- Биохимия эритроцита

Содержание
- 2. АКТУАЛЬНОСТЬ Кровь транспортирует из легких в ткани около 600л О2 в сутки. Весь О2 транспорти-руется в
- 3. ПЛАН ЛЕКЦИИ 1. Биохимия эритроцита 2. Синтез гема 3. Гемоглобинопатии. Талассемии 4. Распад гемоглобина 5. Желтухи
- 4. ЭРИТРОЦИТЫ Эритроциты (erythrosytus) это форменные элементы крови. Функция эритроцитов: 1. Регуляция в крови кислотно-основного состояния, 2.
- 5. ЭРИТРОЦИТ
- 6. Плазмолемма эритроцитов Плазмолемма эритроцитов имеет толщину около 20 нм. Она состоит из примерно равного количества липидов
- 7. Липиды Бислой плазмолеммы образован: глицерофосфолипидами, сфингофосфолипидами, гликолипидами, холестерином. Внешний слой содержит гликолипиды (около 5%) и холин
- 8. Белки В плазмолемме эритроцита идентифицировано 15 главных белков. Более 60% всех мембранных белков приходится на: -
- 9. СПЕКТРИН Спектрин - основной белок цитоскелета эритроцитов. Имеет вид фибриллы, состоящей из двух антипараллельно перекрученных друг
- 10. БЕЛОК ПОЛОСЫ 3 Трансмембранный гликопротеид, его полипептидная цепь много раз пересекает бислой липидов. Является компонентом цитоскелета
- 11. ГЛИКОФОРИН трансмембранный гликопротеин, пронизывающий плазмолемму в виде одиночной спирали. Гликофорины формируют цитоскелет, Выполняют рецепторные функции.
- 12. БЕЛКИ ЭРИТРОЦИТА
- 13. Цитоплазма эритроцитов содержится около 60% воды и 40% сухого остатка. 95% сухого остатка составляет гемоглобин 5%
- 14. Особенность обмена нуклеотидов в эритроцитах Из ФРПФ (из рибозо-5ф) и аденина синтезируется АМФ. В реакциях гликолиза
- 15. Особенность углеводного обмена в эритроцитах 1. В зрелых Эр углеводы не синтезируются. 2. Катаболизм углеводов происходит
- 16. 2,3- Дифосфоглицерат (2,3-ДФГ) Синтезируется из промежуточного продукта гликолиза – 1,3-дифосфоглицерата 2,3- ДФГ - снижает сродство гемоглобина
- 17. Обезвреживание активных форм кислорода в эритроцитах Источники активных форм кислорода (АФК): 1. Ηb (Fe2+) → Мet
- 18. ГЕМ - это порфирин, в центре которого находиться Fe2+. Fe2+ включается в молекулу порфирина с помощью
- 19. Строение гема
- 20. СИНТЕЗ ГЕМА Для синтеза гема требуются: глицин, сукцинил-КоА. Синтез гема протекает в митохондриях и в цитозоле.
- 22. СИНТЕЗ ГЕМА 3. Третья реакция: конденсация 4-х молекул порфобилиногена в тетрапиррол Различают два вида тетрапирролов -
- 23. СИНТЕЗ ГЕМА 4. Уропорфириногены превращаются в соответствующие копропорфириногены. Копропорфириноген III → окисляется в протопорфириноген IX →
- 25. Регуляция синтеза гема 1. Скорость синтеза глобиновых цепей зависит от наличия гема, он ускоряет биосинтез "своих"
- 26. Нарушения синтеза гема. Порфирии Порфирии - гетерогенная группа заболеваний, вызванная нарушениями синтеза гема вследствие дефицита одного
- 27. Классификации порфирий Порфирии делят по причинам на: Наследственные: Возникают при дефекте гена фермента, участвующего в синтезе
- 28. СТРОЕНИЕ ГЕМОГЛОБИНА Нb состоит из 4-х гемсодержащих белковых субъединиц (протомеров) соединенных гидрофобными, ионными, водородными связями по
- 29. Строение гемоглобина А
- 30. Функции гемоглобина 1. Обеспечивают перенос кислорода от легких к тканям; 2. Участвует в переносе углекислого газа
- 31. Производные гемоглобина Оксигемоглобин HbО2 (Fe2+) – соединение молекулярного кислорода с гемоглобином. Карбоксигемоглобин HbСО (Fe2+). Связь гема
- 32. Производные гемоглобина Цианметгемоглобин HbСN (Fe3+). Образуется при присоединении СN- к метгемоглобину. Карбгемоглобин HbСО2 (Fe2+) – соединение
- 33. Виды гемоглобинов Нормальные виды гемоглобина: HbР – примитивный гемоглобин (у эмбриона 7-12 нед.), HbF – фетальный
- 34. Патологические виды гемоглобина HbS – гемоглобин серповидно-клеточной анемии (в β-цепях глу заменен вал) MetHb – метгемоглобин,
- 35. Болезни гемоглобинов - ГЕМОГЛОБИНОЗЫ Описано более 200 гемоглобинозов РАЗЛИЧАЮТ: Гемоглобинопатии - возникают в результате точечных мутаций
- 36. Гемоглобинозы Характерно: 1. Нарушаются пропорции в составе гемоглобина крови. 2. Эритроциты теряют нормальную форму (мишеневидные, каплевидные)
- 37. РАСПАД ГЕМА За сутки у человека распадается около 9 г гемопротеинов, в основном это гемоглобин эритроцитов.
- 38. 4 основные этапа распада гема: I этап - внутри эритроцита старый гемоглобин подвергается частичной денатурации. II
- 39. 1-й этап - внутриэртроцитарный Образование гемоглобин-гаптоглобинового комплекса: Гемоглобин при разрушении эритроцитов в кровеносном русле образует комплекс
- 40. 2-й этап - эритрофагальный Весь эритроцит фагоцитируется эритрофагами - клетками ретикуло-эндотелиальной системы (РЭС) селезенки, печени и
- 41. ОБРАЗОВАНИЕ ГЕМБИЛИРУБИНА В РЭС
- 42. СВОЙСТВА ГЕМБИЛИРУБИНА Не растворим в воде (растворим в жирах), Токсичен для нейронов, Дает непрямую реакцию с
- 43. 3-й этап – гепато-целлюлярный
- 44. СВОЙСТВА ХОЛЕБИЛИРУБИНА Растворим в воде, Малотоксичен для нейронов, Дает прямую реакцию с диазореактивом, Связан с глюкуроновой
- 45. ОБЩИЙ БИЛИРУБИН КРОВИ В крови в норме одновременно присутствуют две формы билирубина: - гембилирубин (свободный), попадающий
- 46. 4-й этап - энтенральный В кишечнике при участии бактериальной β-глюкуронидазы холебилирубин превращаются в гембилирубин. Гембилирубин восстанавливается
- 47. 4-й этап - энтенральный Всосавшийся (10%) в кровоток мезобилиноген попадает в печень, где окисляется до ди-
- 48. 4-й этап - энтенральный В толстом кишечнике: мезобилиноген восстанавливается до стеркобилиногена и выделяется из организма, окрашивая
- 49. КАТАБОЛИЗМ ГЕМОГЛОБИНА
- 50. ГИПЕРБИЛИРУБИНЕМИЯ Увеличение образования билирубина (гемолитическая желтуха) Поражение печени - угнетение конъюгационных или выделительных механизмов печени (паренхиматозная
- 51. ГЕМОЛИТИЧЕСКАЯ ЖЕЛТУХА Гемолитическая или надпеченочная желтуха – причина - внутрисосудистый гемолиз. Кровь: увеличение гембилирубина, холебилирубин в
- 52. ПАРЕНХИМАТОЗНАЯ ЖЕЛТУХА Перенхиматозная или печеночная желтуха – причина – патология печени Кровь: увеличение холебилирубина, увеличение гембилирубина,
- 53. ОБТУРАЦИОННАЯ ЖЕЛТУХА Обтурационная или подпеченочная желтуха – причина – патология желчных путей Кровь: резкое увеличение холебилирубина,
- 54. Физиологическая (транзиторная) желтуха новорожденных Причины относительное снижение активности УДФ-глюкуронилтрансферазы в первые дни жизни, связанное с повышенным
- 55. Гемолитическая болезнь новорожденного Причины: Несовместимость крови матери и плода по группе или по резус-фактору. Накопление гембилирубина
- 56. Наследственные формы паренхиматозной желтухи СИНДРОМЫ: Жильбера-Мейленграхта, Дубина-Джонсона, Криглера-Найяра.
- 57. Синдром Жильбера-Мейленграхта Причины: наследуется по аутосомно-доминантному типу низкая активность УДФ-глюкуронилтрансферазы. Нарушается элиминация билирубина из плазмы крови,
- 58. Синдром Дубина-Джонсона Причины: Аутосомно-доминантная недостаточность выведения конъюгированного билирубина из гепатоцитов в желчные протоки. Встречается чаще у
- 59. Синдром Криглера-Найяра Причины: Полное отсутствие активности УДФ-глюкуронилтрансферазы вследствие аутосомно-рецессивного генетического дефекта. Лабораторная диагностика: Гипербилирубинемия появляется в
- 61. Скачать презентацию

АКТУАЛЬНОСТЬ
Кровь транспортирует из легких в ткани около 600л О2 в сутки.
АКТУАЛЬНОСТЬ
Кровь транспортирует из легких в ткани около 600л О2 в сутки.

ПЛАН ЛЕКЦИИ
1. Биохимия эритроцита
2. Синтез гема
3. Гемоглобинопатии. Талассемии
4. Распад гемоглобина
5. Желтухи
ПЛАН ЛЕКЦИИ
1. Биохимия эритроцита
2. Синтез гема
3. Гемоглобинопатии. Талассемии
4. Распад гемоглобина
5. Желтухи

ЭРИТРОЦИТЫ
Эритроциты (erythrosytus) это форменные элементы крови.
Функция эритроцитов:
1. Регуляция в
ЭРИТРОЦИТЫ
Эритроциты (erythrosytus) это форменные элементы крови.
Функция эритроцитов:
1. Регуляция в
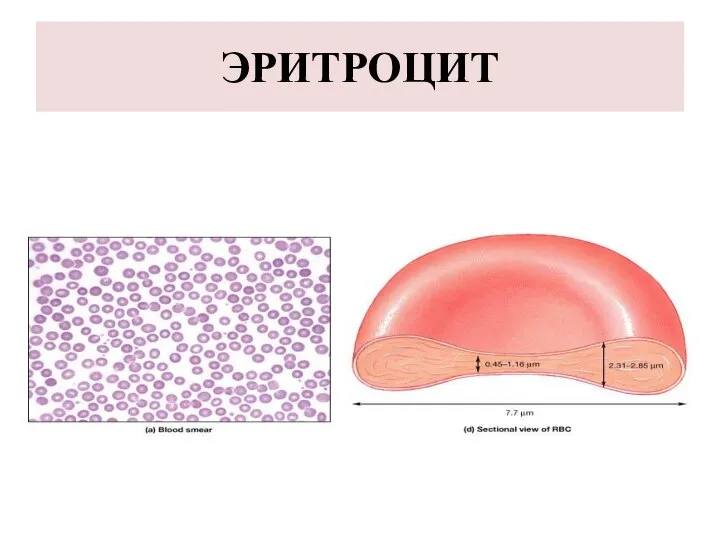
ЭРИТРОЦИТ
ЭРИТРОЦИТ

Плазмолемма эритроцитов
Плазмолемма эритроцитов имеет толщину около 20 нм.
Она состоит из
Плазмолемма эритроцитов
Плазмолемма эритроцитов имеет толщину около 20 нм.
Она состоит из

Липиды
Бислой плазмолеммы образован:
глицерофосфолипидами,
сфингофосфолипидами,
гликолипидами,
холестерином.
Внешний слой содержит гликолипиды (около
Липиды
Бислой плазмолеммы образован:
глицерофосфолипидами,
сфингофосфолипидами,
гликолипидами,
холестерином.
Внешний слой содержит гликолипиды (около

Белки
В плазмолемме эритроцита идентифицировано 15 главных белков.
Более 60% всех мембранных
Белки
В плазмолемме эритроцита идентифицировано 15 главных белков.
Более 60% всех мембранных

СПЕКТРИН
Спектрин - основной белок цитоскелета эритроцитов.
Имеет
СПЕКТРИН
Спектрин - основной белок цитоскелета эритроцитов.
Имеет

БЕЛОК ПОЛОСЫ 3
Трансмембранный гликопротеид, его полипептидная цепь много
БЕЛОК ПОЛОСЫ 3
Трансмембранный гликопротеид, его полипептидная цепь много

ГЛИКОФОРИН
трансмембранный гликопротеин, пронизывающий плазмолемму в виде одиночной спирали.
ГЛИКОФОРИН
трансмембранный гликопротеин, пронизывающий плазмолемму в виде одиночной спирали.
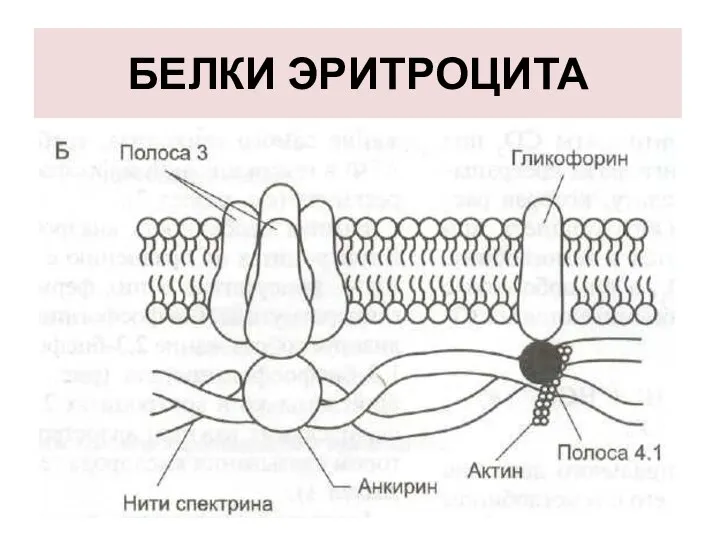
БЕЛКИ ЭРИТРОЦИТА
БЕЛКИ ЭРИТРОЦИТА

Цитоплазма эритроцитов
содержится около 60% воды и 40% сухого остатка.
95%
Цитоплазма эритроцитов
содержится около 60% воды и 40% сухого остатка.
95%

Особенность обмена нуклеотидов в эритроцитах
Из ФРПФ (из рибозо-5ф) и аденина синтезируется
Особенность обмена нуклеотидов в эритроцитах
Из ФРПФ (из рибозо-5ф) и аденина синтезируется

Особенность углеводного обмена в эритроцитах
1. В зрелых Эр углеводы не синтезируются.
2.
Особенность углеводного обмена в эритроцитах
1. В зрелых Эр углеводы не синтезируются.
2.

2,3- Дифосфоглицерат (2,3-ДФГ)
Синтезируется из промежуточного продукта гликолиза – 1,3-дифосфоглицерата
2,3- ДФГ -
2,3- Дифосфоглицерат (2,3-ДФГ)
Синтезируется из промежуточного продукта гликолиза – 1,3-дифосфоглицерата
2,3- ДФГ -

Обезвреживание активных форм кислорода в эритроцитах
Источники активных форм кислорода (АФК):
1. Ηb
Обезвреживание активных форм кислорода в эритроцитах
Источники активных форм кислорода (АФК):
1. Ηb

ГЕМ
- это порфирин, в центре которого находиться Fe2+. Fe2+ включается в
ГЕМ
- это порфирин, в центре которого находиться Fe2+. Fe2+ включается в
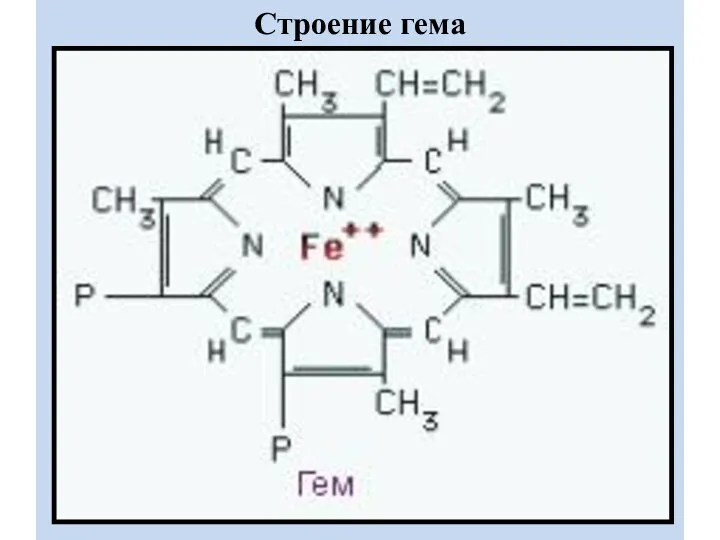
Строение гема
Строение гема

СИНТЕЗ ГЕМА
Для синтеза гема требуются: глицин, сукцинил-КоА. Синтез гема протекает в
СИНТЕЗ ГЕМА
Для синтеза гема требуются: глицин, сукцинил-КоА. Синтез гема протекает в
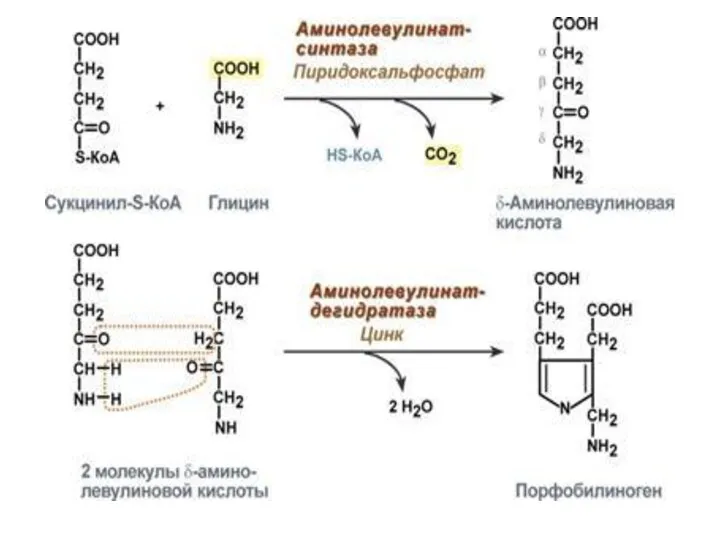

СИНТЕЗ ГЕМА
3. Третья реакция: конденсация 4-х молекул
порфобилиногена в
СИНТЕЗ ГЕМА
3. Третья реакция: конденсация 4-х молекул
порфобилиногена в

СИНТЕЗ ГЕМА
4. Уропорфириногены превращаются в соответствующие копропорфириногены.
Копропорфириноген III
СИНТЕЗ ГЕМА
4. Уропорфириногены превращаются в соответствующие копропорфириногены.
Копропорфириноген III


Регуляция синтеза гема
1. Скорость синтеза глобиновых цепей зависит от наличия гема,
Регуляция синтеза гема
1. Скорость синтеза глобиновых цепей зависит от наличия гема,

Нарушения синтеза гема. Порфирии
Порфирии - гетерогенная группа заболеваний, вызванная нарушениями
Нарушения синтеза гема. Порфирии
Порфирии - гетерогенная группа заболеваний, вызванная нарушениями

Классификации порфирий
Порфирии делят по причинам на:
Наследственные: Возникают при дефекте гена
Классификации порфирий
Порфирии делят по причинам на:
Наследственные: Возникают при дефекте гена

СТРОЕНИЕ ГЕМОГЛОБИНА
Нb состоит из 4-х гемсодержащих белковых субъединиц (протомеров) соединенных гидрофобными,
СТРОЕНИЕ ГЕМОГЛОБИНА
Нb состоит из 4-х гемсодержащих белковых субъединиц (протомеров) соединенных гидрофобными,
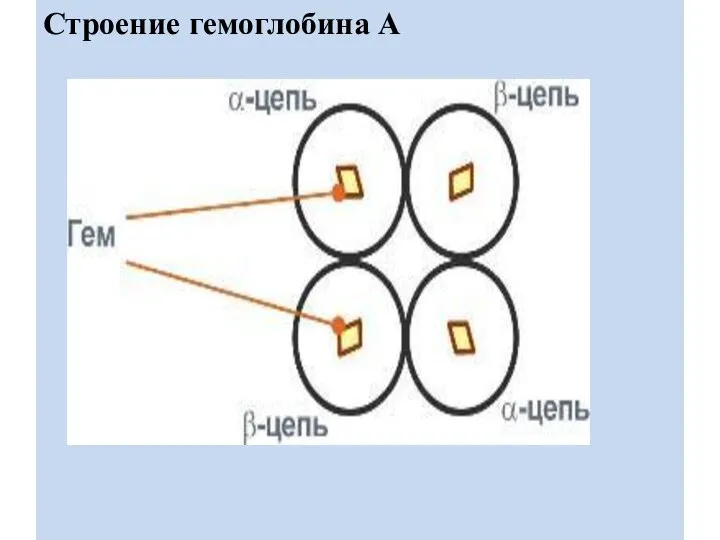
Строение гемоглобина А
Строение гемоглобина А

Функции гемоглобина
1. Обеспечивают перенос кислорода от легких к тканям;
2. Участвует в
Функции гемоглобина
1. Обеспечивают перенос кислорода от легких к тканям;
2. Участвует в

Производные гемоглобина
Оксигемоглобин HbО2 (Fe2+) – соединение молекулярного кислорода с гемоглобином.
Карбоксигемоглобин
Производные гемоглобина
Оксигемоглобин HbО2 (Fe2+) – соединение молекулярного кислорода с гемоглобином.
Карбоксигемоглобин

Производные гемоглобина
Цианметгемоглобин HbСN (Fe3+). Образуется при присоединении СN- к метгемоглобину.
Карбгемоглобин
Производные гемоглобина
Цианметгемоглобин HbСN (Fe3+). Образуется при присоединении СN- к метгемоглобину.
Карбгемоглобин

Виды гемоглобинов
Нормальные виды гемоглобина:
HbР – примитивный гемоглобин (у эмбриона 7-12 нед.),
HbF
Виды гемоглобинов
Нормальные виды гемоглобина:
HbР – примитивный гемоглобин (у эмбриона 7-12 нед.),
HbF

Патологические виды гемоглобина
HbS – гемоглобин серповидно-клеточной анемии (в β-цепях глу заменен
Патологические виды гемоглобина
HbS – гемоглобин серповидно-клеточной анемии (в β-цепях глу заменен

Болезни гемоглобинов - ГЕМОГЛОБИНОЗЫ
Описано более 200 гемоглобинозов
РАЗЛИЧАЮТ:
Гемоглобинопатии - возникают в результате
Болезни гемоглобинов - ГЕМОГЛОБИНОЗЫ
Описано более 200 гемоглобинозов
РАЗЛИЧАЮТ:
Гемоглобинопатии - возникают в результате

Гемоглобинозы
Характерно:
1. Нарушаются пропорции в составе гемоглобина крови.
2. Эритроциты теряют нормальную
Гемоглобинозы
Характерно:
1. Нарушаются пропорции в составе гемоглобина крови.
2. Эритроциты теряют нормальную

РАСПАД ГЕМА
За сутки у человека распадается около 9 г гемопротеинов,
РАСПАД ГЕМА
За сутки у человека распадается около 9 г гемопротеинов,

4 основные этапа распада гема:
I этап - внутри эритроцита старый гемоглобин
4 основные этапа распада гема:
I этап - внутри эритроцита старый гемоглобин

1-й этап - внутриэртроцитарный
Образование гемоглобин-гаптоглобинового комплекса:
Гемоглобин при разрушении эритроцитов
1-й этап - внутриэртроцитарный
Образование гемоглобин-гаптоглобинового комплекса:
Гемоглобин при разрушении эритроцитов

2-й этап - эритрофагальный
Весь эритроцит фагоцитируется эритрофагами - клетками ретикуло-эндотелиальной системы
2-й этап - эритрофагальный
Весь эритроцит фагоцитируется эритрофагами - клетками ретикуло-эндотелиальной системы
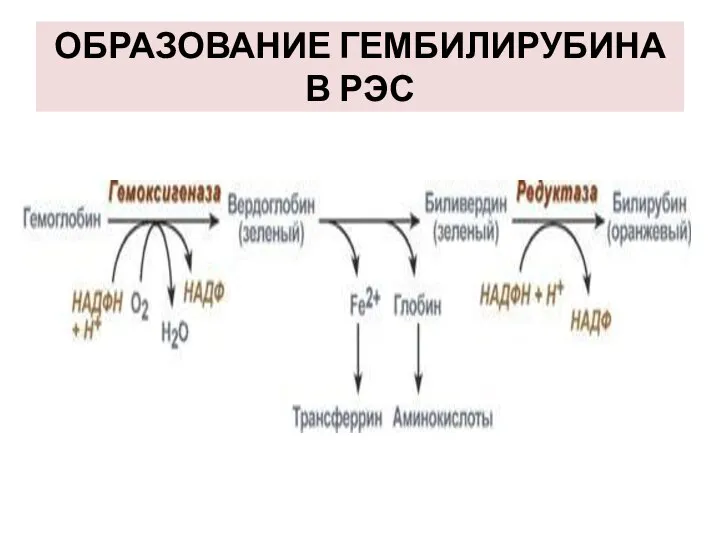
ОБРАЗОВАНИЕ ГЕМБИЛИРУБИНА В РЭС
ОБРАЗОВАНИЕ ГЕМБИЛИРУБИНА В РЭС

СВОЙСТВА ГЕМБИЛИРУБИНА
Не растворим в воде (растворим в жирах),
Токсичен для нейронов,
Дает непрямую
СВОЙСТВА ГЕМБИЛИРУБИНА
Не растворим в воде (растворим в жирах),
Токсичен для нейронов,
Дает непрямую
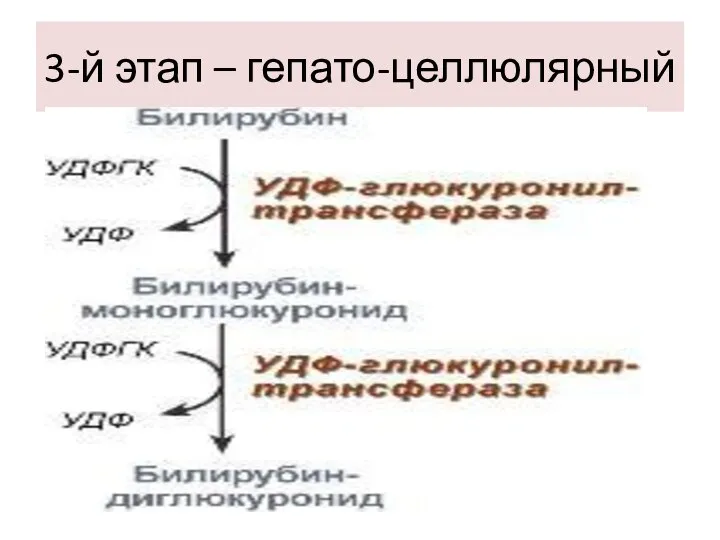
3-й этап – гепато-целлюлярный
3-й этап – гепато-целлюлярный

СВОЙСТВА ХОЛЕБИЛИРУБИНА
Растворим в воде,
Малотоксичен для нейронов,
Дает прямую реакцию с диазореактивом,
Связан с
СВОЙСТВА ХОЛЕБИЛИРУБИНА
Растворим в воде,
Малотоксичен для нейронов,
Дает прямую реакцию с диазореактивом,
Связан с

ОБЩИЙ БИЛИРУБИН КРОВИ
В крови в норме одновременно присутствуют две формы билирубина:
ОБЩИЙ БИЛИРУБИН КРОВИ
В крови в норме одновременно присутствуют две формы билирубина:

4-й этап - энтенральный
В кишечнике при участии бактериальной
β-глюкуронидазы холебилирубин
4-й этап - энтенральный
В кишечнике при участии бактериальной
β-глюкуронидазы холебилирубин

4-й этап - энтенральный
Всосавшийся (10%) в кровоток мезобилиноген попадает в печень,
4-й этап - энтенральный
Всосавшийся (10%) в кровоток мезобилиноген попадает в печень,

4-й этап - энтенральный
В толстом кишечнике:
мезобилиноген восстанавливается до стеркобилиногена и
4-й этап - энтенральный
В толстом кишечнике:
мезобилиноген восстанавливается до стеркобилиногена и

КАТАБОЛИЗМ ГЕМОГЛОБИНА
КАТАБОЛИЗМ ГЕМОГЛОБИНА

ГИПЕРБИЛИРУБИНЕМИЯ
Увеличение образования билирубина
(гемолитическая желтуха)
Поражение печени - угнетение конъюгационных или выделительных
ГИПЕРБИЛИРУБИНЕМИЯ
Увеличение образования билирубина
(гемолитическая желтуха)
Поражение печени - угнетение конъюгационных или выделительных

ГЕМОЛИТИЧЕСКАЯ ЖЕЛТУХА
Гемолитическая или надпеченочная желтуха – причина - внутрисосудистый гемолиз.
ГЕМОЛИТИЧЕСКАЯ ЖЕЛТУХА
Гемолитическая или надпеченочная желтуха – причина - внутрисосудистый гемолиз.

ПАРЕНХИМАТОЗНАЯ ЖЕЛТУХА
Перенхиматозная или печеночная желтуха – причина – патология печени
Кровь: увеличение
ПАРЕНХИМАТОЗНАЯ ЖЕЛТУХА
Перенхиматозная или печеночная желтуха – причина – патология печени
Кровь: увеличение

ОБТУРАЦИОННАЯ ЖЕЛТУХА
Обтурационная или подпеченочная желтуха – причина – патология желчных путей
Кровь:
ОБТУРАЦИОННАЯ ЖЕЛТУХА
Обтурационная или подпеченочная желтуха – причина – патология желчных путей
Кровь:

Физиологическая (транзиторная) желтуха новорожденных
Причины
относительное снижение активности УДФ-глюкуронилтрансферазы в первые дни жизни,
Физиологическая (транзиторная) желтуха новорожденных
Причины
относительное снижение активности УДФ-глюкуронилтрансферазы в первые дни жизни,

Гемолитическая болезнь
новорожденного
Причины: Несовместимость крови матери и плода по группе или
Гемолитическая болезнь
новорожденного
Причины: Несовместимость крови матери и плода по группе или

Наследственные формы паренхиматозной желтухи
СИНДРОМЫ:
Жильбера-Мейленграхта,
Дубина-Джонсона,
Криглера-Найяра.
Наследственные формы паренхиматозной желтухи
СИНДРОМЫ:
Жильбера-Мейленграхта,
Дубина-Джонсона,
Криглера-Найяра.

Синдром Жильбера-Мейленграхта
Причины:
наследуется по аутосомно-доминантному типу низкая активность УДФ-глюкуронилтрансферазы.
Нарушается элиминация
Синдром Жильбера-Мейленграхта
Причины:
наследуется по аутосомно-доминантному типу низкая активность УДФ-глюкуронилтрансферазы.
Нарушается элиминация

Синдром Дубина-Джонсона
Причины:
Аутосомно-доминантная недостаточность выведения конъюгированного билирубина из гепатоцитов в желчные
Синдром Дубина-Джонсона
Причины:
Аутосомно-доминантная недостаточность выведения конъюгированного билирубина из гепатоцитов в желчные

Синдром Криглера-Найяра
Причины:
Полное отсутствие активности УДФ-глюкуронилтрансферазы вследствие аутосомно-рецессивного генетического дефекта.
Лабораторная диагностика:
Гипербилирубинемия
Синдром Криглера-Найяра
Причины:
Полное отсутствие активности УДФ-глюкуронилтрансферазы вследствие аутосомно-рецессивного генетического дефекта.
Лабораторная диагностика:
Гипербилирубинемия
 Тест. Общая характеристика металлов главных подгрупп I-III групп в связи с их положением в Периодической системе
Тест. Общая характеристика металлов главных подгрупп I-III групп в связи с их положением в Периодической системе Репликация ДНК. (Лекция 6)
Репликация ДНК. (Лекция 6) Презентация по Химии "Влияние пищевых добавок на здоровье человека" - скачать смотреть
Презентация по Химии "Влияние пищевых добавок на здоровье человека" - скачать смотреть  Введение в минералогию. Лекция 1
Введение в минералогию. Лекция 1 Каркасные силикаты
Каркасные силикаты Bool-doser team
Bool-doser team Теорія взаємного впливу атомів у молекулах органічних речовин
Теорія взаємного впливу атомів у молекулах органічних речовин Төменгі фосфориттерден азот – фосфорлы тыңайтқыштар алу технологиясын әзірлеу
Төменгі фосфориттерден азот – фосфорлы тыңайтқыштар алу технологиясын әзірлеу Припекание взаимно растворимых твердых тел
Припекание взаимно растворимых твердых тел Изготовление индикаторов растительного происхождения
Изготовление индикаторов растительного происхождения Конденсированные гетероциклы. Нуклеотиды. Нуклеиновые кислоты
Конденсированные гетероциклы. Нуклеотиды. Нуклеиновые кислоты Антидуриан (10 класс)
Антидуриан (10 класс) Катализ в органическом синтезе
Катализ в органическом синтезе Химические свойства кислот
Химические свойства кислот Прочность полимеров
Прочность полимеров Mineralogy. Chemical composition and properties of minerals
Mineralogy. Chemical composition and properties of minerals Классификация химических реакций в органической и неорганической химии
Классификация химических реакций в органической и неорганической химии Углеродистые конструкционные стали
Углеродистые конструкционные стали Альдегиды Выполнила ученица 10В Гимназии №69 Добринских Светлана
Альдегиды Выполнила ученица 10В Гимназии №69 Добринских Светлана Содержание и структура курса органической химии в старшей школе
Содержание и структура курса органической химии в старшей школе Отчет по производственной практике. Место производственной практики ФГБОУ ВО «Пермской ГСХА», кафедра агрохимии
Отчет по производственной практике. Место производственной практики ФГБОУ ВО «Пермской ГСХА», кафедра агрохимии Органические производные аммиака амины
Органические производные аммиака амины Основные вулканические породы. Основные плутонические породы
Основные вулканические породы. Основные плутонические породы Синтез 2,4-диметил-3,5-ди (этоксикар-бонил) пиррол
Синтез 2,4-диметил-3,5-ди (этоксикар-бонил) пиррол 9 класс Урок №2. Предельные углеводороды Составитель презентации – учитель химии МОУ СОШ г. Холма Насонова Т.А.
9 класс Урок №2. Предельные углеводороды Составитель презентации – учитель химии МОУ СОШ г. Холма Насонова Т.А. Получение этиленгликоля
Получение этиленгликоля Презентация по Химии "Взаємозв’язок хімії та енергетики" - скачать смотреть
Презентация по Химии "Взаємозв’язок хімії та енергетики" - скачать смотреть  Склероглюкан
Склероглюкан