- Строение бактериальной клетки. Отличия прокариот от эукариот

Содержание
- 2. Бактериальная клетка состоит из клеточной стенки, цитоплазматической мембраны, цитоплазмы с включениями и ядерного аппарата, называемого нуклеоидом.
- 3. Клеточная стенка В клеточной стенки грамположительных бактерий содержится небольшое количество полисахаридов, липидов, белков. Основным компонентом клеточной
- 4. Цитоплазматическая мембрана Цитоплазматическая мембрана при электронной микроскопии ультратонких срезов представляет собой трехслойную мембрану (2 темных слоя
- 5. Цитоплазма Цитоплазма состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул — рибосом, ответственных
- 6. Нуклеоид Нуклеоид — эквивалент ядра у бактерий. Он расположен в центральной зоне бактерий в виде двунитевой
- 7. Капсула, микрокапсула, слизь Капсула - слизистая структура толщиной более 0,2мкм, прочно связанная с клеточной стенкой бактерий
- 8. Жгутики Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической
- 9. Пили Пили (фимбрии, ворсинки) - нитевидные образования, более тонкие и короткие (3-10нм х 0, 3-10мкм) ,
- 10. Споры Споры - своебразная форма покоящихся фирмикутных бактерий, т.е. бактерий с грамположительным типом строения клеточной стенки.
- 12. Скачать презентацию
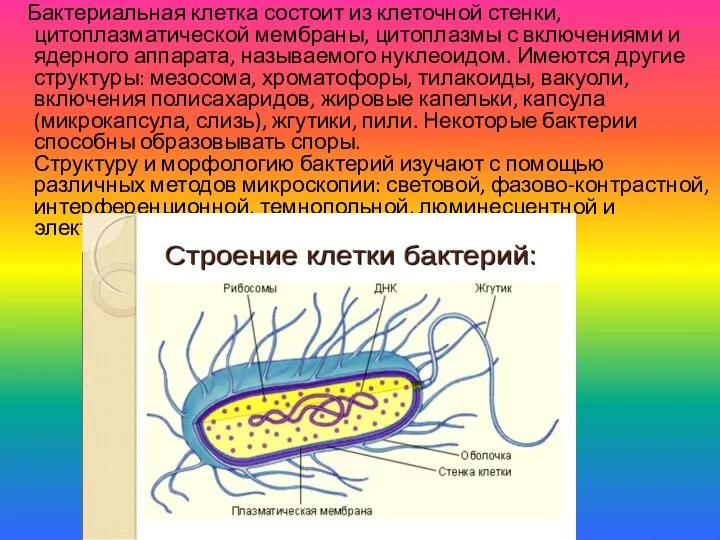
Бактериальная клетка состоит из клеточной стенки, цитоплазматической мембраны, цитоплазмы с
Бактериальная клетка состоит из клеточной стенки, цитоплазматической мембраны, цитоплазмы с

Клеточная стенка
В клеточной стенки грамположительных бактерий содержится небольшое количество полисахаридов, липидов,
Клеточная стенка
В клеточной стенки грамположительных бактерий содержится небольшое количество полисахаридов, липидов,

Цитоплазматическая мембрана
Цитоплазматическая мембрана при электронной микроскопии ультратонких срезов представляет собой трехслойную
Цитоплазматическая мембрана
Цитоплазматическая мембрана при электронной микроскопии ультратонких срезов представляет собой трехслойную

Цитоплазма
Цитоплазма состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких
Цитоплазма
Цитоплазма состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких

Нуклеоид
Нуклеоид — эквивалент ядра у бактерий. Он расположен в центральной зоне
Нуклеоид
Нуклеоид — эквивалент ядра у бактерий. Он расположен в центральной зоне

Капсула, микрокапсула, слизь
Капсула - слизистая структура толщиной более 0,2мкм, прочно связанная
Капсула, микрокапсула, слизь
Капсула - слизистая структура толщиной более 0,2мкм, прочно связанная

Жгутики
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити,
Жгутики
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити,

Пили
Пили (фимбрии, ворсинки) - нитевидные образования, более тонкие и короткие (3-10нм
Пили
Пили (фимбрии, ворсинки) - нитевидные образования, более тонкие и короткие (3-10нм

Споры
Споры - своебразная форма покоящихся фирмикутных бактерий, т.е. бактерий
с грамположительным типом
Споры
Споры - своебразная форма покоящихся фирмикутных бактерий, т.е. бактерий с грамположительным типом
 Болезни аквариумных рыб Выполнил: ученик 9 а класса Волкотрубенко Михаил.
Болезни аквариумных рыб Выполнил: ученик 9 а класса Волкотрубенко Михаил. Развитие жизни на Земле. Эры
Развитие жизни на Земле. Эры Stages of the energy metabolism
Stages of the energy metabolism Пингвины. Семейство пингвиновых
Пингвины. Семейство пингвиновых Тіршіліктің пайда болуы
Тіршіліктің пайда болуы Этапы урока. Организация начала занятия. Проверка выполнения домашнего задания. Подготовка к усвоению новых знаний Изучение
Этапы урока. Организация начала занятия. Проверка выполнения домашнего задания. Подготовка к усвоению новых знаний Изучение  Головной мозг, его строение и функции
Головной мозг, его строение и функции Эмбриональная смертность и спонтанные аборты у собак
Эмбриональная смертность и спонтанные аборты у собак Внешнее строение птиц (мат.к лабор. работе)
Внешнее строение птиц (мат.к лабор. работе) Ю. Герман. К 110-летию со дня рождения писателя
Ю. Герман. К 110-летию со дня рождения писателя Презентация по биологии Строение клетки
Презентация по биологии Строение клетки  Перелётные птицы
Перелётные птицы Презентация на тему Семейство кошачьих Презентация к уроку
Презентация на тему Семейство кошачьих Презентация к уроку МУНИЦИПАЛЬНОЕ ОБЩЕОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ОДИНЦОВСКАЯ ГИМНАЗИЯ № 11 ВИРУСЫ – НЕКЛЕТОЧНАЯ ФОРМА ЖИЗНИ…
МУНИЦИПАЛЬНОЕ ОБЩЕОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ОДИНЦОВСКАЯ ГИМНАЗИЯ № 11 ВИРУСЫ – НЕКЛЕТОЧНАЯ ФОРМА ЖИЗНИ…  Общая характеристика некоторых представителей подкласса Розиды. Лекция 6-7. Часть 2
Общая характеристика некоторых представителей подкласса Розиды. Лекция 6-7. Часть 2 Игра Один-много
Игра Один-много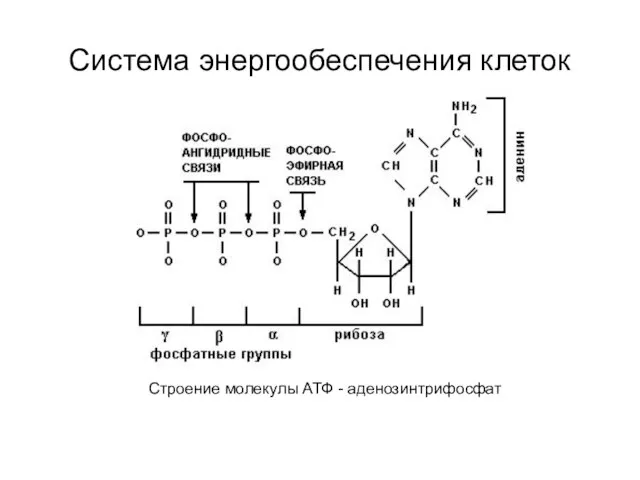 Система энергообеспечения клеток
Система энергообеспечения клеток Скелетная система
Скелетная система Экологическая токсикокинетика и биоаккумуляция ксенобиотиков. Токсическое действие СОЗ
Экологическая токсикокинетика и биоаккумуляция ксенобиотиков. Токсическое действие СОЗ Анатомия. Дыхательная система
Анатомия. Дыхательная система Аскомицеты. Характерные особенности аскомицетов
Аскомицеты. Характерные особенности аскомицетов Интродукция. Инвазивные виды
Интродукция. Инвазивные виды Будова прокаріотів
Будова прокаріотів  Разбор задания 31-32 ОГЭ-9 биология
Разбор задания 31-32 ОГЭ-9 биология Особенности организации многоклеточных животных
Особенности организации многоклеточных животных История болезни. Анамнез болезни животного
История болезни. Анамнез болезни животного Эмбриология. Периоды развития организма
Эмбриология. Периоды развития организма Человек и природа Урок 1. Биологическое и социальное в человеке.
Человек и природа Урок 1. Биологическое и социальное в человеке.