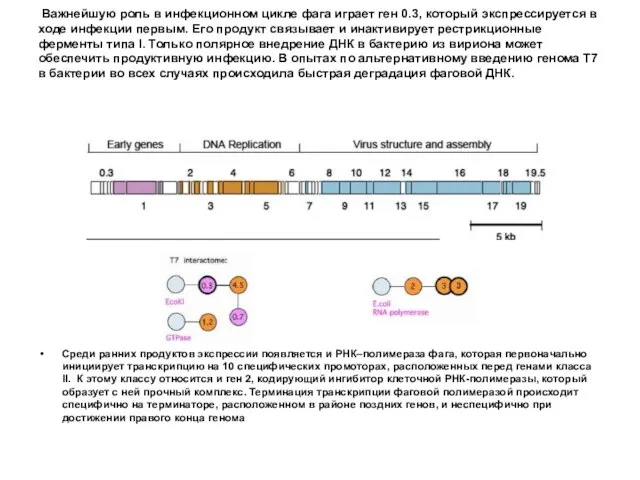
Важнейшую роль в инфекционном цикле фага играет ген 0.3, который
Важнейшую роль в инфекционном цикле фага играет ген 0.3, который
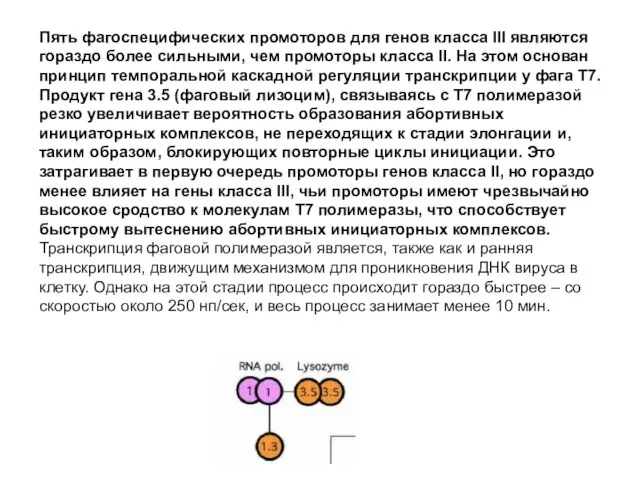
Пять фагоспецифических промоторов для генов класса III являются гораздо более сильными,
Пять фагоспецифических промоторов для генов класса III являются гораздо более сильными,

Регуляция транскрипции у фага N4
Геном этого фага представлен линейной двуцепочечной ДНК
Регуляция транскрипции у фага N4 Геном этого фага представлен линейной двуцепочечной ДНК


Средние гены транскрибируются N4 РНК-полимеразой II, являющейся продуктом ранних генов и
Средние гены транскрибируются N4 РНК-полимеразой II, являющейся продуктом ранних генов и


Поздние гены транскрибируются клеточной σ70-содержащей РНК-полимеразой. Однако, поздние промоторы N4 проявляют
Поздние гены транскрибируются клеточной σ70-содержащей РНК-полимеразой. Однако, поздние промоторы N4 проявляют


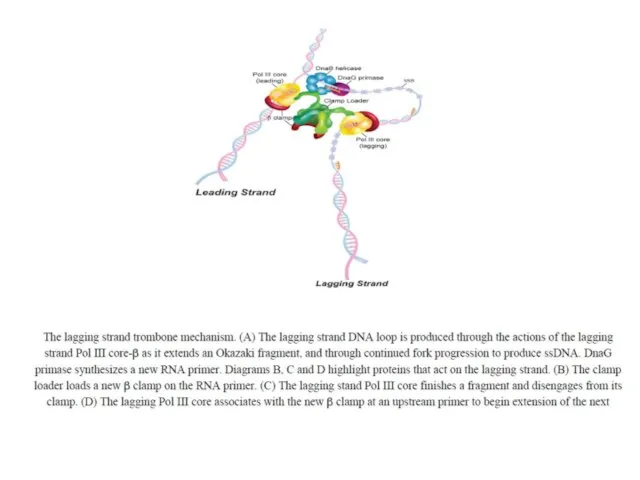
ДНК полимераза III E.coli
Фермент состоит из нескольких субъединиц и представляет собой
ДНК полимераза III E.coli
Фермент состоит из нескольких субъединиц и представляет собой

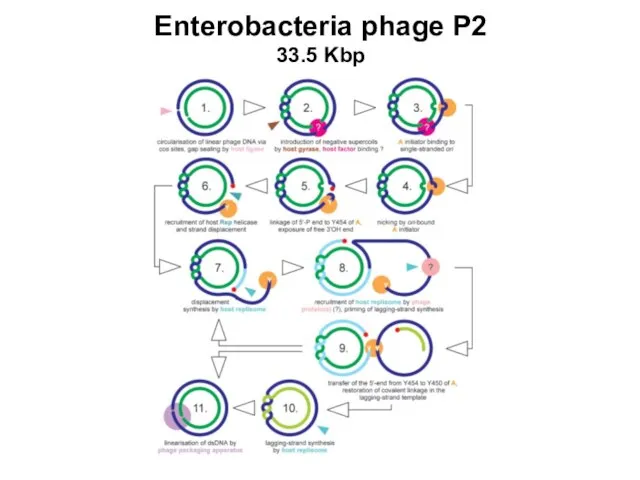
Enterobacteria phage P2
33.5 Kbp
Enterobacteria phage P2
33.5 Kbp
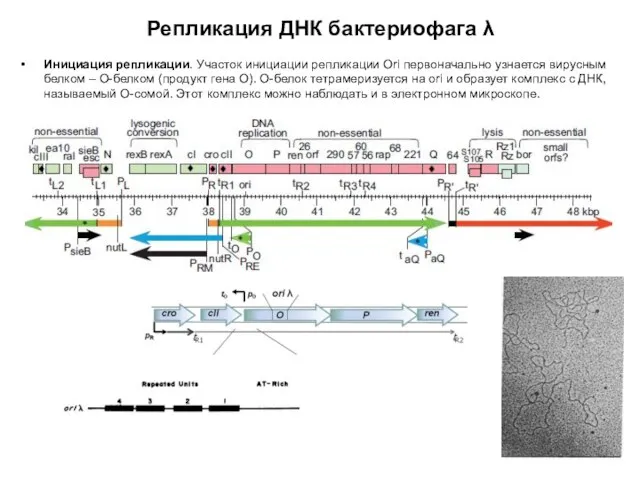
Репликация ДНК бактериофага λ
Инициация репликации. Участок инициации репликации Ori первоначально узнается
Репликация ДНК бактериофага λ
Инициация репликации. Участок инициации репликации Ori первоначально узнается

Если ассоциация белка О с комплексом DnaB-Р-белок не произойдет (в случае
Если ассоциация белка О с комплексом DnaB-Р-белок не произойдет (в случае


В ходе двунаправленной репликации по схеме Кернса в клетках, зараженных фагом
В ходе двунаправленной репликации по схеме Кернса в клетках, зараженных фагом
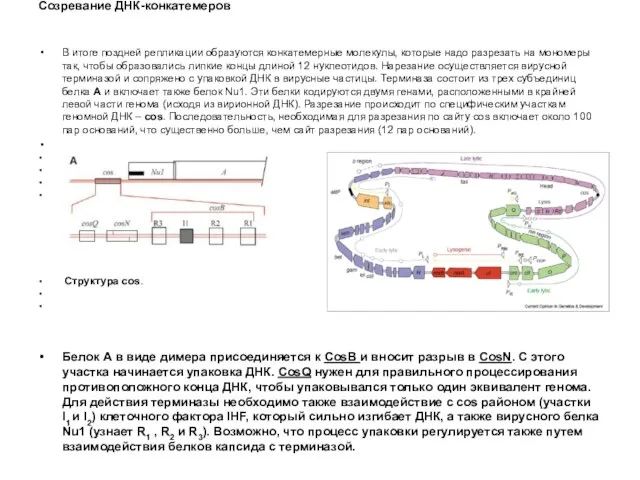
Созревание ДНК-конкатемеров
В итоге поздней репликации образуются конкатемерные молекулы, которые надо разрезать
Созревание ДНК-конкатемеров
В итоге поздней репликации образуются конкатемерные молекулы, которые надо разрезать
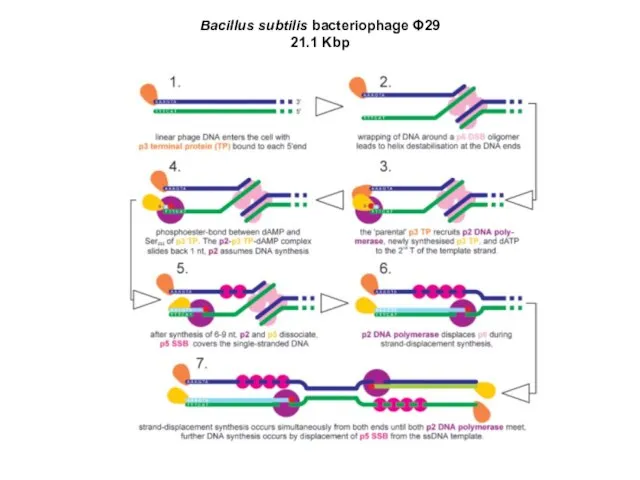
Bacillus subtilis bacteriophage Φ29
21.1 Kbp
Bacillus subtilis bacteriophage Φ29
21.1 Kbp

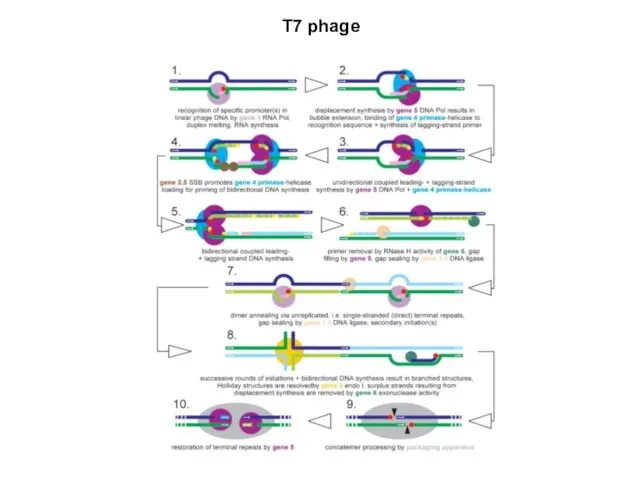
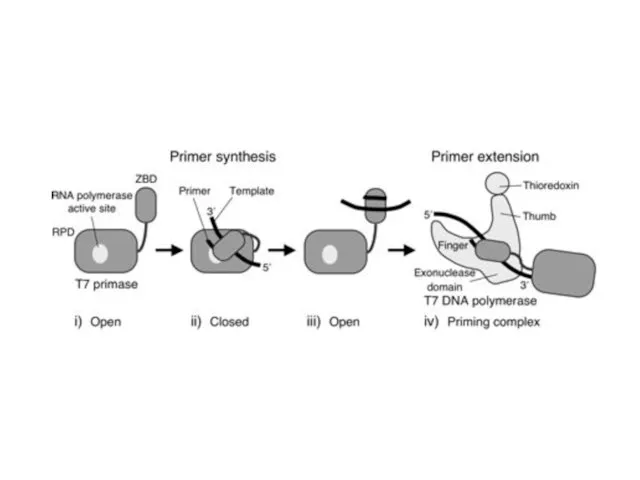
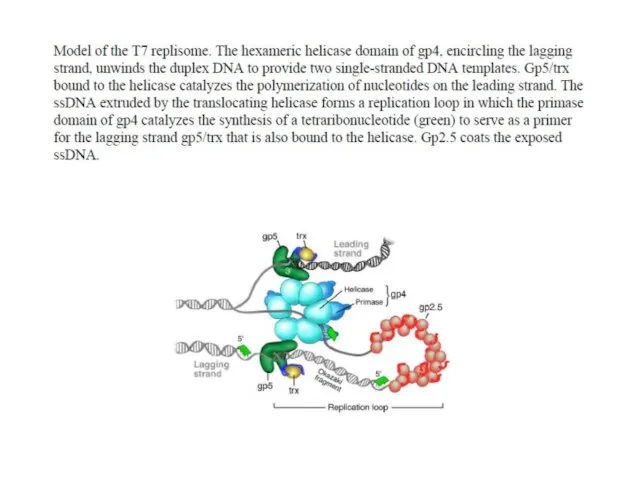
T7 phage
T7 phage

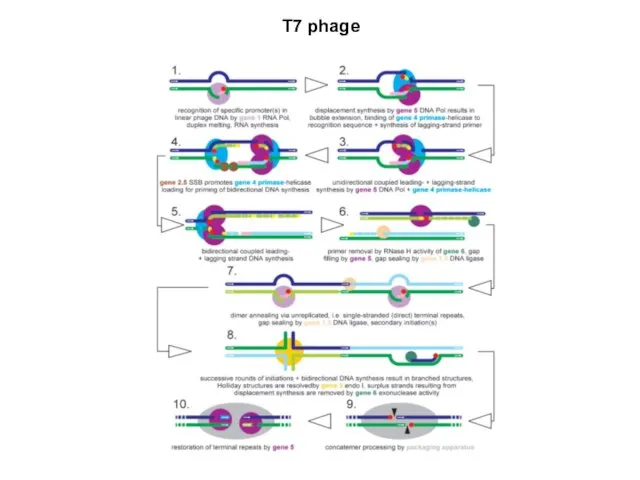
T7 phage
T7 phage




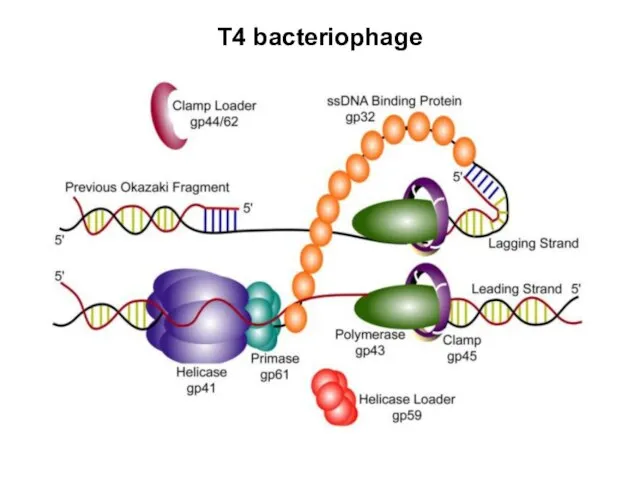
T4 bacteriophage
T4 bacteriophage
 Генетические заболевания
Генетические заболевания Фотосинтез 6 класс
Фотосинтез 6 класс Методы нанодиагностики. Биосенсоры, биочипы
Методы нанодиагностики. Биосенсоры, биочипы Размножение и индивидуальное развитие организмов. Онтогенез
Размножение и индивидуальное развитие организмов. Онтогенез Садовые цветы
Садовые цветы Презентация на тему Симбиоз
Презентация на тему Симбиоз  Подбор растений в уголках природы в разных возрастных группах ДОУ
Подбор растений в уголках природы в разных возрастных группах ДОУ Витамины: за или против
Витамины: за или против Ткани. Типы тканей и их свойства.
Ткани. Типы тканей и их свойства. Тест по теме Биосфера
Тест по теме Биосфера Растения потогонного действия
Растения потогонного действия Развитие эволюционных идей. Доказательства эволюции.
Развитие эволюционных идей. Доказательства эволюции. Атомная энергетика – за и против.
Атомная энергетика – за и против. Тип членистоногие (Arthropoda) Методическая разработка урока Учитель биологии ЦО № 627 ЦАО г. Москвы Сухова Н.И.
Тип членистоногие (Arthropoda) Методическая разработка урока Учитель биологии ЦО № 627 ЦАО г. Москвы Сухова Н.И. Тип Кишечнополостные
Тип Кишечнополостные Геном человека
Геном человека Презентация на тему "Как животные чистят зубы?" - скачать презентации по Биологии
Презентация на тему "Как животные чистят зубы?" - скачать презентации по Биологии Дружда между лошадьми
Дружда между лошадьми Проект урока в 6 классе (УМК Н.И.Сонина) Организм как целостная система Автор: Николаева Славяна Борисовна учит
Проект урока в 6 классе (УМК Н.И.Сонина) Организм как целостная система Автор: Николаева Славяна Борисовна учит Презентация на тему "Условия жизни растений" - скачать презентации по Биологии
Презентация на тему "Условия жизни растений" - скачать презентации по Биологии Викторина по природоведению
Викторина по природоведению Леса России. Деревья и кустарники
Леса России. Деревья и кустарники Живое вещество – источник нафтидов. Геологическая эволюция ЖВ
Живое вещество – источник нафтидов. Геологическая эволюция ЖВ Ayurveda cosmetics. Отражение внутренней гармонии
Ayurveda cosmetics. Отражение внутренней гармонии Ахатина – идеальный домашний питомец для современного человека
Ахатина – идеальный домашний питомец для современного человека Наука о растениях – ботаника. Мир растений
Наука о растениях – ботаника. Мир растений (пособие для учащихся по окружающему миру )
(пособие для учащихся по окружающему миру ) Гиганты и карлики растительного мира
Гиганты и карлики растительного мира