- тұқымқуалайтын ақпараттың жүзеге асырылуы (репликация,транскрипция және трансляция). Дәріс 2

Содержание
- 2. Жоспары: Тұқым қуалайтын ақпараттың берілу бағыттары, Крик постулаты. Репликация, анықтамасы және кезеңдері. Қатысатын ферменттер. Транскрипция, кезеңдері.
- 3. Әдебиеттер: Б. Албертс, Д Брей, Дж. Льюис и др. Молекулярная биология клетки т. 2 «Мир» 1986
- 4. Тұқым қуалайтын ақпараттың берілу бағыттары, Крик постулаты: ДНҚ-нан → ДНҚ-на (репликация); ДНҚ-нан → РНҚ-на (транскрипция); РНҚ-нан
- 5. ДНҚ репликациясы деп оның өзін-өзі еселеуін айтады. Репликация кезінде ДНҚ молекуласынан ұрпақтан-ұрпаққа берілетін қатесіз көшірмесі алынады.
- 6. Эукариоттардағы сызық тәрізді ДНҚ молекуласының еселену жолдары: Оларда полирепликондар түрінде жүреді. Репликация бір немесе бірнеше нүктелерде
- 7. ДНҚ-молекуласының бір және екі ашалық репликациялану түрлері:
- 8. Репликацияның іске асырылу принциптері: Жартылай консервативті; Антипаралельді; Симметриялы; Полярлы; Үзілмелі; Комплементарлы;
- 9. Репликацияның кезеңдері: Инициация. Ферменттері― геликаза, топоизомераза, SSВ- ақуыздары және т.б. Олар аналық ДНҚ молекуласындағы репликация басталатын
- 10. Элонгация. Ферменттері―РНҚ –праймаза, РНҚ праймер,ДНҚ полимеразалар, лигаза және т.б. Олардың қатысуымен ажыратылған аналық тізбектерге комлементарлы,антипаралель жаңа
- 11. РНҚ-праймер ДНҚ тізбегінің 3׳- ұшына ДНҚ-полимеразаны қосады. ДНҚ-полимераза аналық тізбекке комплементарлық принципте жаңа тізбекті синтездейді. 3׳
- 12. ДНҚ РЕПЛИКАЦИЯСЫ
- 13. Репликацияның маңызы: Аналық ДНҚ-ның жаңа екі көшірмесі пайда болады; Олар бөліну кезінде жаңа жасушаларға беріледі; Репликация
- 14. Транскрипция Ақуыз туралы генетикалық информацияның іске асырылуы екі сатыдан – транскрипция және трансляциядан. Транскрипция-тұқымқуалайтын ақпараттың іске
- 15. Транскрипция арнайы РНҚ-полимераза ферментінің қатысуымен жүреді. Ол фермент транскрипциланатын учаскенің басталатын жерін промоторды танып, онымен байланысады;
- 16. РНҚ-полимеразалардың құрылысы мен қызметтері: РНК- полимераза про-және эукариоттық ағзалардың бәрінде табылған. Ішек таяқшасынан (E. coli) бөлініп
- 17. δ – суббірлік РНҚ полимеразаның транскрипция басталатын ДНҚ арнайы учаскесін – промоторды тану (транскрипцияның инициациясы) үшін
- 18. Бактериядағы РНҚ-полимеразамен жүретін ДНҚ-ның транскрипциясы
- 19. РНҚ –полимераза ферментінің ерекшеліктері: - δ-суббірлігінің көмегімен ДНҚ –ның транскрипцияланатын тізбегін таңдап және оның басталу нүктесін
- 20. Эукариот жасушаларында үш түрлі РНҚ –полимеразалар (I,II,III) кездеседі. Олардың құрылыстары күрделі, құрамдарында 10-15 суббірліктері болуы мүмкін.
- 21. Транскрипцияның этаптары: 1.Инициация- δ-факторы бар РНҚ – полимеразаның ДНҚ-тізбегіндегі промоторды танитын және онымен байланысатын транскрипцияның бірінші
- 22. Промотор
- 23. «Көзше» маңындағы ДНҚ матрицалық тізбегіндегі нуклеотидтер рНТФ-пен (рибонуклеозидтрифосфатпен) байланысуына мүмкіндік пайда болады. Синтезделе бастаған РНҚ-ға бірінші
- 24. Транскрипциялық кешен (ДНҚ – РНҚ полимераза- РНҚ ) алдындағы ДНҚ жіпшесі ажырайды; Оның артынан синтезделген РНҚ-ның
- 25. Транскрипцияланатын геннің бойында бірінен кейін бірі қозғалатын бірнеше РНҚ-полимеразалар конвейерлік әдіспен жұмыс істейді. Олардың арасындағы қашықтығы
- 26. Эукариоттар транскрипциясының ерекшеліктері: Эукариоттардың хромосомаларында нуклеосомдық құрылыс болғандықтан, ДНҚ-ның транскрипция жүретін жерлерін гистондардан ажырататын тәсілдер керек.
- 27. РНК – полимераза׀׀ -мен транскрипцияланатын геннің реттеуші бөлігі құрылымының сызба нұсқасы:
- 29. Транскрипцияның басталу нүктесіне ең жақын орналасатыны ТАТА – домен, Хогнесс домені деп аталады. Одан кейін ЦААТ
- 31. ЭУКАРИОТТАРДЫҢ транскрипциялық факторлары: Инициацияға қатысатын ақуыздық факторлар: IF-1, IF-2, IF-3; Элонгацияға қатысатын ақуыздық факторлар: ЕF-1, EF-2;
- 33. Транскрипцияның өнімдері Эукариоттарда транскрипция нәтижесінде пре-м РНҚ, пре-р РНҚ, пре-т РНҚ синтезделеді. Пре – м РНҚ
- 35. мРНҚ процессингі (пісіп жетілуі) Процессингке мРНҚ молекуласының метелденуі және кэптенуі, полиаденилденуі, сплайсинг және тексерілу жатады. Пре-мРНҚ
- 36. Ажыратылған генмен кодталатын ақпараттың ДНҚ-дан ақуызға берілу үрдісі:
- 37. Альтернативті сплайсинг - Бір геннің экзондарының түрліше комбинациялануы нәтижесінде әртүрлі мРНҚ пайда болады. Альтернативтік сплайсинг бір
- 38. Бақылау сұрақтары: Репликация қай уақытта жүреді? Репликацияның қандай этаптары бар ? Репликацияға қайдай ферменттер қатысады ?
- 39. ГЕНЕТИКАЛЫҚ КОД: Полипептидтік тізбектегі аминқышқылдарының орналасу ретін анықтайтын ДНҚ тізбегіндегі нуклеотидтер қатарынан тұратын жүйе. ДНҚ-дағы “4
- 40. ГЕНЕТИКАЛЫҚ КОДТЫҢ ҚАСИЕТТЕРІ: ТРИПЛЕТТІЛІГІ. ДНҚ тізбегіндегі қатар тұрған үш нуклеотид-триплет(кодон) бір аминқышқылын анықтайды. Триплет- генетикалық кодтың
- 41. Ақуыздың синтезі РИБОСОМАДА жүреді. Рибосома екі суббөліктен тұрады. Екі бөлік бірігіп,қосылғанда рибосоманың пішіні жүрекке ұқсайды.Оң жақта
- 42. Рибосоманың функциялық орталықтары: ‹ ̶ Рибосомадағы орталықтардың орналасуы. ‹̶ Функциялық орталықтардың рибосома суббөліктерінде орналасуы.
- 43. ТРАНСЛЯЦИЯҒА ҚАТЫСАТЫН ФАКТОРЛАР: Инициация факторлары (Initiation factor – IF); Элонгация факторлары(Elongation factor – EF); Терминация факторлары
- 44. Ақуыздың синтезіндегі негізгі сатылар: Инициация немесе трансляцияның басталуы; Элангация немесе полипептидтік тізбектің ұзаруы; Терминация немесе трансляцияның
- 45. Трансляцияның басталуы мен инициациялық комплекстың түзілуі қатар жүреді: м-РНҚ (өзінің 5‘ – трансляцияланбайтын учаскесімен) рибосоманың кіші
- 46. Т РНҚ
- 47. Аминқышқылдарының (АҚ) тРНҚ-мен байланысуы; Е – аминоацил-тРНК-синтетаза. антикодон
- 48. Элонгация циклының сатылары:
- 49. ПЕПТИДИЛТРАНСФЕРАЗАЛЫҚ РЕАКЦИЯНЫҢ КЕЗІНДЕ ПЕПТИДТІК БАЙЛАНЫСТЫҢ ТҮЗІЛУІ:
- 50. Трансляцияның терминациясы: (5‘) М- РНК Рибосомада “мағнасыз” кодондардың: УАА, УАГ немесе УГА бірінің пайда болуы терми-
- 51. БҰЛ кодон терминацияның – e RF факторларымен танылады. Мұндай факторлар екеу: 1. УАА и УАГ кодондар
- 52. Ақуыз синтезделу үшін рибосомаға қажет: Ақуыздың полипептидтік тізбегіндегі аминқышқылдары орналасу реті бар бағдарлама; Ақуыз түзілуіне керек
- 53. Посттрансляциялық модификация (фолдинг) Рибосомада синтезделген ақуыз бірінші құрылымдық деңгейде болады. Бұл полипептидтік тізбек, ешбір қызмет атқара
- 55. Скачать презентацию

Жоспары:
Тұқым қуалайтын ақпараттың берілу бағыттары, Крик постулаты.
Репликация, анықтамасы және кезеңдері.
Жоспары:
Тұқым қуалайтын ақпараттың берілу бағыттары, Крик постулаты.
Репликация, анықтамасы және кезеңдері.

Әдебиеттер:
Б. Албертс, Д Брей, Дж. Льюис и др. Молекулярная биология клетки
Әдебиеттер:
Б. Албертс, Д Брей, Дж. Льюис и др. Молекулярная биология клетки

Тұқым қуалайтын ақпараттың берілу бағыттары, Крик постулаты:
ДНҚ-нан → ДНҚ-на (репликация);
ДНҚ-нан →
Тұқым қуалайтын ақпараттың берілу бағыттары, Крик постулаты:
ДНҚ-нан → ДНҚ-на (репликация);
ДНҚ-нан →

ДНҚ репликациясы деп оның өзін-өзі еселеуін айтады.
Репликация кезінде ДНҚ молекуласынан
ДНҚ репликациясы деп оның өзін-өзі еселеуін айтады.
Репликация кезінде ДНҚ молекуласынан

Эукариоттардағы сызық тәрізді ДНҚ молекуласының еселену жолдары:
Оларда полирепликондар түрінде жүреді.
Репликация бір
Эукариоттардағы сызық тәрізді ДНҚ молекуласының еселену жолдары:
Оларда полирепликондар түрінде жүреді.
Репликация бір
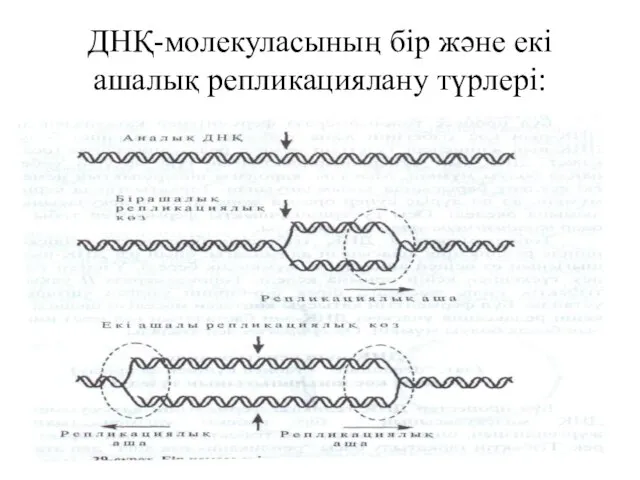
ДНҚ-молекуласының бір және екі ашалық репликациялану түрлері:
ДНҚ-молекуласының бір және екі ашалық репликациялану түрлері:

Репликацияның іске асырылу принциптері:
Жартылай консервативті;
Антипаралельді;
Симметриялы;
Полярлы;
Үзілмелі;
Комплементарлы;
Репликацияның іске асырылу принциптері:
Жартылай консервативті;
Антипаралельді;
Симметриялы;
Полярлы;
Үзілмелі;
Комплементарлы;

Репликацияның кезеңдері:
Инициация. Ферменттері― геликаза, топоизомераза, SSВ- ақуыздары және т.б. Олар аналық
Репликацияның кезеңдері:
Инициация. Ферменттері― геликаза, топоизомераза, SSВ- ақуыздары және т.б. Олар аналық

Элонгация. Ферменттері―РНҚ –праймаза, РНҚ праймер,ДНҚ полимеразалар, лигаза және т.б. Олардың қатысуымен

РНҚ-праймер ДНҚ тізбегінің 3׳- ұшына ДНҚ-полимеразаны қосады.
ДНҚ-полимераза аналық тізбекке комплементарлық принципте
РНҚ-праймер ДНҚ тізбегінің 3׳- ұшына ДНҚ-полимеразаны қосады.
ДНҚ-полимераза аналық тізбекке комплементарлық принципте
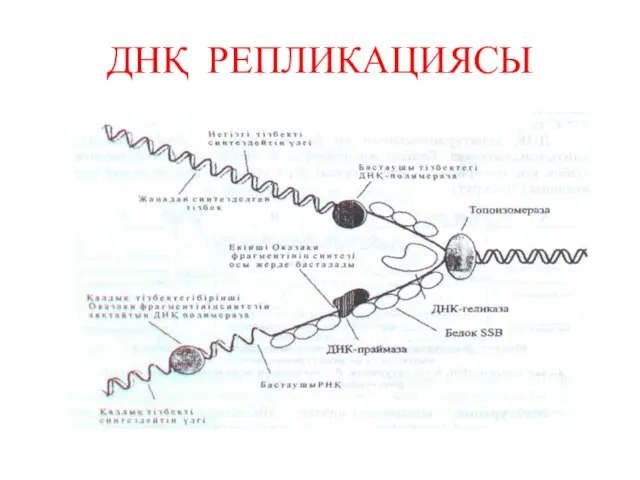
ДНҚ РЕПЛИКАЦИЯСЫ
ДНҚ РЕПЛИКАЦИЯСЫ

Репликацияның маңызы:
Аналық ДНҚ-ның жаңа екі көшірмесі пайда болады;
Олар бөліну кезінде жаңа
Репликацияның маңызы:
Аналық ДНҚ-ның жаңа екі көшірмесі пайда болады;
Олар бөліну кезінде жаңа

Транскрипция
Ақуыз туралы генетикалық информацияның іске асырылуы екі сатыдан – транскрипция және
Транскрипция
Ақуыз туралы генетикалық информацияның іске асырылуы екі сатыдан – транскрипция және

Транскрипция арнайы РНҚ-полимераза ферментінің қатысуымен жүреді.
Ол фермент транскрипциланатын учаскенің
Транскрипция арнайы РНҚ-полимераза ферментінің қатысуымен жүреді.
Ол фермент транскрипциланатын учаскенің

РНҚ-полимеразалардың құрылысы мен қызметтері:
РНК- полимераза про-және эукариоттық ағзалардың бәрінде табылған.
РНҚ-полимеразалардың құрылысы мен қызметтері:
РНК- полимераза про-және эукариоттық ағзалардың бәрінде табылған.

δ – суббірлік РНҚ полимеразаның транскрипция басталатын ДНҚ арнайы учаскесін
δ – суббірлік РНҚ полимеразаның транскрипция басталатын ДНҚ арнайы учаскесін
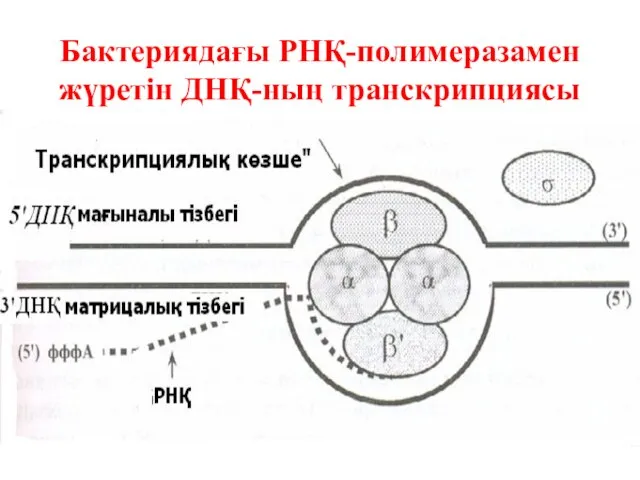
Бактериядағы РНҚ-полимеразамен жүретін ДНҚ-ның транскрипциясы
Бактериядағы РНҚ-полимеразамен жүретін ДНҚ-ның транскрипциясы

РНҚ –полимераза ферментінің ерекшеліктері:
- δ-суббірлігінің көмегімен ДНҚ –ның транскрипцияланатын
РНҚ –полимераза ферментінің ерекшеліктері:
- δ-суббірлігінің көмегімен ДНҚ –ның транскрипцияланатын

Эукариот жасушаларында үш түрлі РНҚ –полимеразалар (I,II,III) кездеседі. Олардың құрылыстары
Эукариот жасушаларында үш түрлі РНҚ –полимеразалар (I,II,III) кездеседі. Олардың құрылыстары

Транскрипцияның этаптары:
1.Инициация- δ-факторы бар РНҚ – полимеразаның ДНҚ-тізбегіндегі промоторды танитын және
Транскрипцияның этаптары:
1.Инициация- δ-факторы бар РНҚ – полимеразаның ДНҚ-тізбегіндегі промоторды танитын және

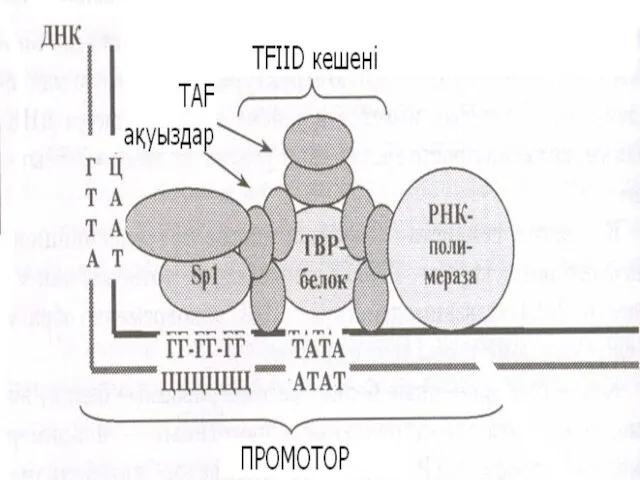
Промотор
Промотор

«Көзше» маңындағы ДНҚ матрицалық тізбегіндегі нуклеотидтер рНТФ-пен (рибонуклеозидтрифосфатпен) байланысуына мүмкіндік
«Көзше» маңындағы ДНҚ матрицалық тізбегіндегі нуклеотидтер рНТФ-пен (рибонуклеозидтрифосфатпен) байланысуына мүмкіндік

Транскрипциялық кешен (ДНҚ – РНҚ полимераза- РНҚ ) алдындағы ДНҚ
Транскрипциялық кешен (ДНҚ – РНҚ полимераза- РНҚ ) алдындағы ДНҚ
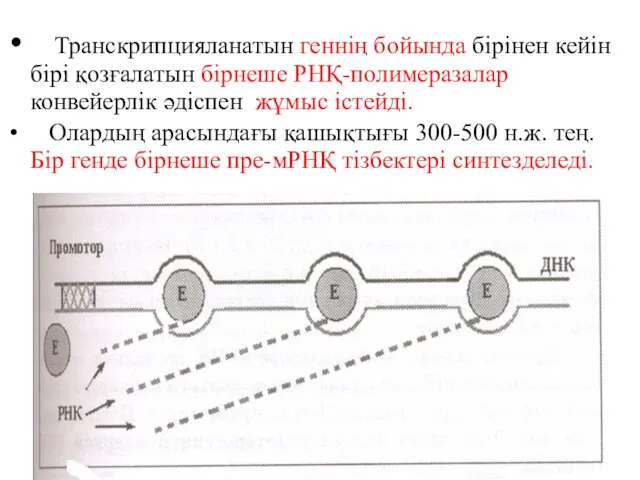
Транскрипцияланатын геннің бойында бірінен кейін бірі қозғалатын бірнеше РНҚ-полимеразалар конвейерлік
Транскрипцияланатын геннің бойында бірінен кейін бірі қозғалатын бірнеше РНҚ-полимеразалар конвейерлік

Эукариоттар транскрипциясының ерекшеліктері:
Эукариоттардың хромосомаларында нуклеосомдық құрылыс болғандықтан, ДНҚ-ның транскрипция жүретін
Эукариоттар транскрипциясының ерекшеліктері:
Эукариоттардың хромосомаларында нуклеосомдық құрылыс болғандықтан, ДНҚ-ның транскрипция жүретін
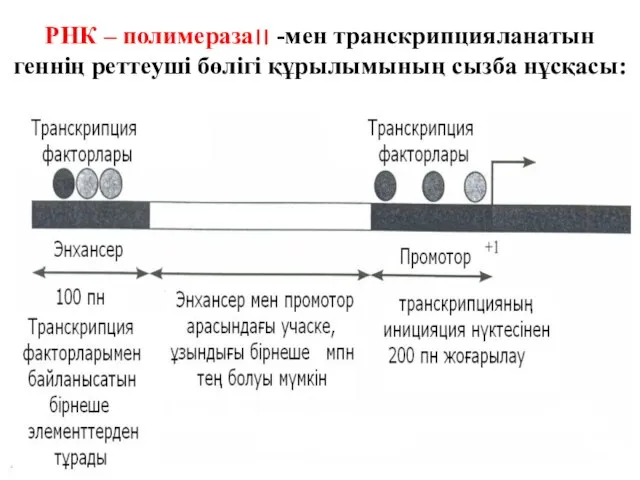
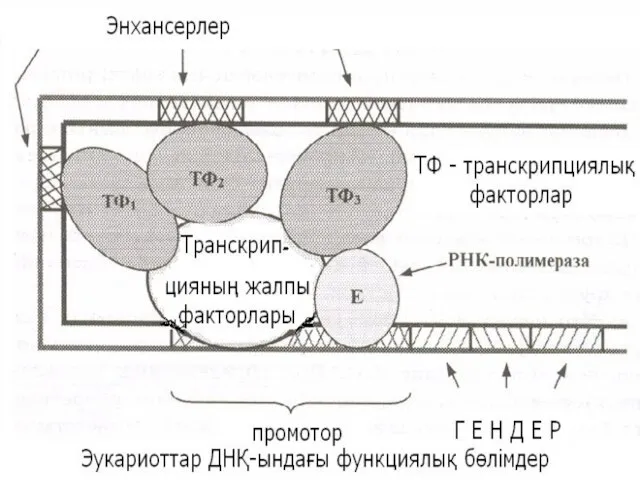
РНК – полимераза׀׀ -мен транскрипцияланатын геннің реттеуші бөлігі құрылымының сызба нұсқасы:
РНК – полимераза׀׀ -мен транскрипцияланатын геннің реттеуші бөлігі құрылымының сызба нұсқасы:
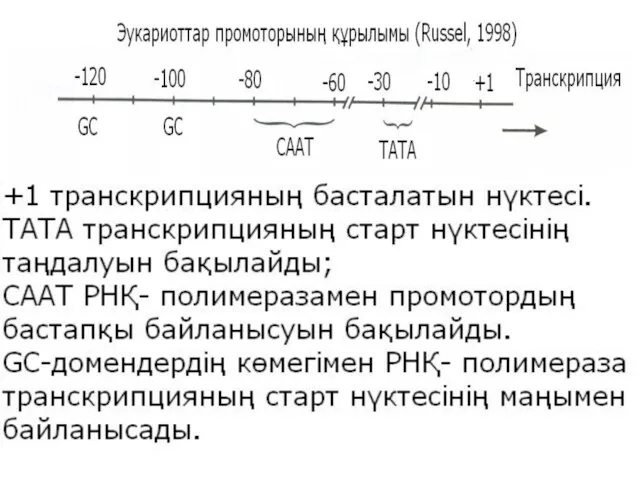

Транскрипцияның басталу нүктесіне ең жақын орналасатыны ТАТА – домен, Хогнесс домені
Транскрипцияның басталу нүктесіне ең жақын орналасатыны ТАТА – домен, Хогнесс домені

ЭУКАРИОТТАРДЫҢ транскрипциялық факторлары:
Инициацияға қатысатын ақуыздық факторлар: IF-1, IF-2, IF-3;
Элонгацияға қатысатын
ЭУКАРИОТТАРДЫҢ транскрипциялық факторлары:
Инициацияға қатысатын ақуыздық факторлар: IF-1, IF-2, IF-3;
Элонгацияға қатысатын

Транскрипцияның өнімдері
Эукариоттарда транскрипция нәтижесінде пре-м РНҚ, пре-р РНҚ, пре-т РНҚ синтезделеді.
Пре
Транскрипцияның өнімдері
Эукариоттарда транскрипция нәтижесінде пре-м РНҚ, пре-р РНҚ, пре-т РНҚ синтезделеді.
Пре
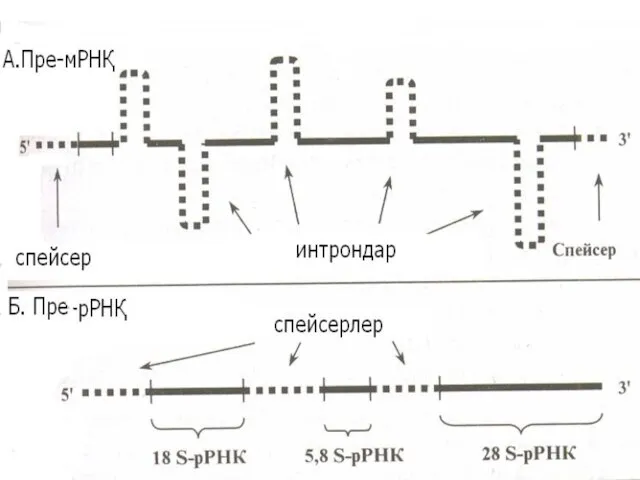

мРНҚ процессингі (пісіп жетілуі)
Процессингке мРНҚ молекуласының метелденуі және кэптенуі, полиаденилденуі, сплайсинг
мРНҚ процессингі (пісіп жетілуі)
Процессингке мРНҚ молекуласының метелденуі және кэптенуі, полиаденилденуі, сплайсинг
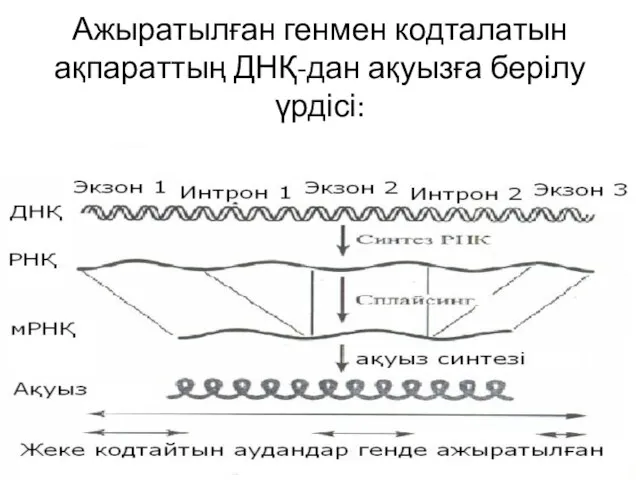
Ажыратылған генмен кодталатын ақпараттың ДНҚ-дан ақуызға берілу үрдісі:
Ажыратылған генмен кодталатын ақпараттың ДНҚ-дан ақуызға берілу үрдісі:

Альтернативті сплайсинг -
Бір геннің экзондарының түрліше комбинациялануы нәтижесінде әртүрлі мРНҚ
Альтернативті сплайсинг -
Бір геннің экзондарының түрліше комбинациялануы нәтижесінде әртүрлі мРНҚ

Бақылау сұрақтары:
Репликация қай уақытта жүреді?
Репликацияның қандай этаптары бар ?
Репликацияға қайдай
Бақылау сұрақтары:
Репликация қай уақытта жүреді?
Репликацияның қандай этаптары бар ?
Репликацияға қайдай

ГЕНЕТИКАЛЫҚ КОД:
Полипептидтік тізбектегі аминқышқылдарының орналасу ретін анықтайтын ДНҚ тізбегіндегі нуклеотидтер қатарынан
ГЕНЕТИКАЛЫҚ КОД:
Полипептидтік тізбектегі аминқышқылдарының орналасу ретін анықтайтын ДНҚ тізбегіндегі нуклеотидтер қатарынан

ГЕНЕТИКАЛЫҚ КОДТЫҢ ҚАСИЕТТЕРІ:
ТРИПЛЕТТІЛІГІ. ДНҚ тізбегіндегі қатар тұрған үш нуклеотид-триплет(кодон) бір аминқышқылын
ГЕНЕТИКАЛЫҚ КОДТЫҢ ҚАСИЕТТЕРІ:
ТРИПЛЕТТІЛІГІ. ДНҚ тізбегіндегі қатар тұрған үш нуклеотид-триплет(кодон) бір аминқышқылын
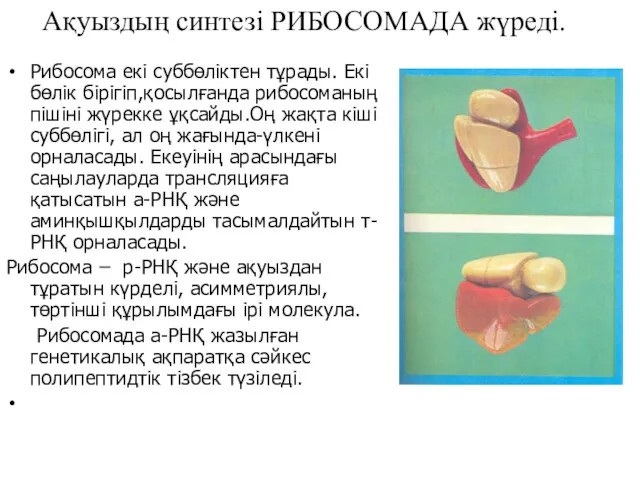
Ақуыздың синтезі РИБОСОМАДА жүреді.
Рибосома екі суббөліктен тұрады. Екі бөлік бірігіп,қосылғанда рибосоманың
Ақуыздың синтезі РИБОСОМАДА жүреді.
Рибосома екі суббөліктен тұрады. Екі бөлік бірігіп,қосылғанда рибосоманың
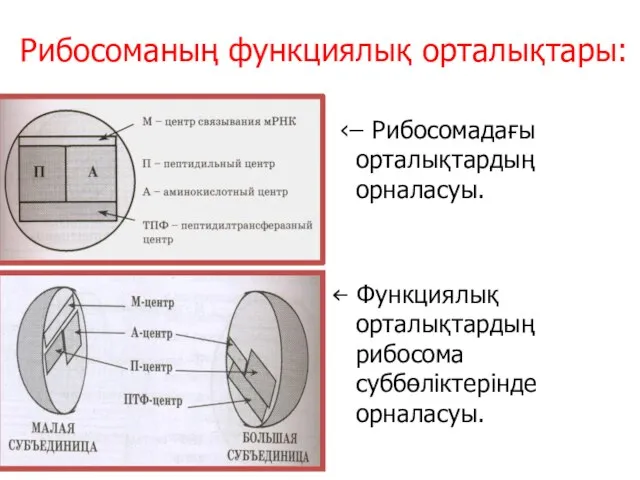
Рибосоманың функциялық орталықтары:
‹ ̶ Рибосомадағы орталықтардың орналасуы.
‹̶ Функциялық орталықтардың
Рибосоманың функциялық орталықтары:
‹ ̶ Рибосомадағы орталықтардың орналасуы.
‹̶ Функциялық орталықтардың

ТРАНСЛЯЦИЯҒА ҚАТЫСАТЫН ФАКТОРЛАР:
Инициация факторлары (Initiation factor – IF);
Элонгация факторлары(Elongation factor –
ТРАНСЛЯЦИЯҒА ҚАТЫСАТЫН ФАКТОРЛАР:
Инициация факторлары (Initiation factor – IF);
Элонгация факторлары(Elongation factor –

Ақуыздың синтезіндегі негізгі сатылар:
Инициация немесе трансляцияның басталуы;
Элангация немесе полипептидтік тізбектің ұзаруы;
Терминация
Ақуыздың синтезіндегі негізгі сатылар:
Инициация немесе трансляцияның басталуы;
Элангация немесе полипептидтік тізбектің ұзаруы;
Терминация

Трансляцияның басталуы мен инициациялық комплекстың түзілуі қатар жүреді:
м-РНҚ (өзінің
Трансляцияның басталуы мен инициациялық комплекстың түзілуі қатар жүреді:
м-РНҚ (өзінің
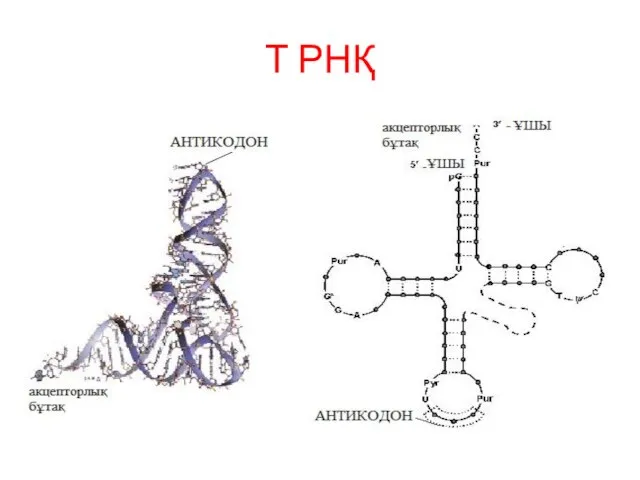
Т РНҚ
Т РНҚ
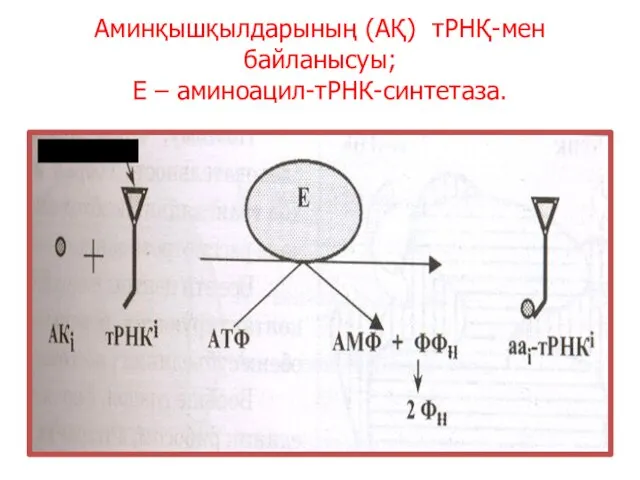
Аминқышқылдарының (АҚ) тРНҚ-мен байланысуы;
Е – аминоацил-тРНК-синтетаза.
антикодон
Аминқышқылдарының (АҚ) тРНҚ-мен байланысуы;
Е – аминоацил-тРНК-синтетаза.
антикодон
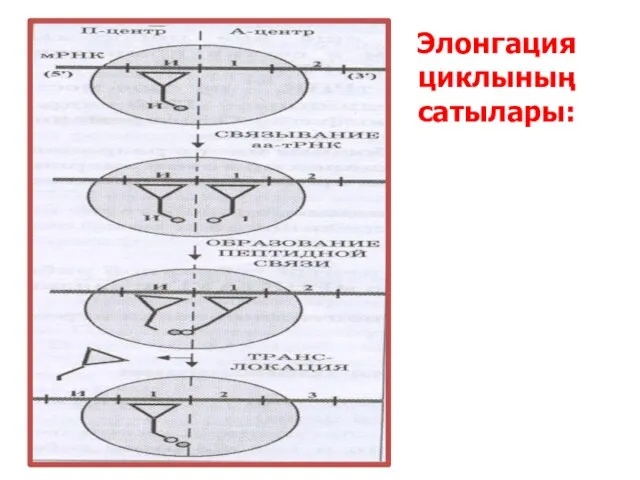
Элонгация циклының сатылары:
Элонгация циклының сатылары:
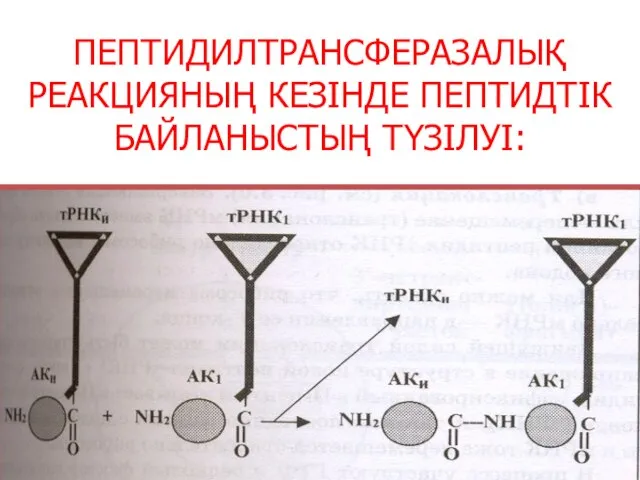
ПЕПТИДИЛТРАНСФЕРАЗАЛЫҚ РЕАКЦИЯНЫҢ КЕЗІНДЕ ПЕПТИДТІК БАЙЛАНЫСТЫҢ ТҮЗІЛУІ:
ПЕПТИДИЛТРАНСФЕРАЗАЛЫҚ РЕАКЦИЯНЫҢ КЕЗІНДЕ ПЕПТИДТІК БАЙЛАНЫСТЫҢ ТҮЗІЛУІ:
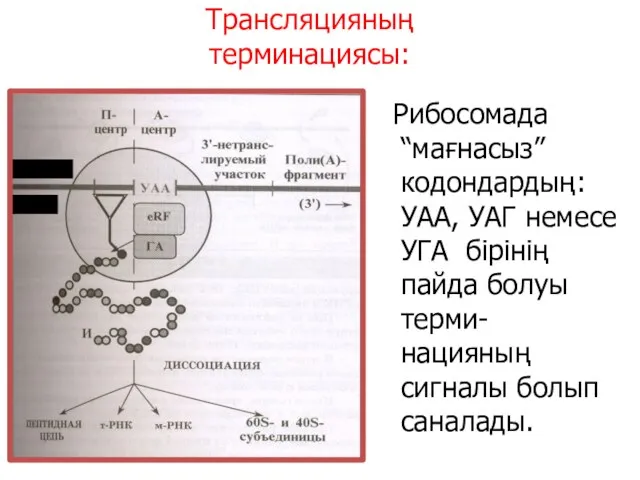
Трансляцияның терминациясы:
(5‘)
М- РНК
Рибосомада “мағнасыз” кодондардың: УАА, УАГ немесе УГА
Трансляцияның терминациясы:
(5‘)
М- РНК
Рибосомада “мағнасыз” кодондардың: УАА, УАГ немесе УГА

БҰЛ кодон терминацияның – e RF факторларымен танылады.
Мұндай факторлар екеу:
1.
БҰЛ кодон терминацияның – e RF факторларымен танылады.
Мұндай факторлар екеу:
1.

Ақуыз синтезделу үшін рибосомаға қажет:
Ақуыздың полипептидтік тізбегіндегі аминқышқылдары орналасу реті бар
Ақуыз синтезделу үшін рибосомаға қажет:
Ақуыздың полипептидтік тізбегіндегі аминқышқылдары орналасу реті бар

Посттрансляциялық модификация (фолдинг)
Рибосомада синтезделген ақуыз бірінші құрылымдық деңгейде болады. Бұл полипептидтік
Посттрансляциялық модификация (фолдинг)
Рибосомада синтезделген ақуыз бірінші құрылымдық деңгейде болады. Бұл полипептидтік
Похожие презентации
 Ракообразные Выполнила: Отряскина Т.А. Неклюдовская сош
Ракообразные Выполнила: Отряскина Т.А. Неклюдовская сош  Грибны
Грибны Отдел Моховидные
Отдел Моховидные Перелетные птицы
Перелетные птицы Органоиды клетки
Органоиды клетки Витамины
Витамины Дивергенция, конвергенция, параллелизм
Дивергенция, конвергенция, параллелизм Цитология старения. Окислительная теория старения
Цитология старения. Окислительная теория старения Прорастание семян
Прорастание семян Основные методы селекции животных
Основные методы селекции животных Тема: «Выделительная система» Задачи: Изучить особенности строения, функции и гигиену органов выделительной системы
Тема: «Выделительная система» Задачи: Изучить особенности строения, функции и гигиену органов выделительной системы Дыхание. Механизм вдоха и выдоха
Дыхание. Механизм вдоха и выдоха Презентация на тему "Нарушения нервной деятельности и профилактика заболеваний ЦНС" - скачать презентации по Биологии
Презентация на тему "Нарушения нервной деятельности и профилактика заболеваний ЦНС" - скачать презентации по Биологии Инфузории
Инфузории  Презентация по экологии Литосфера
Презентация по экологии Литосфера Вегетативная (автономная) нервная система
Вегетативная (автономная) нервная система Скелет человека
Скелет человека Класс Насекомые
Класс Насекомые Презентация на тему Здоровый образ жизни
Презентация на тему Здоровый образ жизни  Аттестационная работа. Лекарственные растения
Аттестационная работа. Лекарственные растения Основы вирусологии
Основы вирусологии Ракоподібні. Цікаві факти
Ракоподібні. Цікаві факти Розовая чайка
Розовая чайка Анатомия, физиология, психология и гигиена человека. Евгений Алексеев |2015 год
Анатомия, физиология, психология и гигиена человека. Евгений Алексеев |2015 год Царство Прокариоты
Царство Прокариоты Психогенетика как наука. Методы психогенетики
Психогенетика как наука. Методы психогенетики Этапы формирования и развития представлений о клетке
Этапы формирования и развития представлений о клетке