- Транскрипция

Содержание
- 2. Транскрипция - это синтез всех видов РНК по матрице ДНК, осуществляемый ферментом ДНК зависимой РНК-полимеразой. Принципы
- 3. Промотор - участок гена, на который происходит посадка РНК-полимеразы Терминатор – участок гена, на котором происходит
- 4. Терминация У бактерий есть два механизма терминации транскрипции: – ро-зависимый механизм, при котором белок Rho (ро)
- 5. Ингибиторы транскрипции • Рифампицин - ингибитор инициации. Связывается с центром инициации E. сoli. • Стрептолидигин -
- 6. Транскрипция у прокариот Прокариоты не имеют ядерной мембраны, поэтому процессы транскрипции, трансляции могут проходить одновременно. Прокариотической
- 7. Регуляция транскрипции у прокариот Лактозный оперон, или lac оперон - участок генома бактерий, отвечающий за усвоение
- 8. В состав оперона входят: • Группа сцепленных структурных генов, кодирующих синтез ферментов для единого метаболического процесса
- 9. Концепцию оперона для прокариот предложили в 1961 году французские ученые Жакоб и Моно, за что получили
- 10. Позитивная и негативная регуляция экспрессии генов Когда благодаря действию специфических регуляторных элементов уровень экспрессии генов возрастает,
- 11. Регуляция экспрессии lac оперона E. coli: а – участок управления транскрипцией, б – в отсутствие лактозы
- 12. Схема негативной индукции Жакоба и Моно Эта схема называется так потому, что контролирующим транскрипцию фактором является
- 13. 1. При отсутствии лактозы активный белок-репрессор связывается с оператором и блокирует синтез мРНК, кодирующей ферменты катаболизма
- 14. Регуляции транскрип-ции у эука-риот
- 15. Амплификация – это увеличение количества генов, точнее многократное копирование одного гена. Естественно, все полученные копии равнозначны
- 16. Энхансеры и сайленсеры - участки связывания активаторов и репрессоров Энхансер + активатор = активация транскрипции Сайленсер
- 17. Транскрип-ционные факторы Транскрипционные факторы регулируют цис-действующие элементы !Цис-действующие элементы (или цис-элементы) — участки ДНК или РНК,
- 18. Высоко консервативные, специальные структуры, связывающие ДНК и названные цинковыми пальцами (Zinc finger), лейциновой застежкой (Leucine zipper),
- 19. Отличия транскрипции у про- и эукариот Разобщение транскрипции и трансляции в пространстве и времени у эукариот,
- 21. В ходе созревания пре-мРНК в ядре сшиваться могут не все экзоны, и в разных условиях разные
- 23. Тканеспецифичность
- 24. Обратная транскрипция (или синтез кДНК) выполняется при участии фермента ревертазы на матрице РНК, в результате чего
- 25. Схема обратной транскрипции ВИЧ. Участок 3’-конца тРНК (сиреневый трилистник) комплементарен области PB (от англ. primer binding
- 27. Скачать презентацию

Транскрипция - это синтез всех видов РНК по матрице ДНК, осуществляемый
Транскрипция - это синтез всех видов РНК по матрице ДНК, осуществляемый
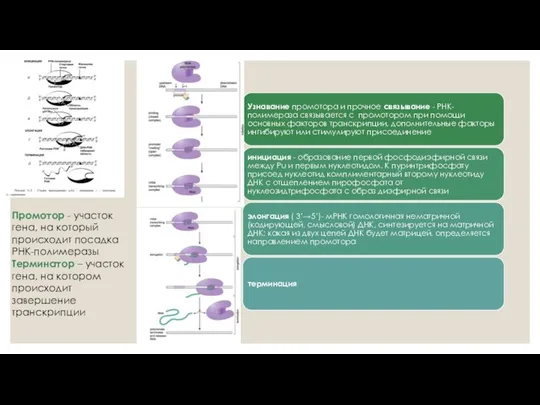
Промотор - участок гена, на который происходит посадка РНК-полимеразы Терминатор –
Промотор - участок гена, на который происходит посадка РНК-полимеразы Терминатор –
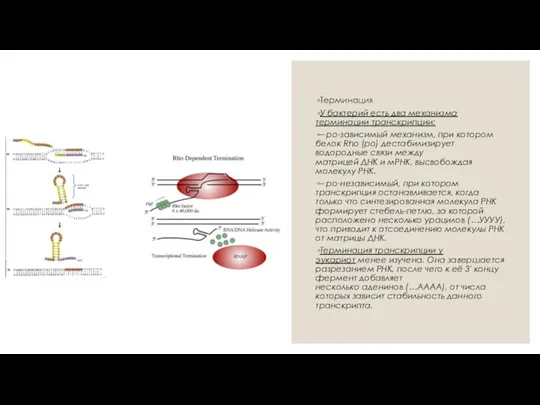
Терминация
У бактерий есть два механизма терминации транскрипции:
– ро-зависимый механизм, при котором белок
Терминация
У бактерий есть два механизма терминации транскрипции:
– ро-зависимый механизм, при котором белок

Ингибиторы транскрипции
• Рифампицин - ингибитор инициации. Связывается с центром инициации
Ингибиторы транскрипции
• Рифампицин - ингибитор инициации. Связывается с центром инициации
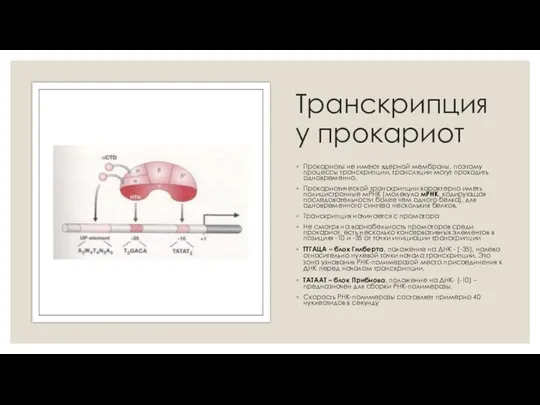
Транскрипция у прокариот
Прокариоты не имеют ядерной мембраны, поэтому процессы транскрипции, трансляции
Транскрипция у прокариот
Прокариоты не имеют ядерной мембраны, поэтому процессы транскрипции, трансляции

Регуляция транскрипции у прокариот
Лактозный оперон, или lac оперон - участок генома бактерий, отвечающий
Регуляция транскрипции у прокариот
Лактозный оперон, или lac оперон - участок генома бактерий, отвечающий

В состав оперона входят:
• Группа сцепленных структурных генов, кодирующих синтез
В состав оперона входят:
• Группа сцепленных структурных генов, кодирующих синтез
Концепцию оперона для прокариот предложили в 1961 году французские ученые Жакоб
Концепцию оперона для прокариот предложили в 1961 году французские ученые Жакоб

Позитивная и негативная регуляция экспрессии генов
Когда благодаря действию специфических регуляторных элементов
Позитивная и негативная регуляция экспрессии генов
Когда благодаря действию специфических регуляторных элементов
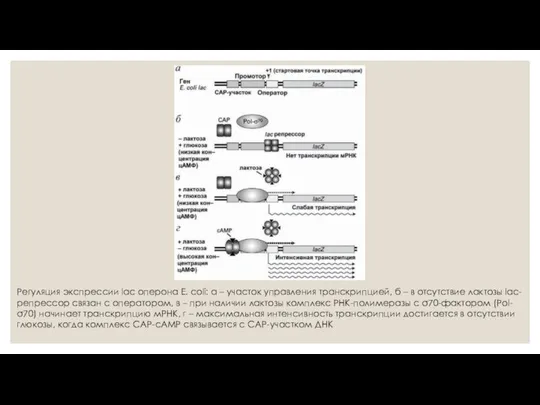
Регуляция экспрессии lac оперона E. coli: а – участок управления транскрипцией,
Регуляция экспрессии lac оперона E. coli: а – участок управления транскрипцией,

Схема негативной индукции Жакоба и Моно
Эта схема называется так потому, что
Схема негативной индукции Жакоба и Моно
Эта схема называется так потому, что
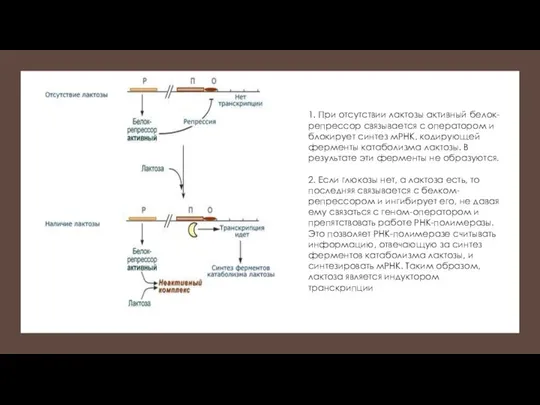
1. При отсутствии лактозы активный белок-репрессор связывается с оператором и блокирует
1. При отсутствии лактозы активный белок-репрессор связывается с оператором и блокирует

Регуляции транскрип-ции у эука-риот
Регуляции транскрип-ции у эука-риот

Амплификация – это увеличение количества генов, точнее многократное копирование одного гена.
Амплификация – это увеличение количества генов, точнее многократное копирование одного гена.

Энхансеры и сайленсеры
- участки связывания активаторов и репрессоров
Энхансер + активатор
Энхансеры и сайленсеры
- участки связывания активаторов и репрессоров
Энхансер + активатор

Транскрип-ционные факторы
Транскрипционные факторы регулируют цис-действующие элементы
!Цис-действующие элементы (или цис-элементы) — участки ДНК или РНК, регулирующие экспрессию
Транскрип-ционные факторы
Транскрипционные факторы регулируют цис-действующие элементы
!Цис-действующие элементы (или цис-элементы) — участки ДНК или РНК, регулирующие экспрессию

Высоко консервативные, специальные структуры, связывающие ДНК и названные цинковыми пальцами (Zinc
Высоко консервативные, специальные структуры, связывающие ДНК и названные цинковыми пальцами (Zinc

Отличия транскрипции у про- и эукариот
Разобщение транскрипции и трансляции в пространстве
Отличия транскрипции у про- и эукариот
Разобщение транскрипции и трансляции в пространстве
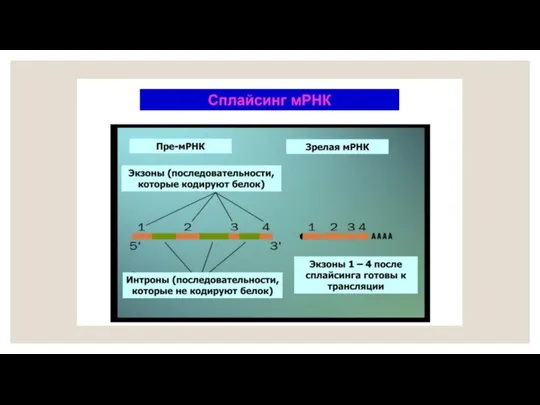

В ходе созревания пре-мРНК в ядре сшиваться могут не все экзоны,
В ходе созревания пре-мРНК в ядре сшиваться могут не все экзоны,


Тканеспецифичность
Тканеспецифичность

Обратная транскрипция (или синтез кДНК) выполняется при участии фермента ревертазы на
Обратная транскрипция (или синтез кДНК) выполняется при участии фермента ревертазы на

Схема обратной транскрипции ВИЧ.
Участок 3’-конца тРНК (сиреневый трилистник) комплементарен области PB (от англ. primer binding site — сайт
Схема обратной транскрипции ВИЧ.
Участок 3’-конца тРНК (сиреневый трилистник) комплементарен области PB (от англ. primer binding site — сайт
 Жан Батист Ламарк
Жан Батист Ламарк Пищеварительная система человека
Пищеварительная система человека Презентацию выполнила учитель высшей категории Севостьянова Г.А. МБОУ «СОШ № 1 им. Б.Н.Куликова» Г. Семикаракорск
Презентацию выполнила учитель высшей категории Севостьянова Г.А. МБОУ «СОШ № 1 им. Б.Н.Куликова» Г. Семикаракорск  Строение животной клетки
Строение животной клетки Биогеохимический цикл Фосфора
Биогеохимический цикл Фосфора  Удивительное изобретение природы
Удивительное изобретение природы  Анатомо-физиологические основы процесса дыхания
Анатомо-физиологические основы процесса дыхания Презентация на тему Половое размножение организмов. Развитие половых клеток. Оплодотворение.
Презентация на тему Половое размножение организмов. Развитие половых клеток. Оплодотворение. Формирование целостной картины мира в старшей группе Весёлые ребята. Тема: Цветы
Формирование целостной картины мира в старшей группе Весёлые ребята. Тема: Цветы Адгезия клеток/ Межклеточные контакты/
Адгезия клеток/ Межклеточные контакты/ Презентация по экологии Влияние городской среды на здоровье человека
Презентация по экологии Влияние городской среды на здоровье человека Перелётные птицы
Перелётные птицы Метаболизм - обмен веществ
Метаболизм - обмен веществ Анализаторы. Функции анализаторов
Анализаторы. Функции анализаторов Класс Двустворчатые моллюски
Класс Двустворчатые моллюски ОРГАНЫ ДЫХАНИЯ
ОРГАНЫ ДЫХАНИЯ  Кожа – наружный покровный орган
Кожа – наружный покровный орган Земноводные
Земноводные Презентация на тему Аквариум – маленькая искусственная экосистема
Презентация на тему Аквариум – маленькая искусственная экосистема  Строение стебля Познакомимся с клеточным строением стебля.
Строение стебля Познакомимся с клеточным строением стебля. Презентация на тему "Формы размножения организмов" - скачать презентации по Биологии
Презентация на тему "Формы размножения организмов" - скачать презентации по Биологии Наши кошки
Наши кошки Немембранные и двумембранные органоиды Задачи: рассмотреть особенности строения и функции немембранных и двумембранных орг
Немембранные и двумембранные органоиды Задачи: рассмотреть особенности строения и функции немембранных и двумембранных орг Методы определения микробной биомассы в почве
Методы определения микробной биомассы в почве Ч. Дарвин о причинах эволюции животного мира
Ч. Дарвин о причинах эволюции животного мира Магония падуболистная
Магония падуболистная Лектор: ассистент кафедры медицинской биологии, к.б.н. Гоч Инна Викторовна
Лектор: ассистент кафедры медицинской биологии, к.б.н. Гоч Инна Викторовна  Презентация на тему "Биоиндикация" - скачать презентации по Биологии
Презентация на тему "Биоиндикация" - скачать презентации по Биологии