- Рецепторы гормонов и передача сигналов: рецепторы клеточной поверхности, внутриклеточные рецепторы

Содержание
- 2. Рецепторы с ферментативной активностью: Взаимодействие внешнего домена рецептора с гормоном усиливает ферментативную активность внутриклеточного каталитического домена,
- 3. Передача сигнала через рецепторы c активностью тирозинкиназы (RTK) Инсулиновые рецепторы (IRs) и их действие IR представляет
- 4. Инсулиновые рецепторы (IRs) и их действие IRS-1 может передавать сигнал инсулина по двум путям: Ras-независимый путь,
- 5. Инсулиновые рецепторы, эффекты инсулина Ras-independent pathway (PI3K) Фосфорилированный IRS-1 взаимодействует с фосфоинозитид-3-киназой (PI3-kinase, PI3K) через SH2-domain:
- 6. Инсулиновые рецепторы, эффекты инсулина Ras-independent pathway(PI3K) Инсулин а также инсулиноподобные факторы роста, IGF-1 и IGF-2) модулируют
- 8. Инсулиновые рецепторы, эффекты инсулина Ras-independent pathwa (PI3K) Активация PKB/Akt приводит к фосфорилированию и активации sterol-regulated element-binding
- 9. Инсулиновые рецепторы, эффекты инсулина Ras-independent pathwa (PI3K) На уровне липидного гомеостаза инсулин ингибирует липолиз и активирует
- 10. Insulin receptors, IR and insulin action Ras-зависимый путь (Ras/MAPK pathway): Фосфорилированный IRS-1 распознается и связывается с
- 11. Insulin receptors, IR and insulin action Ras-зависимый путь (Ras/MAPK pathway): Ras-белки – мономерные G-белков (более 100
- 12. Insulin receptors, IR and insulin action Ras-зависимый путь (Ras/MAPK pathway): Чрезмерная передача сигналов от мутированных или
- 13. При присоединении гормона 1) α-субъединица изменяет свою конформацию, 2) это приводит к появлению тирозинкиназной активности у
- 14. Инсулин активирует использование клетками глюкозы путем: 1. Активирования ключевых ферментов гликолиза (глюкокиназы, фосфофруктокиназы, пируваткиназы); 2. Увеличения
- 15. Эффекты инсулина
- 16. Эффекты инсулина
- 18. Сахарный диабет развивается при нарушении функции инсулина Выделяют два типа сахарного диабета: Тип I – инсулинзависимый
- 28. Внутриклеточные рецепторы Гормон / сигнал проникает через мембрану и активирует транскрипцию генов. рецепторы стероидных гормонов (рецепторы
- 29. Внутриклеточные рецепторы Связывание гормона с рецептором изменяет конформацию и приобретает способность связываться с конкретными последовательностями ДНК
- 30. Внутриклеточные рецепторы являются регуляторами транскрипции генов. Все рецепторы имеют схожие структуры:: ligand binding domain ( for
- 31. Кортикостероиды
- 34. Глюкокортикоиды Надпочечники человека скретируют глюкокортикоиды: кортизол (гидрокортизон), кортизон и кортикостерон. Ткани-мишени: печень, почки, лимфоидная, соединительная и
- 35. Влияние на углеводный обмен: 1. в печени стимулируют синтез гликогена и глюконеогенез (синтез глюкозы из аминокислот);
- 36. Глюкокортикоиды кортизон
- 37. Внутриклеточные рецепторы являются регуляторами транскрипции генов. Все рецепторы имеют схожие структуры:: ligand binding domain ( for
- 38. Стероидные рецепторы - Глюкокортикоидный рецептор (GR) обнаружен в цитоплазме связывается с шаперонами Hsp-90 (ингибитор) в отсутствие
- 39. Цинковый палец (англ. zinc finger) — тип белковой структуры, небольшой белковый мотив, стабилизированный одним или двумя
- 40. Цинковые пальцы и ДНК
- 41. Внутриклеточные рецепторы
- 42. Органы-мишени: Печень АНАБОЛИЧЕСКОЕ ДЕЙСТВИЕ Мышцы Кожа КАТАБОЛИЧЕСКОЕ Жировая ткань ДЕЙСТВИЕ Лимфатическая ткань, Соединительная ткань Данные гормоны
- 43. ГК в обмене углеводов
- 44. Результативные эффекты 1. Уровень глюкозы повышен 2. Синтез белков снижен 3. Снижение биосинтеза γ-глобулинов приводит к
- 45. ГК в обмене липидов
- 46. Результативные эффекты 1. Биосинтез ТАГ в жировых депо снижен 2. Концентрация жирных кислот в крови увеличивается
- 47. При высокой концентрации глюкокортикоиды оказывают следующие эффекты: 1.В лимфоидной ткани подавляют иммунные реакции, вызывая гибель лимфоцитов
- 48. Поражение гипофиза Опухоль надпочечников или Длительный прием ГК Гиперкортицизм
- 49. Гиперкортицизм Встречается гиперкортицизм, сопровождающейся избыточной секрецией преимущественно минералкортикоидов (альдостерона) –гиперальдостеронизм, или болезнь Конна. При этом заболевании
- 50. Гипокортицизм
- 52. Тиреоидные гормоны
- 56. Этапы биосинтеза тиреоидных гормонов I. Поступение «йода» (иодид-ионы) из крови в ЩЖ II. Окисление иодид-ионов под
- 62. Траспорт тиреоидных гормонов: ТСГ- тиреоид связывающий глобулин ТСПА - тиреосвязывающий преальбумин
- 65. Рецепторы гормонов щитовидной железы - TR (и RAR также) находятся в ядре и связываются с ДНК
- 66. Метаболические эффекты Т3 В ядре ↑транскрипцию и трансляцию белков – ферментов ОВР В костной ткани активирует
- 67. Гормон Т2 Рецепторы для этого гормона находятся в митохондриях. По силе своего биологического действия этот гормон
- 70. Эндемический зоб - ИДС
- 72. Пути коррекции йоддефицитных состояний Областная программа «Профилактика эндемического зоба и других йоддефицитных заболеваний на 1999 -
- 74. Скачать презентацию
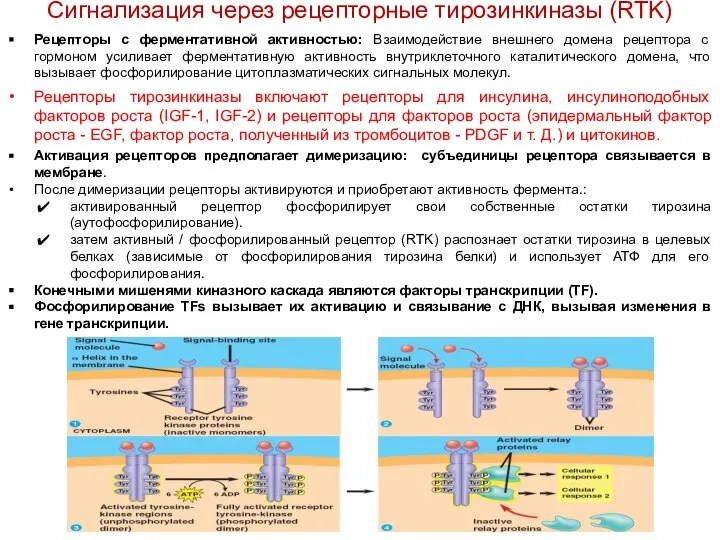
Рецепторы с ферментативной активностью: Взаимодействие внешнего домена рецептора с гормоном усиливает

Передача сигнала через рецепторы c активностью тирозинкиназы (RTK)
Инсулиновые рецепторы (IRs) и
Передача сигнала через рецепторы c активностью тирозинкиназы (RTK)
Инсулиновые рецепторы (IRs) и

Инсулиновые рецепторы (IRs) и их действие
IRS-1 может передавать сигнал инсулина по
IRS-1 может передавать сигнал инсулина по
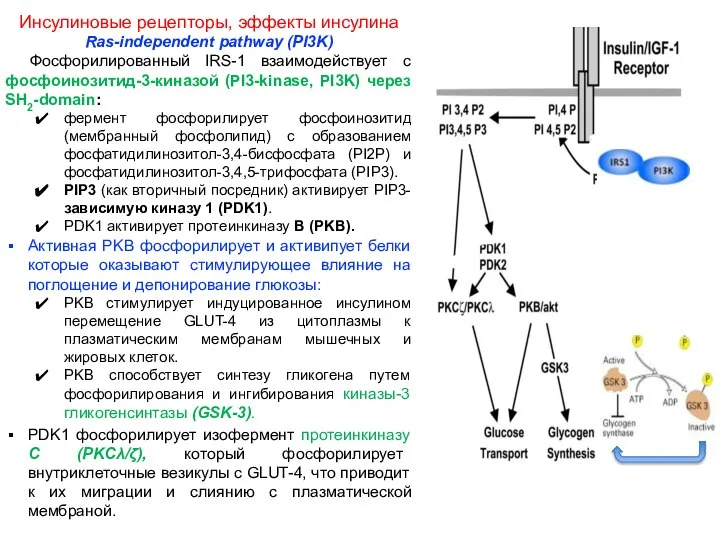
Инсулиновые рецепторы, эффекты инсулина
Ras-independent pathway (PI3K)
Фосфорилированный IRS-1 взаимодействует с фосфоинозитид-3-киназой
Инсулиновые рецепторы, эффекты инсулина
Ras-independent pathway (PI3K)
Фосфорилированный IRS-1 взаимодействует с фосфоинозитид-3-киназой
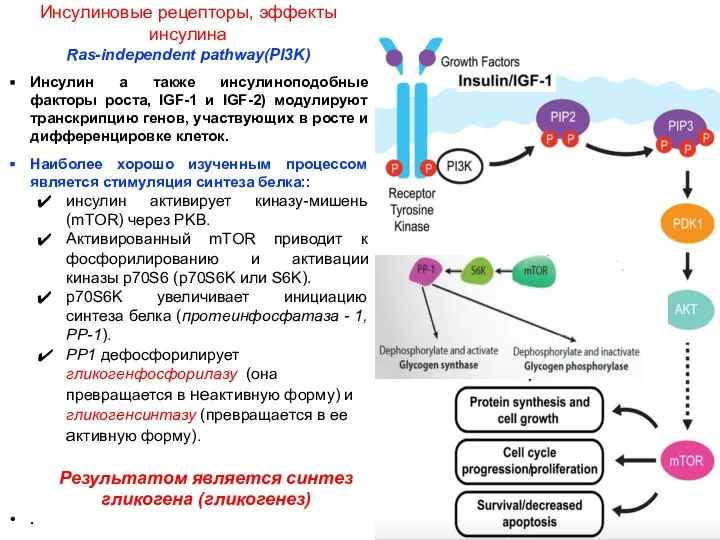
Инсулиновые рецепторы, эффекты инсулина
Ras-independent pathway(PI3K)
Инсулин а также инсулиноподобные факторы роста, IGF-1
Инсулиновые рецепторы, эффекты инсулина
Ras-independent pathway(PI3K)
Инсулин а также инсулиноподобные факторы роста, IGF-1

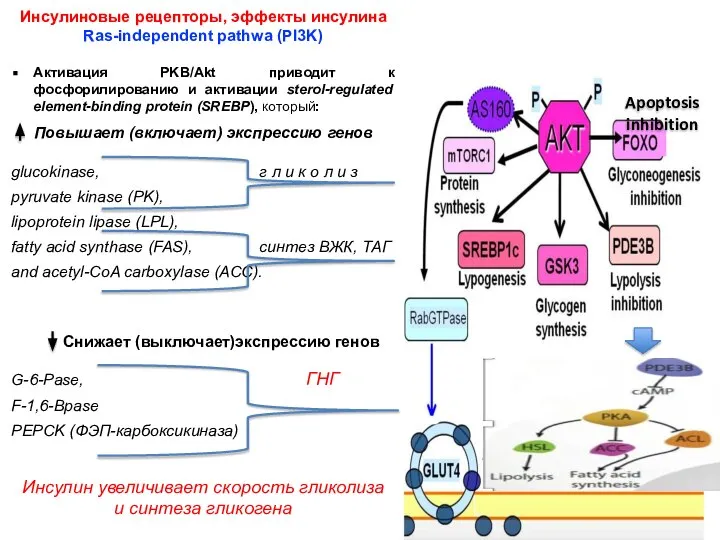
Инсулиновые рецепторы, эффекты инсулина
Ras-independent pathwa (PI3K)
Активация PKB/Akt приводит к фосфорилированию и
Инсулиновые рецепторы, эффекты инсулина
Ras-independent pathwa (PI3K)
Активация PKB/Akt приводит к фосфорилированию и

Инсулиновые рецепторы, эффекты инсулина
Ras-independent pathwa (PI3K)
На уровне липидного гомеостаза инсулин ингибирует
Инсулиновые рецепторы, эффекты инсулина
Ras-independent pathwa (PI3K)
На уровне липидного гомеостаза инсулин ингибирует

Insulin receptors, IR and insulin action
Ras-зависимый путь (Ras/MAPK pathway):
Фосфорилированный IRS-1 распознается
Insulin receptors, IR and insulin action
Ras-зависимый путь (Ras/MAPK pathway):
Фосфорилированный IRS-1 распознается

Insulin receptors, IR and insulin action
Ras-зависимый путь (Ras/MAPK pathway):
Ras-белки – мономерные
Insulin receptors, IR and insulin action
Ras-зависимый путь (Ras/MAPK pathway):
Ras-белки – мономерные

Insulin receptors, IR and insulin action
Ras-зависимый путь (Ras/MAPK pathway):
Чрезмерная передача сигналов
Insulin receptors, IR and insulin action
Ras-зависимый путь (Ras/MAPK pathway):
Чрезмерная передача сигналов
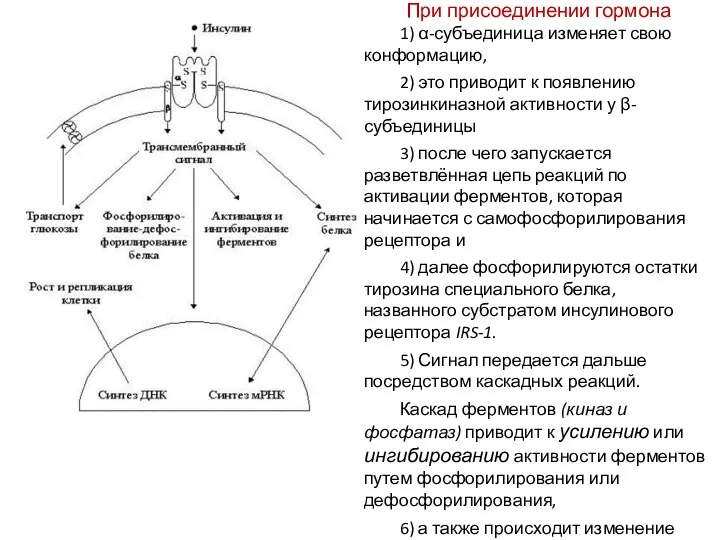
При присоединении гормона
1) α-субъединица изменяет свою конформацию,
2) это приводит
При присоединении гормона
1) α-субъединица изменяет свою конформацию,
2) это приводит

Инсулин активирует использование клетками глюкозы путем:
1. Активирования ключевых ферментов гликолиза (глюкокиназы,
фосфофруктокиназы,
Инсулин активирует использование клетками глюкозы путем:
1. Активирования ключевых ферментов гликолиза (глюкокиназы,
фосфофруктокиназы,

Эффекты инсулина
Эффекты инсулина

Эффекты инсулина
Эффекты инсулина


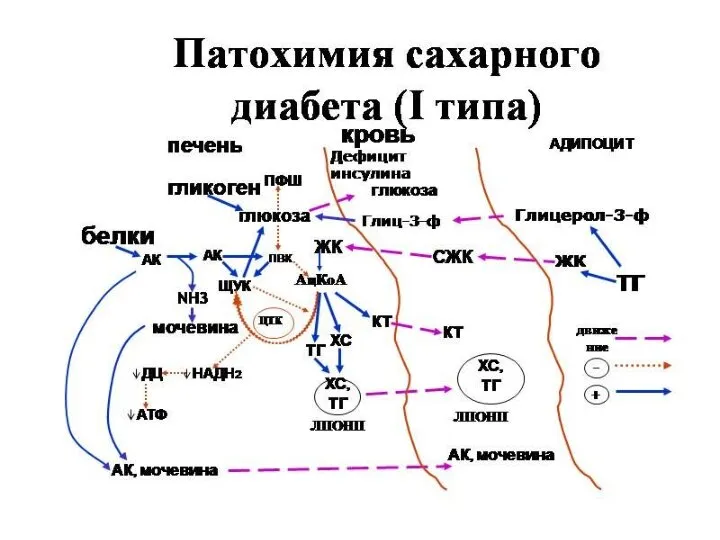
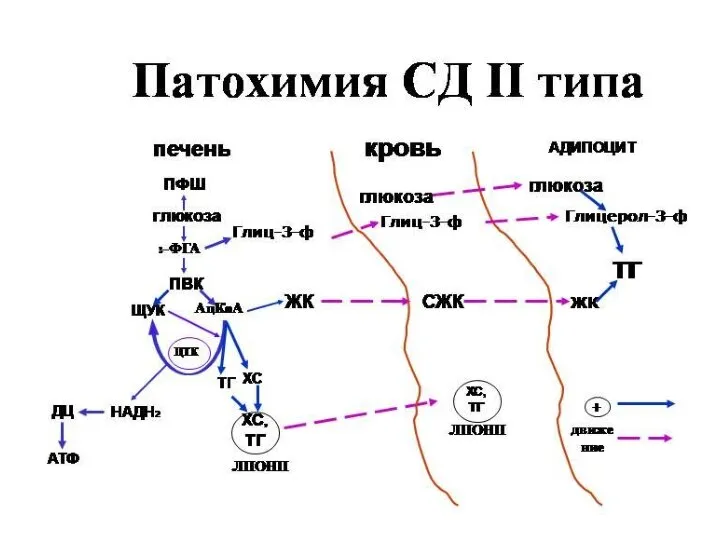
Сахарный диабет развивается при нарушении функции инсулина
Выделяют два типа сахарного диабета:
Тип
Сахарный диабет развивается при нарушении функции инсулина
Выделяют два типа сахарного диабета:
Тип








Внутриклеточные рецепторы
Гормон / сигнал проникает через мембрану и активирует транскрипцию генов.
рецепторы
Внутриклеточные рецепторы
Гормон / сигнал проникает через мембрану и активирует транскрипцию генов.
рецепторы

Внутриклеточные рецепторы
Связывание гормона с рецептором изменяет конформацию и приобретает способность связываться
Внутриклеточные рецепторы
Связывание гормона с рецептором изменяет конформацию и приобретает способность связываться

Внутриклеточные рецепторы являются регуляторами транскрипции генов.
Все рецепторы имеют схожие структуры::
Внутриклеточные рецепторы являются регуляторами транскрипции генов.
Все рецепторы имеют схожие структуры::

Кортикостероиды
Кортикостероиды



Глюкокортикоиды
Надпочечники человека скретируют глюкокортикоиды: кортизол (гидрокортизон), кортизон и кортикостерон.
Ткани-мишени: печень, почки,
Глюкокортикоиды
Надпочечники человека скретируют глюкокортикоиды: кортизол (гидрокортизон), кортизон и кортикостерон.
Ткани-мишени: печень, почки,

Влияние на углеводный обмен:
1. в печени стимулируют синтез гликогена и глюконеогенез
Влияние на углеводный обмен:
1. в печени стимулируют синтез гликогена и глюконеогенез
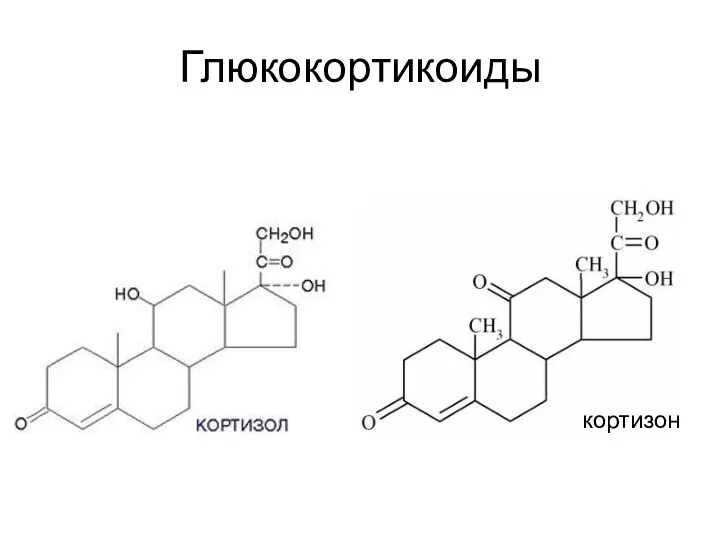
Глюкокортикоиды
кортизон
Глюкокортикоиды
кортизон
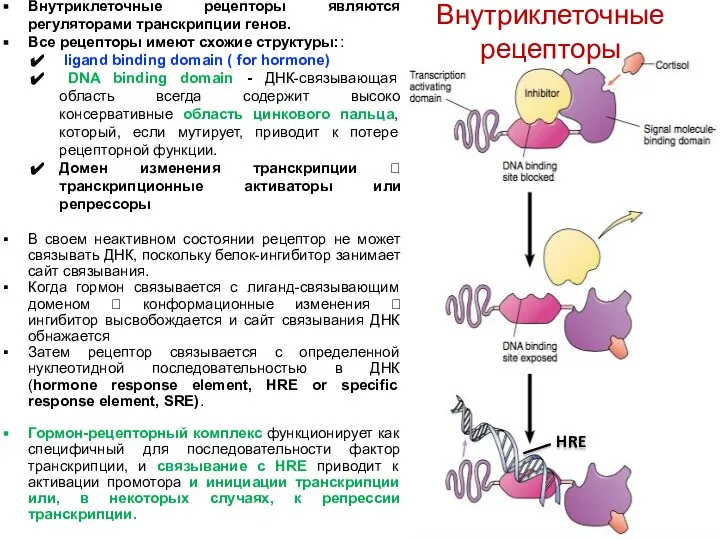
Внутриклеточные рецепторы являются регуляторами транскрипции генов.
Все рецепторы имеют схожие структуры::
Внутриклеточные рецепторы являются регуляторами транскрипции генов.
Все рецепторы имеют схожие структуры::

Стероидные рецепторы - Глюкокортикоидный рецептор (GR) обнаружен в цитоплазме
связывается с шаперонами
Стероидные рецепторы - Глюкокортикоидный рецептор (GR) обнаружен в цитоплазме
связывается с шаперонами
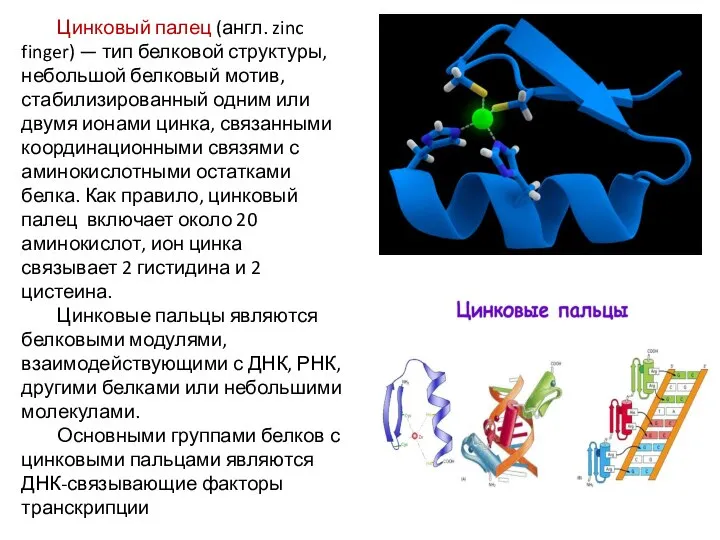
Цинковый палец (англ. zinc finger) — тип белковой структуры, небольшой белковый
Цинковый палец (англ. zinc finger) — тип белковой структуры, небольшой белковый
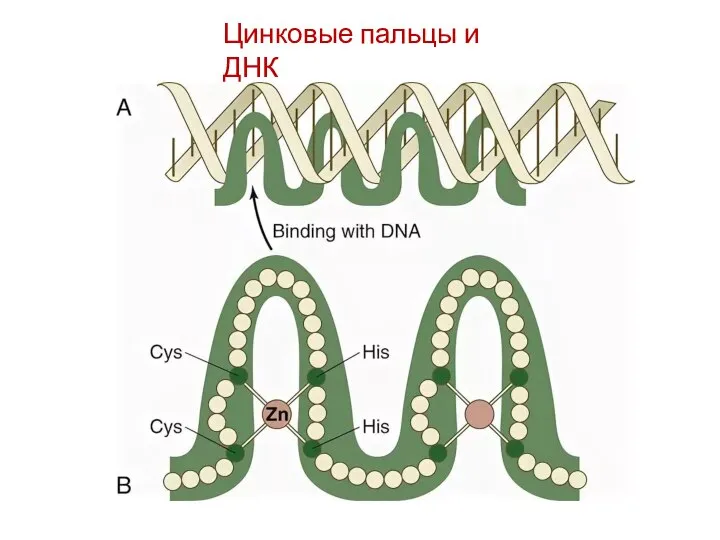
Цинковые пальцы и ДНК
Цинковые пальцы и ДНК

Внутриклеточные рецепторы
Внутриклеточные рецепторы
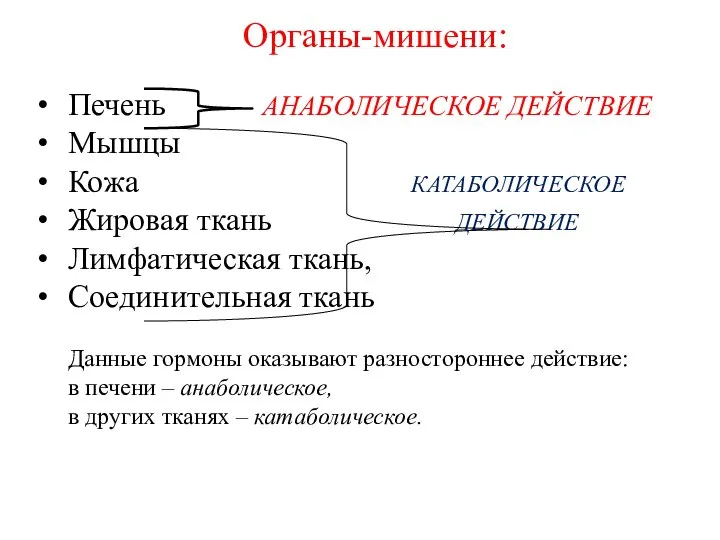
Органы-мишени:
Печень АНАБОЛИЧЕСКОЕ ДЕЙСТВИЕ
Мышцы
Кожа КАТАБОЛИЧЕСКОЕ
Жировая ткань ДЕЙСТВИЕ
Лимфатическая ткань,
Соединительная ткань
Данные
Органы-мишени:
Печень АНАБОЛИЧЕСКОЕ ДЕЙСТВИЕ
Мышцы
Кожа КАТАБОЛИЧЕСКОЕ
Жировая ткань ДЕЙСТВИЕ
Лимфатическая ткань,
Соединительная ткань
Данные

ГК в обмене углеводов
ГК в обмене углеводов

Результативные эффекты
1. Уровень глюкозы повышен
2. Синтез белков снижен
3. Снижение биосинтеза γ-глобулинов
Результативные эффекты
1. Уровень глюкозы повышен
2. Синтез белков снижен
3. Снижение биосинтеза γ-глобулинов

ГК в обмене липидов
ГК в обмене липидов

Результативные эффекты
1. Биосинтез ТАГ в жировых депо снижен
2. Концентрация жирных кислот
Результативные эффекты
1. Биосинтез ТАГ в жировых депо снижен
2. Концентрация жирных кислот

При высокой концентрации глюкокортикоиды оказывают следующие эффекты:
1.В лимфоидной ткани подавляют иммунные
При высокой концентрации глюкокортикоиды оказывают следующие эффекты:
1.В лимфоидной ткани подавляют иммунные

Поражение
гипофиза
Опухоль
надпочечников или
Длительный прием
ГК
Гиперкортицизм
Поражение
гипофиза
Опухоль
надпочечников или
Длительный прием
ГК
Гиперкортицизм

Гиперкортицизм
Встречается гиперкортицизм, сопровождающейся избыточной секрецией преимущественно минералкортикоидов (альдостерона) –гиперальдостеронизм, или болезнь
Гиперкортицизм
Встречается гиперкортицизм, сопровождающейся избыточной секрецией преимущественно минералкортикоидов (альдостерона) –гиперальдостеронизм, или болезнь

Гипокортицизм
Гипокортицизм


Тиреоидные гормоны
Тиреоидные гормоны




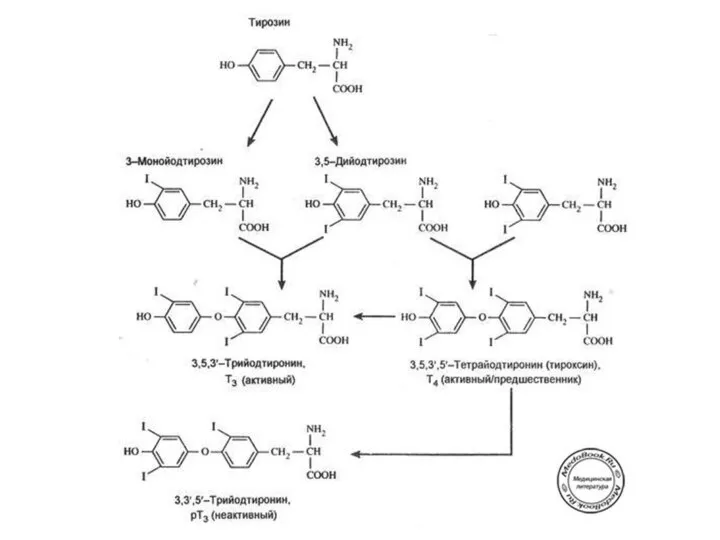
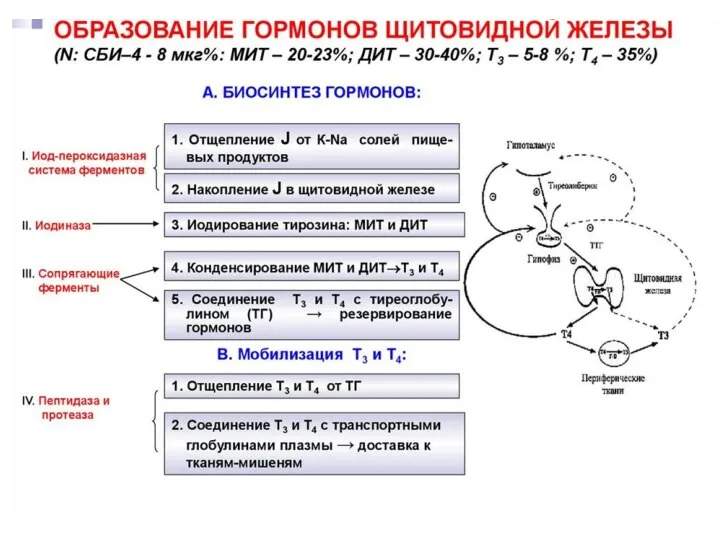
Этапы биосинтеза тиреоидных гормонов
I. Поступение «йода» (иодид-ионы) из крови в ЩЖ
II.
Этапы биосинтеза тиреоидных гормонов
I. Поступение «йода» (иодид-ионы) из крови в ЩЖ
II.
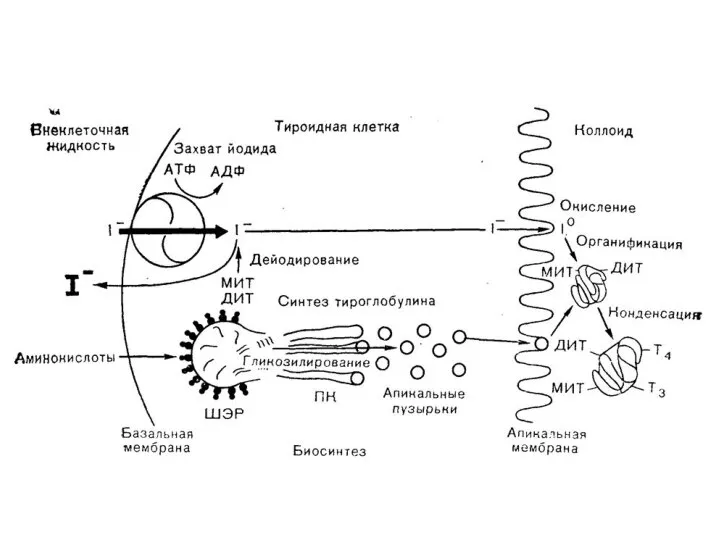



Траспорт тиреоидных гормонов:
ТСГ- тиреоид связывающий глобулин
ТСПА - тиреосвязывающий преальбумин
Траспорт тиреоидных гормонов:
ТСГ- тиреоид связывающий глобулин
ТСПА - тиреосвязывающий преальбумин


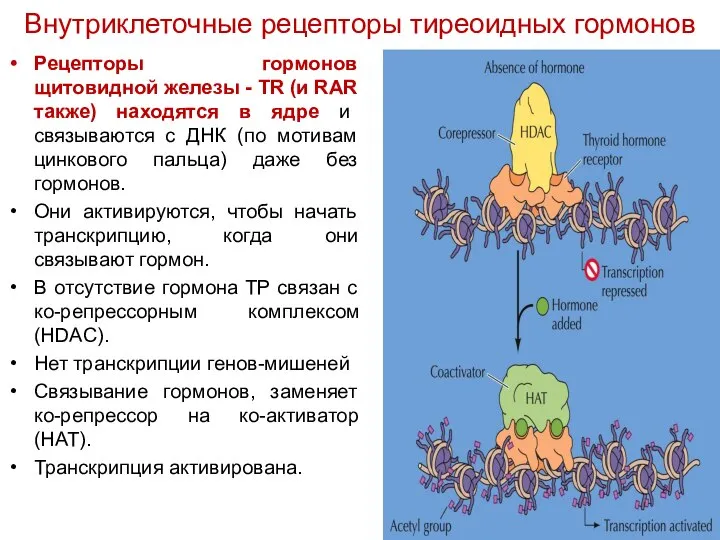
Рецепторы гормонов щитовидной железы - TR (и RAR также) находятся в
Рецепторы гормонов щитовидной железы - TR (и RAR также) находятся в

Метаболические эффекты
Т3
В ядре
↑транскрипцию и трансляцию белков – ферментов ОВР
В костной ткани
Метаболические эффекты
Т3
В ядре
↑транскрипцию и трансляцию белков – ферментов ОВР
В костной ткани

Гормон Т2
Рецепторы для этого гормона находятся в митохондриях.
По силе
Гормон Т2
Рецепторы для этого гормона находятся в митохондриях.
По силе



Эндемический зоб - ИДС
Эндемический зоб - ИДС


Пути коррекции йоддефицитных состояний
Областная программа «Профилактика эндемического зоба и других йоддефицитных
Пути коррекции йоддефицитных состояний
Областная программа «Профилактика эндемического зоба и других йоддефицитных
 Рослинні жири Підготувала учениця 11-А класу Бойко Наталія
Рослинні жири Підготувала учениця 11-А класу Бойко Наталія  Пищеварение. Этапы пищеварения:
Пищеварение. Этапы пищеварения: Немембранные органоиды
Немембранные органоиды Голосовой аппарат Тема урока:
Голосовой аппарат Тема урока:  Презентация на тему Удобрения
Презентация на тему Удобрения Календарь прилёта наших перелётных птиц. Часть четвёртая
Календарь прилёта наших перелётных птиц. Часть четвёртая Органы кровеносной и лимфатической систем. Круги кровообращения
Органы кровеносной и лимфатической систем. Круги кровообращения Видоизменения корней
Видоизменения корней Учение о черепе - краниология
Учение о черепе - краниология Витамины (11 класс)
Витамины (11 класс) Проект освоения участка леса для осуществления рекреационного лесопользования
Проект освоения участка леса для осуществления рекреационного лесопользования Кошачий сосальщик (Opisthorchis felineus), марита
Кошачий сосальщик (Opisthorchis felineus), марита Вітаміни та їх роль в організмі тварин
Вітаміни та їх роль в організмі тварин Биологическая и социальная природа человека. Науки об организме человека
Биологическая и социальная природа человека. Науки об организме человека Развитие психики в онтогенезе. Постнатальный период. Ранний постнатальный период
Развитие психики в онтогенезе. Постнатальный период. Ранний постнатальный период ДЕНЬ ПТИЦ. Весна идет! Рассеялись туманы - Кто знает, где теперь их пелена И вновь видны луга, поля, поляны. Весна идет! В наш край и
ДЕНЬ ПТИЦ. Весна идет! Рассеялись туманы - Кто знает, где теперь их пелена И вновь видны луга, поля, поляны. Весна идет! В наш край и Презентация на тему "Биологически активные соединения живых организмов" - скачать презентации по Биологии
Презентация на тему "Биологически активные соединения живых организмов" - скачать презентации по Биологии Первичнополостные животные. Первичная и вторичная (целом) полости тела
Первичнополостные животные. Первичная и вторичная (целом) полости тела Выращивание картофельных культур в средних суглинистых почвах
Выращивание картофельных культур в средних суглинистых почвах Ядовитые растения. Омела белая
Ядовитые растения. Омела белая Презентация на тему Бактериологическое оружие и его воздействие на организм человека.
Презентация на тему Бактериологическое оружие и его воздействие на организм человека.  Тема: Генетика развития
Тема: Генетика развития Степная Пищуха из Красной книги Челябинской области
Степная Пищуха из Красной книги Челябинской области Отряд Хоботные
Отряд Хоботные Endangered animals
Endangered animals Полушария большого мозга
Полушария большого мозга Зміст і задачі фізіології, її зв’язок з іншими науками
Зміст і задачі фізіології, її зв’язок з іншими науками Бактерии. Признаки бактерий
Бактерии. Признаки бактерий