- Хімія та обмін ліпідів

Содержание
- 2. До ліпідів відносять жири та жироподібні сполуки, які в організмі тварин складають 7-11% від загальної маси,
- 3. Біологічна роль Енергетична. Окиснення 1 г жиру в тканинах дає 39,6 кДж енергії (що майже в
- 4. Структурна роль. Разом з білками вони формують різні біологічні мембрани, ліпіди входять в структуру органоїдів клітини
- 5. Класифікація Прості 1. Жири 2. Стероли і стериди 3. Воски Складні 1. Фосфоліпіди а) Гліцеринфосфоліпіди б)
- 6. ПРОСТІ ЛІПІДИ Жири (тригліцериди, триацилгліцерини) це складні ефіри гліцерину і вищих карбонових кислот. (пальміто-олео-стеарат)
- 7. Якість і чистоту жирів характеризують рядом фізичних і хімічних параметрів: 1. Число омилення жиру – кількість
- 8. 2. Кислотне число жиру – кількість мг КОН, яка необхідна для нейтралізації вільних карбонових кислот, що
- 9. 3. Йодне число жиру – відповідає кількості г йоду, здатного зв'язатися зі 100 г жиру. Воно
- 10. Стероли і стериди Стероли – високомолекулярні напівциклічні ненасичені спирти, основним представником в організмі тварин є холестерол,
- 11. Холестерол Ергостерол (в рослинах)
- 12. Холестерол виконує самостійну роль як структурний компонент багатьох біологічних мембран. В мембранах еритроцитів деяких тварин він
- 13. Стериди – складні ефіри стеролів і вищих карбонових кислот (частіше пальмітинової). Пальмітохолестерид (входить до складу клітинних
- 14. Воски (цериди) - складні ефіри високомолекулярних одноатомних спиртів і вищих карбонових кислот. Бувають воски рослинного, тваринного
- 15. Бджолиний віск міріциловий пальмітинова спирт кислота
- 16. Спермацет – з головного мозку кашалота. цетиловий пальмітинова спирт кислота
- 17. Ланолін (тваринного походження) Складається з цетилового спирту, пальмітинової або стеаринової кислот та вільних карбонових кислот. Застосовується
- 18. СКЛАДНІ ЛІПІДИ Фосфоліпіди До складу фосфоліпідів, окрім спирту гліцерину і карбонових кислот, входять залишки ортофосфатної кислоти
- 19. А). Гліцеринфосфоліпіди основні представники побудовані на основі фосфатидної кислоти, до складу якої входять насичені і ненасичені
- 20. Фосфатидна кислота Залишок холіну → холінфосфоліпід (лецитин). Залишок коламіну → коламінфосфоліпід (кефалін). Залишок серину → серинфосфоліпід.
- 21. Б). Сфінгозинфосфоліпіди (сфінгомієліни) Ці ліпіди побудовані на основі аміноспирту сфінгозину. Ці сполуки зустрічаються в мієлінових оболонках
- 22. залишок сфінгозину карбонова к-та фосфатна залишок к-та холіну
- 23. Гліколіпіди складні ліпіди, які, окрім спирту сфінгозину і карбонової кислоти, в своєму складі містять вуглеводний компонент:
- 24. Цереброзиди в паренхіматозних органах знайдено цереброзиди з глюкозою замість галактози. Залишок сфінгозину Карбонова кислота β-Д-галактоза
- 25. Гангліозиди На відміну від цереброзидів у складі замість моносахариду знаходиться гетерополісахарид. Входить до складу нервової тканини,
- 26. Переварювання жирів Жири складають до 90% ліпідів, що надходять з їжею. Переварювання жирів відбувається в тонкому
- 27. Переварювання жирів — гідроліз жирів панкреатичною ліпазою. Оптимальне значення рН для панкреатичної ліпази ≈8 досягається шляхом
- 28. Эмульгування жирів Так як жири - нерозчинні у воді сполуки, то вони можуть піддаватися дії ферментів,
- 29. Жовчні кислоти діють як детергенти, розташовуючись на поверхні крапель жиру і знижуючи поверхневий натяг. В результаті
- 30. Жовчні кислоти утворюються в печінці з холестеролу і виділяються в складі жовчі. Жовчні кислоти можна представити
- 31. В жовчі тварин і людини переважає холева (тригідроксихоланова) і дезоксихолева (дигідроксихоланова) кислоти:
- 32. Значно менше в жовчі літохолевої (моногідроксихоланової) кислоти. Більша частина жовчних кислот з'єднується (кон’югує) з амінооцтовою кислотою
- 33. Гідроліз триацилгліцеролів панкреатичною ліпазою
- 34. Переварювання фосфогліцеридів Фосфоліпаза А2 каталізує відщеплення молекули карбонової кислоти від фосфатидилхоліну з утворенням лізофосфатидилхоліну. Фосфоліпази А1,
- 35. Гідроліз ефірів холестеролу в тонкій кишці В підшлунковій залозі містяться також ферменти, що забезпечують повний гідроліз
- 36. Утворення змішаних міцел і всмоктування продуктів гідролізу Продукти гідролізу ліпідів - жирні кислоти з довгим вуглеводневим
- 37. Основна частина міцел повністю або після попереднього (пристінкового) розпаду всмоктується через стінку тонкого кишечнику. В подальшому
- 38. Циркуляція жовчних кислот Найбільш активно солі жовчних кислот всмоктуються в клубовій кишці. Жовчні кислоти далі потрапляють
- 39. Ресинтез нейтрального жиру В стінці кишечнику гліцерин при дії гліцеролфосфокінази взаємодіє з АТФ з утворенням гліцеролфосфату.
- 40. Далі активований гліцерин та ацил-S-КоА взаємодіють між собою з утворенням фосфатидної кислоти. Фосфатидна кислота може використовуватися
- 41. Ресинтез нейтрального жиру
- 42. Ресинтез фосфоліпідів До утворення дигліцериду реакції відбуваються аналогічно реакціям ресинтезу тригліцеридів. Далі: Дигліцерид + ЦДФ-холін →
- 43. Загальна характеристика ліпопротеїнів Ліпіди у водному середовищі (а значить, і в крові) нерозчинні, тому для транспорту
- 44. Ліпопротеїни плазми крові
- 45. Апопротеїни Апопротеїни виконують декілька функцій: • формують структуру ліпопротеїнів; • взаємодіють з рецепторами на поверхні клітин
- 46. Ліпопротеїни В організмі синтезуються наступні типи ліпопротеїнів: 1. хіломікрони (ХМ), 2. ліпопротеїни дуже низької щільності (ЛПДНЩ)
- 47. Властивості ліпопротеїнів ЛП добре розчиняються в крові, опалесценція відсутня, так як мають невеликий розмір і негативний
- 48. Гідроліз тканинних жирів
- 49. Окиснення гліцерину Спочатку гліцерин за участі гліцеролфосфокінази перетворюється в α-гліцеролфосфат, що під дією НАД-залежної α-гліцеролфосфат-дегідрогенази перетворюється
- 51. Окиснення жирних кислот Окиснення вищих жирних кислот було вперше вивчено у 1904 р. Кноопом, який, при
- 52. β-Окиснення жирних кислот β-Окиснення - специфічний шлях катаболізму жирних кислот, при якому від карбоксильного кінця жирної
- 53. Активація жирних кислот Перед початком процесу окиснення жирні кислоти повинні бути активовані, тобто зв'язані макроергічним зв'язком
- 54. Транспорт жирних кислот в мітохондрії Оскільки процес активації відбувається поза мітохондрій, то далі є необхідним транспорт
- 56. Розглянемо реакції β-окиснення на прикладі пальмітинової кислоти – С15Н31СООН. Починається цей процес з активації карбонової кислоти
- 57. Активована пальмітинова кислота вступає в першу реакцію β-окиснення згідно наступної схеми: 1.
- 58. 2. Ненасичена кислота приєднує воду і перетворюється в β‑окси-пальмітинову кислоту (активну форму):
- 59. 3. Наступна реакція окиснення проходить біля β-атому Карбону β-окси-пальмітинової кислоти, тому і весь шлях наступних перетворень
- 60. 4. Закінчується перший цикл β-окиснення відщепленням від вихідної кислоти двовуглецевого залишку у вигляді активної оцтової кислоти
- 61. У випадку окиснення пальмітинової кислоти β-окиснення буде повторюватися 7 разів. Так, в молекулі пальмітинової кислоти 16
- 62. Активні залишки оцтової кислоти, які утворилися в результаті β-окиснення, включаються в ЦТК, де окиснюються з утворенням
- 64. В багатьох тканинах окиснення жирних кислот - важливе джерело енергії. Це тканини з високою активністю ферментів
- 65. α-Окиснення жирних кислот відбувається в мікросомальній фракції клітин (особливо нервових) при дії монооксигеназ і завершується скороченням
- 66. Карбонові кислоти, які мають вуглецевий ланцюг середньої довжини (8−12 атомів Карбону) можуть спочатку окиснюватися в ω-положенні.
- 67. Окиснення ненасичених жирних кислот Ненасичені жирні кислоти окиснюються швидше, ніж насичені. Першим етапом окиснення ненасиченої жирної
- 68. Такі кислоти зі спряженими подвійними зв'язками дуже активні і за наявності кисню окиснюються з утворенням гідропероксидів
- 69. БІОСИНТЕЗ ПРОСТИХ ЖИРІВ Біосинтез молекули жиру складається з трьох основних процесів: 1. утворення вищої жирної кислоти
- 70. 1. Біосинтез вищих жирних кислот В клітинах організму жирні кислоти синтезуються з ацетил-КоА, що утворюється з
- 71. Сумарна реакція біосинтезу жирних кислот в цитоплазмі має наступний вигляд (Е – пальмітилсинтетаза): Ацетил-КоА + 7Малоніл-КоА
- 73. З ацетил-КоА утворюється малоніл-КоА:
- 74. Будова мультиферментного комплексу — синтази жирних кислот
- 75. Синтез пальмітинової кислоти Синтаза жирних кислот: в першому протомері SH-група належить цистеїну, у другому − фосфопантетеїну.
- 76. Загальна схема реакцій синтезу пальмітинової кислоти
- 77. Утворення подвійних зв'язків у радикалах жирних кислот Введення подвійних звязків у радикали жирних кислот називається десатурацією.
- 78. Для утворення подвійного зв'язку в радикалі жирної кислоти необхідними є молекулярний кисень, NADH, цитохром b5 і
- 79. Утворення ненасичених жирних кислот
- 80. Синтез жирних кислот має ряд особливостей: на відміну від окиснення синтез локалізований в ендоплазматичному ретикулумі; джерелом
- 81. 2. Біосинтез гліцерину Основним шляхом біосинтезу гліцеролу в організмі людини є відновлення дигідроксиацетонмонофосфату в α‑гліцеролфосфат. Цей
- 82. α-Гліцеролфосфат (гліцеролфосфатна кислота) як активна форма гліцеролу використовується для біосинтезу тригліцеридів, тобто простих жирів, і складних
- 83. 3. Синтез жирів в печінці та жировій тканині (ДАФ –дигідроацетонфосфат, ДАГ – диацилгліцерин)
- 84. Біосинтез кетонових тіл Кетонові тіла утворюються в печінці. Це недоокиснені метаболіти ліпідного, і меншою мірою, білкового
- 85. У випадку прискореного β-окиснення кількість ацетооцтової кислоти різко збільшується. Поряд з нею утворюються патологічні кетонові тіла
- 86. У зв'язку з цим загальна кількість кетонових тіл у крові збільшується в декілька разів і може
- 87. Регуляція ліпідного обміну 1. Зовнішні фактори (тип харчування): надлишок вуглеводів у раціоні прискорює процеси синтезу жирів
- 88. 2. Внутрішні фактори: гормони, які прискорюють процеси ліпогенезу: - інсулін; гормони, які прискорюють процеси ліполізу (розпаду
- 89. Патологія ліпідного обміну 1. Ожиріння Причини: гіпофункція щитоподібної залози (по тироксину); нестача ліпотропного гормону гіпофізу.
- 90. 2. Порушення переварювання і засвоєння ліпідів: захворювання підшлункової залози (недостатня секреція і активність панкреатичної ліпази); порушення
- 91. 3. Кетози – характеризуюьтся кетонемією та кетонурією. Причини: нестача інсуліну; В3-авітаміноз (дефіцит НS-КоА); нестача ЩОК –
- 93. Скачать презентацию

До ліпідів відносять жири та жироподібні сполуки, які в організмі тварин
До ліпідів відносять жири та жироподібні сполуки, які в організмі тварин

Біологічна роль
Енергетична. Окиснення 1 г жиру в тканинах дає 39,6
Біологічна роль
Енергетична. Окиснення 1 г жиру в тканинах дає 39,6

Структурна роль. Разом з білками вони формують різні біологічні мембрани, ліпіди
Структурна роль. Разом з білками вони формують різні біологічні мембрани, ліпіди

Класифікація
Прості
1. Жири
2. Стероли і стериди
3. Воски
Складні
1. Фосфоліпіди
а)
Класифікація
Прості
1. Жири
2. Стероли і стериди
3. Воски
Складні
1. Фосфоліпіди
а)

ПРОСТІ ЛІПІДИ
Жири
(тригліцериди, триацилгліцерини)
це складні ефіри гліцерину і вищих карбонових кислот.
(пальміто-олео-стеарат)
ПРОСТІ ЛІПІДИ
Жири
(тригліцериди, триацилгліцерини)
це складні ефіри гліцерину і вищих карбонових кислот.
(пальміто-олео-стеарат)

Якість і чистоту жирів характеризують рядом фізичних і хімічних параметрів:
1. Число
Якість і чистоту жирів характеризують рядом фізичних і хімічних параметрів:
1. Число

2. Кислотне число жиру – кількість мг КОН, яка необхідна для
2. Кислотне число жиру – кількість мг КОН, яка необхідна для

3. Йодне число жиру – відповідає кількості г йоду, здатного зв'язатися
3. Йодне число жиру – відповідає кількості г йоду, здатного зв'язатися

Стероли і стериди
Стероли – високомолекулярні напівциклічні ненасичені спирти, основним представником в
Стероли і стериди
Стероли – високомолекулярні напівциклічні ненасичені спирти, основним представником в
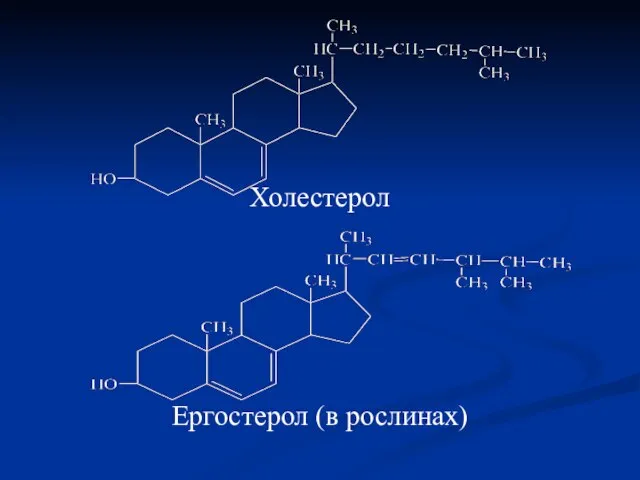
Холестерол
Ергостерол (в рослинах)
Холестерол
Ергостерол (в рослинах)

Холестерол виконує самостійну роль як структурний компонент багатьох біологічних мембран. В
Холестерол виконує самостійну роль як структурний компонент багатьох біологічних мембран. В
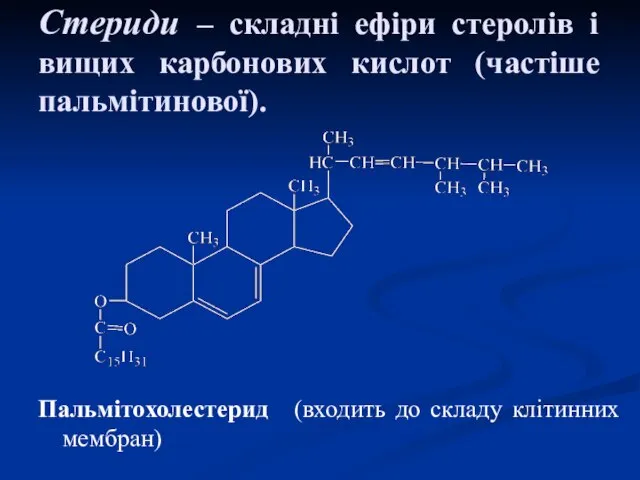
Стериди – складні ефіри стеролів і вищих карбонових кислот (частіше пальмітинової).
Пальмітохолестерид (входить
Стериди – складні ефіри стеролів і вищих карбонових кислот (частіше пальмітинової).
Пальмітохолестерид (входить

Воски (цериди)
- складні ефіри високомолекулярних одноатомних спиртів і вищих карбонових кислот.
Воски (цериди)
- складні ефіри високомолекулярних одноатомних спиртів і вищих карбонових кислот.
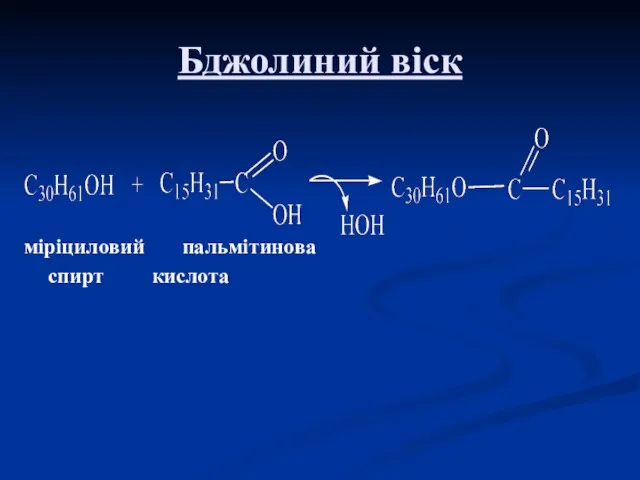
Бджолиний віск
міріциловий пальмітинова
спирт кислота
Бджолиний віск
міріциловий пальмітинова
спирт кислота
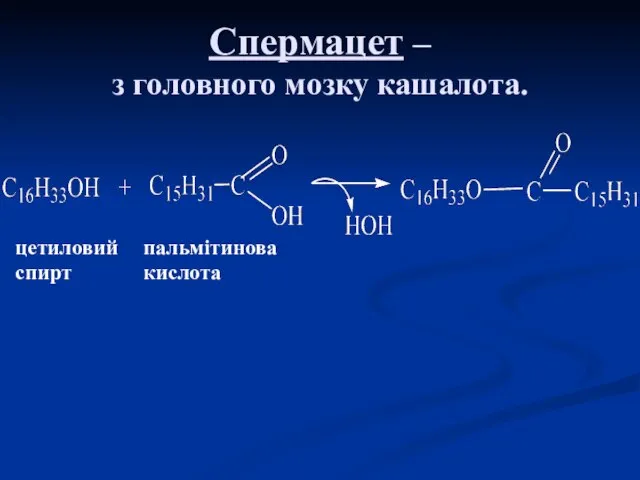
Спермацет –
з головного мозку кашалота.
цетиловий пальмітинова
спирт кислота
Спермацет –
з головного мозку кашалота.
цетиловий пальмітинова
спирт кислота

Ланолін
(тваринного походження)
Складається з цетилового спирту, пальмітинової або стеаринової кислот
Ланолін
(тваринного походження)
Складається з цетилового спирту, пальмітинової або стеаринової кислот

СКЛАДНІ ЛІПІДИ
Фосфоліпіди
До складу фосфоліпідів, окрім спирту гліцерину і карбонових кислот, входять
СКЛАДНІ ЛІПІДИ
Фосфоліпіди
До складу фосфоліпідів, окрім спирту гліцерину і карбонових кислот, входять

А). Гліцеринфосфоліпіди
основні представники побудовані на основі фосфатидної кислоти, до складу якої
А). Гліцеринфосфоліпіди
основні представники побудовані на основі фосфатидної кислоти, до складу якої
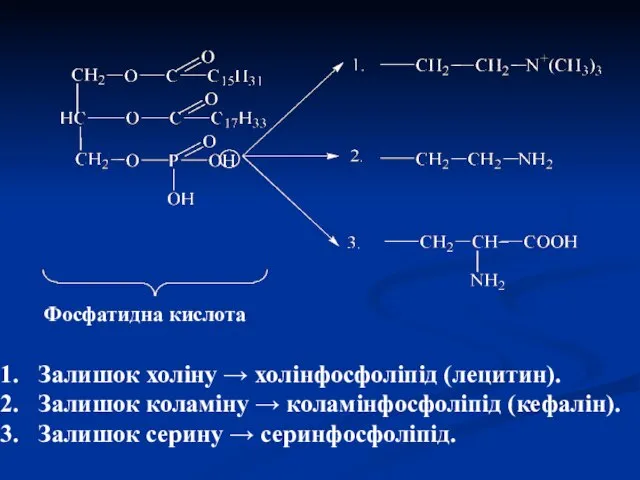
Фосфатидна кислота
Залишок холіну → холінфосфоліпід (лецитин).
Залишок коламіну → коламінфосфоліпід (кефалін).
Залишок
Фосфатидна кислота
Залишок холіну → холінфосфоліпід (лецитин).
Залишок коламіну → коламінфосфоліпід (кефалін).
Залишок

Б). Сфінгозинфосфоліпіди (сфінгомієліни)
Ці ліпіди побудовані на основі аміноспирту сфінгозину.
Ці сполуки
Б). Сфінгозинфосфоліпіди (сфінгомієліни)
Ці ліпіди побудовані на основі аміноспирту сфінгозину.
Ці сполуки
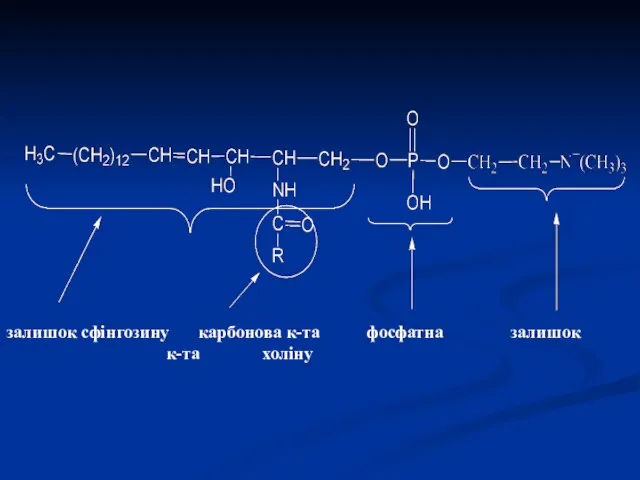
залишок сфінгозину карбонова к-та фосфатна залишок к-та холіну
залишок сфінгозину карбонова к-та фосфатна залишок к-та холіну

Гліколіпіди
складні ліпіди, які, окрім спирту сфінгозину і карбонової кислоти, в своєму
Гліколіпіди
складні ліпіди, які, окрім спирту сфінгозину і карбонової кислоти, в своєму
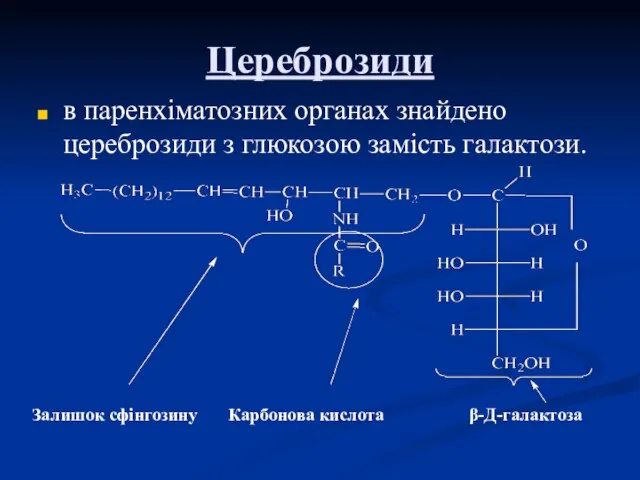
Цереброзиди
в паренхіматозних органах знайдено цереброзиди з глюкозою замість галактози.
Залишок сфінгозину
Цереброзиди
в паренхіматозних органах знайдено цереброзиди з глюкозою замість галактози.
Залишок сфінгозину

Гангліозиди
На відміну від цереброзидів у складі замість моносахариду знаходиться гетерополісахарид. Входить
Гангліозиди
На відміну від цереброзидів у складі замість моносахариду знаходиться гетерополісахарид. Входить

Переварювання жирів
Жири складають до 90% ліпідів, що надходять з їжею. Переварювання
Переварювання жирів
Жири складають до 90% ліпідів, що надходять з їжею. Переварювання

Переварювання жирів — гідроліз жирів панкреатичною ліпазою. Оптимальне значення рН для
Переварювання жирів — гідроліз жирів панкреатичною ліпазою. Оптимальне значення рН для

Эмульгування жирів
Так як жири - нерозчинні у воді сполуки, то вони
Эмульгування жирів
Так як жири - нерозчинні у воді сполуки, то вони

Жовчні кислоти діють як детергенти, розташовуючись на поверхні крапель жиру і
Жовчні кислоти діють як детергенти, розташовуючись на поверхні крапель жиру і
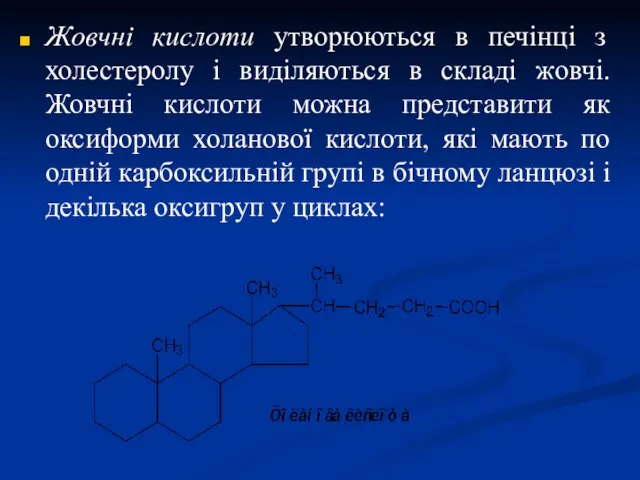
Жовчні кислоти утворюються в печінці з холестеролу і виділяються в складі
Жовчні кислоти утворюються в печінці з холестеролу і виділяються в складі
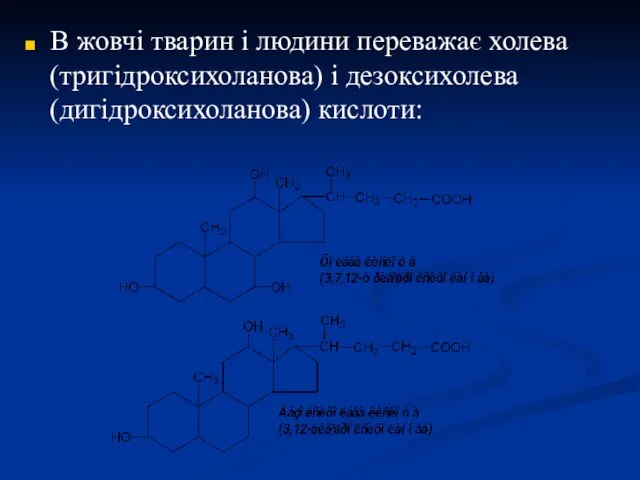
В жовчі тварин і людини переважає холева (тригідроксихоланова) і дезоксихолева (дигідроксихоланова)
В жовчі тварин і людини переважає холева (тригідроксихоланова) і дезоксихолева (дигідроксихоланова)

Значно менше в жовчі літохолевої (моногідроксихоланової) кислоти.
Більша частина жовчних кислот з'єднується
Значно менше в жовчі літохолевої (моногідроксихоланової) кислоти.
Більша частина жовчних кислот з'єднується

Гідроліз триацилгліцеролів панкреатичною ліпазою
Гідроліз триацилгліцеролів панкреатичною ліпазою
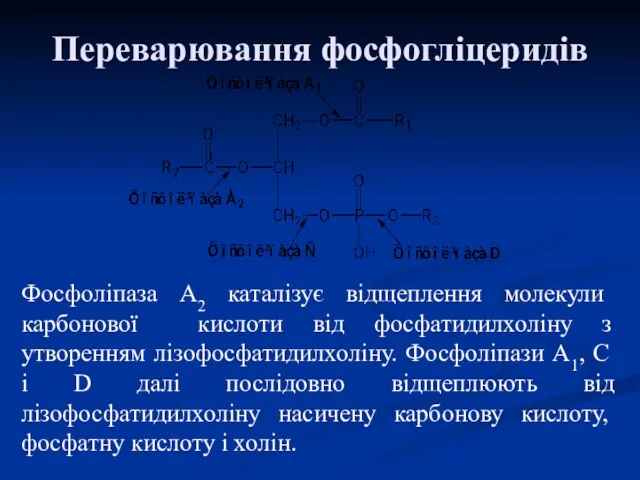
Переварювання фосфогліцеридів
Фосфоліпаза А2 каталізує відщеплення молекули карбонової кислоти від фосфатидилхоліну з
Переварювання фосфогліцеридів
Фосфоліпаза А2 каталізує відщеплення молекули карбонової кислоти від фосфатидилхоліну з
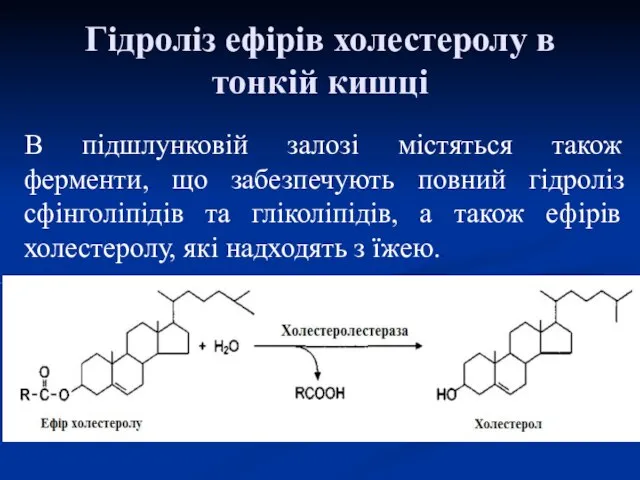
Гідроліз ефірів холестеролу в тонкій кишці
В підшлунковій залозі містяться також
Гідроліз ефірів холестеролу в тонкій кишці
В підшлунковій залозі містяться також

Утворення змішаних міцел і всмоктування продуктів гідролізу
Продукти гідролізу ліпідів - жирні
Утворення змішаних міцел і всмоктування продуктів гідролізу
Продукти гідролізу ліпідів - жирні

Основна частина міцел повністю або після попереднього (пристінкового) розпаду всмоктується через
Основна частина міцел повністю або після попереднього (пристінкового) розпаду всмоктується через

Циркуляція жовчних кислот
Найбільш активно солі жовчних кислот всмоктуються в клубовій кишці.
Циркуляція жовчних кислот
Найбільш активно солі жовчних кислот всмоктуються в клубовій кишці.

Ресинтез нейтрального жиру
В стінці кишечнику гліцерин при дії гліцеролфосфокінази взаємодіє з
Ресинтез нейтрального жиру
В стінці кишечнику гліцерин при дії гліцеролфосфокінази взаємодіє з

Далі активований гліцерин та ацил-S-КоА взаємодіють між собою з утворенням фосфатидної
Далі активований гліцерин та ацил-S-КоА взаємодіють між собою з утворенням фосфатидної
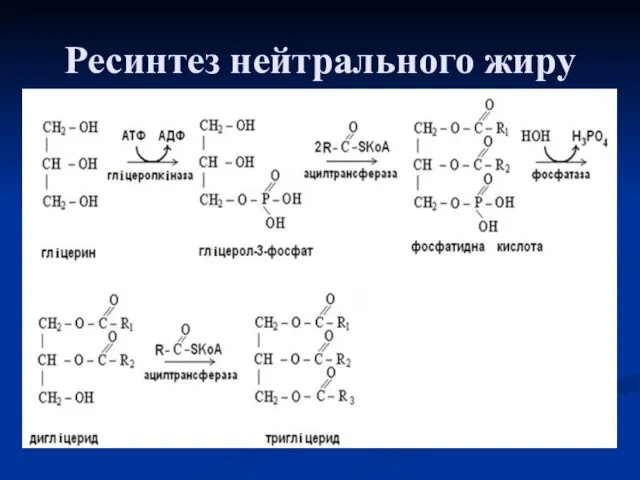
Ресинтез нейтрального жиру
Ресинтез нейтрального жиру

Ресинтез фосфоліпідів
До утворення дигліцериду реакції відбуваються аналогічно реакціям ресинтезу тригліцеридів.
Далі:
Дигліцерид +
Ресинтез фосфоліпідів
До утворення дигліцериду реакції відбуваються аналогічно реакціям ресинтезу тригліцеридів.
Далі:
Дигліцерид +

Загальна характеристика ліпопротеїнів
Ліпіди у водному середовищі (а значить, і в крові)
Загальна характеристика ліпопротеїнів
Ліпіди у водному середовищі (а значить, і в крові)
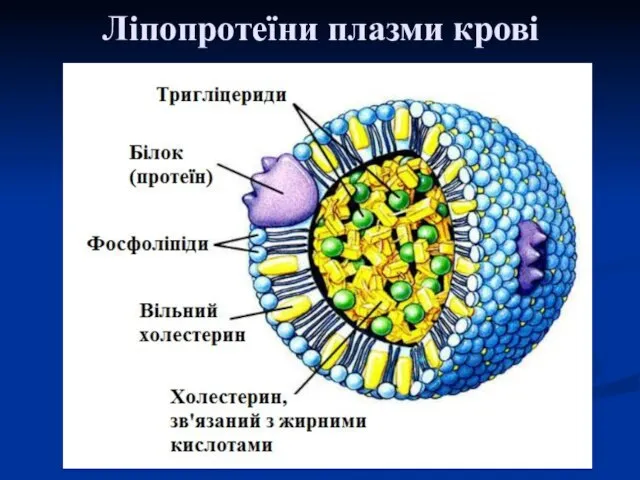
Ліпопротеїни плазми крові
Ліпопротеїни плазми крові

Апопротеїни
Апопротеїни виконують декілька функцій:
• формують структуру ліпопротеїнів;
• взаємодіють з рецепторами на
Апопротеїни
Апопротеїни виконують декілька функцій:
• формують структуру ліпопротеїнів;
• взаємодіють з рецепторами на

Ліпопротеїни
В організмі синтезуються наступні типи ліпопротеїнів:
1. хіломікрони (ХМ),
2. ліпопротеїни
Ліпопротеїни
В організмі синтезуються наступні типи ліпопротеїнів:
1. хіломікрони (ХМ),
2. ліпопротеїни

Властивості ліпопротеїнів
ЛП добре розчиняються в крові, опалесценція відсутня, так як мають
Властивості ліпопротеїнів
ЛП добре розчиняються в крові, опалесценція відсутня, так як мають
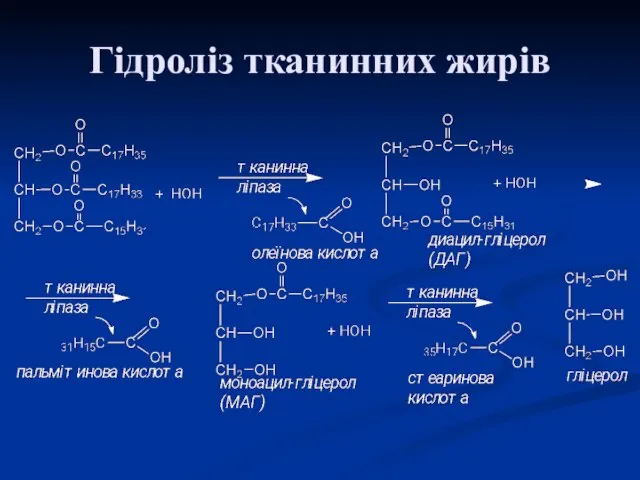
Гідроліз тканинних жирів
Гідроліз тканинних жирів

Окиснення гліцерину
Спочатку гліцерин за участі гліцеролфосфокінази перетворюється в α-гліцеролфосфат, що під
Окиснення гліцерину
Спочатку гліцерин за участі гліцеролфосфокінази перетворюється в α-гліцеролфосфат, що під


Окиснення жирних кислот
Окиснення вищих жирних кислот було вперше вивчено у 1904
Окиснення жирних кислот
Окиснення вищих жирних кислот було вперше вивчено у 1904

β-Окиснення жирних кислот
β-Окиснення - специфічний шлях катаболізму жирних кислот, при якому
β-Окиснення жирних кислот
β-Окиснення - специфічний шлях катаболізму жирних кислот, при якому

Активація жирних кислот
Перед початком процесу окиснення жирні кислоти повинні бути активовані,
Активація жирних кислот
Перед початком процесу окиснення жирні кислоти повинні бути активовані,

Транспорт жирних кислот в мітохондрії
Оскільки процес активації відбувається поза мітохондрій, то
Транспорт жирних кислот в мітохондрії
Оскільки процес активації відбувається поза мітохондрій, то
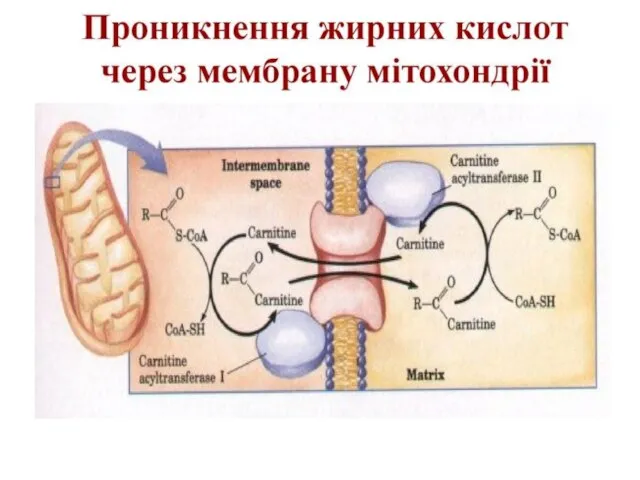

Розглянемо реакції β-окиснення на прикладі пальмітинової кислоти – С15Н31СООН. Починається цей
Розглянемо реакції β-окиснення на прикладі пальмітинової кислоти – С15Н31СООН. Починається цей

Активована пальмітинова кислота вступає в першу реакцію β-окиснення згідно наступної схеми:
1.
Активована пальмітинова кислота вступає в першу реакцію β-окиснення згідно наступної схеми:
1.

2. Ненасичена кислота приєднує воду і перетворюється в β‑окси-пальмітинову кислоту (активну
2. Ненасичена кислота приєднує воду і перетворюється в β‑окси-пальмітинову кислоту (активну

3. Наступна реакція окиснення проходить біля β-атому Карбону β-окси-пальмітинової кислоти, тому
3. Наступна реакція окиснення проходить біля β-атому Карбону β-окси-пальмітинової кислоти, тому

4. Закінчується перший цикл β-окиснення відщепленням від вихідної кислоти двовуглецевого залишку
4. Закінчується перший цикл β-окиснення відщепленням від вихідної кислоти двовуглецевого залишку

У випадку окиснення пальмітинової кислоти β-окиснення буде повторюватися 7 разів. Так,
У випадку окиснення пальмітинової кислоти β-окиснення буде повторюватися 7 разів. Так,

Активні залишки оцтової кислоти, які утворилися в результаті β-окиснення, включаються в
Активні залишки оцтової кислоти, які утворилися в результаті β-окиснення, включаються в


В багатьох тканинах окиснення жирних кислот - важливе джерело енергії. Це
В багатьох тканинах окиснення жирних кислот - важливе джерело енергії. Це

α-Окиснення жирних кислот відбувається в мікросомальній фракції клітин (особливо нервових) при
α-Окиснення жирних кислот відбувається в мікросомальній фракції клітин (особливо нервових) при

Карбонові кислоти, які мають вуглецевий ланцюг середньої довжини (8−12 атомів Карбону)
Карбонові кислоти, які мають вуглецевий ланцюг середньої довжини (8−12 атомів Карбону)

Окиснення ненасичених жирних кислот
Ненасичені жирні кислоти окиснюються швидше, ніж насичені.
Першим
Окиснення ненасичених жирних кислот
Ненасичені жирні кислоти окиснюються швидше, ніж насичені.
Першим

Такі кислоти зі спряженими подвійними зв'язками дуже активні і за наявності
Такі кислоти зі спряженими подвійними зв'язками дуже активні і за наявності

БІОСИНТЕЗ ПРОСТИХ ЖИРІВ
Біосинтез молекули жиру складається з трьох основних процесів:
1.
БІОСИНТЕЗ ПРОСТИХ ЖИРІВ
Біосинтез молекули жиру складається з трьох основних процесів:
1.

1. Біосинтез вищих жирних кислот
В клітинах організму жирні кислоти синтезуються з
1. Біосинтез вищих жирних кислот
В клітинах організму жирні кислоти синтезуються з

Сумарна реакція біосинтезу жирних кислот в цитоплазмі має наступний вигляд (Е
Сумарна реакція біосинтезу жирних кислот в цитоплазмі має наступний вигляд (Е
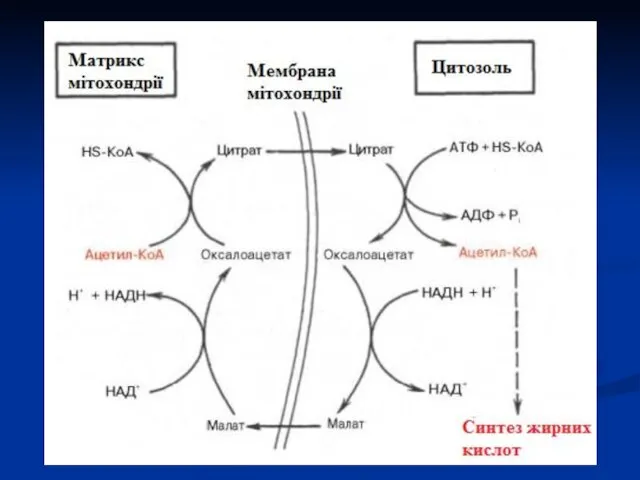

З ацетил-КоА утворюється малоніл-КоА:
З ацетил-КоА утворюється малоніл-КоА:
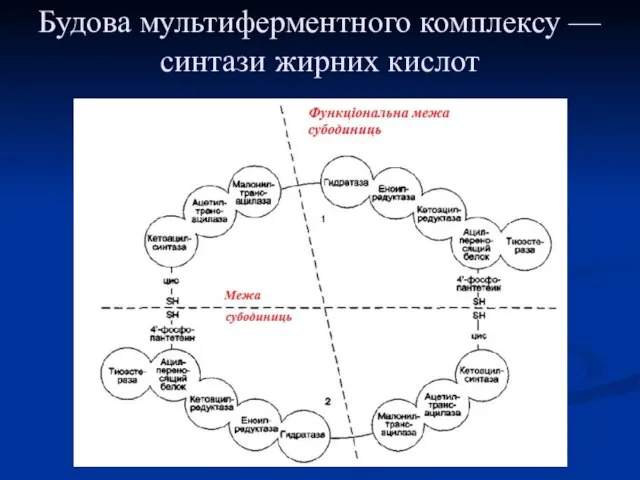
Будова мультиферментного комплексу — синтази жирних кислот
Будова мультиферментного комплексу — синтази жирних кислот
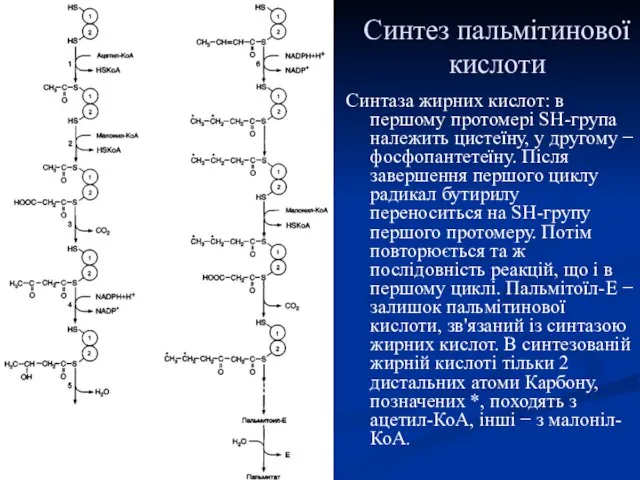
Синтез пальмітинової кислоти
Синтаза жирних кислот: в першому протомері SH-група належить цистеїну,
Синтез пальмітинової кислоти
Синтаза жирних кислот: в першому протомері SH-група належить цистеїну,

Загальна схема реакцій синтезу пальмітинової кислоти
Загальна схема реакцій синтезу пальмітинової кислоти

Утворення подвійних зв'язків у радикалах жирних кислот
Введення подвійних звязків у радикали
Утворення подвійних зв'язків у радикалах жирних кислот
Введення подвійних звязків у радикали

Для утворення подвійного зв'язку в радикалі жирної кислоти необхідними є молекулярний
Для утворення подвійного зв'язку в радикалі жирної кислоти необхідними є молекулярний
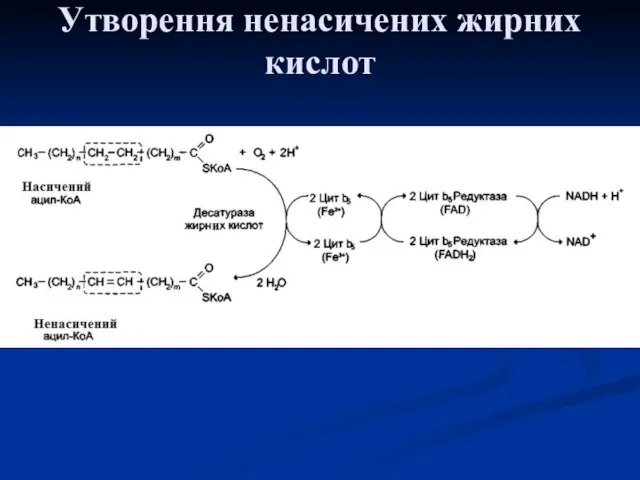
Утворення ненасичених жирних кислот
Утворення ненасичених жирних кислот

Синтез жирних кислот має ряд особливостей:
на відміну від окиснення синтез локалізований
Синтез жирних кислот має ряд особливостей:
на відміну від окиснення синтез локалізований

2. Біосинтез гліцерину
Основним шляхом біосинтезу гліцеролу в організмі людини є відновлення
2. Біосинтез гліцерину
Основним шляхом біосинтезу гліцеролу в організмі людини є відновлення

α-Гліцеролфосфат (гліцеролфосфатна кислота) як активна форма гліцеролу використовується для біосинтезу тригліцеридів,
α-Гліцеролфосфат (гліцеролфосфатна кислота) як активна форма гліцеролу використовується для біосинтезу тригліцеридів,
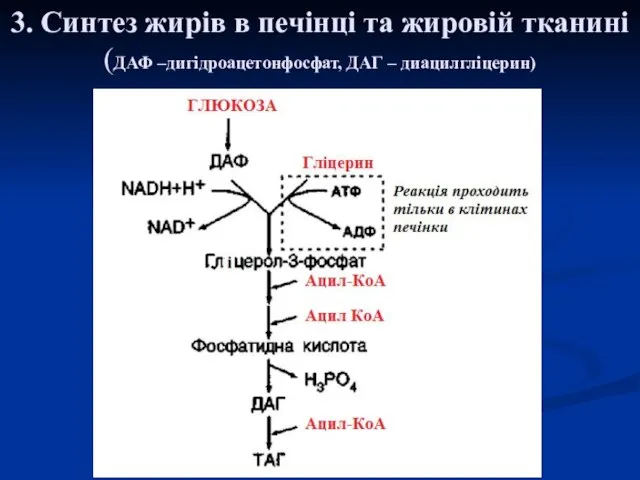
3. Синтез жирів в печінці та жировій тканині
(ДАФ –дигідроацетонфосфат, ДАГ –
3. Синтез жирів в печінці та жировій тканині (ДАФ –дигідроацетонфосфат, ДАГ –

Біосинтез кетонових тіл
Кетонові тіла утворюються в печінці. Це недоокиснені метаболіти ліпідного,
Біосинтез кетонових тіл
Кетонові тіла утворюються в печінці. Це недоокиснені метаболіти ліпідного,
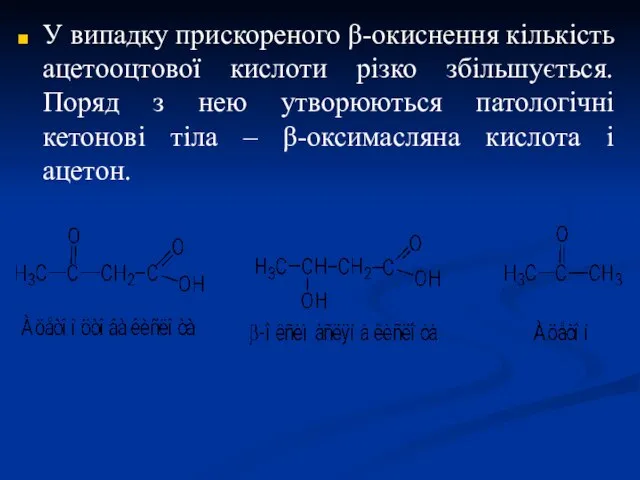
У випадку прискореного β-окиснення кількість ацетооцтової кислоти різко збільшується. Поряд з
У випадку прискореного β-окиснення кількість ацетооцтової кислоти різко збільшується. Поряд з

У зв'язку з цим загальна кількість кетонових тіл у крові збільшується
У зв'язку з цим загальна кількість кетонових тіл у крові збільшується

Регуляція ліпідного обміну
1. Зовнішні фактори (тип харчування):
надлишок вуглеводів у раціоні прискорює
Регуляція ліпідного обміну
1. Зовнішні фактори (тип харчування):
надлишок вуглеводів у раціоні прискорює

2. Внутрішні фактори:
гормони, які прискорюють процеси ліпогенезу:
- інсулін;
гормони, які прискорюють процеси
2. Внутрішні фактори:
гормони, які прискорюють процеси ліпогенезу:
- інсулін;
гормони, які прискорюють процеси

Патологія ліпідного обміну
1. Ожиріння
Причини:
гіпофункція щитоподібної залози (по тироксину);
нестача ліпотропного гормону
Патологія ліпідного обміну
1. Ожиріння
Причини:
гіпофункція щитоподібної залози (по тироксину);
нестача ліпотропного гормону

2. Порушення переварювання і засвоєння ліпідів:
захворювання підшлункової залози (недостатня секреція і
2. Порушення переварювання і засвоєння ліпідів:
захворювання підшлункової залози (недостатня секреція і

3. Кетози – характеризуюьтся кетонемією та кетонурією. Причини:
нестача інсуліну;
В3-авітаміноз (дефіцит НS-КоА);
нестача
3. Кетози – характеризуюьтся кетонемією та кетонурією. Причини:
нестача інсуліну;
В3-авітаміноз (дефіцит НS-КоА);
нестача
 Бытовая химия в квартире
Бытовая химия в квартире Кислоты
Кислоты Центрифугирование в почвоведении
Центрифугирование в почвоведении Реакции замещения. Электрохимический ряд напряжений. Класс: 8 Учитель: Осиевская Инна Анатольевна
Реакции замещения. Электрохимический ряд напряжений. Класс: 8 Учитель: Осиевская Инна Анатольевна  Презентация по теме Полисахариды. Крахмал и целлюлоза
Презентация по теме Полисахариды. Крахмал и целлюлоза Галогены
Галогены Загальна характеристика неметалічних елементів. Неметали як прості речовини. Явище алотропії
Загальна характеристика неметалічних елементів. Неметали як прості речовини. Явище алотропії π-комплексы металлов. Природа связи металлуглерод в π-комплексах металлов. Отличия в свойствах от обычной σ-связи
π-комплексы металлов. Природа связи металлуглерод в π-комплексах металлов. Отличия в свойствах от обычной σ-связи Personal сare. Akypo Foam Rl 40
Personal сare. Akypo Foam Rl 40 Амінокислоти. Класифікація та номенклатура
Амінокислоти. Класифікація та номенклатура Аминокислоты «Единственный путь, ведущий к знанию – это деятельность» Б. Шоу
Аминокислоты «Единственный путь, ведущий к знанию – это деятельность» Б. Шоу Биосинтез гема
Биосинтез гема Спирты
Спирты Химические свойства металлов
Химические свойства металлов Презентация Витамин С
Презентация Витамин С Источники сырья для производства полимера полиуретана, области применения, его свойства, объемы его производства
Источники сырья для производства полимера полиуретана, области применения, его свойства, объемы его производства Применение этанола
Применение этанола Сущность процесса электролиза
Сущность процесса электролиза Альдегиды и кетоны
Альдегиды и кетоны Внесение удобрений разбрасыванием
Внесение удобрений разбрасыванием Медьсодержащие соединения
Медьсодержащие соединения Главная подгруппа V группы. Фосфор
Главная подгруппа V группы. Фосфор Теоретическая электрохимия
Теоретическая электрохимия Пространственное строение. Стереоизомерия. Структура и функции биолекул
Пространственное строение. Стереоизомерия. Структура и функции биолекул Виды изомерии. Углеводороды
Виды изомерии. Углеводороды Виды топлива и их характеристика
Виды топлива и их характеристика Менделєєв Дмитро Іванович
Менделєєв Дмитро Іванович  Целестин мінерал Підготувала учениця 8 – А класу Мостєпан Ксенія
Целестин мінерал Підготувала учениця 8 – А класу Мостєпан Ксенія